

Exploring the Benefits of Phycocyanin: From Spirulina Cultivation to Its Widespread Applications

 ,
,  ,
,  , and
, and 
Abstract
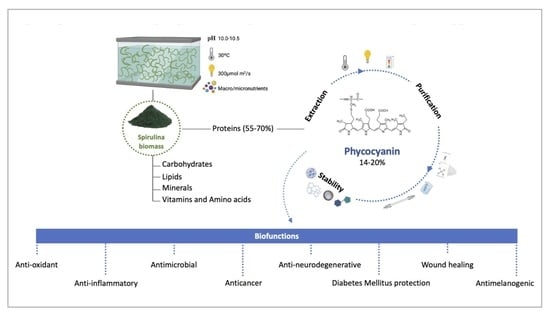
1. Introduction
2. Overview of Spirulina
3. Phycocyanin
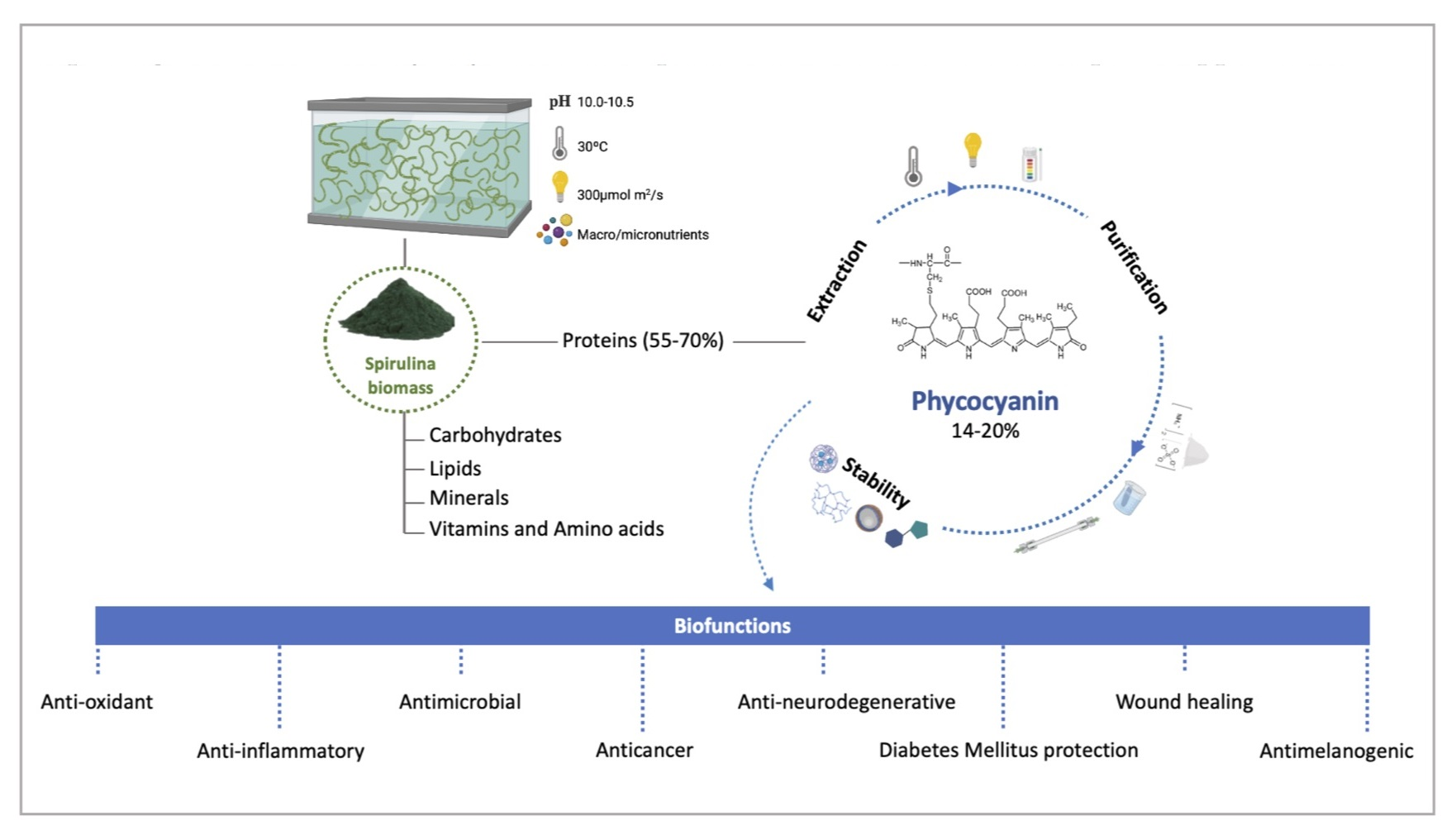
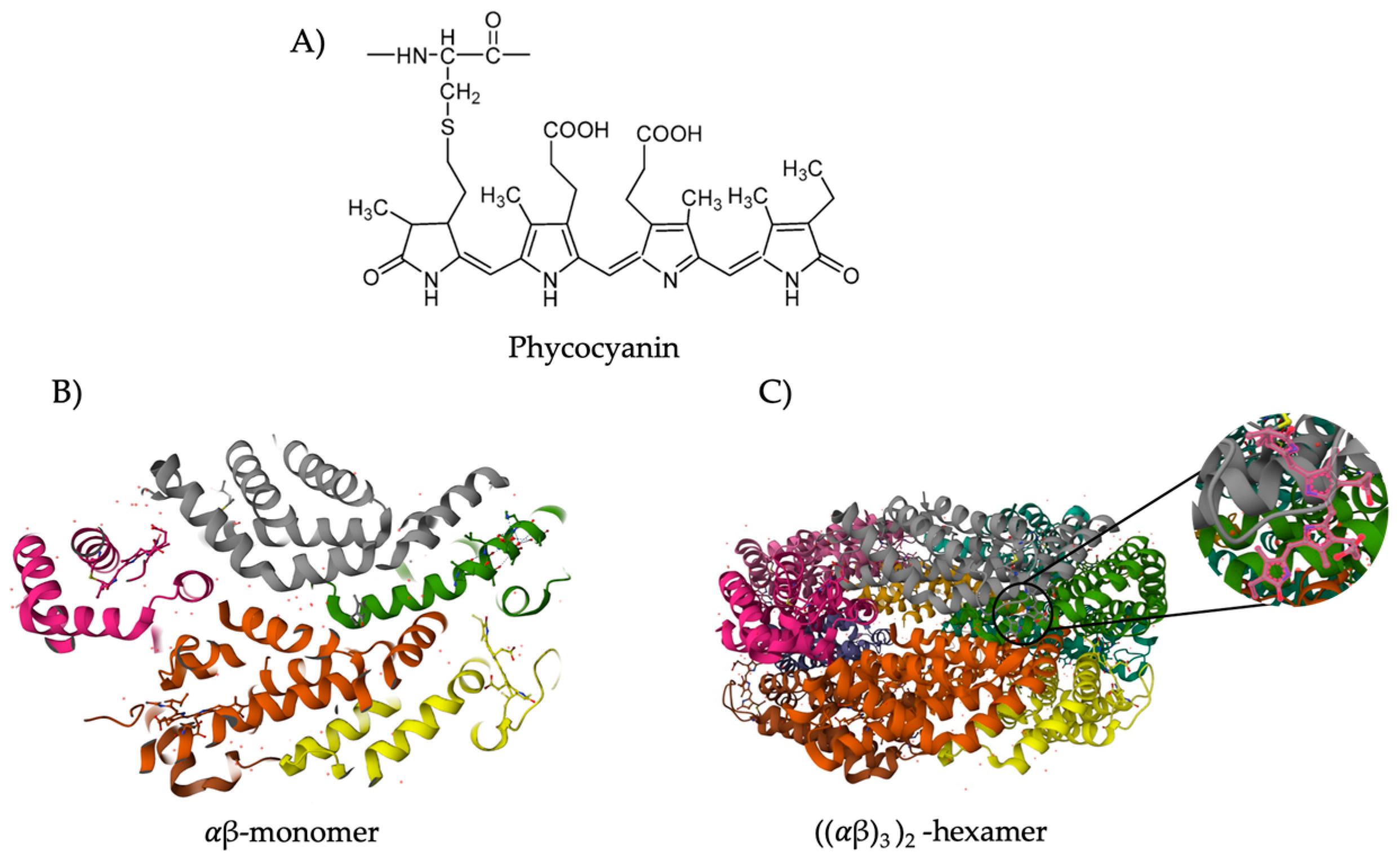
3.1. Classification and Structure
3.2. Biological Functions and Applicability
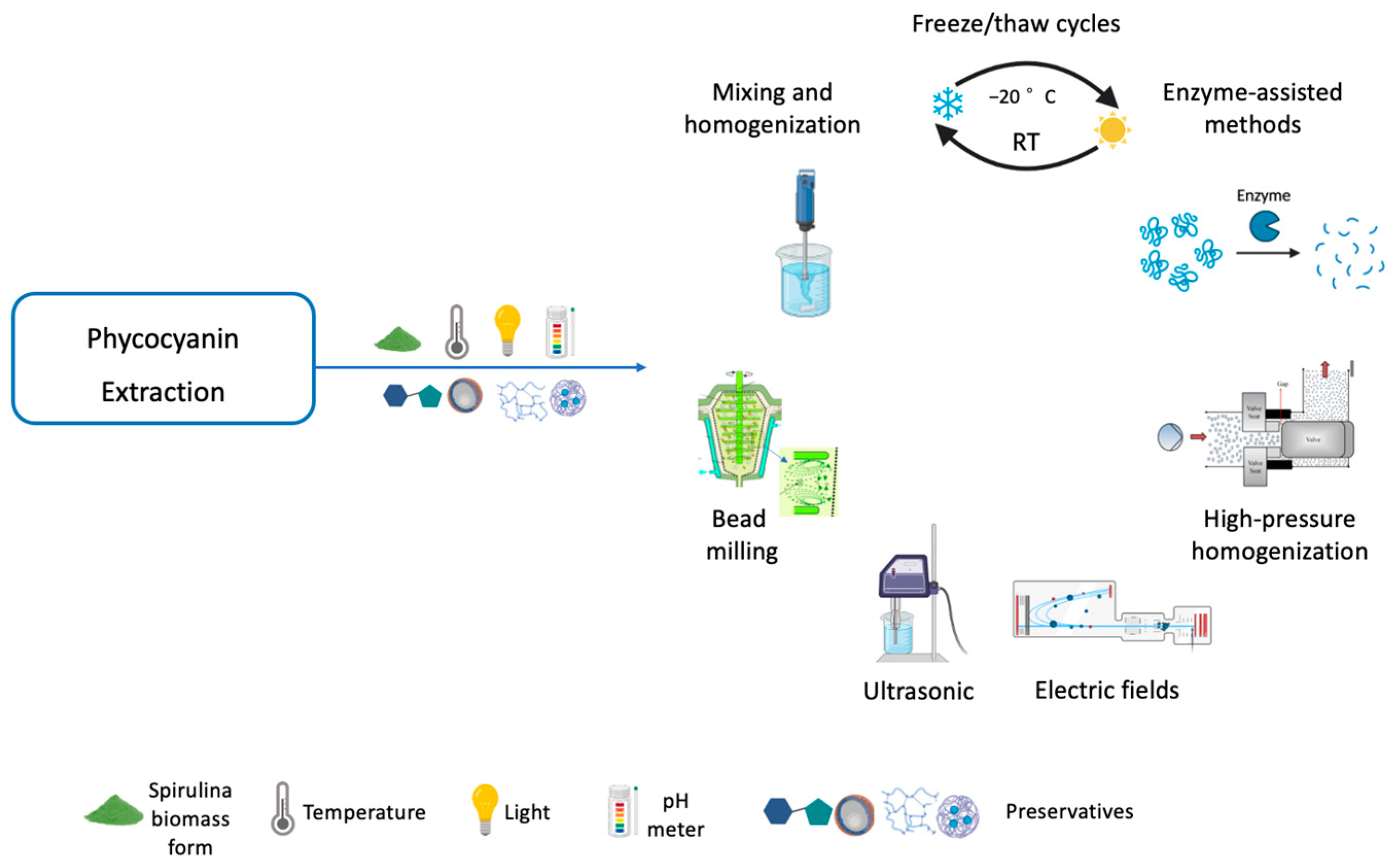
3.3. Phycocyanin Extraction Methods
3.3.1. Cell Disruption Methods
Freeze/Thaw Cycles
Mixing and Homogenization
Bead Milling
Ultrasonic
Electric Fields
High-Pressure Homogenization
Enzyme-Assisted Methods
3.4. Key Parameters for Phycocyanin Extraction and Stability
3.4.1. Biomass Form
3.4.2. Temperature
3.4.3. Light
3.4.4. pH
3.4.5. Type of Solvent
3.4.6. Biomass/Solvent Ratio
3.4.7. Preservatives
3.5. Phycocyanin Purification Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. Available online: https://www.un.org/en/desa/world-population-projected-reach-98-billion-2050-and-112-billion-2100 (accessed on 3 March 2023).
