Marine Microalgae with Anti-Cancer Properties





Abstract
1. Introduction
2. Marine Microalgae
3. Active Fractions from Marine Microalgae
3.1. Carotenoid Extract from Chlorella Ellipsoidea
3.2. Ethanol and Ethyl Acetate Extracts from Chaetoceros Calcitrans
3.3. Organic Fractions from Amphidinium Carterae
3.4. Methanolic Extracts from Amphidinium Carterae, Prorocentrum Rhathymum, Symbiodinium sp., Coolia Malayensis, Ostreopsis Ovata, Amphidinium operculatum and Heterocapsa psammophila
3.5. Hydrophobic Fraction from Skeletonema Marinoi
3.6. Aqueous Extract from a Canadian Marine Microalgal Pool
3.7. Aqueous Extract from Chlorella Sorokiniana
4. Active Compounds from Marine Microalgae
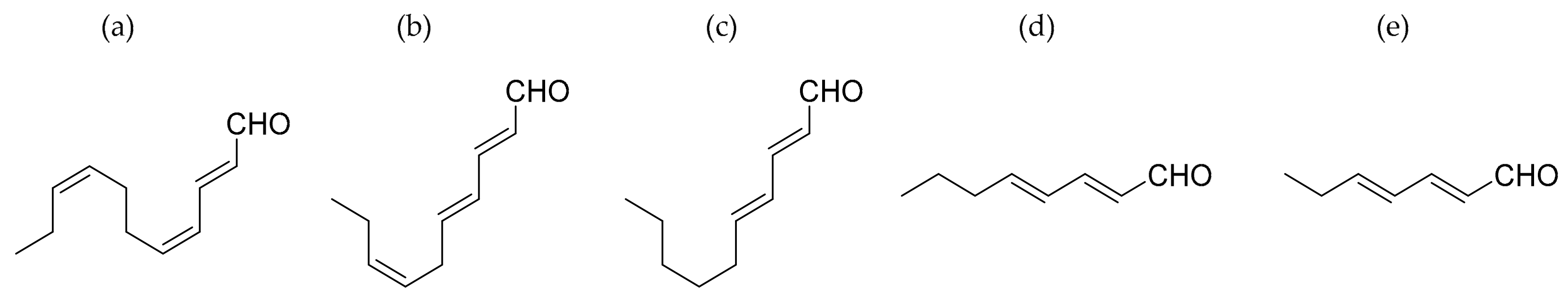
4.1. Polyunsaturated Aldehides (PUAs)
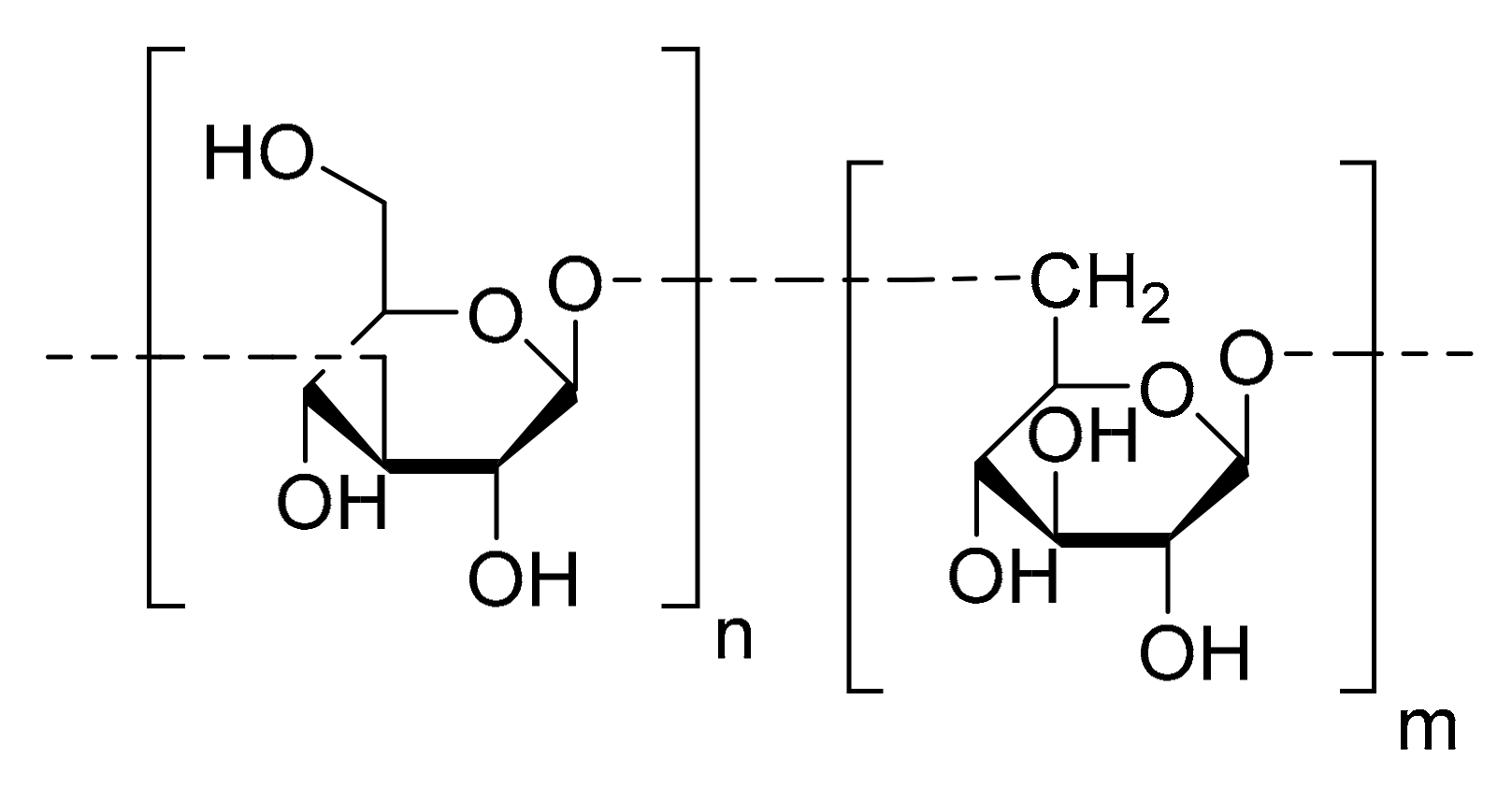
4.2. Chrysolaminaran Polysaccharide
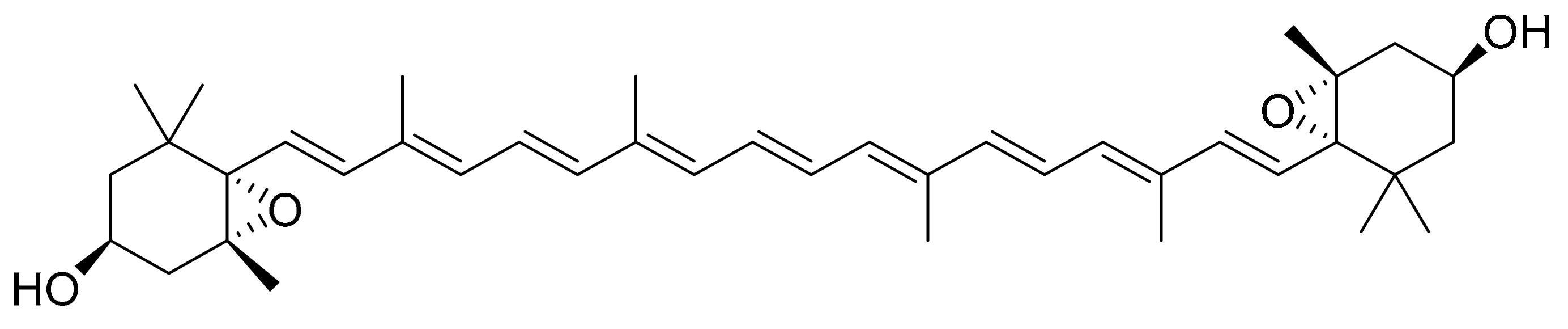
4.3. Violaxanthin
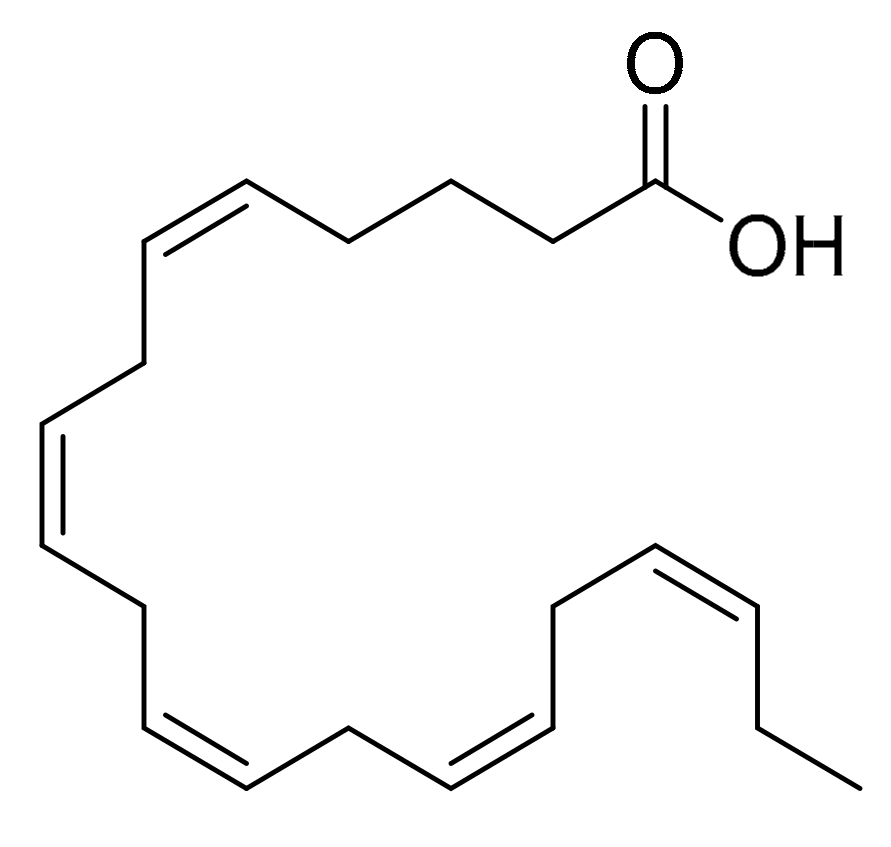
4.4. Eicosapentaenoic Acid (EPA)
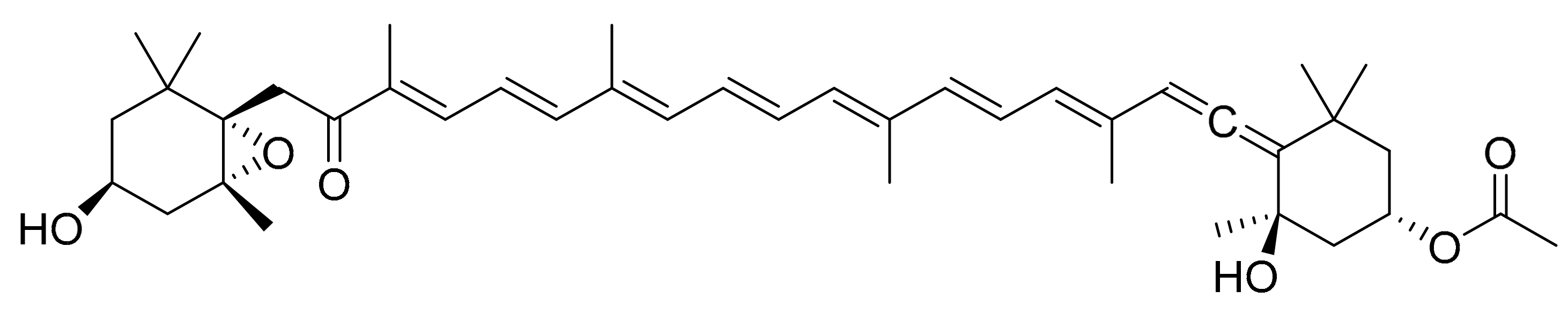
4.5. Fucoxanthin
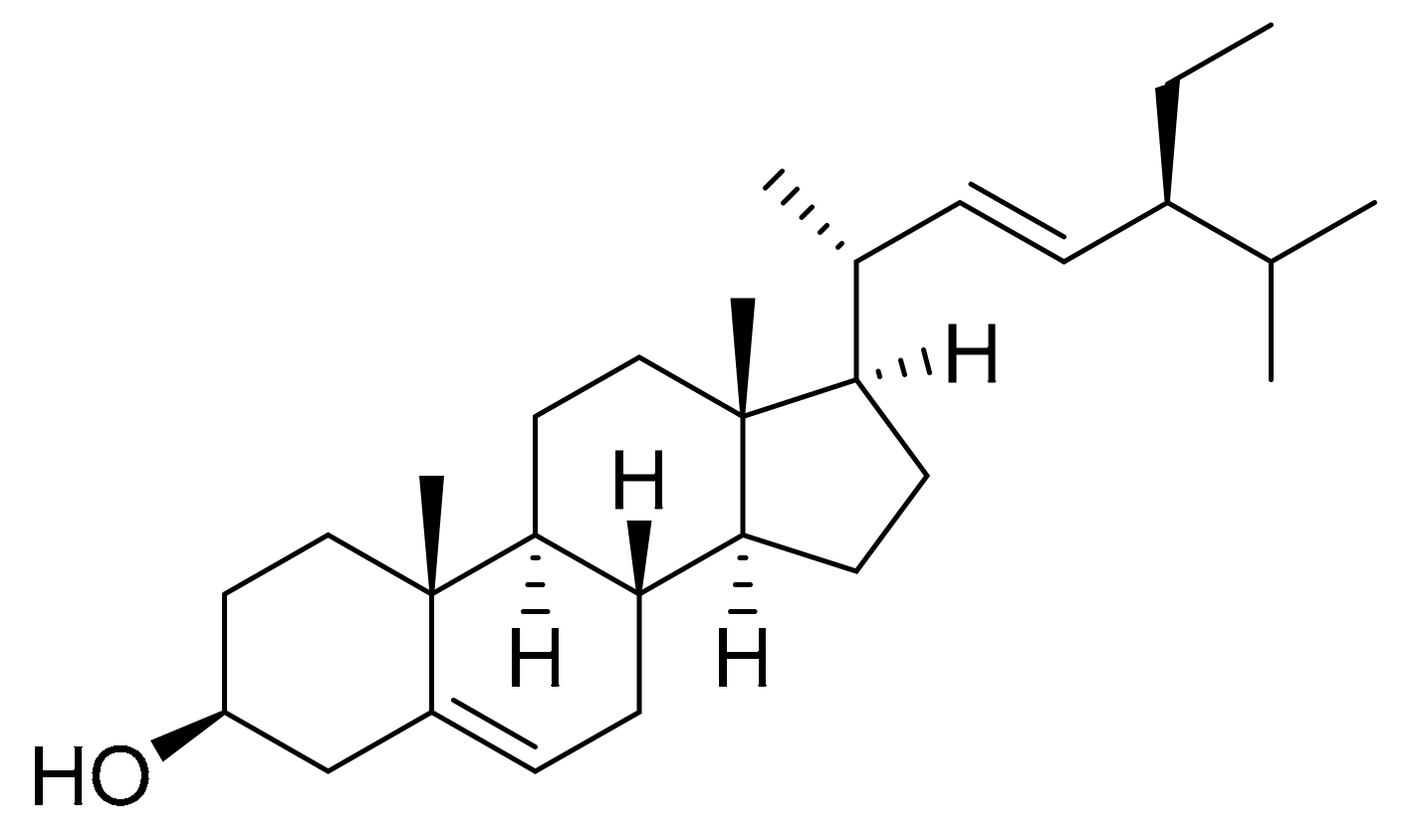
4.6. Stigmasterol
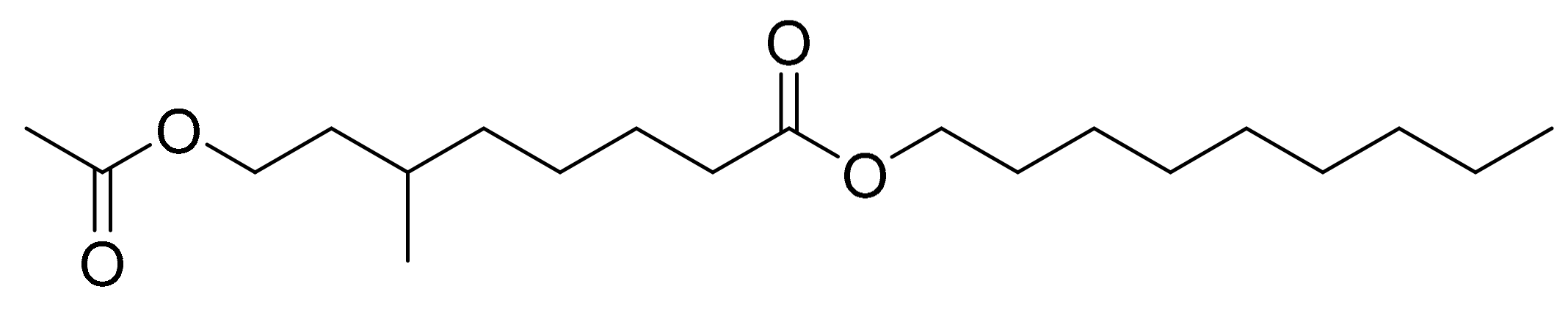
4.7. NAMO (Nonyl 8-acetoxy-6-methyloctanoate)
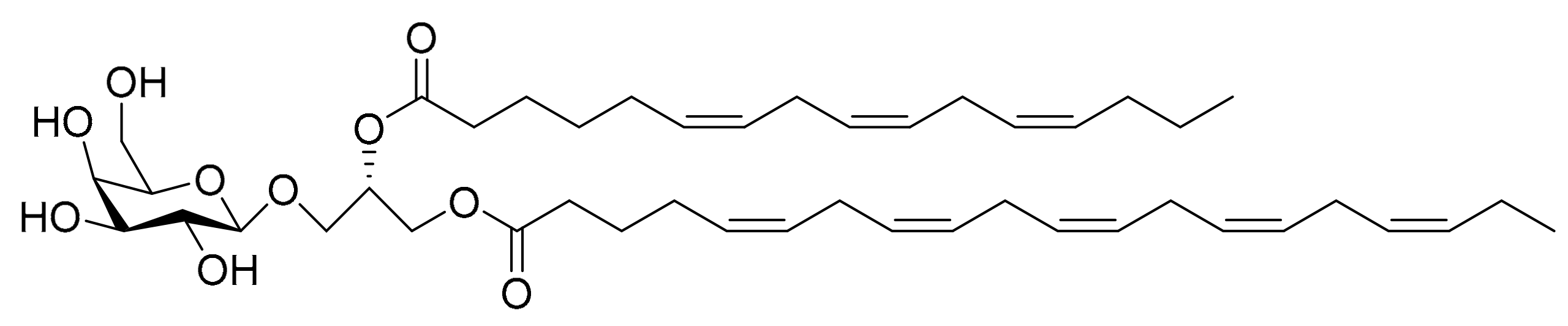
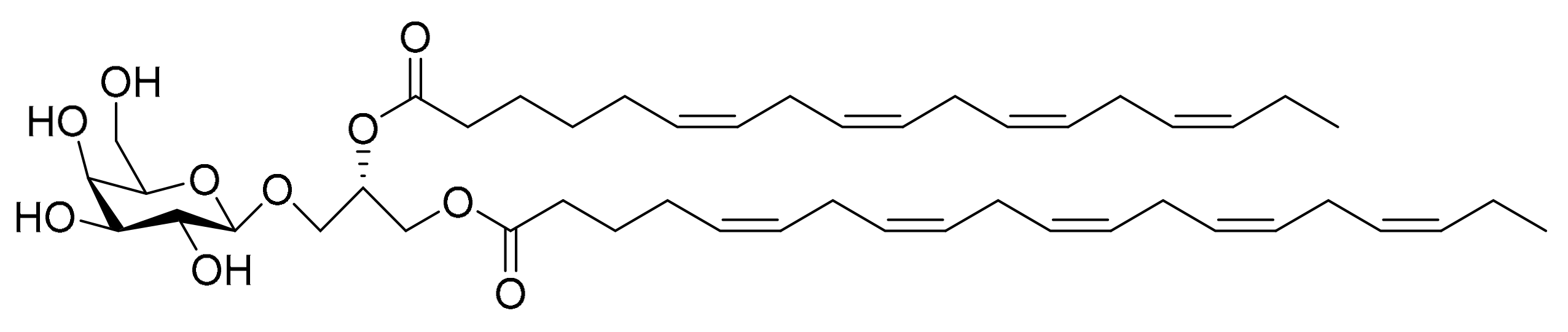
4.8. Monogalactosyl Glycerols
5. Active Compounds from Other Marine Organisms
6. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- What Is Cancer. Available online: https://www.cancer.gov/about-cancer/understanding/what-is-cancer (accessed on 12 April 2018).
- European Cancer Observatory. Available online: http://eco.iarc.fr/ (accessed on 12 April 2018).
- Dyshlovoy, S.A.; Honecker, F. Marine compounds and cancer: Where do we stand? Mar. Drugs 2015, 13, 5657–5665. [Google Scholar] [CrossRef] [PubMed]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. UK 2016, 96, 151–158. [Google Scholar] [CrossRef]
