
Comparison and Performance Validation of Calculated and Established Anaerobic Lactate Thresholds in Running



Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Design
2.3. Isokinetic Sprint Test and VLamax Determination (Performance Capacity of Glycolysis)
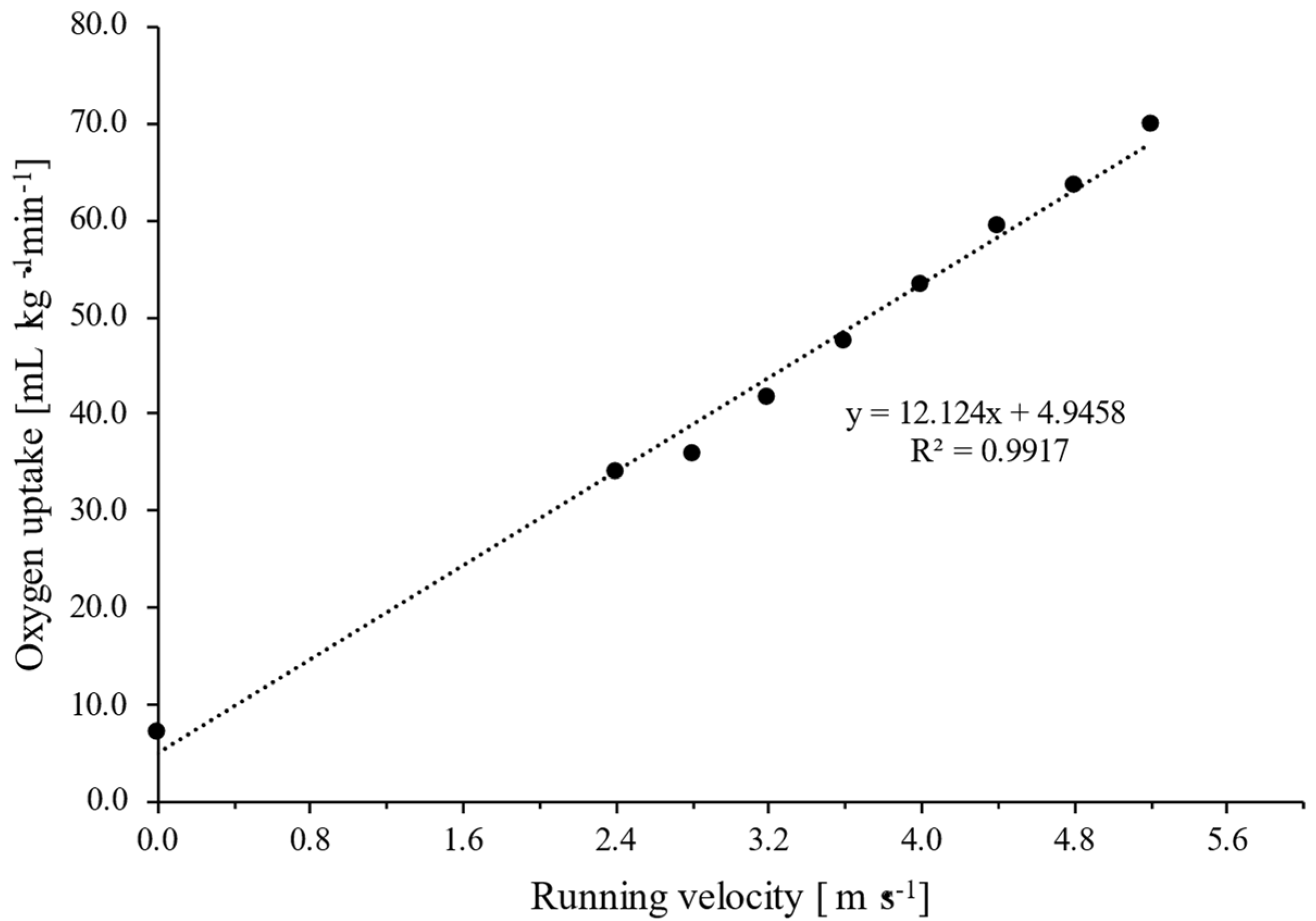
2.4. Graded Exercise Running Test and VO2max and LTAn Determination
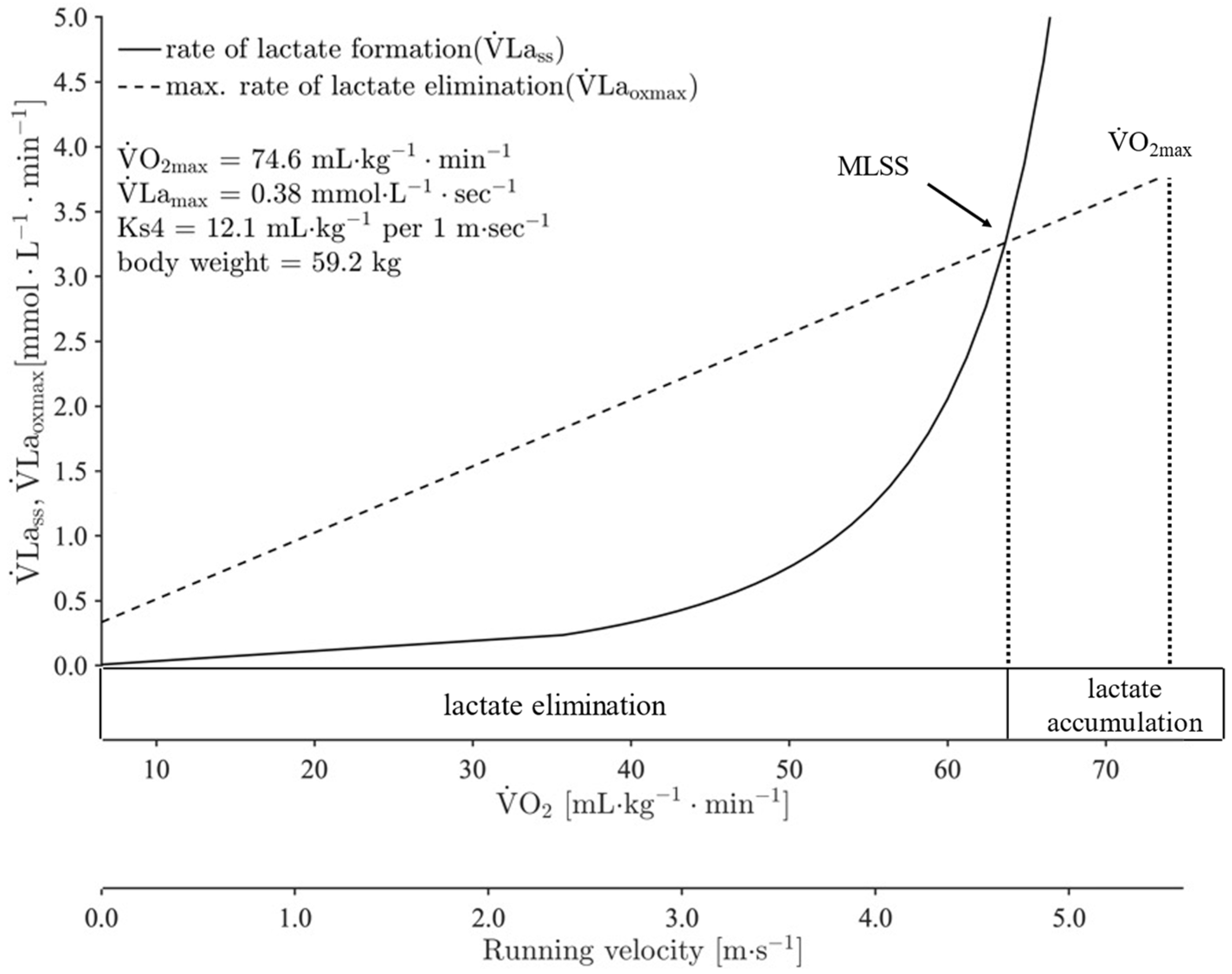
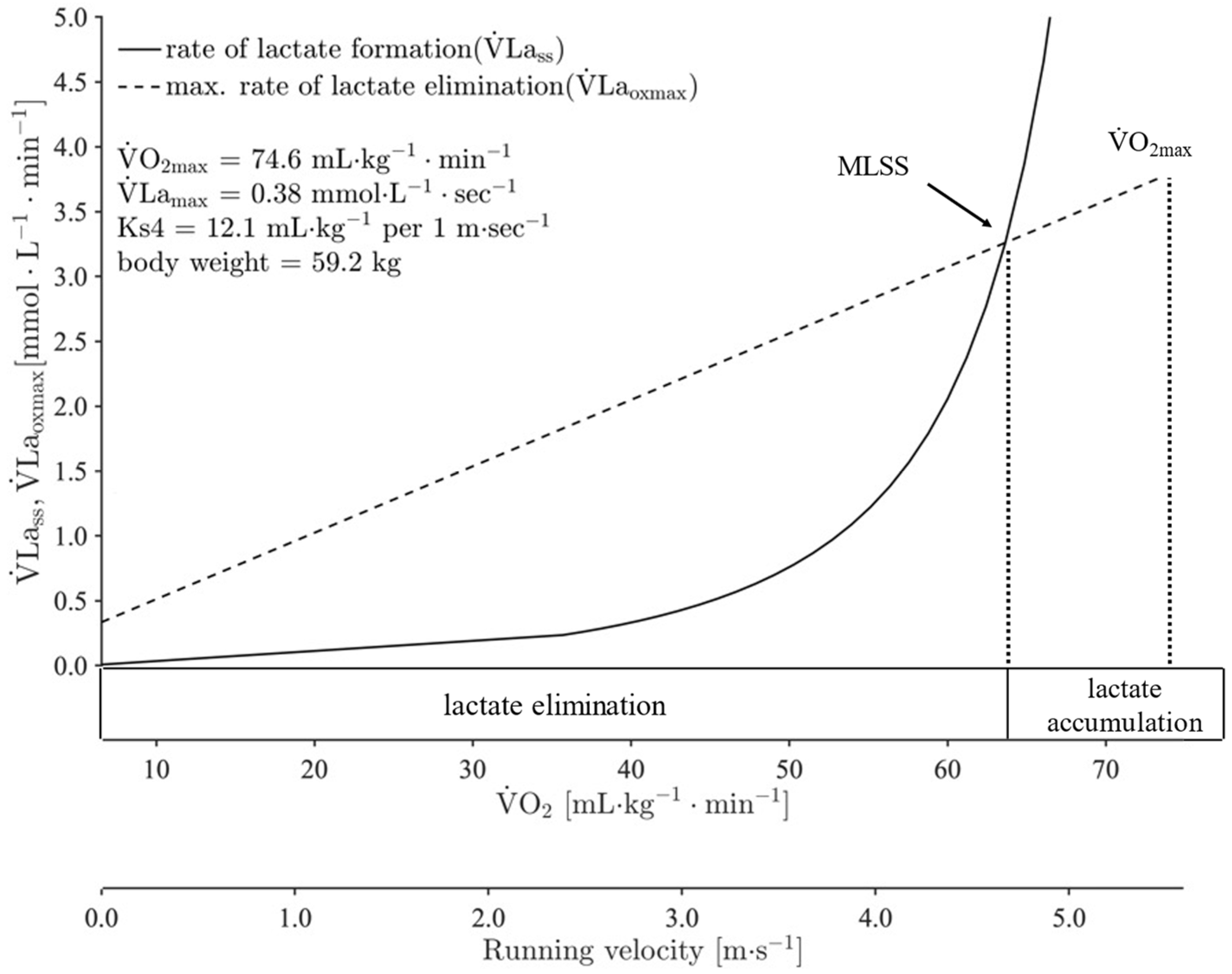
2.5. Calculation of Running Velocity at cLTAn
2.6. Statistical Analysis
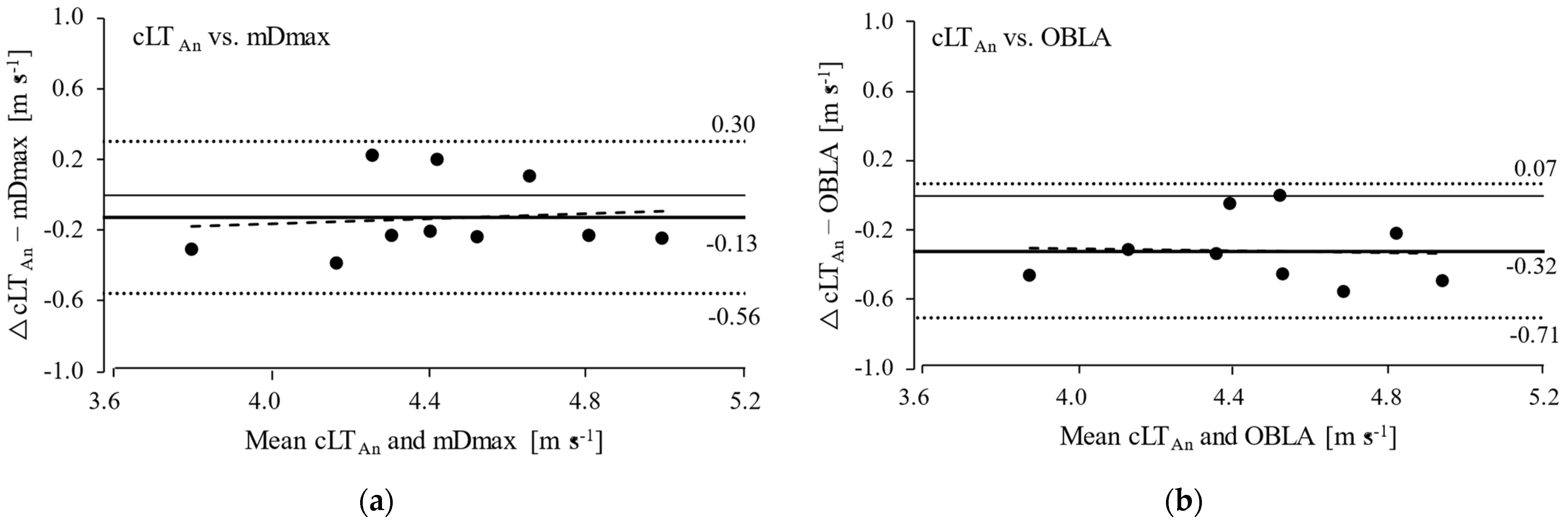
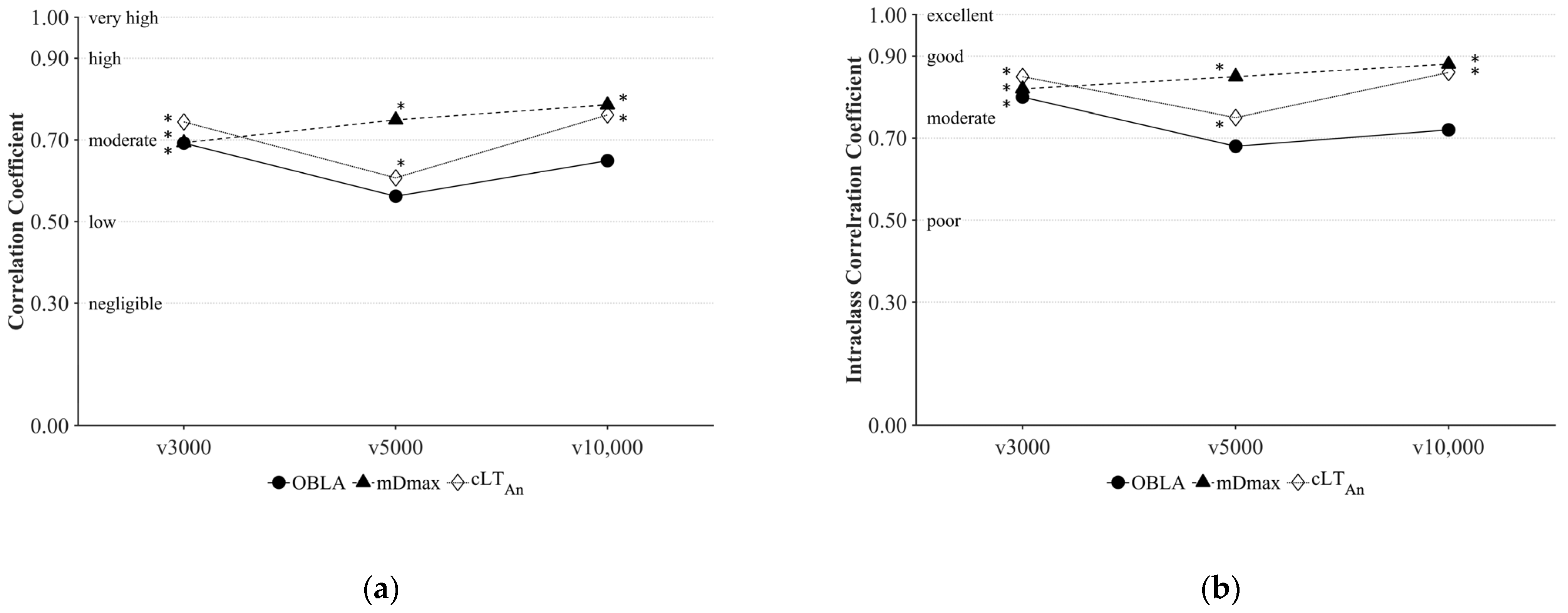
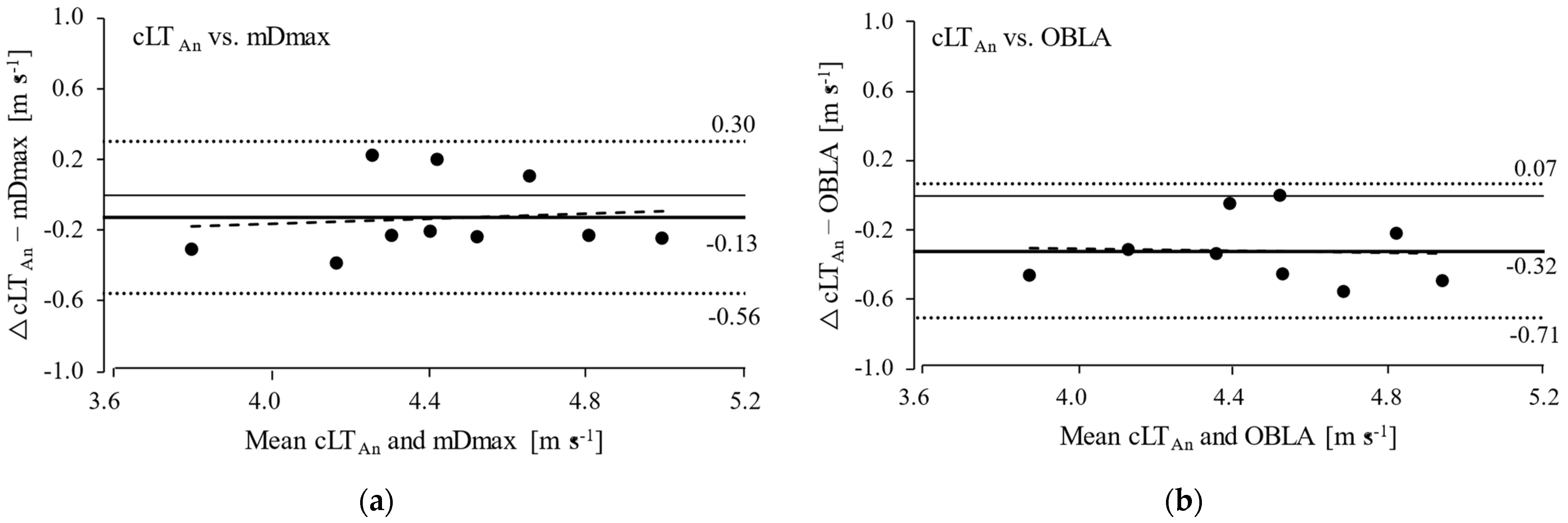
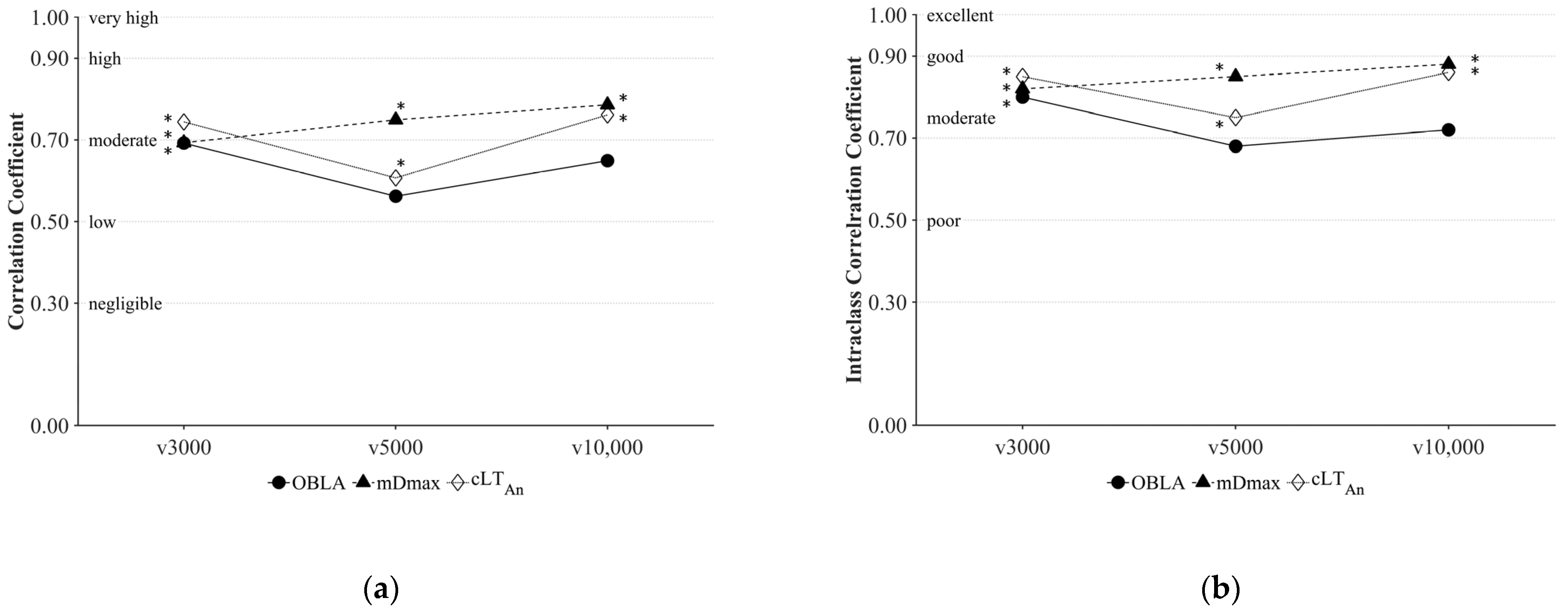
3. Results
4. Discussion
5. Practical Applications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Billat, V.L. Use of Blood Lactate Measurements for Prediction of Exercise Performance and for Control of Training. Recommendations for Long-Distance Running. Sports Med. 1996, 22, 157–175. [Google Scholar] [CrossRef]
- Jacobs, I. Blood Lactate. Sports Med. 1986, 3, 10–25. [Google Scholar] [CrossRef]
- Mader, A.; Heck, H. A Theory of the Metabolic Origin of “Anaerobic Threshold”. Int. J. Sports Med. 1986, 7, 45–65. [Google Scholar] [CrossRef]
- Heck, H.; Mader, A.; Hess, G.; Mücke, S.; Müller, R.; Hollmann, W. Justification of the 4-Mmol/l Lactate Threshold. Int. J. Sports Med. 1985, 6, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Lactate glycolytic end product and oxidative substrate during sustained exercise in mammals—The “lactate shuttle”. In Circulation, Respiration, and Metabolism; Springer: Berlin/Heidelberg, Germany, 1985; pp. 208–218. [Google Scholar]