- Lim, H.R.; Khoo, K.S.; Chew, K.W.; Chang, C.-K.; Munawaroh, H.S.H.; Kumar, P.S.; Huy, N.D.; Show, P.L. Perspective of Spirulina culture with wastewater into a sustainable circular bioeconomy. Environ. Pollut. 2021, 284, 117492. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T. Cultured Microalgae and Compounds Derived Thereof for Food Applications: Strain Selection and Cultivation, Drying, and Processing Strategies. Food Rev. Int. 2020, 36, 559–583. [Google Scholar] [CrossRef]
- Jaeschke, D.P.; Teixeira, I.R.; Marczak, L.D.F.; Mercali, G.D. Phycocyanin from Spirulina: A review of extraction methods and stability. Food Res. Int. 2021, 143, 110314. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. New Report Calls for Urgent Action to Avert Antimicrobial Resistance Crisis. International Organizations Unite on Critical Recommendations to Combat Drug-Resistant Infections and Prevent Staggering Number of Deaths Each Year. Available online: https://www.who.int/news/item/29-04-2019-new-report-calls-for-urgent-action-to-avert-antimicrobial-resistance-crisis (accessed on 3 March 2023).
- Nowruzi, B. Cyanobacteria Natural Products as Sources for Future Directions in Antibiotic Drug Discovery. In Cyanobacteria—Recent Advances and New Perspectives; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Saxena, R.; Rodríguez-Jasso, R.M.; Chávez-Gonzalez, M.L.; Aguilar, C.N.; Quijano, G.; Ruiz, H.A. Strategy Development for Microalgae Spirulina platensis Biomass Cultivation in a Bubble Photobioreactor to Promote High Carbohydrate Content. Fermentation 2022, 8, 374. [Google Scholar] [CrossRef]
- AlFadhly, N.K.Z.; Alhelfi, N.; Altemimi, A.B.; Verma, D.K.; Cacciola, F. Tendencies Affecting the Growth and Cultivation of Genus Spirulina: An Investigative Review on Current Trends. Plants 2022, 11, 3063. [Google Scholar] [CrossRef]
- Algae Products Market Size, Growth and Forecast 2028. Available online: https://www.credenceresearch.com/report/algae-products-market (accessed on 3 March 2023).
- Paniagua-Michel, J. Microalgal Nutraceuticals. In Handbook of Marine Microalgae: Biotechnology Advances; Academic Press: Cambridge, MA, USA, 2015; pp. 255–267. [Google Scholar] [CrossRef]
- Sorrenti, V.; Castagna, D.A.; Fortinguerra, S.; Buriani, A.; Scapagnini, G.; Willcox, D.C. Spirulina Microalgae and Brain Health: A Scoping Review of Experimental and Clinical Evidence. Mar. Drugs 2021, 19, 293. [Google Scholar] [CrossRef]
- de Morais, M.G.; da Fontoura Prates, D.; Moreira, J.B.; Duarte, J.H.; Costa, J.A.V. Phycocyanin from Microalgae: Properties, Extraction and Purification, with Some Recent Applications. Ind. Biotechnol. 2018, 14, 30–37. [Google Scholar] [CrossRef]
- Yuan, B.; Li, Z.; Shan, H.; Dashnyam, B.; Xu, X.; McClements, D.J.; Zhang, B.; Tan, M.; Wang, Z.; Cao, C. A review of recent strategies to improve the physical stability of phycocyanin. Curr. Res. Food Sci. 2022, 5, 2329–2337. [Google Scholar] [CrossRef]
- Jung, C.H.G.; Braune, S.; Waldeck, P.; Küpper, J.-H.; Petrick, I.; Jung, F. Morphology and Growth of Arthrospira platensis during Cultivation in a Flat-Type Bioreactor. Life 2021, 11, 536. [Google Scholar] [CrossRef]
- Trotta, T.; Porro, C.; Cianciulli, A.; Panaro, M.A. Beneficial Effects of Spirulina Consumption on Brain Health. Nutrients 2022, 14, 676. [Google Scholar] [CrossRef]
- A Review on Culture, Production and Use of Spirulina as Food for Humans and Feeds for Domestic Animals and Fish. Food and Agriculture Organization of the United Nations: Rome, Italy, 2008. Available online: https://www.fao.org/3/i0424e/i0424e00.pdf (accessed on 3 March 2023).
- Economic and Social Council Substantive Session For 2008 High-Level Segment. This Year’s Theme is “Achieving Sustainable Development”. Available online: https://www.un.org/en/ecosoc/docs/statement08/iimsam.pdf (accessed on 3 March 2023).
- OPINION of the French Agency for Food, Environmental and Occupational Health & Safety on the “Risks Associated with the Consumption of Food Supplements Containing Spirulina”. Available online: https://www.anses.fr/en/system/files/NUT2014SA0096EN.pdf (accessed on 3 March 2023).
- GRAS Notice (GRN) No. 742. Available online: https://www.fda.gov/media/113614/download (accessed on 3 March 2023).
- Code of Federal Regulations. Available online: https://www.ecfr.gov/ (accessed on 3 March 2023).
- Algae as Novel Food in Europe. Available online: https://www.algae-novel-food.com/output/algae-novel-food/download.pdf (accessed on 3 March 2023).
- Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001 (Text with EEA Relevance); 2015; Volume 327. Available online: http://data.europa.eu/eli/reg/2015/2283/oj/eng (accessed on 3 March 2023).
- Bortolini, D.G.; Maciel, G.M.; Fernandes, I.D.A.A.; Pedro, A.C.; Rubio, F.T.V.; Branco, I.G.; Haminiuk, C.W.I. Functional properties of bioactive compounds from Spirulina spp.: Current status and future trends. Food Chem. Mol. Sci. 2022, 5, 100134. [Google Scholar] [CrossRef] [PubMed]
- Grosshagauer, S.; Kraemer, K.; Somoza, V. The True Value of Spirulina. J. Agric. Food Chem. 2020, 68, 4109–4115. [Google Scholar] [CrossRef] [PubMed]
- Blue-Green Algae: MedlinePlus Supplements. Available online: https://medlineplus.gov/druginfo/natural/923.html (accessed on 3 March 2023).
- AlFadhly, N.K.Z.; Alhelfi, N.; Altemimi, A.B.; Verma, D.K.; Cacciola, F.; Narayanankutty, A. Trends and Technological Advancements in the Possible Food Applications of Spirulina and Their Health Benefits: A Review. Molecules 2022, 27, 5584. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Fernández-Sevilla, J.M.; González-López, C.; Acién-Fernández, F.G. Spirulina for the food and functional food industries. Food Res. Int. 2020, 137, 109356. [Google Scholar] [CrossRef]
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina platensis, a super food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Manimekalan, A.; Parthiban, M.S. Reducing the Carbon Footprint by Cultivating and Consuming Spirulina: A Mini-review. Int. J. Environ. Clim. Chang. 2022, 12, 3069–3076. [Google Scholar] [CrossRef]
- de Medeiros, V.P.B.; da Costa, W.K.A.; da Silva, R.T.; Pimentel, T.C.; Magnani, M. Microalgae as source of functional ingredients in new-generation foods: Challenges, technological effects, biological activity, and regulatory issues. Crit. Rev. Food Sci. Nutr. 2022, 62, 4929–4950. [Google Scholar] [CrossRef]
- Ragusa, I.; Nardone, G.; Zanatta, S.; Bertin, W.; Amadio, E. Spirulina for Skin Care: A Bright Blue Future. Cosmetics 2021, 8, 7. [Google Scholar] [CrossRef]
- State of Technology Review—Algae Bioenergy. Available online: https://www.ieabioenergy.com/blog/publications/state-of-technology-review-algae-bioenergy (accessed on 3 March 2023).
- Spirulina Market by Distribution Channel (Consumer Channel, Business Channel), Product Type (Powder, Tablets, Capsules, Flakes, Phycocyanin Extract), Application (Nutraceuticals, Food and Beverages, Agriculture, Animal Feed)-Global Forecast to 2028. Available online: https://www.giiresearch.com/report/meti1018472-spirulina-market-by-distribution-channel-consumer.html (accessed on 3 March 2023).
- Thevarajah, B.; Nishshanka, G.K.S.H.; Premaratne, M.; Nimarshana, P.; Nagarajan, D.; Chang, J.-S.; Ariyadasa, T.U. Large-scale production of Spirulina-based proteins and c-phycocyanin: A biorefinery approach. Biochem. Eng. J. 2022, 185, 108541. [Google Scholar] [CrossRef]
- Altmann, B.A.; Rosenau, S. Spirulina as Animal Feed: Opportunities and Challenges. Foods 2022, 11, 965. [Google Scholar] [CrossRef]
- Costa, J.A.V.; Freitas, B.C.B.; Rosa, G.M.; Moraes, L.; Morais, M.G.; Mitchell, B.G. Operational and economic aspects of Spirulina-based biorefinery. Bioresour. Technol. 2019, 292, 121946. [Google Scholar] [CrossRef]
- MacColl, R. Cyanobacterial Phycobilisomes. J. Struct. Biol. 1998, 124, 311–334. [Google Scholar] [CrossRef]
- Fabris, M.; Abbriano, R.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiumparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef] [PubMed]
- Giwa, A.; Abuhantash, F.; Chalermthai, B.; Taher, H. Bio-Based Circular Economy and Polygeneration in Microalgal Produc-tion from Food Wastes: A Concise Review. Sustainability 2022, 14, 10759. [Google Scholar] [CrossRef]
- Bayramoglu, G.; Akbulut, A.; Arica, M.Y. Study of polyethyleneimine- and amidoxime-functionalized hybrid biomass of Spirulina (Arthrospira) platensis for adsorption of uranium (VI) ion. Environ. Sci. Pollut. Res. 2015, 22, 17998–18010. [Google Scholar] [CrossRef] [PubMed]
- Hsieh-Lo, M.; Castillo, G.; Ochoa-Becerra, M.A.; Mojica, L. Phycocyanin and phycoerythrin: Strategies to improve production yield and chemical stability. Algal Res. 2019, 42, 101600. [Google Scholar] [CrossRef]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- de Amarante, M.C.A.; Braga, A.R.C.; Sala, L.; Moraes, C.C.; Kalil, S.J. Design strategies for C-phycocyanin purification: Process influence on purity grade. Sep. Purif. Technol. 2020, 252, 117453. [Google Scholar] [CrossRef]
- C-Phycocyanin | Sigma-Aldrich. Available online: http://www.sigmaaldrich.com/ (accessed on 4 March 2023).