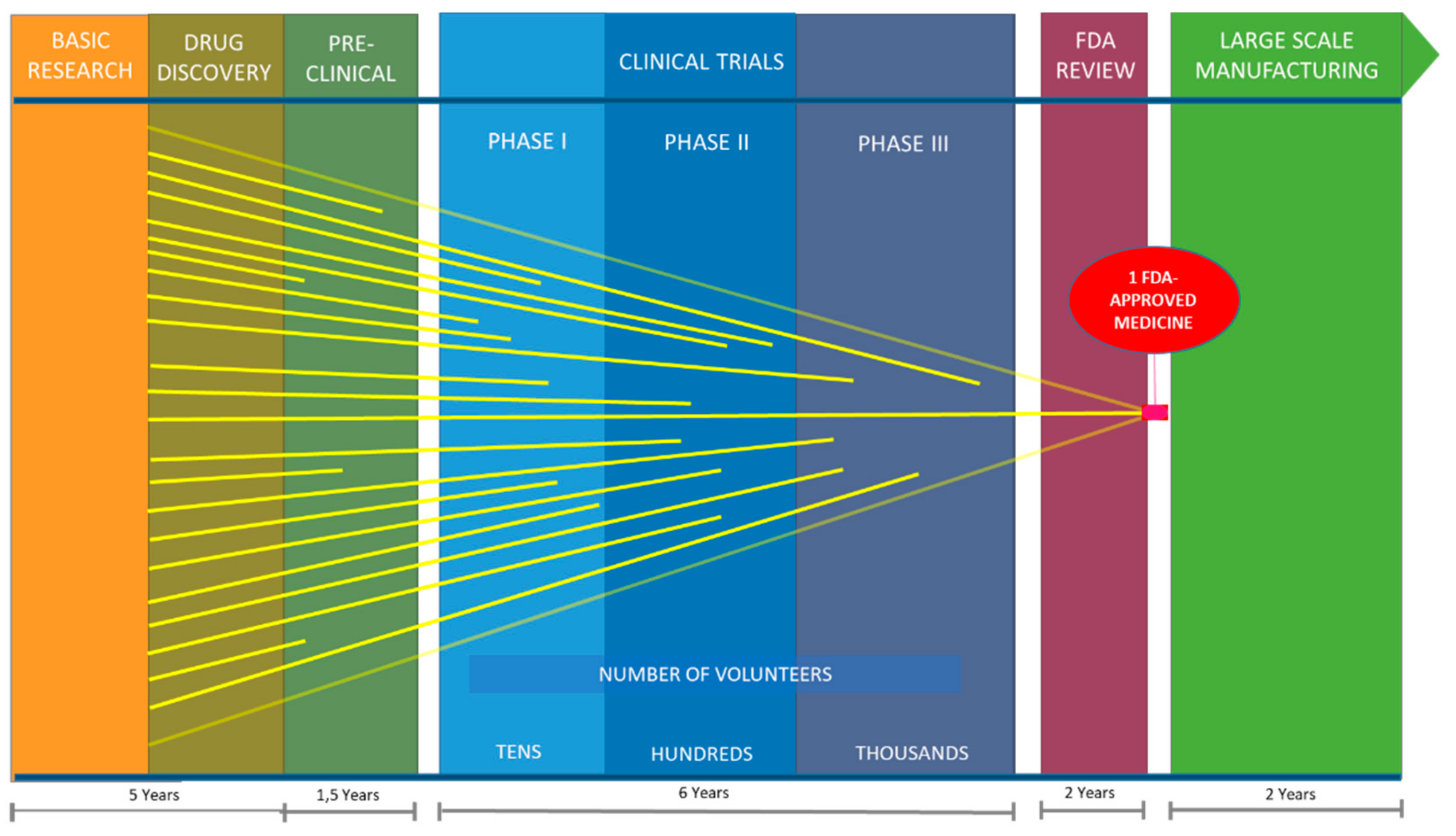
- Van Norman, G.A. Drugs, Devices, and the FDA: Part 1: An Overview of Approval Processes for Drugs. JACC Basic Transl. Sci. 2016, 1, 170–179. [Google Scholar] [CrossRef]
- Biopharmaceutical Research & Development. Available online: http://phrma-docs.phrma.org/sites/default/files/pdf/rd_brochure_022307.pdf (accessed on 12 April 2018).
- Moreno-Garrido, I. Microalgae immobilization: Current techniques and uses. Bioresour. Technol. 2008, 99, 3949–3964. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes and antibacterial activities. Front. Mar. Sci. 2016, 3, 1–12. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef] [PubMed]
- De Morais, M.G.; Vaz, B.D.S.; De Morais, E.G.; Costa, J.A.V. Biologically active metabolites synthesized by microalgae. Biomed. Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Sansone, C.; Braca, A.; Ercolesi, E.; Romano, G.; Palumbo, A.; Casotti, R.; Francone, M.; Ianora, A. Diatom-derived polyunsaturated aldehydes activate cell death in human cancer cell lines but not normal cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kwang, H.C.; Song, Y.I.K.; Lee, D.U. Antiproliferative effects of carotenoids extracted from Chlorella ellipsoidea and Chlorella vulgaris on human colon cancer cells. J. Agric. Food Chem. 2008, 56, 10521–10526. [Google Scholar] [CrossRef]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Pasquet, V.; Morisset, P.; Ihammouine, S.; Chepied, A.; Aumailley, L.; Berard, J.B.; Serive, B.; Kaas, R.; Lanneluc, I.; Thiery, V.; et al. Antiproliferative activity of violaxanthin isolated from bioguided fractionation of Dunaliella tertiolecta extracts. Mar. Drugs 2011, 9, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Nappo, M.; Berkov, S.; Massucco, C.; Di Maria, V.; Bastida, J.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Zupo, S. Apoptotic activity of the marine diatom Cocconeis scutellum and eicosapentaenoic acid in BT20 cells. Pharm. Biol. 2012, 50, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef] [PubMed]
