
Acute Exhaustive Exercise under Normoxic and Normobaric Hypoxic Conditions Differentially Regulates Angiogenic Biomarkers in Humans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
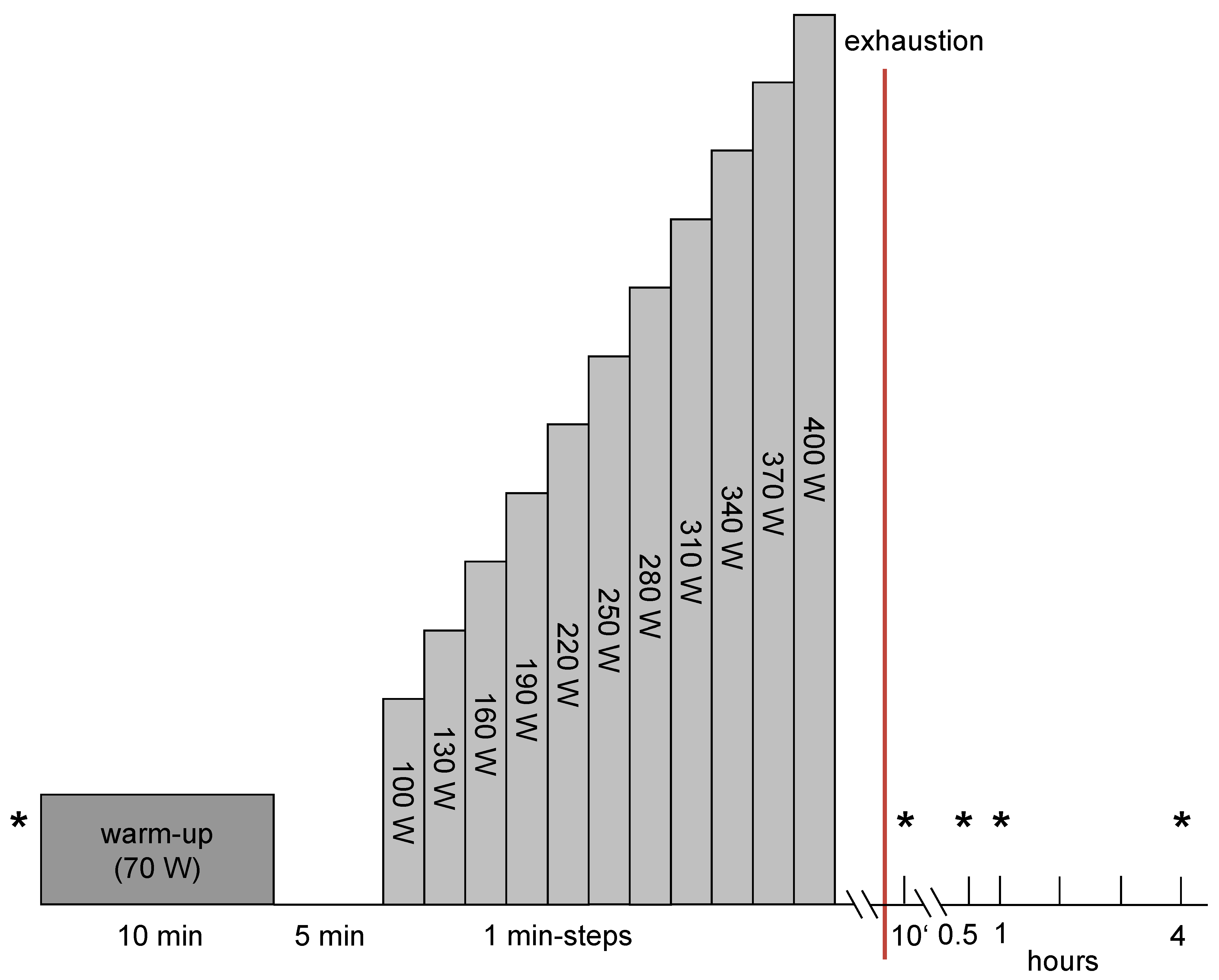
2.3. Exercise Protocol
2.4. Measurement of VEGF, MMP-2, MMP-9 and Endostatin Serum Levels
2.5. Statistics
3. Results
3.1. Pmax and Relative VO2max
3.2. SaO2
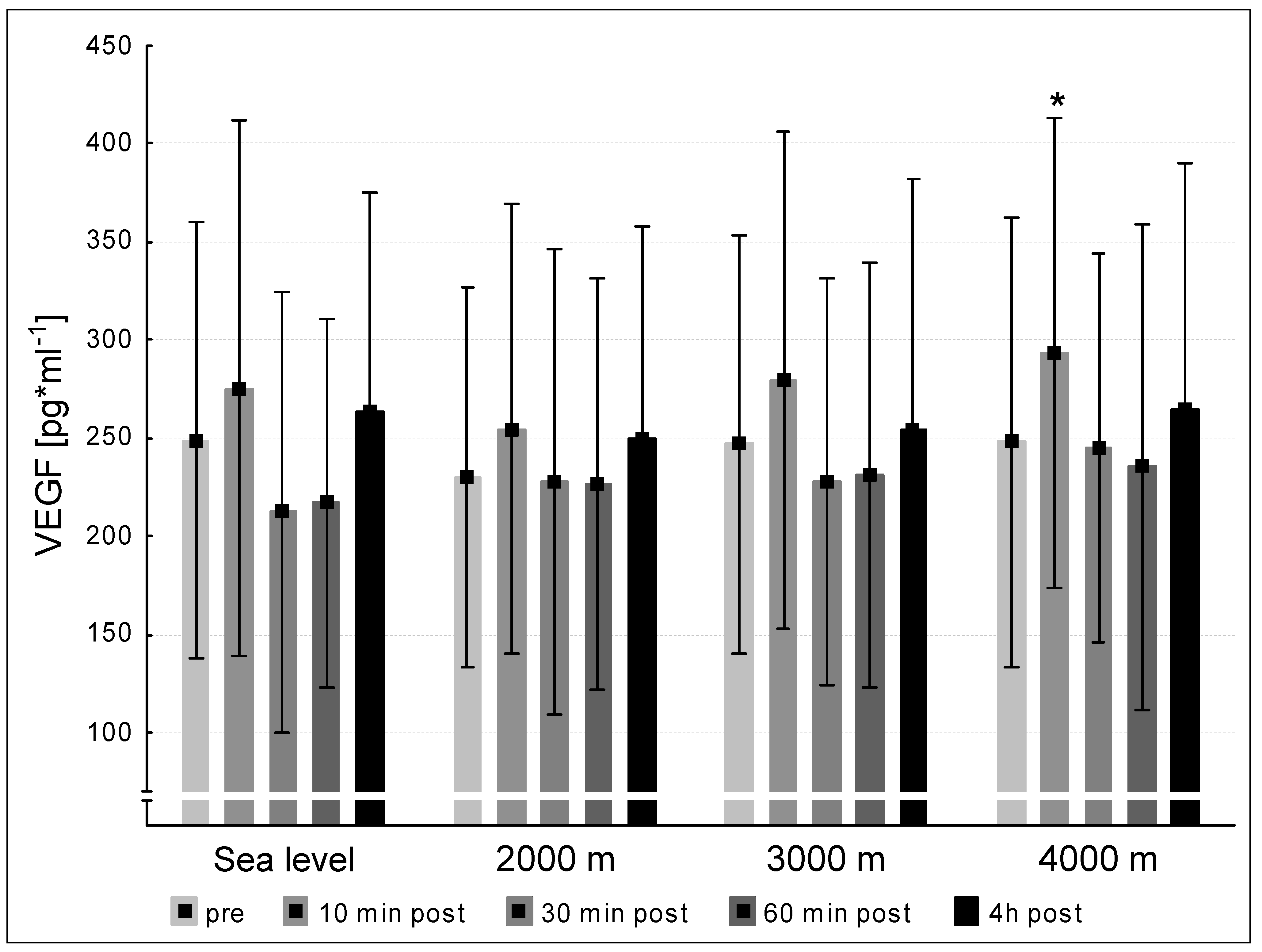
3.3. VEGF
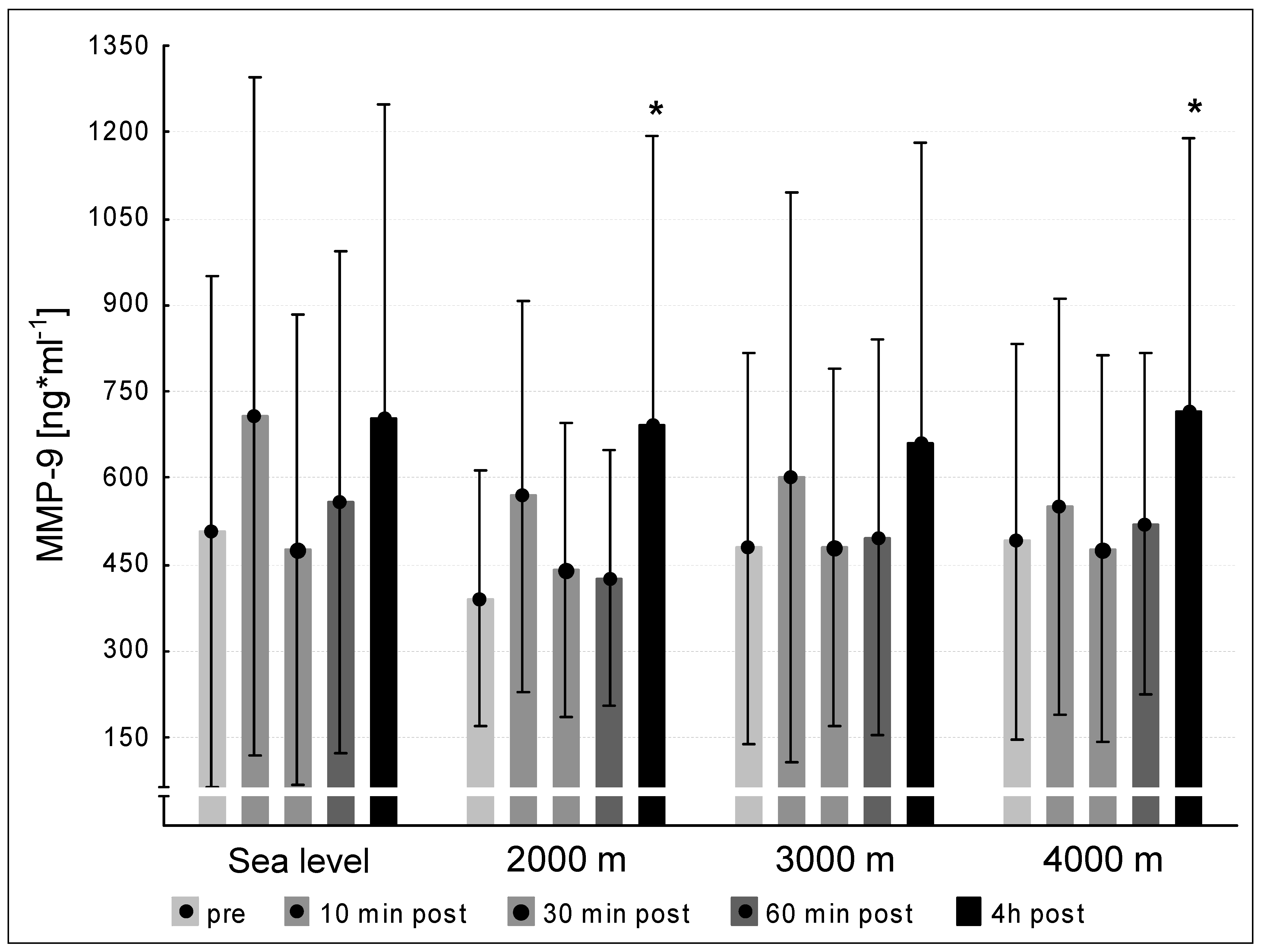
3.4. MMP-2 and -9
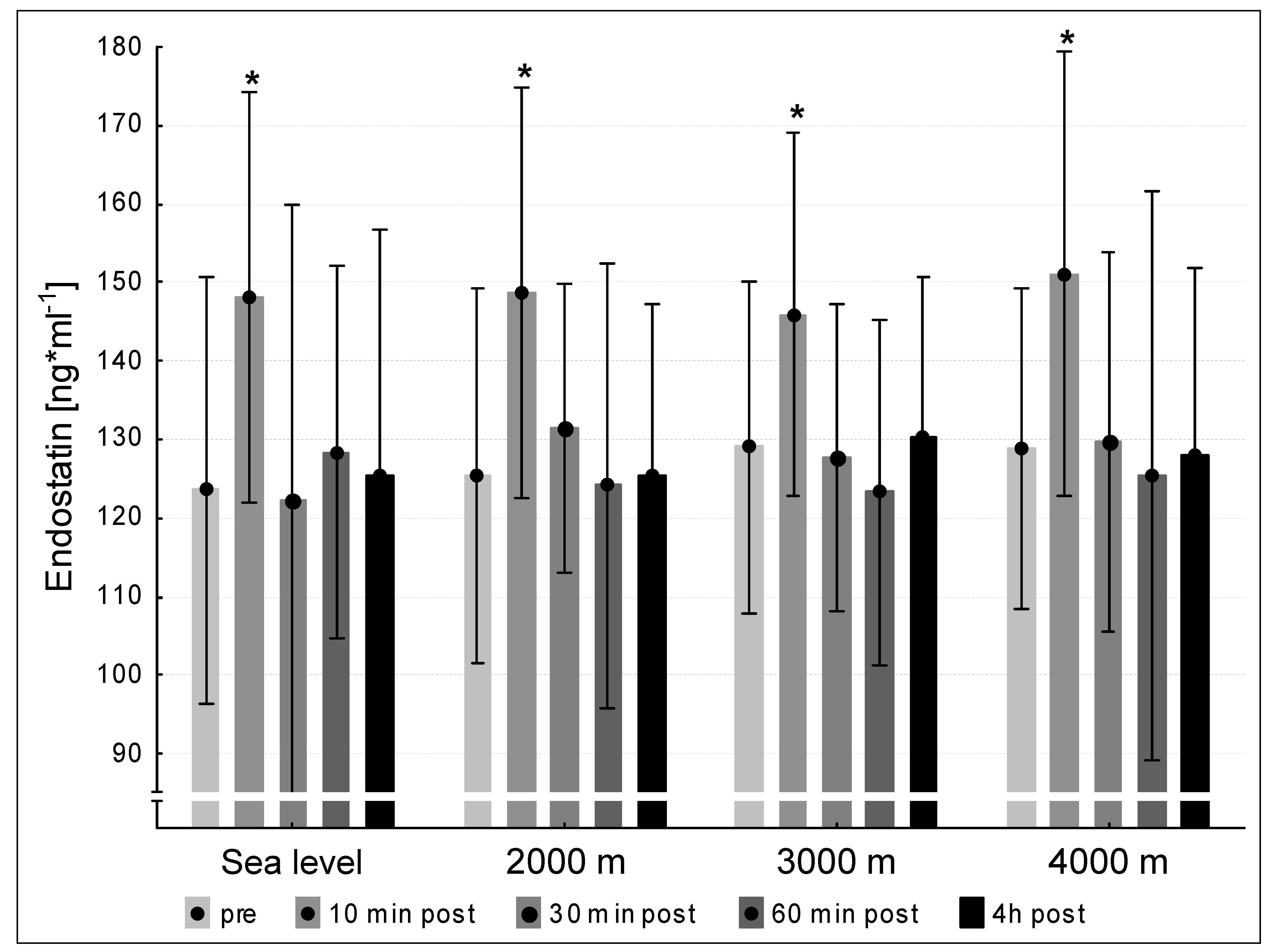
3.5. Endostatin
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hargreaves, M. Skeletal muscle metabolism during exercise in humans. Clin. Exp. Pharmacol. Physiol. 2000, 27, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. Angiogenesis in life, disease and medicine. Nature 2005, 438, 932–936. [Google Scholar] [CrossRef] [PubMed]
- Prior, B.M.; Yang, H.T.; Terjung, R.L. What makes vessels grow with exercise training? J. Appl. Physiol. 2004, 97, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, D.; Schmidt, A.; Reimann, K.; Hescheler, J.; Pfitzer, G.; Bloch, W.; Fleischmann, B.K. Endostatin, the proteolytic fragment of collagen XVIII, induces vasorelaxation. Circ. Res. 2006, 98, 1203–1211. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P.; Ferreira, V.; Breier, G.; Pollefeyt, S.; Kieckens, L.; Gertsenstein, M.; Fahrig, M.; Vandenhoeck, A.; Harpal, K.; Eberhardt, C.; et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nat. Cell Biol. 1996, 380, 435–439. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix Metalloproteinases: Regulators of the Tumor Microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Suhr, F.; Brixius, K.; Bloch, W. Angiogenic and vascular modulation by extracellular matrix cleavage products. Curr. Pharm. Des. 2009, 15, 389–410. [Google Scholar] [CrossRef]
- Hynes, R.O. The extracellular matrix: Not just pretty fibrils. Science 2009, 326, 1216–1219. [Google Scholar] [CrossRef] [Green Version]
- Faye, C.; Chautard, E.; Olsen, B.R.; Ricard-Blum, S. The first draft of the endostatin interaction network. J. Biol. Chem. 2009, 284, 22041–22047. [Google Scholar] [CrossRef] [Green Version]
- Suhr, F.; Brixius, K.; De Marées, M.; Bölck, B.; Kleinöder, H.; Achtzehn, S.; Bloch, W.; Mester, J. Effects of short-term vibration and hypoxia during high-intensity cycling exercise on circulating levels of angiogenic regulators in humans. J. Appl. Physiol. 2007, 103, 474–483. [Google Scholar] [CrossRef]
- Suhr, F.; Rosenwick, C.; Vassiliadis, A.; Bloch, W.; Brixius, K. Regulation of extracellular matrix compounds involved in angiogenic processes in short- and long-track elite runners. Scand. J. Med. Sci. Sports 2009, 20, 441–448. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The biology of VEGF and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.