- Messonnier, L.A.; Emhoff, C.A.W.; Fattor, J.A.; Horning, M.A.; Carlson, T.J.; Brooks, G.A. Lactate Kinetics at the Lactate Threshold in Trained and Untrained Men. J. Appl. Physiol. 2013, 114, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Hering, G.O.; Hennig, E.M.; Riehle, H.J.; Stepan, J. A Lactate Kinetics Method for Assessing the Maximal Lactate Steady State Workload. Front. Physiol. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate Threshold Concepts: How Valid Are They? Sports Med. 2009, 39, 469–490. [Google Scholar] [CrossRef]
- Svedahl, K.; MacIntosh, B.R. Anaerobic Threshold: The Concept and Methods of Measurement. Can. J. Appl. Physiol. 2003, 28, 299–323. [Google Scholar] [CrossRef] [Green Version]
- Kindermann, W.; Simon, G.; Keul, J. The Significance of the Aerobic-Anaerobic Transition for the Determination of Work Load Intensities during Endurance Training. Eur. J. Appl. Physiol. Occup. Physiol. 1979, 42, 25–34. [Google Scholar] [CrossRef]
- Cheng, B.; Kuipers, H.; Snyder, A.C.; Keizer, H.A.; Jeukendrup, A.; Hesselink, M. A New Approach for the Determination of Ventilatory and Lactate Thresholds. Int. J. Sports Med. 1992, 13, 518–522. [Google Scholar] [CrossRef]
- Bishop, D.; Jenkins, D.G.; Mackinnon, L.T. The Relationship between Plasma Lactate Parameters, Wpeak and 1-h Cycling Performance in Women. Med. Sci. Sports Exerc. 1998, 30, 1270–1275. [Google Scholar] [CrossRef] [PubMed]
