
Dietary Agaricus blazei Spent Substrate Improves Disease Resistance of Nile Tilapia (Oreochromis niloticus) against Streptococcus agalactiae In Vivo


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Preparation of ABSS and Crude Extract Preparation
2.3. In Vitro Trials
2.3.1. Cell Viability Assay
2.3.2. Analysis of Phagocytic Activity
2.3.3. Gene Expression Analysis
2.3.4. Bacterial Inhibition Assay
2.4. In Vivo Experiments
2.4.1. First Feeding Trial
2.4.2. Second Feeding Trial
2.5. Statistical Analysis
3. Results
3.1. Proximate Composition of ABSS
3.2. Approximate Composition of ABSSE
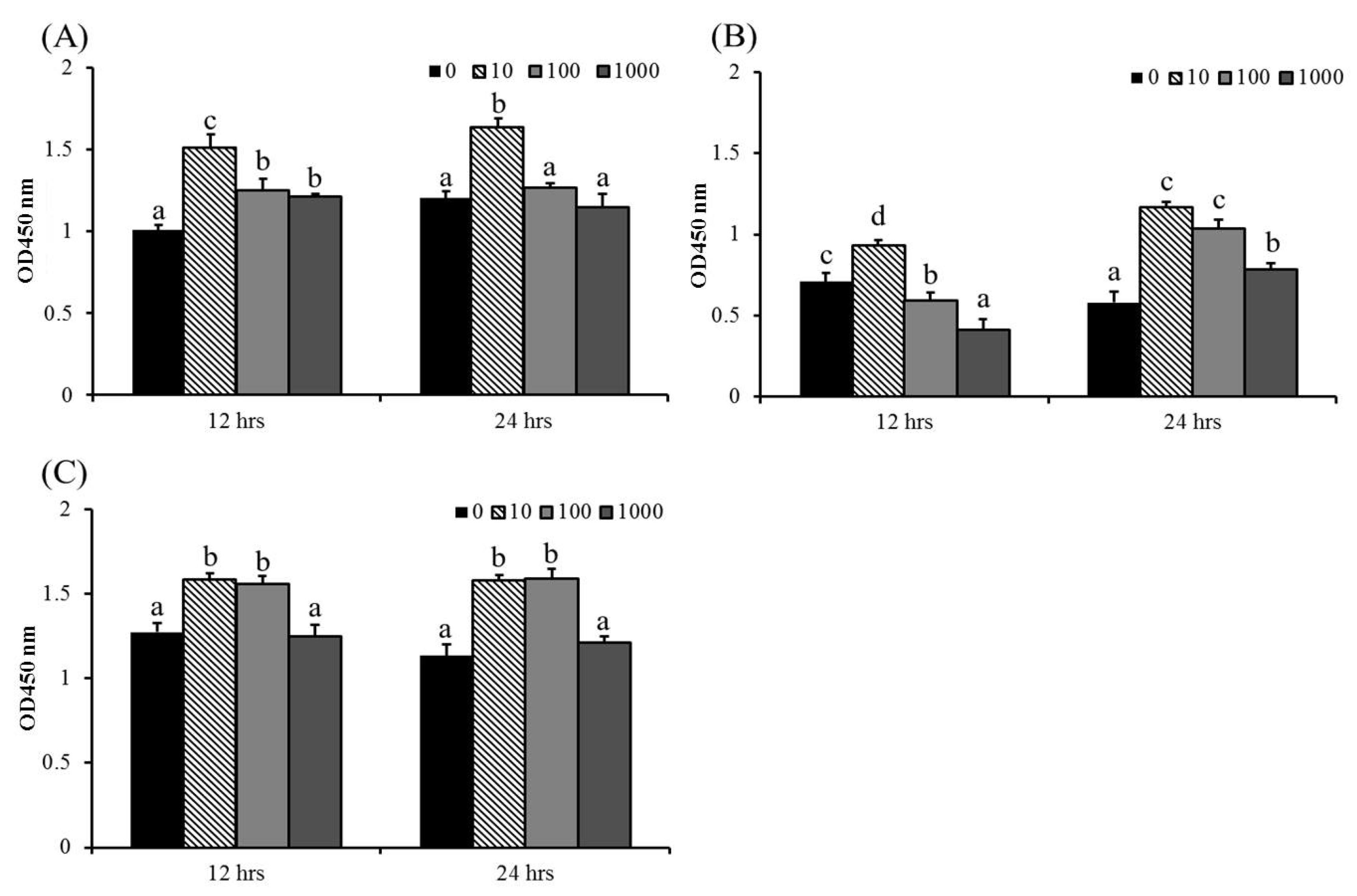
3.3. ABSSE Promotes Proliferation of Hepatic and Splenic Macrophages and the THK Cell Line
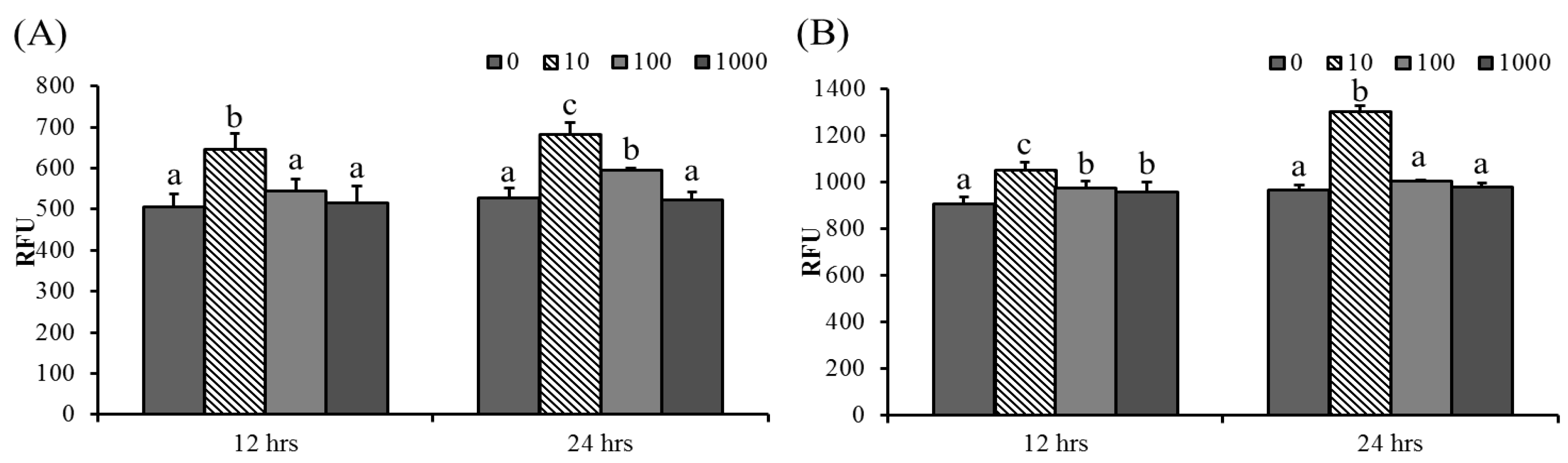
3.4. ABSSE Induce Phagocytic Activity in Hepatic and Splenic Macrophages and THK Cells
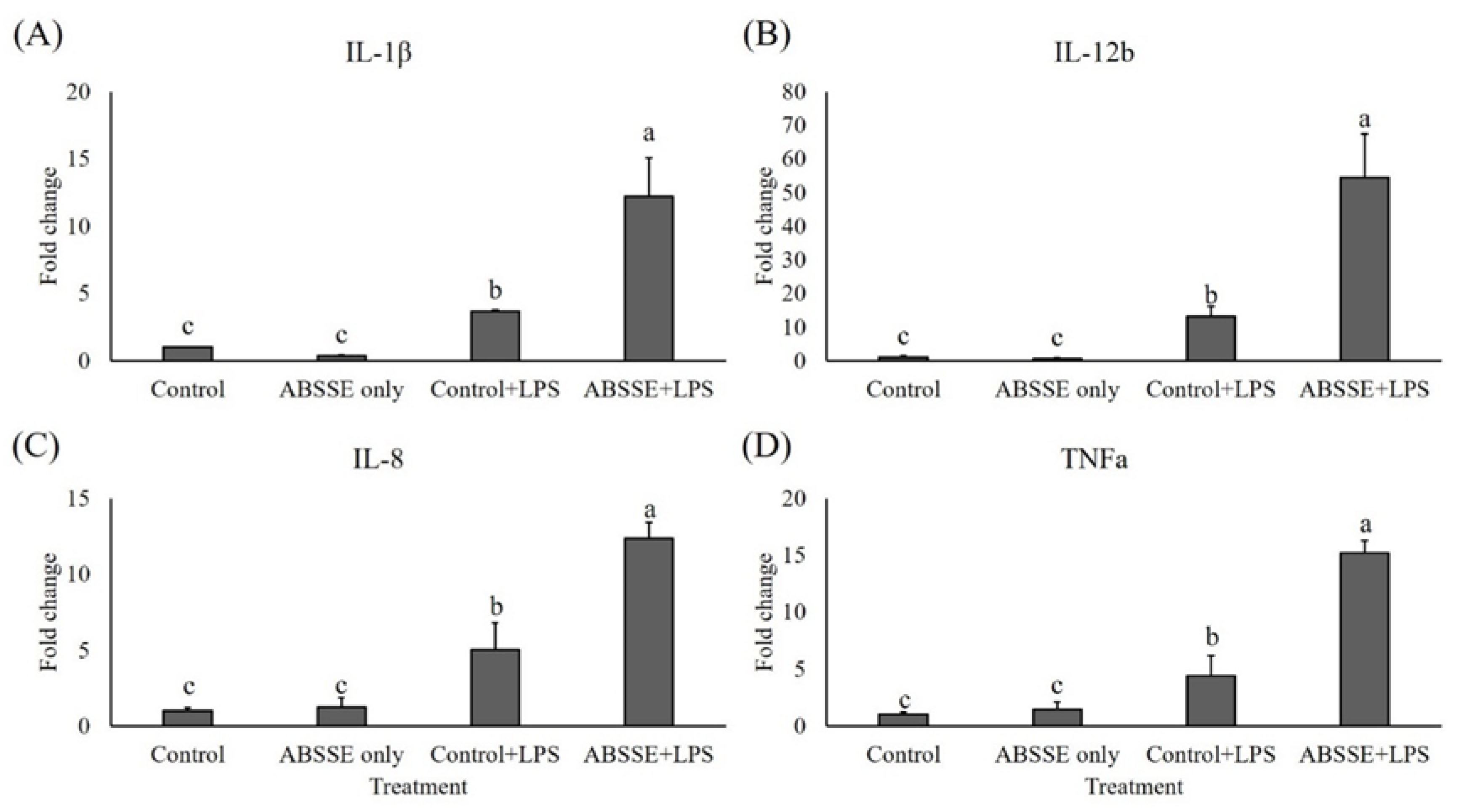
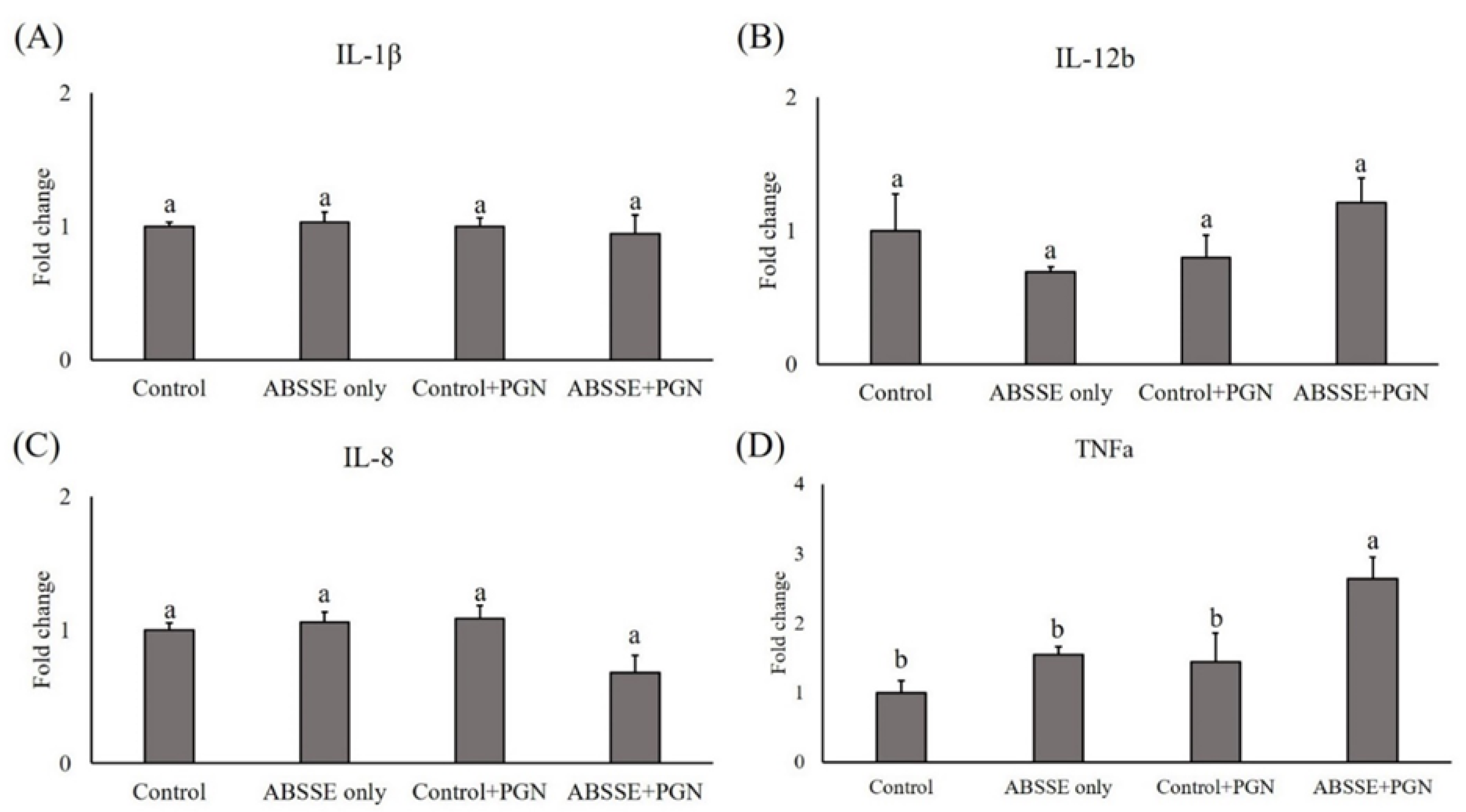
3.5. Priming THK Cells with ABSSE Enhances the Expression of Proinflammatory Cytokines When Bacterial Ligands Are Encountered
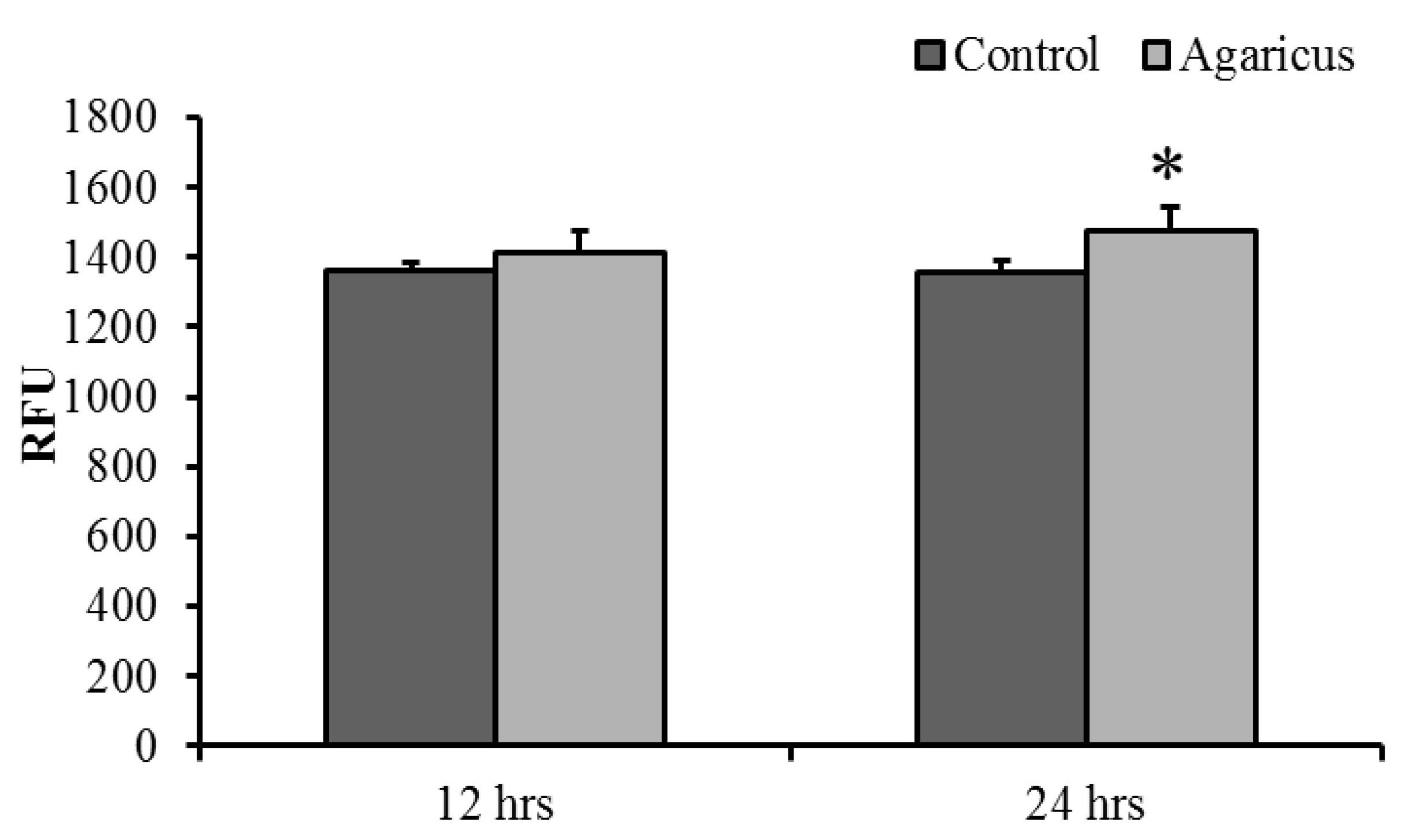
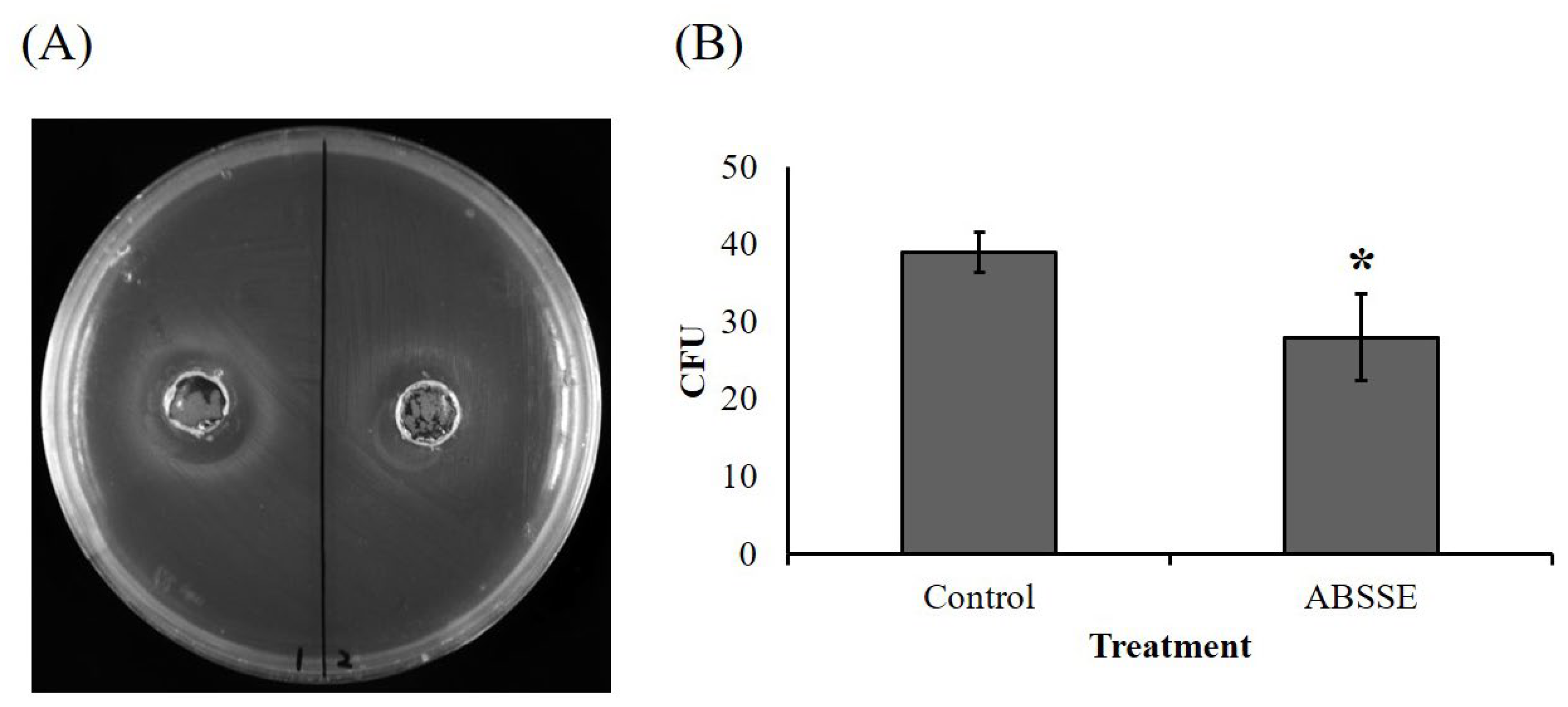
3.6. ABSSE Inhibits the Growth of S. agalactiae
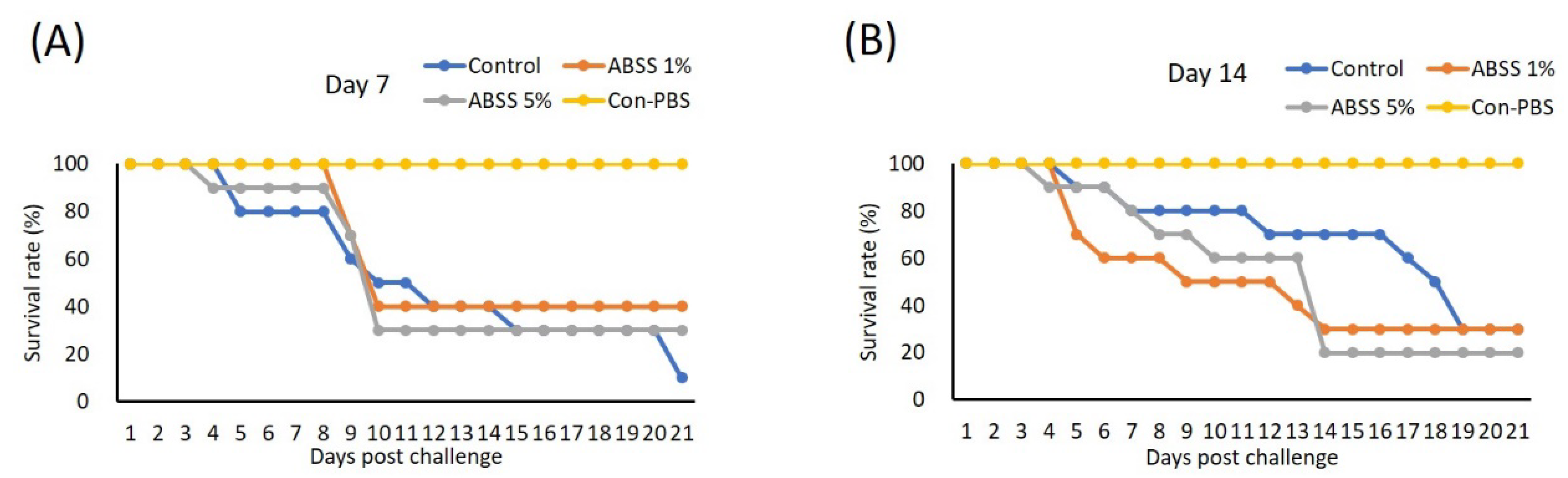
3.7. Protection of Nile Tilapia against Bacterial Infection
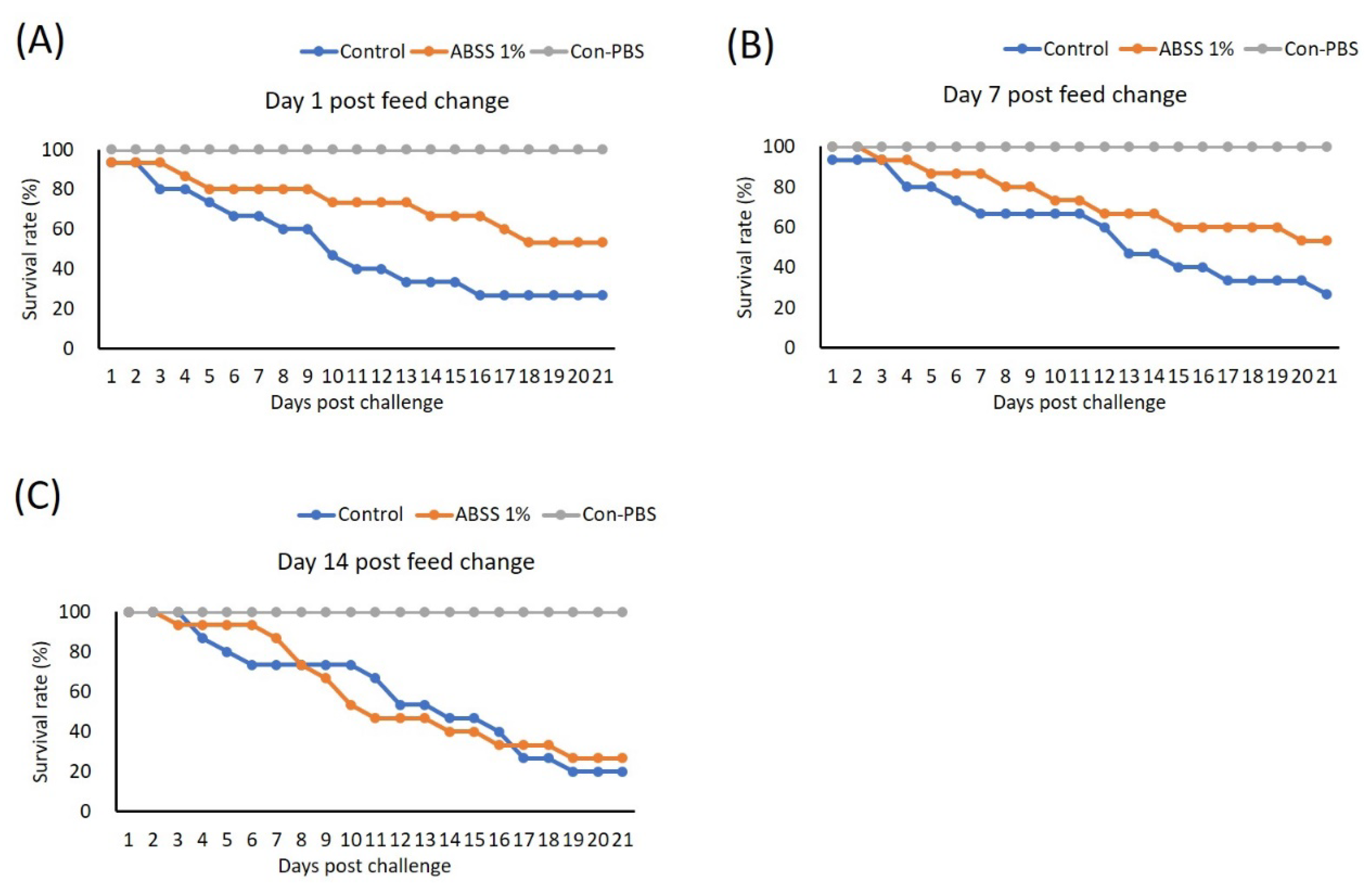
3.8. ABSS Provides Sustained Protection against S. agalactiae Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Report on the State of Global Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020. [Google Scholar]
- El-Sayed, A.-F.M. Current state and future potential. In Tilapia Culture, 2nd ed.; El-Sayed, A.-F.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; Chapter 1; pp. 1–20. [Google Scholar]
- Abu-Elala, N.; Ali, T.; Ragaa, N.; Ali, S.; Abd-Elsalam, R.; Younis, N.; Abdel-Moneam, D.; Hamdien, A.; Bonato, M.; Dawood, M. Analysis of the Productivity, Immunity, and Health Performance of Nile Tilapia (Oreochromis niloticus) Broodstock-fed Dietary Fermented Extracts Sourced from Saccharomyces cerevisiae (Hilyses): A Field Trial. Animals 2021, 11, 815. [Google Scholar] [CrossRef]
- Su, Y.-L.; Feng, J.; Li, Y.-W.; Bai, J.-S.; Li, A.-X. Development of a quantitative PCR assay for monitoring Streptococcus agalactiae colonization and tissue tropism in experimentally infected tilapia. J. Fish Dis. 2016, 39, 229–238. [Google Scholar] [CrossRef]
- Neamat-Allah, A.N.; Mahmoud, E.; El Hakim, Y.A. Efficacy of dietary Nano-selenium on growth, immune response, antioxidant, transcriptomic profile and resistance of Nile tilapia, Oreochromis niloticus against Streptococcus iniae infection. Fish Shellfish Immunol. 2019, 94, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Aly, S.M.; Albutti, A.S.; Rahmani, A.H.; Atti, N.M.A. The response of New-season Nile tilapia to Aeromonas hydrophila vaccine. Int. J. Clin. Exp. Med. 2015, 8, 4508–4514. [Google Scholar]