-W.; Gadonski, G.; Wang, J.; Makey, I.; Adair, T.H. Exercise increases endostatin in circulation of healthy volunteers. BMC Physiol. 2004, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Kraus, R.M.; Stallings, H.W.; Yeager, R.C.; Gavin, T.P. Circulating plasma VEGF response to exercise in sedentary and endurance-trained men. J. Appl. Physiol. 2004, 96, 1445–1450. [Google Scholar] [CrossRef]
- Birot, O.J.G.; Koulmann, N.; Peinnequin, A.; Bigard, X.A. Exercise-induced expression of vascular endothelial growth factor mRNA in rat skeletal muscle is dependent on fibre type. J. Physiol. 2003, 552, 213–221. [Google Scholar] [CrossRef]
- Brown, M.; Hudlicka, O. Modulation of physiological angiogenesis in skeletal muscle by mechanical forces: Involvement of VEGF and metalloproteinases. Angiogenesis 2003, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Mester, J. A model analysis of internal loads, energetics, and effects of wobbling mass during the whole-body vibration. J. Biomech. 2002, 35, 639–647. [Google Scholar] [CrossRef]
- Berceli, S.A.; Jiang, Z.; Klingman, N.V.; Schultz, G.S.; Ozaki, C.K. Early Differential MMP-2 and -9 Dynamics During Flow-Induced Arterial and Vein Graft Adaptations. J. Surg. Res. 2006, 134, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Zatterstrom, U.K.; Felbor, U.; Fukai, N.; Olsen, B.R. Collagen XVIII/endostatin structure and functional role in angiogenesis. Cell Struct. Funct. 