- Riboli, A.; Rampichini, S.; Cè, E.; Limonta, E.; Coratella, G.; Esposito, F. Effect of Ramp Slope on Different Methods to Determine Lactate Threshold in Semi-Professional Soccer Players. Res. Sports Med. 2019, 27, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Jamnick, N.A.; Botella, J.; Pyne, D.B.; Bishop, D.J. Manipulating Graded Exercise Test Variables Affects the Validity of the Lactate Threshold and VO2peak. PLoS ONE 2018, 13, e0199794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riboli, A.; Rampichini, S.; Cè, E.; Limonta, E.; Borrelli, M.; Coratella, G.; Esposito, F. Training Status Affects between-Protocols Differences in the Assessment of Maximal Aerobic Velocity. Eur. J. Appl. Physiol. 2021, 121, 3083–3093. [Google Scholar] [CrossRef] [PubMed]
- Gladden, L.B. Lactate Metabolism: A New Paradigm for the Third Millennium. J. Physiol. 2004, 558, 5–30. [Google Scholar] [CrossRef]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleicher, A.; Mader, A.; Mester, J. Zur Interpretation von Laktatleistungskurven—Experimentelle Ergebnisse Mit Computergestützten Nachber. Spectr. Sportwiss. 1998, 10, 92–104. [Google Scholar]
- Olbrecht, J. Lactate production and metabolism in swimming. In World Book of Swimming. From Science to Performance; Nova: New York, NY, USA, 2011; pp. 255–276. [Google Scholar]
- Mader, A. Eine Theorie zur Berechnung der Dynamik und des Steady State von Phosphorylierungszustand und Stoffwechselaktivität der Muskelzelle Als Folge des Energiebedarfs; Deutsche Sporthochschule Köln: Köln, Germany, 1984. [Google Scholar]