- Iregui, C.A.; Guarín, M.; Tibatá, V.M.; Ferguson, H.W. Novel brain lesions caused by Edwardsiella tarda in a red tilapia (Oreochromis spp.). J. Veter. Diagn. Investig. 2012, 24, 446–449. [Google Scholar] [CrossRef]
- Hsieh, C.; Tung, M.; Tu, C.; Chang, C.; Tsai, S. Enzootics of visceral granulomas associated with Francisella-like organism infection in tilapia (Oreochromis spp.). Aquaculture 2006, 254, 129–138. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Protective effect of clove oil-supplemented fish diets on experimental Lactococcus garvieae infection in tilapia. Biosci. Biotechnol. Biochem. 2009, 73, 2085–2089. [Google Scholar] [CrossRef] [PubMed]
- Acharya, V.; Chakraborty, H.J.; Rout, A.K.; Balabantaray, S.; Behera, B.K.; Das, B.K. Structural Characterization of Open Reading Frame-Encoded Functional Genes from Tilapia Lake Virus (TiLV). Mol. Biotechnol. 2019, 61, 945–957. [Google Scholar] [CrossRef]
- Pridgeon, J.W.; Klesius, P.H. Major bacterial diseases in aquaculture and their vaccine development. In Animal Science Review; CABI: Boston, MA, USA, 2012; pp. 141–156. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, J.; Chen, K.; Gao, T.; Yao, H.; Liu, Y.; Zhang, W.; Lu, C. Development of Streptococcus agalactiae vaccines for tilapia. Dis. Aquat. Org. 2016, 122, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Bromage, E.; Owens, L. Infection of barramundi Lates calcarifer with Streptococcus iniae: Effects of different routes of exposure. Dis. Aquat. Org. 2002, 52, 199–205. [Google Scholar] [CrossRef]
- Firenzuoli, F.; Gori, L.; Lombardo, G. The Medicinal Mushroom Agaricus blazei Murrill: Review of Literature and Pharmaco-Toxicological Problems. Evid. Based Complement. Altern. Med. 2008, 5, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Hetland, G.; Tangen, J.-M.; Mahmood, F.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Nentwich, I.; Therkelsen, S.P.; Tjønnfjord, G.E.; Johnson, E. Antitumor, Anti-inflammatory and Antiallergic Effects of Agaricus blazei Mushroom Extract and the Related Medicinal Basidiomycetes Mushrooms, Hericium erinaceus and Grifola frondosa: A Review of Preclinical and Clinical Studies. Nutrients 2020, 12, 1339. [Google Scholar] [CrossRef]
- Sorimachi, K.; Akimoto, K.; Ikehara, Y.; Inafuku, K.; Okubo, A.; Yamazaki, S. Secretion of TNF-α, IL-8 and Nitric Oxide by Macrophages Activated with Agaricus blazei Murill Fractions In Vitro. Cell Struct. Funct. 2001, 26, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kawashita, A.; Ishi, S.; Mazumder, T.K.; Nagai, S.; Tsuji, K.; Dan, T. Antihypertensive Effect of GAMMA-Aminobutyric Acid-Enriched Agaricus blazei on Mild Hypertensive Human Subjects. Nippon. Shokuhin Kagaku Kogaku Kaishi 2003, 50, 167–173. [Google Scholar] [CrossRef][Green Version]