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- CFR-Code of Federal Regulations Title 21. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/CFRSearch.cfm?fr=73.530 (accessed on 4 March 2023).
- Provision of Scientific and Technical Support with Respect to the Classification of Extracts/Concentrates with Colouring Properties Either as Food Colours (Food Additives Falling under Regulation (EC) No 1333/2008) or Colouring Foods. JRC Publications Repository. Available online: https://publications.jrc.ec.europa.eu/repository/handle/JRC96974 (accessed on 4 March 2023).
- GRAS Notice (GRN) No. 1000. Available online: https://www.fda.gov/media/155000/download (accessed on 4 March 2023).
- Enciso, P.; Cabrerizo, F.M.; Gancheff, J.S.; Denis, P.A.; Cerdá, M.F. Phycocyanin as Potential Natural Dye for its Use in Photovoltaic Cells. J. Appl. Sol. Chem. Model. 2013, 2, 225–233. Available online: https://www.semanticscholar.org/paper/Phycocyanin-as-Potential-Natural-Dye-for-its-Use-in-Enciso-Cabrerizo/677db30d4b30102e704d1f311f05e526fc277a7e (accessed on 4 March 2023).
- Kraseasintra, O.; Tragoolpua, Y.; Pandith, H.; Khonkarn, R.; Pathom-Aree, W.; Pekkoh, J.; Pumas, C. Application of phycocyanin from Arthrospira (Spirulina) platensis as a hair dye. Front. Mar. Sci. 2022, 9, 1024988. [Google Scholar] [CrossRef]
- Dewi, E.N.; Kurniasih, R.A.; Purnamayati, L. The Application of Microencapsulated Phycocyanin as a Blue Natural Colorant to the Quality of Jelly Candy. IOP Conf. Ser. Earth Environ. Sci. 2018, 116, 012047. [Google Scholar] [CrossRef]
- Vernes, L.; Granvillain, P.; Chemat, F.; Vian, M. Phycocyanin from Arthrospira platensis. Production, Extraction and Analysis. Curr. Biotechnol. 2015, 4, 481–491. [Google Scholar] [CrossRef]
- Romay, C.; Armesto, J.; Remirez, D.; González, R.; Ledon, N.; García, I. Antioxidant and anti-inflammatory properties of C-phycocyanin from blue-green algae. Inflamm. Res. 1998, 47, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Safari, R.; Amiri, Z.R.; Kenari, R.E. Antioxidant and antibacterial activities of C-phycocyanin from common name Spirulina platensis. Iran. J. Fish. Sci. 2019, 19, 1911–1927. [Google Scholar] [CrossRef]
- Wu, H.-L.; Wang, G.-H.; Xiang, W.-Z.; Li, T.; He, H. Stability and Antioxidant Activity of Food-Grade Phycocyanin Isolated fromSpirulina platensis. Int. J. Food Prop. 2016, 19, 2349–2362. [Google Scholar] [CrossRef]
- Namasivayam, S.K.R.; Shivaramakrishnan, K.; Bharani, R.S.A. Potential antioxidative protein-pigment complex Spirulina platensis mediated food grade phycocyanin C -Extraction, purification, antioxidative activity and biocompatibility. Indian J. Biochem. Biophys. 2019, 56, 230–239. [Google Scholar]
- Grover, P.; Bhatnagar, A.; Kumari, N.; Bhatt, A.N.; Nishad, D.K.; Purkayastha, J. C-Phycocyanin-a novel protein from Spirulina platensis- In vivo toxicity, antioxidant and immunomodulatory studies. Saudi J. Biol. Sci. 2021, 28, 1853–1859. [Google Scholar] [CrossRef]
- Jang, Y.; Kim, B. Protective Effect of Spirulina-Derived C-Phycocyanin against Ultraviolet B-Induced Damage in HaCaT Cells. Medicina 2021, 57, 273. [Google Scholar] [CrossRef]
- Hsiao, G.; Chou, P.-H.; Shen, M.-Y.; Chou, D.-S.; Lin, C.-H.; Sheu, J.-R. C-Phycocyanin, a Very Potent and Novel Platelet Aggregation Inhibitor from Spirulina platensis. J. Agric. Food Chem. 2005, 53, 7734–7740. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.G.; Stankova, K.G.; Nikolov, V.N.; Georgieva, R.T.; Minkova, K.M.; Gigova, L.G.; Rupova, I.T.; Boteva, R.N. The biliprotein C-phycocyanin modulates the early radiation response: A pilot study. Mutat. Res. Toxicol. Environ. Mutagen. 2010, 695, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Varadharaj, S.; Ganesan, L.P.; Shobha, J.C.; Naidu, M.U.; Parinandi, N.L.; Tridandapani, S.; Kutala, V.K.; Kuppusamy, P. C-phycocyanin protects against ischemia-reperfusion injury of heart through involvement of p38 MAPK and ERK signaling. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H2136–H2145. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Zheng, S.; Lin, L.; Jiang, Q.; Yang, X. Protective effect of C-phycocyanin against carbon tetrachloride-induced hepatocyte damage in vitro and in vivo. Chem. Biol. Interact. 2010, 185, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Farooq, S.M.; Boppana, N.B.; Asokan, D.; Sekaran, S.D.; Shankar, E.M.; Li, C.; Gopal, K.; Bakar, S.A.; Karthik, H.S.; Ebrahim, A.S. C-Phycocyanin Confers Protection against Oxalate-Mediated Oxidative Stress and Mitochondrial Dysfunctions in MDCK Cells. PLoS ONE 2014, 9, e93056. [Google Scholar] [CrossRef]
- Ravi, M.; Tentu, S.; Baskar, G.; Prasad, S.R.; Raghavan, S.; Jayaprakash, P.; Jeyakanthan, J.; Rayala, S.K.; Venkatraman, G. Molecular mechanism of anti-cancer activity of phycocyanin in triple-negative breast cancer cells. BMC Cancer 2015, 15, 768. [Google Scholar] [CrossRef]
- Subhashini, J.; Mahipal, S.V.; Reddy, M.C.; Reddy, M.M.; Rachamallu, A.; Reddanna, P. Molecular mechanisms in C-Phycocyanin induced apoptosis in human chronic myeloid leukemia cell line-K562. Biochem. Pharmacol. 2004, 68, 453–462. [Google Scholar] [CrossRef]
- Arunasree, K.M.; Roy, K.R.; Reddy, N.P.; Dheeraj, B.; Reddy, G.V.; Reddanna, P. Alteration of mitochondrial membrane potential by Spirulina platensis C-phycocyanin induces apoptosis in the doxorubicinresistant human hepatocellular-carcinoma cell line HepG2. Biotechnol. Appl. Biochem. 2007, 47, 159–167. [Google Scholar] [CrossRef]
- Kunte, M.; Desai, K. The Inhibitory Effect of C-phycocyanin Containing Protein Extract (C-PC Extract) on Human Matrix Metalloproteinases (MMP-2 and MMP-9) in Hepatocellular Cancer Cell Line (HepG2). Protein J. 2017, 36, 186–195. [Google Scholar] [CrossRef]
- Saini, M.K.; Sanyal, S.N. Targeting angiogenic pathway for chemoprevention of experimental colon cancer using C-phycocyanin as cyclooxygenase-2 inhibitor. Biochem. Cell Biol. 2014, 92, 206–218. [Google Scholar] [CrossRef]
- Chen, T.; Wong, Y.-S. In Vitro Antioxidant and Antiproliferative Activities of Selenium-Containing Phycocyanin from Selenium-Enriched Spirulina platensis. J. Agric. Food Chem. 2008, 56, 4352–4358. [Google Scholar] [CrossRef] [PubMed]
- Nishanth, R.P.; Ramakrishna, B.; Jyotsna, R.G.; Roy, K.R.; Reddy, G.V.; Reddy, P.K.; Reddanna, P. C-Phycocyanin inhibits MDR1 through reactive oxygen species and cyclooxygenase-2 mediated pathways in human hepatocellular carcinoma cell line. Eur. J. Pharmacol. 2010, 649, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.-M.; Cheng, S.-N.; Wong, C.-S.; Kuo, Y.-L.; Chou, T.-C. Antiinflammatory and Antihyperalgesic Activity of C-Phycocyanin. Anesth. Analg. 2009, 108, 1303–1310. [Google Scholar] [CrossRef]
- Cherng, S.-C.; Cheng, S.-N.; Tarn, A.; Chou, T.-C. Anti-inflammatory activity of c-phycocyanin in lipopolysaccharide-stimulated RAW 264.7 macrophages. Life Sci. 2007, 81, 1431–1435. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.-O.; Lee, H.-H.; Kung, Y.-C.; Tsai, M.-F.; Chou, T.-C. Therapeutic Effect of C-Phycocyanin Extracted from Blue Green Algae in a Rat Model of Acute Lung Injury Induced by Lipopolysaccharide. Evid. Based Complement. Altern. Med. 2013, 2013, 916590. [Google Scholar] [CrossRef]
- Zhang, L.; Kong, D.; Huang, J.; Wang, Q.; Shao, L. The Therapeutic Effect and the Possible Mechanism of C-Phycocyanin in Lipopolysaccharide and Seawater-Induced Acute Lung Injury. Drug Des. Dev. Ther. 2022, 16, 1025–1040. [Google Scholar] [CrossRef]
- Pattarayan, D.; Rajarajan, D.; Ayyanar, S.; Palanichamy, R.; Subbiah, R. C-phycocyanin suppresses transforming growth factor-β1-induced epithelial mesenchymal transition in human epithelial cells. Pharmacol. Rep. 2017, 69, 426–431. [Google Scholar] [CrossRef]
- Liu, R.-Z.; Li, W.-J.; Zhang, J.-J.; Liu, Z.-Y.; Li, Y.; Liu, C.; Qin, S. The Inhibitory Effect of Phycocyanin Peptide on Pulmonary Fibrosis In Vitro. Mar. Drugs 2022, 20, 696. [Google Scholar] [CrossRef]
- Mohite, Y.; Shrivastava, N.; Sahu, D. Antimicrobial Activity of C-Phycocyanin from Arthrospira Platensis Isolated from Extreme Haloalkaline Environment of Lonar Lake. Semantic Scholar 2015. Available online: https://www.semanticscholar.org/paper/Antimicrobial-Activity-of-C-Phycocyanin-from-From-Mohite-Shrivastava/0ce72a6894d9670db805c76ac10247d48f3076a6 (accessed on 4 March 2023).