2000, 25, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Koskinen, S.O.A.; Hoyhtya, M.; Turpeenniemi-Hujanen, T.; Martikkala, V.; Makinen, T.T.; Oksa, J.; Rintamaki, H.; Lofberg, M.; Somer, H.; Takala, T.E.S. Serum concentrations of collagen degrading enzymes and their inhibitors after downhill running. Scand. J. Med. Sci. Sports 2001, 11, 9–15. [Google Scholar] [CrossRef]
- Koskinen, S.O.A.; Heinemeier, K.M.; Olesen, J.L.; Langberg, H.; Kjaer, M. Physical exercise can influence local levels of matrix metalloproteinases and their inhibitors in tendon-related connective tissue. J. Appl. Physiol. 2004, 96, 861–864. [Google Scholar] [CrossRef] [Green Version]
- Bergers, G.; Brekken, R.; McMahon, G.; Vu, T.H.; Itoh, T.; Tamaki, K.; Tanzawa, K.; Thorpe, P.; Itohara, S.; Werb, Z.; et al. Matrix metalloproteinase-9 triggers the angiogenic switch during carcinogenesis. Nat. Cell Biol. 2000, 2, 737–744. [Google Scholar] [CrossRef]
- Bloch, W.; Huggel, K.; Sasaki, T.; Grose, R.; Bugnon, P.; Addicks, K.; Timpl, R.; Werner, S. The angiogenesis inhibitor endostatin impairs blood vessel maturation during wound healing. FASEB J. 2000, 14, 2373–2376. [Google Scholar] [CrossRef]