- Kweon, M.H.; Kwon, S.T.; Kwon, S.H.; Ma, M.S.; Park, Y.I. Lowering effects in plasma cholesterol and body weight by mycelial extracts of two mushrooms: Agaricus blazai and Lentinus edodes. Korean J. Microbiol. Biotechnol. 2002, 30, 402–409. [Google Scholar]
- Inuzuka, H.; Yoshida, T. Clinical utility of ABCL (Agalicus Mushroom Extract) treatment for C-type hepatitis. Jpn. Pharmacol. Ther. 2002, 30, 103–107. [Google Scholar]
- Takaku, T.; Kimura, Y.; Okuda, H. Isolation of an Antitumor Compound from Agaricus blazei Murill and Its Mechanism of Action. J. Nutr. 2001, 131, 1409–1413. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Martins, A.; Barros, L.; Ferreira, I. Antioxidant properties and phenolic profile of the most widely appreciated cultivated mushrooms: A comparative study between in vivo and in vitro samples. Food Chem. Toxicol. 2012, 50, 1201–1207. [Google Scholar] [CrossRef]
- Ahmed, M.; Abdullah, N.; Shuib, A.S.; Razak, S.A. Influence of raw polysaccharide extract from mushroom stalk waste on growth and pH perturbation induced-stress in Nile tilapia, Oreochromis niloticus. Aquaculture 2017, 468, 60–70. [Google Scholar] [CrossRef]
- Phan, C.-W.; Sabaratnam, V. Potential uses of spent mushroom substrate and its associated lignocellulosic enzymes. Appl. Microbiol. Biotechnol. 2012, 96, 863–873. [Google Scholar] [CrossRef]
- Mayolo-Deloisa, K.; Trejo-Hernández, M.D.R.; Rito-Palomares, M. Recovery of laccase from the residual compost of Agaricus bisporus in aqueous two-phase systems. Process Biochem. 2009, 44, 435–439. [Google Scholar] [CrossRef]
- Munusamy, U.; Sabaratnam, V.; Muniandy, S.; Abdullah, N.; Pandey, A.; Jones, E. Biodegradation of Polycyclic Aromatic Hydrocarbons by Laccase of Pycnoporus sanguineus and Toxicity Evaluation of Treated PAH. Biotechnology 2008, 7, 669–677. [Google Scholar] [CrossRef]
- Trejo-Hernandez, M.; Lopez-Munguia, A.; Ramirez, R.Q. Residual compost of Agaricus bisporus as a source of crude laccase for enzymic oxidation of phenolic compounds. Process Biochem. 2001, 36, 635–639. [Google Scholar] [CrossRef]
- Chiu, S.; Ching, M.; Fong, K.; Moore, D. Spent oyster mushroom substrate performs better than many mushroom mycelia in removing the biocide pentachlorophenol. Mycol. Res. 1998, 102, 1553–1562. [Google Scholar] [CrossRef]
- Adamović, M.; Grubić, G.; Milenković, I.; Jovanović, R.; Protić, R.; Sretenović, L.; Stoićević, L. The biodegradation of wheat straw by Pleurotus ostreatus mushrooms and its use in cattle feeding. Anim. Feed. Sci. Technol. 1998, 71, 357–362. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Reyes-López, F.; Teles, M.; MacKenzie, S. The response of fish to immunostimulant diets. Fish Shellfish. Immunol. 2016, 56, 34–69. [Google Scholar] [CrossRef] [PubMed]