- Nigjeh, S.E.; Yusoff, F.; Banu, N.; Alitheen, M.; Rasoli, M.; Keong, Y.S.; Rahman, A. Cytotoxic effect of ethanol extract of microalga, Chaetoceros calcitrans, and its mechanisms in inducing apoptosis in human breast cancer cell line. Biomed. Res. Int. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.H.; Alitheen, N.B.; Yusoff, F.M.; Yap, S.K.; Loh, S.P. Crude ethyl acetate extract of marine microalga, Chaetoceros calcitrans, induces Apoptosis in MDA-MB-231 breast cancer cells. Pharmacogn. Mag. 2014, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.W.; Ko, J.Y.; Shah, M.M.R.; Lee, J.H.; Kang, M.C.; O-Nam, K.; Lee, J.B.; Jeon, Y.J. In vitro studies of anti-inflammatory and anticancer activities of organic solvent extracts from cultured marine microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Shah, M.R.; Kalpa, W.S.; Ju-Young, K.; Lakmal, H.H.C.; Ji-Hyeok, L.; So-Jeong, A.; You-Jin, J.; Joon-Baek, L. Potentiality of benthic dinoflagellate cultures and screening of their bioactivities in Jeju Island, Korea. Afr. J. Biotechnol. 2014, 13, 792–805. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Li, X.-F.; Kang, K.-H.; Ryu, B.; Kim, S.-K. Stigmasterol isolated from marine microalgae Navicula incerta induces apoptosis in human hepatoma HepG2 cells. BMB Rep. 2014, 47, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Samarakoon, K.W.; Ko, J.Y.; Lee, J.H.; Kwon, O.N.; Kim, S.W.; Jeon, Y.J. Apoptotic anticancer activity of a novel fatty alcohol ester isolated from cultured marine diatom, Phaeodactylum tricornutum. J. Funct. Foods 2014, 6, 231–240. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Haramaty, L.; Vardi, A.; White, E.; Lutz, R.; Falkowski, P. Apoptosis-inducing galactolipids from a cultured marine diatom, Phaeodactylum tricornutum. J. Nat. Prod. 2008, 4883, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Somasekharan, S.P.; El-Naggar, A.; Sorensen, P.H.; Wang, Y.; Cheng, H. An aqueous extract of marine microalgae exhibits antimetastatic activity through preferential killing of suspended cancer cells and anticolony forming activity. Evid. Based Complement. Altern. Med. 2016, 2016, 9730654. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.-Y.; Tsai, C.-T.; Chuang, W.-L.; Chao, Y.-H.; Pan, I.-H.; Chen, Y.-K.; Lin, C.-C.; Wang, B.-Y. Chlorella sorokiniana induces mitochondrial-mediated apoptosis in human non-small cell lung cancer cells and inhibits xenograft tumor growth in vivo. BMC Complement. Altern. Med. 2017, 17, 88. [Google Scholar] [CrossRef] [PubMed]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and temperature effects on bioactivity in diatoms. J. Appl. Phycol. 2016, 28, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef] [PubMed]
- Dockery, N.; Higashida, K.; Verdes, R.P.; Mooneyham, T.P. Chlorella Containing Nutritional Supplement Having Improved Digestability. Patent Application US 10/795,804, 8 September 2005. [Google Scholar]
- Hildebrand, M.; Manandhar-Shrestha, K.; Abbriano, R. Effects of chrysolaminarin synthase knockdown in the diatom Thalassiosira pseudonana: Implications of reduced carbohydrate storage relative to green algae. Algal Res. 2017, 23, 66–77. [Google Scholar] [CrossRef]
- Chajès, V.; Sattler, W.; Stranzl, A.; Kostner, G.M. Influence of n-3 fatty acids on the growth of human breast cancer cells in vitro: Relationship to peroxides and vitamin-E. Breast Cancer Res. Treat. 1995, 34, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Kao, N.; Hu, J.; Wu, C. Fucoxanthin-rich brown algae extract decreases inflammation and attenuates colitis-associated colon cancer in mice. J. Food Nutr. Res. 2016, 4, 137–147. [Google Scholar] [CrossRef]