- Pufe, T.; Petersen, W.J.; Miosge, N.; Goldring, M.B.; Mentlein, R.; Varoga, D.J.; Tillmann, B.N. Endostatin/collagen XVIII—An inhibitor of angiogenesis—Is expressed in cartilage and fibrocartilage. Matrix Biol. 2004, 23, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Addicks, K.; Bloch, W. Opposite effects of endostatin on different endothelial cells. Cancer Biol. Ther. 2004, 3, 1162–1166. [Google Scholar] [CrossRef] [Green Version]
- Wickström, S.A.; Alitalo, K.; Keski-Oja, J. Endostatin signaling and regulation of endothelial cell-matrix interactions. Adv. Cancer Res. 2005, 94, 197–229. [Google Scholar] [CrossRef]
- Paddenberg, R.; Faulhammer, P.; Goldenberg, A.; Kummer, W. Hypoxia-induced increase of endostatin in murine aorta and lung. Histochem. Cell Biol. 2006, 125, 497–508. [Google Scholar] [CrossRef]
- Timpl, R. Macromolecular organization of basement membranes. Curr. Opin. Cell Biol. 1996, 8, 618–624. [Google Scholar] [CrossRef]
- Kyparos, A.; Riganas, C.; Nikolaidis, M.G.; Sampanis, M.; Koskolou, M.D.; Grivas, G.; Kouretas, D.; Vrabas, I.S. The effect of exercise-induced hypoxemia on blood redox status in well-trained rowers. Graefe’s Arch. Clin. Exp. Ophthalmol. 2011, 112, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, M.J.; Almar, M.; García-Glez, J.C.; García-López, D.; De Paz, J.A.; Alvear-Ordenes, I.; González-Gallego, J. Changes in oxidative stress markers and NF-κB activation induced by sprint exercise. Free Radic. Res. 2005, 39, 431–439. [Google Scholar] [CrossRef]