- Yudiati, E.; Isnansetyo, A.; Handayani, C.R. Alginate from Sargassum siliquosum Simultaneously Stimulates Innate Immunity, Upregulates Immune Genes, and Enhances Resistance of Pacific White Shrimp (Litopenaeus vannamei) against White Spot Syndrome Virus (WSSV). Mar. Biotechnol. 2019, 21, 503–514. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Dawood, M.; Chitmanat, C.; Tayyamath, K. Effects of Cordyceps militaris spent mushroom substrate and Lactobacillus plantarum on mucosal, serum immunology and growth performance of Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Pohlenz, C.; Gatlin, D.M. Interrelationships between fish nutrition and health. Aquaculture 2014, 431, 111–117. [Google Scholar] [CrossRef]
- Van Doan, H.; Hoseinifar, S.H.; Tapingkae, W.; Chitmanat, C.; Mekchay, S. Effects of Cordyceps militaris spent mushroom substrate on mucosal and serum immune parameters, disease resistance and growth performance of Nile tilapia, (Oreochromis niloticus). Fish Shellfish. Immunol. 2017, 67, 78–85. [Google Scholar] [CrossRef]
- Horm, V.; Ohga, S. Potential of Compost with Some Added Supplementary Materials on the Development of Agaricus blazei Murill. J. Fac. Agric. Kyushu Univ. 2008, 53, 417–422. [Google Scholar] [CrossRef]
- Largeteau, M.L.; Llarena-Hernandez, R.C.; Regnault-Roger, C.; Savoie, J.-M. The medicinal Agaricus mushroom cultivated in Brazil: Biology, cultivation and non-medicinal valorisation. Appl. Microbiol. Biotechnol. 2011, 92, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Rózsa, S.; Măniuțiu, D.-N.; Poșta, G.; Gocan, T.-M.; Andreica, I.; Bogdan, I.; Lazăr, V. Influence of the Culture Substrate on the Agaricus blazei Murrill Mushrooms Vitamins Content. Plants 2019, 8, 316. [Google Scholar] [CrossRef]
- Cunniff, P.; AOAC International. Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 1997. [Google Scholar]
- Bhadja, P.; Tan, C.-Y.; Ouyang, J.-M.; Yu, K. Repair Effect of Seaweed Polysaccharides with Different Contents of Sulfate Group and Molecular Weights on Damaged HK-2 Cells. Polymers 2016, 8, 188. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Nan, F.-H.; Huang, S.-L.; Hsiao, C.-M.; Lai, K.-C.; Lu, C.-L.; Chen, S.-N. Studies of macrophage cellular response to the extracellular hydrogen peroxide by tilapia model. Fish Shellfish Immunol. 2014, 36, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.-M. Development and characterization of a cell line from tilapia head kidney with melanomacrophage characteristics. Fish Shellfish. Immunol. 2016, 49, 442–449. [Google Scholar] [CrossRef]