- Stainsby, W.N.; Brooks, G.A. Control of Lactic Acid Metabolism in Contracting Muscles and during Exercise. Exerc. Sport Sci. Rev. 1990, 18, 29–64. [Google Scholar] [CrossRef]
- Hauser, T.; Adam, J.; Schulz, H. Comparison of Calculated and Experimental Power in Maximal Lactate-Steady State during Cycling. Theor. Biol. Med. Model. 2014, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Adam, J.; Öhmichen, M.; Öhmichen, E.; Rother, J.; Müller, U.M.; Hauser, T.; Schulz, H. Reliability of the Calculated Maximal Lactate Steady State in Amateur Cyclists. Biol. Sport 2015, 32, 97–102. [Google Scholar] [CrossRef]
- Zwingmann, L.; Strütt, S.; Martin, A.; Volmary, P.; Bloch, W.; Wahl, P. Modifications of the Dmax Method in Comparison to the Maximal Lactate Steady State in Young Male Athletes. Physician Sportsmed. 2019, 47, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Faria, E.W.; Parker, D.L.; Faria, I.E. The Science of Cycling Physiology and Training-Part 1. Sports Med. 2005, 35, 285–312. [Google Scholar] [CrossRef] [PubMed]
- Wahl, P.; Zinner, C.; Grosskopf, C.; Rossmann, R.; Bloch, W.; Mester, J. Passive Recovery Is Superior to Active Recovery during a High-Intensity Shock Microcycle. J. Strength Cond. Res. 2013, 27, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Heck, H.; Schulz, H.; Bartmus, U. Diagnostics of Anaerobic Power and Capacity. Eur. J. Sport Sci. 2003, 3, 1–23. [Google Scholar] [CrossRef]