- Kadekaru, T.; Toyama, H.; Yasumoto, T. Safety evaluation of Fucoxanthin purified from Undaria pinnatifida. Nippon Shokuhin Kagaku Kogaku Kaishi 2008, 55, 304–308. [Google Scholar] [CrossRef]
- Ishikawa, C.; Tafuku, S.; Kadekaru, T.; Sawada, S.; Tomita, M.; Okudaira, T.; Nakazato, T.; Toda, T.; Uchihara, J.N.; Taira, N.; et al. Antiadult T-cell leukemia effects of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. Int. J. Cancer 2008, 123, 2702–2712. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Wanezaki, S.; Miyauchi, K.; Kurihara, H.; Kohno, H.; Kawabata, J.; Odashima, S.; Takahashi, K. Apoptosis-inducing effect of fucoxanthin on human leukemia cell line HL-60. Food Sci. Technol. Res. 1999, 5, 243–246. [Google Scholar] [CrossRef]
- Kotake-nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids affect proliferation of human prostate cancer cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.; Li, Y.; Qian, Z.J.; Kim, M.M.; Kim, S.K. Differentiation of human osteosarcoma cells by isolated phlorotannins is subtly linked to COX-2, iNOS, MMPs, and MAPK signaling: Implication for chronic articular disease. Chem. Biol. Interact. 2009, 179, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Ghielmini, M.; Colli, E.; Erba, E.; Bergamaschi, D.; Pampallona, S.; Jimeno, J.; Faircloth, G.; Sessa, C. In vitro schedule-dependency of myelotoxicity and cytotoxicity of Ecteinascidin 743 (ET-743). Ann. Oncol. 1998, 9, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Francisco, J.A.; Cerveny, C.G.; Meyer, D.L.; Mixan, B.J.; Klussman, K.; Chace, D.F.; Rejniak, S.X.; Gordon, K.A.; DeBlanc, R.; Toki, B.E.; et al. cAC10-vcMMAE, an anti-CD30-monomethyl auristatin E conjugate with potent and selective antitumor activity. Blood 2003, 102, 1458–1465. [Google Scholar] [CrossRef] [PubMed]