- Lee, P.-T.; Wen, C.-M.; Nan, F.-H.; Yeh, H.-Y.; Lee, M.-C. Immunostimulatory effects of Sarcodia suiae water extracts on Nile tilapia Oreochromis niloticus and its resistance against Streptococcus agalactiae. Fish Shellfish. Immunol. 2020, 103, 159–168. [Google Scholar] [CrossRef]
- Trung, N.B.; Lee, P.-T. Functional characterization of myeloid differentiation factor 88 in Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 250, 110485. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Diaz-Rosales, P.; Costa, M.M.; Campbell, S.; Snow, M.; Collet, B.; Martin, S.; Secombes, C.J. Functional Characterization of a Nonmammalian IL-21: Rainbow TroutOncorhynchus mykissIL-21 Upregulates the Expression of the Th Cell Signature Cytokines IFN-γ, IL-10, and IL-22. J. Immunol. 2010, 186, 708–721. [Google Scholar] [CrossRef]
- Hao, H.; Fu, M.; Yan, R.; He, B.; Li, M.; Liu, Q.; Cai, Y.; Zhang, X.; Huang, R. Chemical composition and immunostimulatory properties of green alga Caulerpa racemosa var peltata. Food Agric. Immunol. 2019, 30, 937–954. [Google Scholar] [CrossRef]
- Bernart, M.W. Mushrooms. Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; Chang, S.-T., Miles, P.G., Eds.; CRC Press: Boca Raton, FL, USA, 2004; 451p. [Google Scholar] [CrossRef]
- Cardozo, F.T.G.D.S.; Camelini, C.M.; Mascarello, A.; Rossi, M.J.; Nunes, R.J.; Barardi, C.R.M.; de Mendonça, M.M.; Simões, C.M.O. Antiherpetic activity of a sulfated polysaccharide from Agaricus brasiliensis mycelia. Antivir. Res. 2011, 92, 108–114. [Google Scholar] [CrossRef]
- Ker, Y.-B.; Chen, K.-C.; Chyau, C.-C.; Chen, C.-C.; Guo, J.-H.; Hsieh, C.-L.; Wang, H.-E.; Peng, C.-C.; Chang, C.-H.; Peng, R.Y. Antioxidant Capability of Polysaccharides Fractionated from Submerge-Cultured Agaricus blazei Mycelia. J. Agric. Food Chem. 2005, 53, 7052–7058. [Google Scholar] [CrossRef]
- Cheng, F.; Yan, X.; Zhang, M.; Chang, M.; Yun, S.; Meng, J.; Liu, J.; Feng, C.-P. Regulation of RAW 264.7 cell-mediated immunity by polysaccharides from Agaricus blazei Murill via the MAPK signal transduction pathway. Food Funct. 2017, 8, 1475–1480. [Google Scholar] [CrossRef] [PubMed]
- Navegantes, K.C.; Albuquerque, R.F.V.; Dalla-Santa, H.S.; Soccol, C.R.; Monteiro, M.C. Agaricus brasiliensismycelium and its polysaccharide modulate the parameters of innate and adaptive immunity. Food Agric. Immunol. 2013, 24, 393–408. [Google Scholar] [CrossRef]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Miyagawa, M.; Hirono, Y.; Kawazoe, A.; Shigeyoshi, E.; Nose, M.; Sakura, M.; Pinkerton, K.E.; Takeuchi, M. Effect of Hot Water Extract from Agaricus blazei Murill on Chemotaxis of Neutrophils. J. Cosmet. Dermatol. Sci. Appl. 2013, 3, 12–17. [Google Scholar] [CrossRef]
- Yasuma, T.; Toda, M.; Kobori, H.; Tada, N.; D’Alessandro-Gabazza, C.; Gabazza, E. Subcritical Water Extracts from Agaricus blazei Murrill’s Mycelium Inhibit the Expression of Immune Checkpoint Molecules and Axl Receptor. J. Fungi 2021, 7, 590. [Google Scholar] [CrossRef]