- Hauser, T. Untersuchungen zur Validität und Praktikabilität des Mathematisch Bestimmten Maximalen Laktat-Steady-States Bei Radergometrischen Belastungen; Chemnitz University of Technology: Chemnitz, Germany, 2013. [Google Scholar]
- Saunders, P.U.; Pyne, D.B.; Telford, R.D.; Hawley, J.A. Factors Affecting Running Economy in Trained Distance Runners. Sports Med. 2004, 34, 465–485. [Google Scholar] [CrossRef] [PubMed]
- Swinnen, W.; Kipp, S.; Kram, R. Comparison of Running and Cycling Economy in Runners, Cyclists, and Triathletes. Eur. J. Appl. Physiol. 2018, 118, 1331–1338. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Taylor and Francis: Hoboken, NJ, USA, 1988. [Google Scholar]
- Koo, T.K.; Li, M.Y. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Mukaka, M.M. A Guide to Appropriate Use of Correlation Coefficient in Medical Research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar]
- Barnes, K.R.; Kilding, A.E. Running Economy: Measurement, Norms, and Determining Factors. Sports Med.-Open 2015, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.; Baldini, F.; Martin, P.; Kohrt, W. Ten Kilometer Performance and Predicted Velocity at VO2max among Well-Trained Male Runners. Med. Sci. Sports Exerc. 1989, 21, 78–83. [Google Scholar] [CrossRef]
- Hommel, J.; Öhmichen, S.; Rudolph, U.M.; Hauser, T.; Schulz, H. Effects of Six-Week Sprint Interval or Endurance Training on Calculated Power in Maximal Lactate Steady State. Biol. Sport 2019, 36, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Buchfuhrer, M.J.; Hansen, J.E.; Robinson, T.E.; Sue, D.Y.; Wasserman, K.; Whipp, B.J. Optimizing the Exercise Protocol for Cardiopulmonary Assessment. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 1558–1564. [Google Scholar] [CrossRef]