- Towle, M.J.; Salvato, K.A.; Budrow, J.; Wels, B.F.; Kuznetsov, G.; Aalfs, K.K.; Welsh, S.; Zheng, W.; Seletsky, B.M.; Palme, M.H.; et al. In vitro and in vivo anticancer activities of synthetic macrocyclic ketone analogues of halichondrin B. Cancer Res. 2001, 61, 1013–1021. [Google Scholar] [PubMed]
- Desai, U.; Shah, K.; Mirza, S.; Panchal, D.; Parikh, S.; Rawal, R. Enhancement of the cytotoxic effects of Cytarabine in synergism with Hesperidine and Silibinin in Acute Myeloid Leukemia: An in-vitro approach. J. Cancer Res. Ther. 2015, 11, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Bandi, M.; Raje, N.; Richardson, P.; Palladino, M.A.; Anderson, K.C.; Singh, A.V.; Bandi, M.; Raje, N.; Richardson, P.; et al. A novel vascular disrupting agent plinabulin triggers JNK-mediated apoptosis and inhibits angiogenesis in multiple myeloma cells A novel vascular disrupting agent plinabulin triggers JNK-mediated apoptosis and inhibits angiogenesis in multiple myeloma cells. Blood 2011, 117, 5692–5700. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Fabre, P.M.; De Pedro, E.; Medina, M.A.; Núñez De Castro, I.; Márquez, J. Polyamine contents of human breast cancer cells treated with the cytotoxic agents chlorpheniramine and dehydrodidemnin B. Cancer Lett. 1997, 113, 141–144. [Google Scholar] [CrossRef]
- Takahashi, R.; Mabuchi, S.; Kawano, M.; Sasano, T.; Matsumoto, Y.; Kuroda, H.; Kozasa, K.; Hashimoto, K.; Sawada, K.; Kimura, T. Preclinical investigations of PM01183 (Lurbinectedin) as a single agent or in combination with other anticancer agents for clear cell carcinoma of the ovary. PLoS ONE 2016, 11, e0151050. [Google Scholar] [CrossRef] [PubMed]
- Monomethyl Auristatin E (MMAE). Available online: https://adcreview.com/adc-university/adcs-101/cytotoxic-agents/monomethyl-auristatin-e-mmae/ (accessed on 12 April 2018).