- Mohamed, S.A.; Osman, A.; Abo Eita, A.; Sitohy, M.Z. Estimation of antibacterial and antioxidant activities of phycocyanin isolated from Spirulina. Zagazig J. Agric. Res. 2018, 45, 657–666. [Google Scholar] [CrossRef]
- Sarada, D.V.L.; Kumar, C.S.; Rengasamy, R. Purified C-phycocyanin from Spirulina platensis (Nordstedt) Geitler: A novel and potent agent against drug resistant bacteria. World J. Microbiol. Biotechnol. 2011, 27, 779–783. [Google Scholar] [CrossRef]
- Nihal, B.; Gupta, N.V.; Gowda, D.V.; Manohar, M. Formulation and development of topical anti acne formulation of spirulina extract. Int. J. Appl. Pharm. 2018, 10, 229–233. [Google Scholar] [CrossRef]
- Murugan, T.; Radhamadhavan. Screening for Antifungal and Antiviral Activity of C-Phycocyanin from Spirulina Platensis. Association of Pharmaceutical Innovators 2011. Available online: https://scholar.google.com/scholar_lookup?title=Screening+for+Antifungal+and+Antiviral+activity+of+C-phycocyanin+from+Spirulina+platensis&author=T.Murugan+1+&publication_year=2011 (accessed on 4 March 2023).
- Pentón-Rol, G.; Martínez-Sánchez, G.; Cervantes-Llanos, M.; Lagumersindez-Denis, N.; Acosta-Medina, E.F.; Falcón-Cama, V.; Alonso-Ramírez, R.; Valenzuela-Silva, C.; Rodríguez-Jiménez, E.; Llópiz-Arzuaga, A.; et al. C-Phycocyanin ameliorates experimental autoimmune encephalomyelitis and induces regulatory T cells. Int. Immunopharmacol. 2011, 11, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Koh, E.-J.; Kim, K.-J.; Choi, J.; Kang, D.-H.; Lee, B.-Y. Spirulina maxima extract prevents cell death through BDNF activation against amyloid beta 1-42 (Aβ 1-42) induced neurotoxicity in PC12 cells. Neurosci. Lett. 2018, 673, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Perumal, Y.; Bansal, S.; Arora, S.; Chopra, K. Phycocyanin alleviates ICV-STZ induced cognitive and molecular deficits via PI3-Kinase dependent pathway. Food Chem. Toxicol. 2020, 145, 111684. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gan, L.; Yan, S.; Yan, Y.; Huang, W. Effect of C-phycocyanin on HDAC3 and miRNA-335 in Alzheimer’s disease. Transl. Neurosci. 2020, 11, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jovcevski, B.; Pukala, T.L. C-Phycocyanin from Spirulina Inhibits α-Synuclein and Amyloid-β Fibril Formation but Not Amorphous Aggregation. J. Nat. Prod. 2019, 82, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, G.; Sampathkumar, P.; Kavisri, M.; Moovendhan, M. Extraction and characterization of phycocyanin from Spirulina platensis and evaluation of its anticancer, antidiabetic and antiinflammatory effect. Int. J. Biol. Macromol. 2020, 153, 256–263. [Google Scholar] [CrossRef]
- Husain, A.; Alouffi, S.; Khanam, A.; Akasha, R.; Farooqui, A.; Ahmad, S. Therapeutic Efficacy of Natural Product ‘C-Phycocyanin’ in Alleviating Streptozotocin-Induced Diabetes via the Inhibition of Glycation Reaction in Rats. Int. J. Mol. Sci. 2022, 23, 14235. [Google Scholar] [CrossRef]
- Ou, Y.; Lin, L.; Yang, X.; Pan, Q.; Cheng, X. Antidiabetic potential of phycocyanin: Effects on KKAy mice. Pharm. Biol. 2013, 51, 539–544. [Google Scholar] [CrossRef]
- Hao, S.; Li, F.; Li, Q.; Yang, Q.; Zhang, W. Phycocyanin Protects against High Glucose High Fat Diet Induced Diabetes in Mice and Participates in AKT and AMPK Signaling. Foods 2022, 11, 3183. [Google Scholar] [CrossRef]
- El-Sayed, E.-S.M.; Hikal, M.S.; Abo El-Khair, B.E.; El-Ghobashy, R.E.; El-Assar, A.M. Hypoglycemic and hypolipidemic effects of spirulina platensis, phycocyanin, phycocyanopeptide and phycocyanobilin on male diabetic rats. Arab. Univ. J. Agric. Sci. 2018, 26, 1121–1134. [Google Scholar] [CrossRef]
- Khan, M.; Shobha, J.C.; Mohan, I.K.; Naidu, M.U.R.; Prayag, A.; Kutala, V.K. Spirulina attenuates cyclosporine-induced nephrotoxicity in rats. J. Appl. Toxicol. 2006, 26, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.B.; Madyastha, K. C-Phycocyanin: A Potent Peroxyl Radical Scavenger in Vivo and in Vitro. Biochem. Biophys. Res. Commun. 2000, 275, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rojas, B.; Medina-Campos, O.N.; Hernández-Pando, R.; Negrette-Guzmán, M.; Huerta-Yepez, S.; Pedraza-Chaverri, J. C-Phycocyanin prevents cisplatin-induced nephrotoxicity through inhibition of oxidative stress. Food Funct. 2014, 5, 480–490. [Google Scholar] [CrossRef]
- Zheng, J.; Inoguchi, T.; Sasaki, S.; Maeda, Y.; McCarty, M.F.; Fujii, M.; Ikeda, N.; Kobayashi, K.; Sonoda, N.; Takayanagi, R. Phycocyanin and phycocyanobilin from Spirulina platensis protect against diabetic nephropathy by inhibiting oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R110–R120. [Google Scholar] [CrossRef]
- Lim, B.J.; Jeong, J.Y.; Chang, Y.-K.; Na, K.R.; Lee, K.W.; Shin, Y.-T.; Choi, D.E. C-Phycocyanin Attenuates Cisplatin-Induced Nephrotoxicity in Mice. Ren. Fail. 2012, 34, 892–900. [Google Scholar] [CrossRef]
- Strasky, Z.; Zemankova, L.; Nemeckova, I.; Rathouska, J.; Wong, R.J.; Muchova, L.; Subhanova, I.; Vanikova, J.; Vanova, K.; Vitek, L.; et al. Spirulina platensis and phycocyanobilin activate atheroprotective heme oxygenase-1: A possible implication for atherogenesis. Food Funct. 2013, 4, 1586–1594. [Google Scholar] [CrossRef]
- Blas-Valdivia, V.; Moran-Dorantes, D.N.; Rojas-Franco, P.; Franco-Colin, M.; Mirhosseini, N.; Davarnejad, R.; Halajisani, A.; Tavakoli, O.; Cano-Europa, E. C-Phycocyanin prevents acute myocardial infarction-induced oxidative stress, inflammation and cardiac damage. Pharm. Biol. 2022, 60, 755–763. [Google Scholar] [CrossRef]
- Sheu, M.-J.; Hsieh, Y.-Y.; Lai, C.-H.; Chang, C.-C.; Wu, C.-H. Antihyperlipidemic and Antioxidant Effects of C-phycocyanin in Golden Syrian Hamsters Fed with a Hypercholesterolemic Diet. J. Tradit. Complement. Med. 2013, 3, 41–47. [Google Scholar] [CrossRef]
- Ichimura, M.; Kato, S.; Tsuneyama, K.; Matsutake, S.; Kamogawa, M.; Hirao, E.; Miyata, A.; Mori, S.; Yamaguchi, N.; Suruga, K.; et al. Phycocyanin prevents hypertension and low serum adiponectin level in a rat model of metabolic syndrome. Nutr. Res. 2013, 33, 397–405. [Google Scholar] [CrossRef]
- Seo, Y.-J.; Kim, K.-J.; Choi, J.; Koh, E.-J.; Lee, B.-Y. Spirulina maxima Extract Reduces Obesity through Suppression of Adipogenesis and Activation of Browning in 3T3-L1 Cells and High-Fat Diet-Induced Obese Mice. Nutrients 2018, 10, 712. [Google Scholar] [CrossRef] [PubMed]
- Madhyastha, H.K.; Radha, K.S.; Nakajima, Y.; Omura, S.; Maruyama, M. uPA dependent and independent mechanisms of wound healing by C-phycocyanin. J. Cell. Mol. Med. 2008, 12, 2691–2703. [Google Scholar] [CrossRef]
- Madhyastha, H.; Radha, K.; Sugiki, M.; Omura, S.; Maruyama, M. C-phycocyanin transcriptionally regulates uPA mRNA through cAMP mediated PKA pathway in human fibroblast WI-38 cells. Biochim. Biophys. Acta Gen. Subj. 2006, 1760, 1624–1630. [Google Scholar] [CrossRef]
- Kose, A.; Oncel, S.S. Design of melanogenesis regulatory peptides derived from phycocyanin of the microalgae Spirulina platensis. Peptides 2022, 152, 170783. [Google Scholar] [CrossRef]
- Wu, L.-C.; Lin, Y.-Y.; Yang, S.-Y.; Weng, Y.-T.; Tsai, Y.-T. Antimelanogenic effect of c-phycocyanin through modulation of tyrosinase expression by upregulation of ERK and downregulation of p38 MAPK signaling pathways. J. Biomed. Sci. 2011, 18, 74. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rojas, B.; Hernández-Juárez, J.; Pedraza-Chaverri, J. Nutraceutical properties of phycocyanin. J. Funct. Foods 2014, 11, 375–392. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Pizarro, M.; Lissi, E. Kinetics of c-Phycocyanin Reaction with Hypochlorite. J. Protein Chem. 2000, 19, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Bhat, V.B.; Madyastha, K. Scavenging of Peroxynitrite by Phycocyanin and Phycocyanobilin from Spirulina platensis: Protection against Oxidative Damage to DNA. Biochem. Biophys. Res. Commun. 2001, 285, 262–266. [Google Scholar] [CrossRef]
- Pleonsil, P.; Soogarun, S.; Suwanwong, Y. Anti-oxidant activity of holo- and apo-c-phycocyanin and their protective effects on human erythrocytes. Int. J. Biol. Macromol. 2013, 60, 393–398. [Google Scholar] [CrossRef]
- Patel, A.; Mishra, S.; Pawar, R.; Ghosh, P. Purification and characterization of C-Phycocyanin from cyanobacterial species of marine and freshwater habitat. Protein Expr. Purif. 2005, 40, 248–255. [Google Scholar] [CrossRef]
- Xu, F.; Zhang, Y.; Qiu, Y.; Yang, F.; Liu, G.; Dong, X.; Chen, G.; Cao, C.; Zhang, Q.; Zhang, S.; et al. Three novel antioxidant peptides isolated from C-phycocyanin against H2O2-induced oxidative stress in zebrafish via Nrf2 signaling pathway. Front. Mar. Sci. 2022, 9, 1098091. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Qin, S. Therapeutic effect of phycocyanin on acute liver oxidative damage caused by X-ray. Biomed. Pharmacother. 2020, 130, 110553. [Google Scholar] [CrossRef] [PubMed]
- Dranseikienė, D.; Balčiūnaitė-Murzienė, G.; Karosienė, J.; Morudov, D.; Juodžiukynienė, N.; Hudz, N.; Gerbutavičienė, R.J.; Savickienė, N. Cyano-Phycocyanin: Mechanisms of Action on Human Skin and Future Perspectives in Medicine. Plants 2022, 11, 1249. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, Y.; Yin, Q.; Liu, G.; Liu, H.; Huang, Y.; Li, B. Phycocyanin: A Potential Drug for Cancer Treatment. J. Cancer 2017, 8, 3416–3429. [Google Scholar] [CrossRef] [PubMed]
- Morcos, N.C.; Berns, M.; Henry, W.L. Phycocyanin: Laser activation, cytotoxic effects, and uptake in human atherosclerotic plaque. Lasers Surg. Med. 1988, 8, 10–17. [Google Scholar] [CrossRef]
- Thangam, R.; Suresh, V.; Princy, W.A.; Rajkumar, M.; SenthilKumar, N.; Gunasekaran, P.; Rengasamy, R.; Anbazhagan, C.; Kaveri, K.; Kannan, S. C-Phycocyanin from Oscillatoria tenuis exhibited an antioxidant and in vitro antiproliferative activity through induction of apoptosis and G0/G1 cell cycle arrest. Food Chem. 2013, 140, 262–272. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- AMR Review Paper—Tackling a Crisis for the Health and Wealth of Nations_1.Pdf. Available online: https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tackling%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf (accessed on 4 March 2023).