- González-Alonso, J.; Calbet, J.A.L. Reductions in Systemic and Skeletal Muscle Blood Flow and Oxygen Delivery Limit Maximal Aerobic Capacity in Humans. Circulation 2003, 107, 824–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentley, D.J.; Newell, J.; Bishop, D. Incremental Exercise Test Design and Analysis: Implications for Performance Diagnostics in Endurance Athletes. Sports Med. 2007, 37, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Sperlich, P.F.; Holmberg, H.-C.; Reed, J.L.; Zinner, C.; Mester, J.; Sperlich, B. Individual versus Standardized Running Protocols in the Determination of VO2max. J. Sports Sci. Med. 2015, 14, 386. [Google Scholar] [PubMed]
- Poole, D.C.; Jones, A.M. Measurement of the Maximum Oxygen Uptake VO2max: VO2peak Is No Longer Acceptable. J. Appl. Physiol. 2017, 122, 997–1002. [Google Scholar] [CrossRef]
- Schaun, G.Z. The Maximal Oxygen Uptake Verification Phase: A Light at the End of the Tunnel? Sports Med.-Open 2017, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Krüger, M.; de Marees, M.; Dittmar, K.-H.; Sperlich, B.; Mester, J. Whole-Body Cryotherapy’s Enhancement of Acute Recovery of Running Performance in Well-Trained Athletes. Int. J. Sports Physiol. Perform. 2015, 10, 605–612. [Google Scholar] [CrossRef]