- Cuevas, C.; Francesch, A. Development of Yondelis® (trabectedin, ET-743). A semisynthetic process solves the supply problem. Nat. Prod. Rep. 2009, 26, 322. [Google Scholar] [CrossRef] [PubMed]
- Amaro, H.; Guedes, A.; Malcata, F. Antimicrobial activities of microalgae: An invited review. Sci. Microb. Pathog. Commun. Curr. Res. Technol. Adv. 2011, 1272–1280. [Google Scholar] [CrossRef]
- Paul, C.; Reunamo, A.; Lindehoff, E.; Bergkvist, J.; Mausz, M.A.; Larsson, H.; Richter, H.; Wängberg, S.Å.; Leskinen, P.; Bam̊stedt, U.; et al. Diatom derived polyunsaturated aldehydes do not structure the planktonic microbial community in a mesocosm study. Mar. Drugs 2012, 10, 775–792. [Google Scholar] [CrossRef] [PubMed]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.J.; Chang, J.S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, Y.; Zhang, Y.; Zhang, S.; Qu, J.; Wang, X.; Kong, R.; Han, C.; Liu, Z. Fucoxanthin: A promising medicinal and nutritional ingredient. Evid. Based Complement. Altern. Med. 2015, 2015, 723515. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Su, P.; Zhang, W. Advances in microalgae-derived phytosterols for functional food and pharmaceutical applications. Mar. Drugs 2015, 13, 4231–4254. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Jafri, M.; Kar, A.; Meheta, B.K. Thyroid inhibitory, antiperoxidative and hypoglycemic effects of stigmasterol isolated from Butea monosperma. Fitoterapia 2009, 80, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S. The Story of Beta-sitosterol—A Review. Eur. J. Med. Plants 2014, 4, 590–609. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgae | Fraction/Compound | Target Cells | Active Concentration | Reference |
|---|---|---|---|---|
| Thalassiosira rotula, Skeletonema costatum and Pseudonitzschia delicatissima. Commercial source, not from microalgae | Polyunsaturated aldehydes (PUAs) | Colon adenocarcinoma (Caco-2) Lung adenocarcinoma (A549) Colon adenocarcinoma (COLO 205) | 11 to 17 µg/mL (arrest of cell growth) 0.22 to 1.5 µg/mL (CV of 80% to 0% depending on the conditions) | [11] [12] |
| Chlorella ellipsoidea | Carotenoid extract | Colon carcinoma (HCT-116) | 40 µg/mL (IC50) | [13] |
| Synedra acus | Chrysolaminaran (polysaccharide) | Colorectal adenocarcinoma (HT-29 and DLD-1) | 54.5 and 47.7 µg/mL (IC50 for HT-29 and DLD-1) | [14] |
| Dunaliella tertiolecta | Violaxanthin (carotenoid already identified in C. ellipsoidea) | Breast adenocarcinoma (MCF-7) | 40 µg/mL (to observe cytostatic activity) | [15] |
| Cocconeis scutellum | Eicosapentaenoic acid (EPA) | Breast carcinoma (BT20) | Not clarified | [16] |
| Chaetoseros sp., Cylinrotheca closterium, Odontella aurita and Phaeodactylum tricornutum | Fucoxanthin (carotenoid) | Promyelocytic leukemia (HL-60), Caco-2, colon adenocarcinoma (HT-29), DLD-1 and prostate cancer (PC-3, DU145 and LNCaP) | 29.78 µg/mL (CV of 17.3% for HL-60) 10.01 µg/mL (CV of 14.8%, 29.4% and 50.8% for Caco-2, DLD-1 and HT-29) 13.18 µg/mL (CV of 14.9%, 5.0% and 9.8% for PC-3, DU145 and LNCaP) | [17] |
| Chaetoceros calcitrans | EtOH extract AcOEt extract | MCF-7 Breast adenocarcinoma (MDA-MB-231) | 3.00 µg/mL (IC50) 60 µg/mL (IC50) | [18] [19] |
| Amphidinium carterae | CH3Cl fraction Hexane fraction AcOEt fraction | HL-60 HL60, Skin melanoma (B16F10), A549 | 50 µg/mL (CV of 40%) 25–50 µg/mL (CV between 50% and 90%) | [20] |
| Eleven strains of benthic diatoms Ostreopsis ovata Amphidinium operculatum | MeOH extract | HL-60 | 50 µg/mL (CV of 48% for O. ovata and 58% for A. operculatumi) | [21] |
| Navicula incerta | Stigmasterol (phytosterol) | Liver hepatocellular carcinoma (HepG2) | 8.25 μg/mL (CV of 54%) | [22] |
| Phaeodactylum tricornutum | Nonyl-8-acetoxy-6-methyloctanoate (NAMO, fatty alcohol ester) Monogalactosyl glycerols 1 | HL-60 Mouse epithelial cell lines (W2, D3) | 22.3 μg/mL (IC50) 40-50 μg/mL (concentration necessary to induce apoptosis) | [23] [24] |
| Skeletonema costatum Skeletonema marinoi | Hydrophobic fraction and PUAs Hydrophobic fraction | Caco-2 (A2058 not affected) Skin melanoma (A2058) | 11 to 17 µg/mL (PUAs) 50 µg/mL (CV of 60%) | [11] [8] |