- Lee, Y.B.; Byun, E.J.; Kim, H.S. Potential Role of the Microbiome in Acne: A Comprehensive Review. J. Clin. Med. 2019, 8, 987. [Google Scholar] [CrossRef]
- Habib, R.; Noureen, N.; Nadeem, N. Decoding Common Features of Neurodegenerative Disorders: From Differentially Expressed Genes to Pathways. Curr. Genom. 2018, 19, 300–312. [Google Scholar] [CrossRef]
- Pentón-Rol, G.; Marín-Prida, J.; McCarty, M.F. C-Phycocyanin-derived Phycocyanobilin as a Potential Nutraceutical Approach for Major Neurodegenerative Disorders and COVID-19-induced Damage to the Nervous System. Curr. Neuropharmacol. 2021, 19, 2250–2275. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Tanaka, M.; Ooike, M.; Tsunomura, T.; Sakaguchi, M. Antioxidant activities of phycocyanobilin prepared from Spirulina platensis. J. Appl. Phycol. 2000, 12, 435–439. [Google Scholar] [CrossRef]
- Pentón-Rol, G.; Marín-Prida, J.; Falcón-Cama, V. C-Phycocyanin and Phycocyanobilin as Remyelination Therapies for Enhancing Recovery in Multiple Sclerosis and Ischemic Stroke: A Preclinical Perspective. Behav. Sci. 2018, 8, 15. [Google Scholar] [CrossRef]
- Hua, P.; Yu, Z.; Xiong, Y.; Liu, B.; Zhao, L. Regulatory Efficacy of Spirulina platensis Protease Hydrolyzate on Lipid Metabolism and Gut Microbiota in High-Fat Diet-Fed Rats. Int. J. Mol. Sci. 2018, 19, 4023. [Google Scholar] [CrossRef]
- Xie, Y.; Li, W.; Zhu, L.; Zhai, S.; Qin, S.; Du, Z. Effects of phycocyanin in modulating the intestinal microbiota of mice. Microbiologyopen 2019, 8, e00825. [Google Scholar] [CrossRef] [PubMed]
- Elbialy, Z.I.; Assar, D.H.; Abdelnaby, A.; Abu Asa, S.; Abdelhiee, E.Y.; Ibrahim, S.S.; Abdel-Daim, M.M.; Almeer, R.; Atiba, A. Healing potential of Spirulina platensis for skin wounds by modulating bFGF, VEGF, TGF-ß1 and α-SMA genes expression targeting angiogenesis and scar tissue formation in the rat model. Biomed. Pharmacother. 2021, 137, 111349. [Google Scholar] [CrossRef]
- Munawaroh, H.S.H.; Gumilar, G.G.; Nurjanah, F.; Yuliani, G.; Aisyah, S.; Kurnia, D.; Wulandari, A.P.; Kurniawan, I.; Ningrum, A.; Koyandev, A.K.; et al. In-vitro molecular docking analysis of microalgae extracted phycocyanin as an anti-diabetic candidate. Biochem. Eng. J. 2020, 161, 107666. [Google Scholar] [CrossRef]
- Ashaolu, T.J.; Samborska, K.; Lee, C.C.; Tomas, M.; Capanoglu, E.; Tarhan, Ö.; Taze, B.; Jafari, S.M. Phycocyanin, a super functional ingredient from algae; properties, purification characterization, and applications. Int. J. Biol. Macromol. 2021, 193 Pt B, 2320–2331. [Google Scholar] [CrossRef]
- Wen, X.; Han, Z.; Liu, S.-J.; Hao, X.; Zhang, X.-J.; Wang, X.-Y.; Zhou, C.-J.; Ma, Y.-Z.; Liang, C.-G. Phycocyanin Improves Reproductive Ability in Obese Female Mice by Restoring Ovary and Oocyte Quality. Front. Cell Dev. Biol. 2020, 8, 595373. [Google Scholar] [CrossRef]
- Syarina, P.N.A.; Karthivashan, G.; Abas, F.; Arulselvan, P.; Fakurazi, S. Wound healing potential of Spirulina platensis extracts on human dermal fibroblast cells. EXCLI J. 2015, 14, 385–393. [Google Scholar] [CrossRef]
- Liu, P.; Choi, J.; Lee, M.; Choi, Y.H.; Nam, T. Spirulina protein promotes skin wound repair in a mouse model of full-thickness dermal excisional wound. Int. J. Mol. Med. 2020, 46, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Chen, J.; Lu, J.; Yi, L.; Tong, X.; Kang, L.; Pei, S.; Ouyang, Y.; Jiang, L.; Ding, Y.; et al. Roles of inflammation factors in melanogenesis. Mol. Med. Rep. 2020, 21, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Favas, R.; Morone, J.; Martins, R.; Vasconcelos, V.; Lopes, G. Cyanobacteria and microalgae bioactive compounds in skin-ageing: Potential to restore extracellular matrix filling and overcome hyperpigmentation. J. Enzym. Inhib. Med. Chem. 2021, 36, 1829–1838. [Google Scholar] [CrossRef] [PubMed]
- Nicolaidou, E.; Katsambas, A.D. Pigmentation disorders: Hyperpigmentation and hypopigmentation. Clin. Dermatol. 2014, 32, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D.; Cassidy, C.; Horga, G.; Kang, U.J.; Fahn, S.; Casella, L.; Pezzoli, G.; Langley, J.; Hu, X.P.; Zucca, F.A.; et al. Neuromelanin detection by magnetic resonance imaging (MRI) and its promise as a biomarker for Parkinson’s disease. npj Park. Dis. 2018, 4, 11. [Google Scholar] [CrossRef]
- Mahendra, J.; Mahendra, L.; Muthu, J.; John, L.; Romanos, G.E. Clinical Effects of Subgingivally Delivered Spirulina Gel in Chronic Periodontitis Cases: A Placebo Controlled Clinical Trial. J. Clin. Diagn. Res. 2013, 7, 2330–2333. [Google Scholar] [CrossRef]
- Patil, S.; Al-Zarea, B.K.; Maheshwari, S.; Sahu, R. Comparative evaluation of natural antioxidants spirulina and aloe vera for the treatment of oral submucous fibrosis. J. Oral Biol. Craniofacial Res. 2015, 5, 11–15. [Google Scholar] [CrossRef]
- Ge, Y.; Kang, Y.-K.; Dong, L.; Liu, L.-H.; An, G.-Y. The efficacy of dietary Spirulina as an adjunct to chemotherapy to improve immune function and reduce myelosuppression in patients with malignant tumors. Transl. Cancer Res. 2019, 8, 1065–1073. [Google Scholar] [CrossRef]
- Tavanandi, H.A.; Mittal, R.; Chandrasekhar, J.; Raghavarao, K. Simple and efficient method for extraction of C-Phycocyanin from dry biomass of Arthospira platensis. Algal Res. 2018, 31, 239–251. [Google Scholar] [CrossRef]
- Silveira, S.T.; Burkert, J.F.D.M.; Costa, J.A.V.; Burkert, C.A.V.; Kalil, S.J. Optimization of phycocyanin extraction from Spirulina platensis using factorial design. Bioresour. Technol. 2007, 98, 1629–1634. [Google Scholar] [CrossRef]
- Chentir, I.; Hamdi, M.; Li, S.; Doumandji, A.; Markou, G.; Nasri, M. Stability, bio-functionality and bio-activity of crude phycocyanin from a two-phase cultured Saharian Arthrospira sp. strain. Algal Res. 2018, 35, 395–406. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Nunes, R.; De Biasio, F.; Spigno, G.; Gorgoglione, D.; Teixeira, J.A.; Rocha, C.M. Influence of thermal and electrical effects of ohmic heating on C-phycocyanin properties and biocompounds recovery from Spirulina platensis. LWT 2020, 128, 109491. [Google Scholar] [CrossRef]
- Ilter, I.; Akyıl, S.; Demirel, Z.; Koç, M.; Conk-Dalay, M.; Kaymak-Ertekin, F. Optimization of phycocyanin extraction from Spirulina platensis using different techniques. J. Food Compos. Anal. 2018, 70, 78–88. [Google Scholar] [CrossRef]
- Pan-Utai, W.; Iamtham, S. Extraction, purification and antioxidant activity of phycobiliprotein from Arthrospira platensis. Process. Biochem. 2019, 82, 189–198. [Google Scholar] [CrossRef]
- Käferböck, A.; Smetana, S.; de Vos, R.; Schwarz, C.; Toepfl, S.; Parniakov, O. Sustainable extraction of valuable components from Spirulina assisted by pulsed electric fields technology. Algal Res. 2020, 48, 101914. [Google Scholar] [CrossRef]
- Pott, R.W.M. The release of the blue biological pigment C-phycocyanin through calcium-aided cytolysis of live Spirulina sp. Color. Technol. 2019, 135, 17–21. [Google Scholar] [CrossRef]
- Martínez, J.M.; Luengo, E.; Saldaña, G.; Álvarez, I.; Raso, J. C-phycocyanin extraction assisted by pulsed electric field from Artrosphira platensis. Food Res. Int. 2017, 99 Pt 3, 1042–1047. [Google Scholar] [CrossRef]
- Jaeschke, D.P.; Mercali, G.D.; Marczak, L.D.F.; Müller, G.; Frey, W.; Gusbeth, C. Extraction of valuable compounds from Arthrospira platensis using pulsed electric field treatment. Bioresour. Technol. 2019, 283, 207–212. [Google Scholar] [CrossRef]
- Liao, X.; Zhang, B.; Wang, X.; Yan, H.; Zhang, X. Purification of C-Phycocyanin from Spirulina platensis by Single-Step Ion-Exchange Chromatography. Chromatographia 2011, 73, 291–296. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Paciulli, M.; Abbaspourrad, A. Extraction of phycocyanin—A natural blue colorant from dried spirulina biomass: Influence of processing parameters and extraction techniques. J. Food Sci. 2020, 85, 727–735. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Jáuregui, M.; Medina, E.; Jaime, C.; Cerezal, P. Rapid Green Extractions of C-Phycocyanin from Arthrospira maxima for Functional Applications. Appl. Sci. 2019, 9, 1987. [Google Scholar] [CrossRef]
- Giannoglou, M.; Andreou, V.; Thanou, I.; Markou, G.; Katsaros, G. High pressure assisted extraction of proteins from wet biomass of Arthrospira platensis (spirulina)—A kinetic approach. Innov. Food Sci. Emerg. Technol. 2022, 81, 103138. [Google Scholar] [CrossRef]
- Tavanandi, H.A.; Raghavarao, K. Ultrasound-assisted enzymatic extraction of natural food colorant C-Phycocyanin from dry biomass of Arthrospira platensis. LWT 2020, 118, 108802. [Google Scholar] [CrossRef]
- Acker, J.P.; McGann, L.E. Protective effect of intracellular ice during freezing? Cryobiology 2003, 46, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Ores, J.D.C.; de Amarante, M.C.A.; Kalil, S.J. Co-production of carbonic anhydrase and phycobiliproteins by Spirulina sp. and Synechococcus nidulans. Bioresour. Technol. 2016, 219, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.S.; Lo, C.; Eppink, M.; Wijffels, R.; van den Berg, C. Understanding mild cell disintegration of microalgae in bead mills for the release of biomolecules. Chem. Eng. Sci. 2019, 203, 380–390. [Google Scholar] [CrossRef]
- Günerken, E.; D’Hondt, E.; Eppink, M.; Garcia-Gonzalez, L.; Elst, K.; Wijffels, R. Cell disruption for microalgae biorefineries. Biotechnol. Adv. 2015, 33, 243–260. [Google Scholar] [CrossRef]
- Montalescot, V.; Rinaldi, T.; Touchard, R.; Jubeau, S.; Frappart, M.; Jaouen, P.; Bourseau, P.; Marchal, L. Optimization of bead milling parameters for the cell disruption of microalgae: Process modeling and application to Porphyridium cruentum and Nannochloropsis oculata. Bioresour. Technol. 2015, 196, 339–346. [Google Scholar] [CrossRef]
- Soto-Sierra, L.; Stoykova, P.; Nikolov, Z.L. Extraction and fractionation of microalgae-based protein products. Algal Res. 2018, 36, 175–192. [Google Scholar] [CrossRef]
- Tiwari, B.K. Ultrasound: A clean, green extraction technology. TrAC Trends Anal. Chem. 2015, 71, 100–109. [Google Scholar] [CrossRef]
- Vernès, L.; Abert-Vian, M.; El Maâtaoui, M.; Tao, Y.; Bornard, I.; Chemat, F. Application of ultrasound for green extraction of proteins from spirulina. Mechanism, optimization, modeling, and industrial prospects. Ultrason. Sonochem. 2019, 54, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Vorobiev, E.; Lebovka, N. 2–Extraction from Foods and Biomaterials Enhanced by Pulsed Electric Energy. In Innovative Food Processing Technologies; Knoerzer, K., Juliano, P., Smithers, G., Eds.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2016; pp. 31–56. [Google Scholar] [CrossRef]
- Lebovka, N.; Vorobiev, E. (Eds.) Electrotechnologies for Extraction from Food Plants and Biomaterials; Food Engineering Series; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Song, W.; Zhao, C.; Wang, S. A Large-Scale Preparation Method of High Purity C-Phycocyanin. Int. J. Biosci. Biochem. Bioinform. 2013, 3, 293–297. Available online: http://www.ijbbb.org/papers/216-S007.pdf. (accessed on 4 March 2023).