- Cleary, J.A.; Kelly, G.E.; Husband, A.J. The effect of molecular weight and β-1,6-linkages on priming of macrophage function in mice by (1,3)-β-d-glucan. Immunol. Cell Biol. 1999, 77, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Thornton, B.P.; Ross, G.D. Soluble beta-glucan polysaccharide binding to the lectin site of neutrophil or natural killer cell complement receptor type 3 (CD11b/CD18) generates a primed state of the receptor capable of mediating cytotoxicity of iC3b-opsonized target cells. J. Clin. Investig. 1996, 98, 50–61. [Google Scholar] [CrossRef]
- Huang, T.-T.; Ojcius, D.; Young, J.D.; Wu, Y.-H.; Ko, Y.-F.; Wong, T.-Y.; Wu, C.-Y.; Lu, C.-C.; Lai, H.-C. The Anti-Tumorigenic Mushroom Agaricus blazei Murill Enhances IL-1β Production and Activates the NLRP3 Inflammasome in Human Macrophages. PLoS ONE 2012, 7, e41383. [Google Scholar] [CrossRef]
- Bernardshaw, S.; Hetland, G.; Ellertsen, L.K.; Tryggestad, A.M.A.; Johnson, E. An Extract of the Medicinal Mushroom Agaricus blazei Murill Differentially Stimulates Production of Pro-inflammatory Cytokines in Human Monocytes and Human Vein Endothelial Cells In Vitro. Inflammation 2006, 29, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.; Ozinsky, A.; Hajjar, A.; Stevens, A.; Wilson, C.B.; Bassetti, M.; Aderem, A. The Toll-like receptor 2 is recruited to macrophage phagosomes and discriminates between pathogens. Nature 1999, 401, 811–815. [Google Scholar] [CrossRef]
- Brown, G.D.; Herre, J.; Williams, D.L.; Willment, J.; Marshall, A.; Gordon, S. Dectin-1 Mediates the Biological Effects of β-Glucans. J. Exp. Med. 2003, 197, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Bailey, E.C.; Wheeler, R.T.; De Oliveira, C.A.F.; Forlenza, M.; Wiegertjes, G.F. Studies Into β-Glucan Recognition in Fish Suggests a Key Role for the C-Type Lectin Pathway. Front. Immunol. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.J.G.; Pugh, N.; Pasco, D.S.; Ross, S.A. Isolation of a Galactomannan That Enhances Macrophage Activation from the Edible Fungus Morchella esculenta. J. Agric. Food Chem. 2002, 50, 5683–5685. [Google Scholar] [CrossRef]
- Hsu, J.-W.; Huang, H.-C.; Chen, S.-T.; Wong, C.-H.; Juan, H.-F. Ganoderma lucidumPolysaccharides Induce Macrophage-like Differentiation in Human Leukemia THP-1 Cells via Caspase and p53 Activation. Evid. Based Complement. Altern. Med. 2011, 2011, 358717. [Google Scholar] [CrossRef]
- Carvajal, A.E.S.; Koehnlein, E.A.; Soares, A.A.; Eler, G.J.; Nakashima, A.T.A.; Bracht, A.; Peralta, R.M. Bioactives of fruiting bodies and submerged culture mycelia of Agaricus brasiliensis (A. blazei) and their antioxidant properties. LWT Food Sci. Technol. 2012, 46, 493–499. [Google Scholar] [CrossRef]