- Pialoux, V.; Mounier, R.; Rock, E.; Mazur, A.; Schmitt, L.; Richalet, J.-P.; Robach, P.; Brugniaux, J.; Coudert, J.F.; Fellmann, N. Effects of the “live high-train low” method on prooxidant/antioxidant balance on elite athletes. Eur. J. Clin. Nutr. 2008, 63, 756–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pialoux, V.; Mounier, R.; Brugniaux, J.V.; Rock, E.; Mazur, A.; Richalet, J.-P.; Robach, P.; Coudert, J.; Fellmann, N. Thirteen days of “live high-train low” does not affect prooxidant/antioxidant balance in elite swimmers. Eur. J. Appl. Physiol. 2009, 106, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Pialoux, V.; Brugniaux, J.; Rock, E.; Mazur, A.; Schmitt, L.; Richalet, J.-P.; Robach, P.; Clottes, E.; Coudert, J.; Fellmann, N.; et al. Antioxidant status of elite athletes remains impaired 2 weeks after a simulated altitude training camp. Eur. J. Nutr. 2010, 49, 285–292. [Google Scholar] [CrossRef]
- Tominaga, T.; Ma, S.; Sugama, K.; Kanda, K.; Omae, C.; Choi, W.; Hashimoto, S.; Aoyama, K.; Yoshikai, Y.; Suzuki, K. Changes in urinary biomarkers of organ damage, inflammation, oxidative stress, and bone turnover following a 3000-m time trial. Antioxidants 2021, 10, 79. [Google Scholar] [CrossRef]
- Sugama, K.; Suzuki, K.; Yoshitani, K.; Shiraishi, K.; Miura, S.; Yoshioka, H.; Mori, Y.; Kometani, T. Changes of thioredoxin, oxidative stress markers, inflammation and muscle/renal damage following intensive endurance exercise. Exerc. Immunol. Rev. 2015, 21, 130–142. [Google Scholar] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nat. Cell Biol. 