| Canadian marine microalgal pool | Aqueous extract | A549, lung carcinoma (H460), prostate carcinoma (PC-3, DU145), stomach carcinoma (N87), MCF-7, pancreas adenocarcinoma (BxPC-3) and osteosarcoma (MNNG) | 5000 µg/mL (CV between 30% and 80% depending on the cell line) | [25] |
| Chlorella sorokiniana | Aqueous extract | A549 and lung adenocarcinoma (CL1-5) | 0.0156 to 1 µg/mL (CV reduced down to 20% progressively) | [26] |
| Microalgae | Source | Culturing Conditions | Harvesting Time | Reference |
|---|---|---|---|---|
| Synedra acus | Lake Baikal | Culture medium consisting of (mg/L) Ca(NO3)2·4H2O (20), KH2PO4 (2), MgSO4 (12), NaHCO3 (30), Na2EDTA (2.2), H3BO3 (2.4), MnCl2·4H2O (1.3), (NH4)6Mo7O24·4H2O (1), Na2SiO3·9H2O (25), FeCl3 (1.6), cyanocobalamine (0.04), thiamine (0.04), and biotin (0.04). 12 °C and 250–300 µmol·m−2·s−1 light intensity. | Not provided | [14] |
| Dunaliella tertiolecta | DT strain CCMP364 (NCMA, USA) | Conway medium. 20 °C, 180 μmol·m−2·s−1 light intensity. | Late exponential phase | [15] |
| Cocconeis scutellum | Mediterranean Sea, Stazione Zoologica A. Dohrn | Guillard’s F/2 medium.18 °C, 140 µmol·m−2·s−1 light intensity and 12 h:12 h photoperiod. | Not provided | [16] |
| Chaetoceros calcitrans | Strain UPMAAHU10 University Putra Malaysi | Conway medium. 24 °C, 120 μmol·m−2·s−1 light intensity, automatic oscillating shaker at 110 rpm and harvested at stationary phase (6–7 days). Conway medium. Conditions not provided. | Stationary phase Not provided | [18] [19] |
| Amphidinium carterae | Korea Marine Microalgae Culture Center | Conway medium. 20 °C, 34 μmol·m−2·s−1 light intensity and 24 h:0 h photoperiod. | Days 8–10 | [20] |
| Eleven strains of benthic dinoflagellates | Coast of Jeju Island (Korea) | Daigo IMK medium (Nihon Pharmaceutical Co., Ltd.) and Guillard’s F/2 medium. 20 °C, 180 μmol·m−2·s−1 light intensity and 12 h:12 h photoperiod. | Exponential phase. | [21] |
| Navicula incerta | Korea Marine Microalgae Culture Center. | Guillard’s F/2 medium. Conditions not provided. | Not provided | [22] |
| Phaeodactylum tricornutum | Korea Marine Microalgae Culture Center Provasoli-Guillard National Center | Conway medium. 20 °C, 34 μmol·m−2·s−1 light intensity and 24 h:0 h photoperiod. Guillard’s F/2 medium. 18 °C and 100 μmol·m−2·s−1 light intensity. | Days 8–10 Not provided | [23] [24] |
| Skeletonema marinoi FE6 (1997) FE60 (2005) | Adriatic Sea (Mediterranean Sea) | Guillard’s F/2 medium. 19 °C, 100 μmol·m−2·s−1 light intensity and 12 h:12 h photoperiod. | Late stationary phase | [8] |
| Marine Organism | Compound | Target | Active Concentration | Reference |
|---|---|---|---|---|
| Ecteinascidia turbinata | Ecteinascidin/ Trabectedin (alkaloid) | MFC7 A549 | 0.6 ng/mL (IC70) 5.6 ng/mL (IC70) | [38] |
| Dolabella auricularia/Symploca sp. VP642 | Brentuximab vedotin (antibody-drug conjugate) | Non-Hodgkin’s lymphoma cells (Karpas 299) | 2.5 ng/mL (IC50) | [39] |
| Halichondria okadai | Eribulin mesylate (macrolide) | DLD-1 LNCaP HL-60 | 6.934 ng/mL (IC50) 0.365 ng/mL (IC50) 0.657 ng/mL (IC50) | [40] |
| Cryptotheca crypta | Cytarabine (nucleoside) | Acute Myeloid Leukemia (AML) cells | 272 ng/mL (IC50) | [41] |
| Marine Organism | Compound | Target | Active Concentration | Reference |
|---|---|---|---|---|
| Aspergillus sp. CNC139 | Plinabulin (diketopiperazine) | Multiple myeloma cells (MM.1S, MM.1R, RPMI8226, and INA-6) | 2.7 to 3.375 ng/mL (IC50) | [42] |
| Aplidium albicans | Plitidepsin (depsipeptide) | MCF-7 | 55.5 ng/mL (IC50) | [43] |
| Halichondria okadai | Lurbinectedin (alkaloid) | Ovarian cancer cells (RMG1, RMG2, KOC7C, HAC2, A2780, HeyA8 and SKOV-3) | 0.78 to 2.34 ng/mL (IC50) | [44] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. https://doi.org/10.3390/md16050165
Martínez Andrade KA, Lauritano C, Romano G, Ianora A. Marine Microalgae with Anti-Cancer Properties. Marine Drugs. 2018; 16(5):165. https://doi.org/10.3390/md16050165
Chicago/Turabian StyleMartínez Andrade, Kevin A., Chiara Lauritano, Giovanna Romano, and Adrianna Ianora. 2018. "Marine Microalgae with Anti-Cancer Properties" Marine Drugs 16, no. 5: 165. https://doi.org/10.3390/md16050165
APA StyleMartínez Andrade, K. A., Lauritano, C., Romano, G., & Ianora, A. (2018). Marine Microalgae with Anti-Cancer Properties. Marine Drugs, 16(5), 165. https://doi.org/10.3390/md16050165