- Lee, C.-W.; Bae, G.Y.; Bae, S.-H.; Suh, H.J.; Jo, K. Increased thermal stability of phycocyanin extracted from Spirulina platensis by cysteine addition during enzyme extraction. Food Sci. Technol. 2022, 42, e15021. [Google Scholar] [CrossRef]
- Patil, G.; Raghavarao, K.S.M.S. Aqueous two phase extraction for purification of C-phycocyanin. Biochem. Eng. J. 2007, 34, 156–164. [Google Scholar] [CrossRef]
- Chaiklahan, R.; Chirasuwan, N.; Bunnag, B. Stability of phycocyanin extracted from Spirulina sp.: Influence of temperature, pH and preservatives. Process. Biochem. 2012, 47, 659–664. [Google Scholar] [CrossRef]
- Antelo, F.S.; Costa, J.A.; Kalil, S.J. Thermal degradation kinetics of the phycocyanin from Spirulina platensis. Biochem. Eng. J. 2008, 41, 43–47. [Google Scholar] [CrossRef]
- Patel, A.; Pawar, R.; Mishra, S.; Sonawane, S.; Ghosh, P.K. Kinetic studies on thermal denaturation of C-phycocyanin. Indian J. Biochem. Biophys. 2004, 41, 254–257. [Google Scholar]
- Böcker, L.; Ortmann, S.; Surber, J.; Leeb, E.; Reineke, K.; Mathys, A. Biphasic short time heat degradation of the blue microalgae protein phycocyanin from Arthrospira platensis. Innov. Food Sci. Emerg. Technol. 2019, 52, 116–121. [Google Scholar] [CrossRef]
- Su, C.-H.; Liu, C.-S.; Yang, P.-C.; Syu, K.-S.; Chiuh, C.-C. Solid–liquid extraction of phycocyanin from Spirulina platensis: Kinetic modeling of influential factors. Sep. Purif. Technol. 2014, 123, 64–68. [Google Scholar] [CrossRef]
- Wicaksono, H.A.; Satyantini, W.H.; Masithah, E.D. The spectrum of light and nutrients required to increase the production of phycocyanin Spirulina platensis. IOP Conf. Ser. Earth Environ. Sci. 2019, 236, 012008. [Google Scholar] [CrossRef]
- Walter, A.; De Carvalho, J.C.; Soccol, V.T.; De Faria, A.B.B.; Ghiggi, V.; Soccol, C.R. Study of phycocyanin production from Spirulina platensis under different light spectra. Braz. Arch. Biol. Technol. 2011, 54, 675–682. [Google Scholar] [CrossRef]
- Escalante, F.M.E.; Pérez-Rico, D.A.; Alarcón-Jiménez, J.L.; González-Morales, E.; Guerra-Álvarez, L.F.; Ramírez-Vázquez, J.C.; Gutiérrez-Pulido, H. Phycocyanin Thermo-photostability: An Accelerated Life-test Analysis. J. Mex. Chem. Soc. 2020, 64, 218–229. [Google Scholar] [CrossRef]
- Choi, W.Y.; Lee, H.Y. Kinetic Analysis of Stabilizing C-Phycocyanin in the Spirulina platensis Extracts from Ultrasonic Process Associated with Effects of Light and Temperature. Appl. Sci. 2018, 8, 1662. [Google Scholar] [CrossRef]
- Aftari, R.V.; Rezaei, K.; Mortazavi, A.; Bandani, A.R. The Optimized Concentration and Purity of Spirulina platensis C-Phycocyanin: A Comparative Study on Microwave-Assisted and Ultrasound-Assisted Extraction Methods. J. Food Process. Preserv. 2015, 39, 3080–3091. [Google Scholar] [CrossRef]
- Wachda; Harjanto, G.D.; Huzain, M.L.; Hadiyanto, H.; Aji, R.W. Production of antioxidant C-phycocyanin using extraction process of Spirulina platensis in large scale industry. IOP Conf. Ser. Mater. Sci. Eng. 2019, 633, 012025. [Google Scholar] [CrossRef]
- Khandual, S.; Sanchez, E.O.L.; Andrews, H.E.; de la Rosa, J.D.P. Phycocyanin content and nutritional profile of Arthrospira platensis from Mexico: Efficient extraction process and stability evaluation of phycocyanin. BMC Chem. 2021, 15, 24. [Google Scholar] [CrossRef] [PubMed]
- Gorgich, M.; Passos, M.L.; Mata, T.M.; Martins, A.A.; Saraiva, M.L.M.; Caetano, N.S. Enhancing extraction and purification of phycocyanin from Arthrospira sp. with lower energy consumption. Energy Rep. 2020, 6, 312–318. [Google Scholar] [CrossRef]
- Adjali, A.; Clarot, I.; Chen, Z.; Marchioni, E.; Boudier, A. Physicochemical degradation of phycocyanin and means to improve its stability: A short review. J. Pharm. Anal. 2022, 12, 406–414. [Google Scholar] [CrossRef]
- Faieta, M.; Neri, L.; Sacchetti, G.; Di Michele, A.; Pittia, P. Role of saccharides on thermal stability of phycocyanin in aqueous solutions. Food Res. Int. 2020, 132, 109093. [Google Scholar] [CrossRef]
- Hadiyanto; Christwardana, M.; Sutanto, H.; Suzery, M.; Amelia, D.; Aritonang, R.F. Kinetic study on the effects of sugar addition on the thermal degradation of phycocyanin from Spirulina sp. Food Biosci. 2018, 22, 85–90. [Google Scholar] [CrossRef]
- Pan-utai, W.; Kahapana, W.; Iamtham, S. Extraction of C-phycocyanin from Arthrospira (Spirulina) and its thermal stability with citric acid. J. Appl. Phycol. 2018, 30, 231–242. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shrivastav, A.; Mishra, S. Effect of preservatives for food grade C-PC from Spirulina platensis. Process. Biochem. 2008, 43, 339–345. [Google Scholar] [CrossRef]
- Sun, L.; Wang, S.; Qiao, Z. Chemical stabilization of the phycocyanin from cyanobacterium Spirulina platensis. J. Biotechnol. 2006, 121, 563–569. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Abbaspourrad, A. Improvement of the colloidal stability of phycocyanin in acidified conditions using whey protein-phycocyanin interactions. Food Hydrocoll. 2020, 105, 105747. [Google Scholar] [CrossRef]
- Zhang, Z.; Cho, S.; Dadmohammadi, Y.; Li, Y.; Abbaspourrad, A. Improvement of the storage stability of C-phycocyanin in beverages by high-pressure processing. Food Hydrocoll. 2020, 110, 106055. [Google Scholar] [CrossRef]
- Moreira, J.B.; Lim, L.-T.; Zavareze, E.D.R.; Dias, A.R.G.; Costa, J.A.V.; de Morais, M.G. Antioxidant ultrafine fibers developed with microalga compounds using a free surface electrospinning. Food Hydrocoll. 2019, 93, 131–136. [Google Scholar] [CrossRef]
- Hadiyanto, H.; Christwardana, M.; Suzery, M.; Sutanto, H.; Nilamsari, A.M.; Yunanda, A. Effects of Carrageenan and Chitosan as Coating Materials on the Thermal Degradation of Microencapsulated Phycocyanin from Spirulina sp. Int. J. Food Eng. 2019, 15, 20180290. [Google Scholar] [CrossRef]
- Pan-Utai, W.; Iamtham, S. Enhanced Microencapsulation of C-Phycocyanin from Arthrospira by Freeze-Drying with Different Wall Materials. Food Technol. Biotechnol. 2020, 58, 423–432. [Google Scholar] [CrossRef]
- Schmatz, D.A.; da Silveira Mastrantonio, D.J.; Costa, J.A.V.; de Morais, M.G. Encapsulation of phycocyanin by electrospraying: A promising approach for the protection of sensitive compounds. Food Bioprod. Process. 2019, 119, 206–215. [Google Scholar] [CrossRef]
- Pradeep, H.N.; Nayak, C.A. Enhanced stability of C-phycocyanin colorant by extrusion encapsulation. J. Food Sci. Technol. 2019, 56, 4526–4534. [Google Scholar] [CrossRef]
- Ilter, I.; Koç, M.; Demirel, Z.; Dalay, M.C.; Ertekin, F.K. Improving the stability of phycocyanin by spray dried microencapsulation. J. Food Process. Preserv. 2021, 45, e15646. [Google Scholar] [CrossRef]
- He, S.; Joseph, N.; Feng, S.; Jellicoe, M.; Raston, C.L. Application of microfluidic technology in food processing. Food Funct. 2020, 11, 5726–5737. [Google Scholar] [CrossRef]
- Yan, S.-G.; Zhu, L.-P.; Su, H.-N.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhang, Y.-Z. Single-step chromatography for simultaneous purification of C-phycocyanin and allophycocyanin with high purity and recovery from Spirulina (Arthrospira) platensis. J. Appl. Phycol. 2011, 23, 1–6. [Google Scholar] [CrossRef]
- Minic, S.L.; Stanic-Vucinic, D.; Mihailovic, J.; Krstic, M.; Nikolic, M.R.; Velickovic, T.C. Digestion by pepsin releases biologically active chromopeptides from C-phycocyanin, a blue-colored biliprotein of microalga Spirulina. J. Proteom. 2016, 147, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.-H.; Wang, S.S.-S.; Show, P.-L.; Hsu, S.-L.; Chang, Y.-K. Rapid and efficient recovery of C-phycocyanin from highly turbid Spirulina platensis algae using stirred fluidized bed ion exchange chromatography. Sep. Purif. Technol. 2019, 209, 636–645. [Google Scholar] [CrossRef]
- de Amarante, M.C.A.; Júnior, L.C.S.C.; Sala, L.; Kalil, S.J. Analytical grade C-phycocyanin obtained by a single-step purification process. Process. Biochem. 2020, 90, 215–222. [Google Scholar] [CrossRef]
- Lauceri, R.; Zittelli, G.C.; Maserti, B.; Torzillo, G. Purification of phycocyanin from Arthrospira platensis by hydrophobic interaction membrane chromatography. Algal Res. 2018, 35, 333–340. [Google Scholar] [CrossRef]
- Figueira, F.D.S.; Moraes, C.C.; Kalil, S.J. C-Phycocyanin purification: Multiple processes for different applications. Braz. J. Chem. Eng. 2018, 35, 1117–1128. [Google Scholar] [CrossRef]