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Schröder, K. Redox Control of Angiogenesis. Antioxidants Redox Signal. 2019, 30, 960–971. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suhr, F.; Knuth, S.; Achtzehn, S.; Mester, J.; de Marees, M. Acute Exhaustive Exercise under Normoxic and Normobaric Hypoxic Conditions Differentially Regulates Angiogenic Biomarkers in Humans. Medicina 2021, 57, 727. https://doi.org/10.3390/medicina57070727
Suhr F, Knuth S, Achtzehn S, Mester J, de Marees M. Acute Exhaustive Exercise under Normoxic and Normobaric Hypoxic Conditions Differentially Regulates Angiogenic Biomarkers in Humans. Medicina. 2021; 57(7):727. https://doi.org/10.3390/medicina57070727
Chicago/Turabian StyleSuhr, Frank, Sarah Knuth, Silvia Achtzehn, Joachim Mester, and Markus de Marees. 2021. "Acute Exhaustive Exercise under Normoxic and Normobaric Hypoxic Conditions Differentially Regulates Angiogenic Biomarkers in Humans" Medicina 57, no. 7: 727. https://doi.org/10.3390/medicina57070727
APA StyleSuhr, F., Knuth, S., Achtzehn, S., Mester, J., & de Marees, M. (2021). Acute Exhaustive Exercise under Normoxic and Normobaric Hypoxic Conditions Differentially Regulates Angiogenic Biomarkers in Humans. Medicina, 57(7), 727. https://doi.org/10.3390/medicina57070727