- Nitzsche, N.; Baumgärtel, L.; Schulz, H. Comparison of Maximum Lactate Formation Rates in Ergometer Sprint and Maximum Strength Loads. Dtsch. Z. Sportmed. 2018, 69, 13–17. [Google Scholar] [CrossRef]
- Quittmann, O.J.; Appelhans, D.; Abel, T.; Strüder, H.K. Evaluation of a Sport-Specific Field Test to Determine Maximal Lactate Accumulation Rate and Sprint Performance Parameters in Running. J. Sci. Med. Sport 2020, 23, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Helgerud, J.; Støren, Ø.; Hoff, J. Are There Differences in Running Economy at Different Velocities for Well-Trained Distance Runners? Eur. J. Appl. Physiol. 2010, 108, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Helgerud, J. Maximal Oxygen Uptake, Anaerobic Threshold and Running Economy in Women and Men with Similar Performances Level in Marathons. Eur. J. Appl. Physiol. Occup. Physiol. 1994, 68, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Daniels, J.; Daniels, N. Running Economy of Elite Male and Elite Female Runners. Med. Sci. Sports Exerc. 1992, 24, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Batliner, M.; Kipp, S.; Grabowski, A.; Kram, R.; Byrnes, W. Does Metabolic Rate Increase Linearly with Running Speed in All Distance Runners? Sports Med. Int. Open 2018, 2, E1–E8. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Body Mass (kg) | VO2max (mL kg−1∙min−1) | VLamax (mmol L−1 s−1) | Ks4 (mL kg−1∙min−1 per 1 m s−1) | OBLA (m s−1) | mDmax (m s−1) | cLTAn (m s−1) |
|---|---|---|---|---|---|---|---|
| 1 | 59.2 | 74.6 | 0.38 | 12.1 | 5.19 | 4.93 | 4.70 |
| 2 | 64.4 | 70.0 | 0.32 | 12.1 | 4.97 | 4.65 | 4.42 |