- Moraes, C.C.; Kalil, S.J. Strategy for a protein purification design using C-phycocyanin extract. Bioresour. Technol. 2009, 100, 5312–5317. [Google Scholar] [CrossRef]
- Kumar, D.; Dhar, D.W.; Pabbi, S.; Kumar, N.; Walia, S. Extraction and purification of C-phycocyanin from Spirulina platensis (CCC540). Indian J. Plant Physiol. 2014, 19, 184–188. [Google Scholar] [CrossRef]
- Julianti, E.; Susanti, S.; Singgih, M.; Mulyani, L.N. Optimization of Extraction Method and Characterization of Phycocyanin Pigment from Spirulina platensis. J. Math. Fundam. Sci. 2019, 51, 168–176. [Google Scholar] [CrossRef]
- Chew, K.W.; Chia, S.R.; Krishnamoorthy, R.; Tao, Y.; Chu, D.-T.; Show, P.L. Liquid biphasic flotation for the purification of C-phycocyanin from Spirulina platensis microalga. Bioresour. Technol. 2019, 288, 121519. [Google Scholar] [CrossRef] [PubMed]
- Patil, G.; Chethana, S.; Sridevi, A.S.; Raghavarao, K.S.M.S. Method to obtain C-phycocyanin of high purity. J. Chromatogr. A 2006, 1127, 76–81. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Amount | Ref. | Compound | Amount | Ref. |
|---|---|---|---|---|---|
| General composition (in dry weight Spirulina) | Inositol | 6.4 mg | [28] | ||
| Proteins | Folic acid | [22,28] | Pantothenic acid | 10 μg | |
| Carbohydrates | Biotin | Folic acid | 1.0 μg | ||
| Total lipids | 5.0–6.0% | Biotin | 0.5 μg | ||
| Fibers | 3.6–6.0% | Minerals (per 10 g Spirulina) | |||
| Minerals | 7.0–13% | Potassium | 140 mg | [28] | |
| Vitamins (per 10 g Spirulina) | Sodium | 90 mg | |||
| Vitamin A | 23000 IU | [28] | Calcium | 70 mg | |
| Vitamin B1, B2, B3 | 0.4–1.4 mg | Phosphorus | 60 mg | ||
| Vitamin B12, B6 | 20–60 μg | Magnesium | 40 mg | ||
| Vitamin C | 0.8 mg | Iron | 15 mg | ||
| Vitamin D | 1200 IU | Manganese | 0.5 mg | ||
| Vitamin E | 1.0 mg | Zinc | 0.3 mg | ||
| Vitamin K1, K2 | 200 μg | Cooper | 120 μg | ||
| Germanium | 60 μg | [28] | Histidine | 100 mg | [29] |
| Iodine | 55 μg | Tryptophan | 90 mg | ||
| Chrome | 25 μg | Nonessential amino acid (per 10 g Spirulina) | |||
| Selenium | 10 μg | Glutamic acid | 910 mg | [29] | |
| Essential amino acid (per 10 g Spirulina) | Aspartic acid | 610 mg | |||
| Leucine | 540 mg | Alanine | 470 mg | ||
| Valine | 400 mg | Arginine | 430 mg | ||
| Isoleucine | 350 mg | Glycine | 320 mg | ||
| Threonine | 320 mg | Serine | 320 mg | ||
| Lysine | 290 mg | Tyrosine | 300 mg | ||
| Phenylalanine | 280 mg | Proline | 270 mg | ||
| Methionine | 140 mg | Cystine | 60 mg | ||
| Role | Effect | Dose or Concentration (Route of Administration) | Analysis Methodology | Ref. |
|---|---|---|---|---|
| Anti-oxidative | Scavenging of free radicals, lipid peroxidation inhibitor and metal chelator | 1–3 mg/mL | Luminol-enhanced chemiluminescence | [54] |
| 0–0.16 mM | Deoxyribose assay | |||
| 8–20 mg/mL | Inhibition of liver microsomal lipid peroxidation induced by Fe-ascorbic acid | |||
| 50–200 mg/kg (oral) | Glucose oxidase-induced inflammation in vivo | |||
| 62.34 mg/g | DPPH, FRAP and Fe2+—chelating activity | [55] | ||
| 0.125–2.00 mg/mL 0.3125–5.00 mg/mL | ABTS and DPPH | [56] | ||
| 10–100 μg/mL | DPPH | [57] | ||
| Serum antioxidant | 200–1000 mg/kg (oral) | SOD and catalase activity in vivo | [58] | |
| Attenuation of MMPs and ROS | 20–80 μg/mL | MMP-1 and MMP-9 and DCFDA staining | [59] | |
| Attenuation of platelet aggregation by decreasing hydroxyl radicals | 0.5–10 nM | Electron Spin Resonance Spectrometry | [60] | |
| Increase of antioxidant enzymes | 5 μM + 2 Gy radiation | RANDOX kit | [61] | |
| Attenuation of ROS | 10 μM (rat heart perfusion) | Electron paramagnetic resonance spectroscopy | [62] | |
| Attenuation of ROS, MDA and GSH, and maintenance of SOD activity | 100–400 mg/kg (intraperitoneal injection) 31–250 μg/mL | DCFDA staining and histopathologic analysis | [63] | |
| Free radical scavenger | 5–50 mM pre-treatment for 1 h and then co-treatment | DCFDA staining | [64] | |
| Anticancer | Cell cycle arrest in G0/G1, attenuation of proliferation and stimulation of apoptosis | 1–20 μM | Propidium iodide, annexin V-PE, 7-AAD, proliferative, and apoptotic markers | [65] |
| 10–100 μM | MTT assay, cytochrome c, ethidium bromide | [66] | ||
| Alteration of the mitochondrial membrane potential | 10–100 μM | Rhodamine 123 | [67] | |
| Attenuation of MMPs | 5–40 μg/mL | MMP-1, MMP-2, MMP-9, TIMP-1, TIMP-2 | [68] | |
| Attenuation of metastasis | 200 mg/kg (oral) | MMPs, VEGF-A and HIF-1α, activity of MMPs and HIF-1α | [69] | |
| Stimulation of mitochondria-mediated apoptosis | 5–40 μg | Depolarized mitochondria, apoptotic, and proliferative markers | [70] | |
| Drug resistance by preventing the induction of multidrug resistance protein | 1–100 μM | ROS production and COX-2 expression | [71] | |
| Anti-inflammatory | Attenuation of pro-inflammatory mediators and neutrophil infiltration | 30–50 mg/kg | TNF-α, IL-1β, IL-10, nitrite, nitrate, PGE2, COX-2, iNOS, MPO, and NF-kB activity | [72] |
| 0–250 μg/mL | [73] | |||
| Attenuation of lung injury | 50 mg/kg 100–400 mg/kg (intraperitoneal injection) | Lung injury, nitrate/nitrite, pro-inflammatory cytokines in BALF, MPO and NF-kB activity, iNOS, COX-2, lung edema, proapoptotic proteins | [74,75] | |
| Prevention of fibrosis | 10–50 μg/mL 0–200 μg/mL | Nrf2, NQO-1, EMT evaluated through the expression of vimentin, type-1-collagen, fibronectin, α-SMA, N-cadherin, and E-cadherin | [76,77] | |
| Antimicrobial | Decrease the growth of Escherichia coli, Bacillus sp., Staphylococcus aureus, and Salmonella Typhi | 35 μg/mL | Disc diffusion assay and determination of MIC. Comparation with Antibiotic Assay Medium (Himedia). | [78] |
| Attenuate the growth of Listeria monocytogenes, S. aureus, Yersinia ruckeri, E. coli, and Streptococcus iniae | 25 μg/mL | Agar well diffusion assay, MIC and MBC. Comparation with Tetracycline, Amikacin, and Doxycycline | [55] | |
| Impair the growth of S. aureus, Aeromonas hydrofila, and Salmonella Enteritidis. No effect in Enterococcus faecalis | 320 μg/mL | Agar well diffusion method and turbidity liquid media assay. MIC and turbidity at 600 nm | [79] | |
| Antibacterial activity against Pseudomonas aeruginosa MTCC 1034, Klebsiella pneumoniae (ESBL-KP) ATCC 700603, E. coli (ATCC 25922), and S. aureus ATCC 25,923 (MRSA). No effect on Acinetobacter baumanii, Enterococcus durans (P502). | 1000 μg/mL | Mueller–Hinton Agar plates and MIC using broth microdilution method | [80] | |
| Attenuation of acne symptoms and reduction of Propionibacterium acnes and Staphylococcus epidermidis | 10% extract | Disc diffusion method and MIC | [81] | |
| Inhibition of the growth of Candida albicans, Aspergillus niger, Aspergillus flavus, Penicillium sp., and Rhizopus sp. | 40–80 μg/mL | Agar block method and MIC | [82] | |
| Anti-neurodegenerative | Promotor of remyelination | 25 mg/kg (intraperitoneal injection) | Brain biopsies, pro-inflammatory mediators and populations, lipid peroxidation | [83] |
| Attenuation of Alzheimer’s disease markers | 0–20 μg/mL | Intracellular GSH, APP, BACE2, GSH-Px, SOD2, GR, BDNF, α-tubulin | [84] | |
| 50 or 100 mg/kg (intraperitoneal injection) | Morris water maze, novel object recognition and open field test, ChAT, inflammatory and apoptotic mediators, IRS-1, INS, PI3K/AKT, and PTEN gene expression | [85] | ||
| 200 mg/kg (intraperitoneal injection) | Eight-arm radial maze, HAC3, pro-inflammatory, and proapoptic mediators | [86] | ||
| Attenuation of Parkinson’s disease markers | 2.5–7.5 μM | Fibril formation of αS or Aβ40/42, ADH, catalase | [87] | |
| Antidiabetic | Antidiabetic and antiglycation | 100–500 μg/mL | Inhibitory effect of PPA and β-glucosidase | [88] |
| 100 and 200 mg/kg 100 mg/kg 200 mg/kg 50 mg/kg (oral) | Blood glucose, glycosylated hemoglobin HbA1c, BUN, urea, serum creatinine, SGOT/AST, SGPT/ALT, alkaline phosphatase, total bilirubin, TGs, LDL-C, TC, and HDL-C NBT assay, carbonyl content, reduced GSH | [89,90,91,92] | ||
| Hepatoprotection | Attenuation of nephrotoxicity | Not described | Plasma urea, creatinine, urinary N-acetyl-β-D-glucosaminidase, creatinine and lithium, histomorphology evaluation | [93] |
| Reduction of hepatocyte damage | 50–200 mg/kg (intraperitoneal injection) | Hepatic lipid peroxidation assayed by measuring malondialdehyde | [94] | |