- Sorimachi, K.; Ikehara, Y.; Maezato, G.; Okubo, A.; Yamazaki, S.; Akimoto, K.; Niwa, A. Inhibition by Agaricus blazei Murill Fractions of Cytopathic Effect Induced by Western Equine Encephalitis (WEE) Virus on VERO Cells In Vitro. Biosci. Biotechnol. Biochem. 2001, 65, 1645–1647. [Google Scholar] [CrossRef][Green Version]
- Alves, M.J.; Ferreira, I.; Dias, J.; Teixeira, V.; Martins, A.; Pintado, M.M. A Review on Antimicrobial Activity of Mushroom (Basidiomycetes) Extracts and Isolated Compounds. Planta Med. 2012, 78, 1707–1718. [Google Scholar] [CrossRef]
- Lima, C.U.; Gris, E.F.; Karnikowski, M. Antimicrobial properties of the mushroom Agaricus blazei—Integrative review. Rev. Bras. Farm. 2016, 26, 780–786. [Google Scholar] [CrossRef][Green Version]
- Bernardshaw, S.; Hetland, G.; Grinde, B.; Johnson, E. An extract of the mushroom Agaricus blazei Murill protects against lethal septicemia in a mouse model of fecal peritonitis. Shock 2006, 25, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Osaki, Y.; Kato, T.; Yamamoto, K.; Okubo, J.; Miyazaki, T. Antimutagenic and Bactericidal Substances in the Fruit Body of a Basidiomycete Agaricus blazei, Jun-17. Yakugaku Zasshi J. Pharm. Soc. Jpn. 1994, 114, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Stojković, D.; Reis, F.S.; Glamočlija, J.; Ćirić, A.; Barros, L.; Van Griensven, L.J.L.D.; Ferreira, I.C.F.R.; Soković, M. Cultivated strains of Agaricus bisporus and A. brasiliensis: Chemical characterization and evaluation of antioxidant and antimicrobial properties for the final healthy product—Natural preservatives in yoghurt. Food Funct. 2014, 5, 1602–1612. [Google Scholar] [CrossRef]
- Soković, M.; Ćirić, A.; Glamočlija, J.; Nikolić, M.; Van Griensven, L.J.L.D. Agaricus blazei Hot Water Extract Shows Anti Quorum Sensing Activity in the Nosocomial Human Pathogen Pseudomonas Aeruginosa. Molecules 2014, 19, 4189–4199. [Google Scholar] [CrossRef]
- Mazzutti, S.; Ferreira, S.R.S.; Riehl, C.A.; Smania, A.; Smania, F.A.; Martínez, J. Supercritical fluid extraction of Agaricus brasiliensis: Antioxidant and antimicrobial activities. J. Supercrit. Fluids 2012, 70, 48–56. [Google Scholar] [CrossRef]
- Bernardshaw, S.; Johnson, E.; Hetland, G. An Extract of the Mushroom Agaricus blazei Murill Administered Orally Protects against Systemic Streptococcus pneumoniae Infection in Mice. Scand. J. Immunol. 2005, 62, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; Wiegertjes, G.F. Long-lived effects of administering β-glucans: Indications for trained immunity in fish. Dev. Comp. Immunol. 2016, 64, 93–102. [Google Scholar] [CrossRef]
- Bagni, M.; Archetti, L.; Amadori, M.; Marino, G. Effect of Long-term Oral Administration of an Immunostimulant Diet on Innate Immunity in Sea Bass (Dicentrarchus labrax). J. Veter-Med. Ser. B 2000, 47, 745–751. [Google Scholar] [CrossRef]