| 3 | 64.4 | 80.3 | 0.33 | 13.2 | - | 5.12 | 4.87 |
| 4 | 72.6 | 68.0 | 0.33 | 11.8 | 4.43 | 4.16 | 4.38 |
| 5 | 72.3 | 65.4 | 0.42 | 11.3 | 4.54 | 4.43 | 4.20 |
| 6 | 68.5 | 62.3 | 0.46 | 10.7 | 4.31 | 4.37 | 3.99 |
| 7 | 59.1 | 80.1 | 0.31 | 14.2 | 4.53 | 4.33 | 4.53 |
| 8 | 73.2 | 67.4 | 0.55 | 10.8 | 4.77 | 4.52 | 4.31 |
| 9 | 58.4 | 68.7 | 0.33 | 11.0 | 4.94 | 4.61 | 4.72 |
| 10 | 65.4 | 60.8 | 0.50 | 11.0 | 4.13 | 3.97 | 3.66 |
| Mean ± SD | 65.8 ± 5.8 | 69.8 ± 6.7 | 0.39 ± 0.09 | 11.8 ± 1.1 | 4.65 ± 0.35 | 4.44 ± 0.28 * | 4.32 ± 0.34 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, S.; Sommer, A.; Bloch, W.; Wahl, P. Comparison and Performance Validation of Calculated and Established Anaerobic Lactate Thresholds in Running. Medicina 2021, 57, 1117. https://doi.org/10.3390/medicina57101117
Ji S, Sommer A, Bloch W, Wahl P. Comparison and Performance Validation of Calculated and Established Anaerobic Lactate Thresholds in Running. Medicina. 2021; 57(10):1117. https://doi.org/10.3390/medicina57101117
Chicago/Turabian StyleJi, Sanghyeon, Aldo Sommer, Wilhelm Bloch, and Patrick Wahl. 2021. "Comparison and Performance Validation of Calculated and Established Anaerobic Lactate Thresholds in Running" Medicina 57, no. 10: 1117. https://doi.org/10.3390/medicina57101117
APA StyleJi, S., Sommer, A., Bloch, W., & Wahl, P. (2021). Comparison and Performance Validation of Calculated and Established Anaerobic Lactate Thresholds in Running. Medicina, 57(10), 1117. https://doi.org/10.3390/medicina57101117