- Chang, C.-S.; Huang, S.-L.; Chen, S.; Chen, S.-N. Innate immune responses and efficacy of using mushroom beta-glucan mixture (MBG) on orange-spotted grouper, Epinephelus coioides, aquaculture. Fish Shellfish Immunol. 2013, 35, 115–125. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′→3′) | Reverse (5′→3′) | Accession Number |
|---|---|---|---|
| β-actin | TCCTTCCTTGGTATGGAATCC | GTGGGGCAATGATCTTGATC | KJ126772 |
| IL-1β | CAGTGAAGACCGCAAAGTGC | TATCCGTCACCTCCTCCAG | XM_019365841 |
| IL-8 | TCGCCACCTGTGAAGGCAT | TCCTTTTCAGTGTGGCAATGAT | XM_019359413 |
| IL-12b | CAACAGTGACAATCAAATAATTAATAT | CGTTATGTTTGTTCACTGTGCA | XM_019364048 |
| TNF-α | GAACACTGGCGACAAAACAGA | TTGAGTCGCTGCCTTCTAGA | AY428948 |
| Moisture | 10.12 |
|---|---|
| Ash | 11.29 |
| Crude protein | 4.34 |
| Crude lipid | 6.25 |
| Fiber | 4.33 |
| NFE * | 63.67 |
| Content | Percentage |
|---|---|
| Total carbohydrate (w/w, %) | >95 |
| Sulfate (%) | 6.3 |
| β-glucan (%) | 1.08 |
| Monosaccharide content (%) | 65.4 (Sucrose) |
| Polysaccharide content (%) | 34.6 |
| Glucose | 62.4 |
| Galactose | 37.6 |
| Others | 0 |
| Proximate Analysis (%) | Control | ABSS * 1% | ABSS * 5% |
|---|---|---|---|
| Crude protein | 28.0 | 27.8 | 26.8 |
| Crude lipid | 9.7 | 9.8 | 9.3 |
| Carbohydrate | 49.6 | 49.8 | 51.0 |
| Moisture | 6.4 | 6.3 | 6.6 |
| Ash | 6.3 | 6.3 | 6.3 |
| Calorie (kcal/100 g) | 398 | 399 | 395 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, P.-T.; Wu, Y.-S.; Tseng, C.-C.; Lu, J.-Y.; Lee, M.-C. Dietary Agaricus blazei Spent Substrate Improves Disease Resistance of Nile Tilapia (Oreochromis niloticus) against Streptococcus agalactiae In Vivo. J. Mar. Sci. Eng. 2022, 10, 100. https://doi.org/10.3390/jmse10010100
Lee P-T, Wu Y-S, Tseng C-C, Lu J-Y, Lee M-C. Dietary Agaricus blazei Spent Substrate Improves Disease Resistance of Nile Tilapia (Oreochromis niloticus) against Streptococcus agalactiae In Vivo. Journal of Marine Science and Engineering. 2022; 10(1):100. https://doi.org/10.3390/jmse10010100
Chicago/Turabian StyleLee, Po-Tsang, Yu-Sheng Wu, Chung-Chih Tseng, Jia-Yu Lu, and Meng-Chou Lee. 2022. "Dietary Agaricus blazei Spent Substrate Improves Disease Resistance of Nile Tilapia (Oreochromis niloticus) against Streptococcus agalactiae In Vivo" Journal of Marine Science and Engineering 10, no. 1: 100. https://doi.org/10.3390/jmse10010100
APA StyleLee, P.-T., Wu, Y.-S., Tseng, C.-C., Lu, J.-Y., & Lee, M.-C. (2022). Dietary Agaricus blazei Spent Substrate Improves Disease Resistance of Nile Tilapia (Oreochromis niloticus) against Streptococcus agalactiae In Vivo. Journal of Marine Science and Engineering, 10(1), 100. https://doi.org/10.3390/jmse10010100