| 1–100 μgM | ROS, MDA, GSH, GSH-Px, ALT, AST, SOD, TGF-β1, HGF | [71] | ||
| Nephroprotection | Prevention of cisplatin induced nephrotoxicity by reducing oxidative stress | 5–30 mg/kg (intraperitoneal injection) | Blood urea nitrogen, plasma glutathione peroxidase, plasma creatinine quantification, N-acetyl-β-D-glucosaminidase, apoptosis and histopathological changes; glutathione, malondialdehyde, 4-hydroxynonenal, and oxidized proteins quantification | [95] |
| Protection of Type 2 diabetes mice against oxidative stress and renal dysfunction | 300 mg/kg (oral) | Urinary 8-hydroxy-2-deoxyguanosine, 8-iso-prostaglandin F2α and albumin quantification; immunohistochemistry | [96] | |
| Recovery of cisplatin-induced renal injury in renal tissue and HK-2 cell and reduction of p-ERK, p-JNK, p-p38, Bax, caspase-9, and caspase-3 | 50 mg/kg (intraperitoneal injection) | Light microscopy examination, cell viability Assay, western blot, caspase-3 activity assay, and apoptosis detection by the terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling method | [97] | |
| Cardiovascular protection | Reduction of atherosclerotic disease | 200 μM | HMOX-1, eNOS, P22, VCAM-1 | [98] |
| Prevention of AMI-induced oxidative stress, inflammation and heart damage | 50 mg/kg (subcutaneous injection) | CK, AST, ALT, ROS, nitrites, oxidized glutathione, pro-inflammatory and proapoptotic cytokines, lipid peroxidation | [99] | |
| Prevention of cardiovascular diseases and atherosclerotic formation | 0.25% and 1.25% (oral) | Cholesterol, MDA, GOT, GPT, catalase, SOD, GSH-Px, HMG CoA | [100] | |
| Anti-obesity | Prevention of endothelial dysfunction and attenuation of obesity | 2500, 5000, or 10,000 mg/kg (oral) | Serum triglyceride, total cholesterol, HDL-C, and glucose, insulin and leptin, immunohistochemistry analysis | [101] |
| Reduction of adipogenesis and lipogenesis | 0, 0.625, 1.25, 2.5, 5, 10, or 20 μg/mL (oral) | Western blots of adipogenic proteins (C/EBPα, PPARγ, and aP2) and lipogenic proteins (SREBP1, ACC, FAS, LPAATβ, Lipin1, and DGAT1) | [102] | |
| Wound healing | Proliferation of fibroblasts, synthesis of ECM components and regeneration | 10–200 μg/mL (superficial collagen films) 50 μg/mL | Cytotoxicity and proliferation/viability of fibroblasts, cdK1, cdK2, uPA, PI3K, and in vivo wound healing analysis | [103,104] |
| Antimelanogenic | Attenuation of melanin production | Not described | Cellular tyrosinase, production of melanin, DPPH | [105] |
| 0.05–2.00 mg/mL | Tyrosinase activity, melanin, intracellular cAMP, MITF, tyrosinase, ERK, pERK1/2, MEK1/2, p38, CREB | [106] |
| Cell Disruption Method | Purity * | Yield (mg/g) | Conditions | Advantages | Limitations | Ref. |
|---|---|---|---|---|---|---|
| Freeze/thaw cycles | 0.56–0.66- | 73.73–74.51 | 6 cycles, −40 °C/4 h + room temperature/1 h, 0.1 M phosphate buffer pH (6.8), 1:6, 1:8 and 1:10 S/L ratios | Continuous damage to the plasmatic membrane, easy to perform, availability | Time and energy consuming, and often require high solvent, leading to an increase in the production costs. Not suitable for industrial scale | [141] |
| 0.4 | ND | 25 °C/4 h, distilled water, 1:25 S/L ratio | [142] | |||
| 0.77 | 217.18 | 3 cycles, −20 °C + room temperature/24 h, 20 mM of sodium acetate and 50 mM of NaCl buffer (pH 5.1) 1:20 S/L ratio | [143] | |||
| 2.10 | 41.90 | 4 cycles, −20 °C/4 h + room temperature/1.5 h, sodium hydroxide, (pH 6.8), 1:20 S/L ratio | [144] | |||
| Mixing and homogenization | 0.6 | 52.11 | 25,200× g/10 min, 0.1 M phosphate buffer (pH 6.8), 1:6 S/L ratio | Simple, availability, reproducibility | Increased temperatures during the process, time consuming, not suitable for industrial scale, cell debris released | [141] |
| ND | ND | Rotary shaker at 30 °C, 10 mM sodium phosphate buffer (pH 7.0), 10 mM sodium acetate buffer (pH 5.0), NaCl 0.15 M and CaCl2 10 g/L, 1:25 S/L ratio | [142] | |||
| ND | 67.61 | 1.71% biomass/solvent ratio, 6237.66 homogenization rate, 15 min extraction time | [145] | |||
| 0.67 | 103.07 | Oven-dried biomass preparation, 70 °C/4 h, extraction at 25 °C/24 h by 0.01 M phosphate buffer using homogenization assisted method at 0.02 g/mL biomass concentration | [146] | |||
| Bead milling | 0.46 | 217.14 | Low-density beads for low viscosity media, 80–85% degree of bead filling | Low time, high biomass disruption, low energy | Time very dependent of the type of bead, low purities, cell debris, additional purification steps needed | [144] |
| ND | 119.48 mg/g | Bead diameter 0.3 mm, glass beads at a speed of 3580 rpm. 4 cycles of milling each 25 s and subsequent cooling at 4 °C | [147] | |||
| ND | 90% recovery | Dakot Zirconia beads (0.5–1.4 mm), 330 rpm agitation, 8 h under 0.5 M Ca (II) in a 0.35 M acetate buffer (pH 6.8) | [148] | |||
| 0.21 | ND | Bead Beater, diameter 0.1 mm, agitation of 4800 rpm, 10 cycles of 10 s. Following each cycle, samples cool down in water at 0 °C | [149] | |||
| ND | 94.90 | Glass beads 0.25–0.5 mm of diameter) in 2 mL flasks, 4 cycles of 25 s at 30 Hz of vibrational frequency | [150] | |||
| Ultrasound | 0.62 | 51.51 | Pre-soaked for 120 min, ultrasonication amplitude of 50%, 2.5 min, 1:6 S/L | High purities, reproducibility, suitable for industrial scale, temperature could be controlled | Increased temperatures during the process, complex process, expensive, specific equipment | [141] |
| ND | 98.84 | 1% biomass/solvent ratio, 60% amplitude, 16.23 min extraction time | [145] | |||
| 0.67–0.93 | 90.00 | Frequencies of 20–100 kHz, power intensities >1 W/cm2, PBS soaking | [146] | |||
| 0.65 | 18.20 | Power 60 W, extraction for 10 s, 30 cycles in total, in ice bath | [151] | |||
| Electric field | 2.45 | 143.33 | Freeze/thaw and pulsed electric field maximum charging voltage of 30 kV, square bipolar pulses with a variable pulse width of 4–32 μs and a pulse frequency up to 300 Hz. | Increased permeability of the membrane | Long time to optimization, complex equipment, intracellular compounds might not be completely released | [141] |
| 0.51 | 151.94 | 40 °C, 25 kV/cm, 150 μs | [149] | |||
| ND | ND | 50 to 200 pulses at 20 kV | [152] | |||
| High-pressure homogenization | ND | ND | 3.5 min with pressures between 50 and 600 MPa, distilled water ratio of 6% (wt%) | Simpler, scalable for industrial application, environmentally friendly, high recovery | Expensive, not useful in extracting dry biomass, can lead to protein denaturation | [152] |
| ND | 291.90 | 100 mM Na-phosphate solvent (pH 7), 1400 bar | [153] | |||
| 1.2–1.4 | 90% recovery | 300 MPa for 10 min, deionized water or phosphate buffer (pH 6.8), 1/20 (w/v) ratio | [154] | |||
| Enzyme-assisted | 0.80–0.90 | 20–25 | 1 mg/mL lysozyme, high pressure homogenizer D-15M at 10–12,000 p.s.i., 4–8 °C | Stable, efficient, eco-friendly | More efficient when combined with other methods | [153] |
| 1.19 | 82.07 | 1.0% enzyme concentration, 16 h incubation time, 1:6 S/L ratio | [155] | |||
| 1.09 | 92.73 | 2.5 min Ultrasonication at 50% Amplitude, 0.6% enzyme concentration, 16 h incubation, 1:6 S/L ratio |
| Purification Method | Conditions | Purity * | Recovery (%) | Ref. |
|---|---|---|---|---|
| Ammonium sulfate (AS) precipitation | 50–65% AS | 2.11 | 86 | [197] |
| Chitosan and activated charcoal | 0.24% chitosan/8.4% activated charcoal | 3.14 | 79 | [198] |
| Stirred fluidized bed IEC | 1%, dw/v in STREAMLINE DEAE 10%, dw/v in STREAMLINE DEAE | 2.70 3.00 | 90 64 | [199] |
| IEC | IEC with pH gradient using an anion-exchanger Q-Sepharose Fast Flow column | 4.20 | 49 | [200] |
| Combined methods | 50–65% AS, dialysis in sodium acetate buffer, IEC on a DEAE-Sepharose Fast Flow column | 5.59 | 67 | [197] |
| 50% AS, dialysis, ultrafiltration with MWCO of 50 kDa and IEC (anion-exchanger resin Q-Sepharose Fast Flow column) with pH gradient | 4.00 | 80 | [44] | |
| 65% AS, 65% dialysis with 12–14 kDa membranes, microfiltration, IEC in a Sephadex-G-100 column, and HPLC with a reverse column | 92% | 53 | [88] | |
| 1.113 M AS, filtration with a PVDF membrane and two hydrophobic interaction membrane chromatography steps | 4.20 | 67 | [201] | |
| 2% w/v chitosan solution, 80 g/L activated charcoal, ultrafiltration, and IEC on DEAE Sephadex A-25 | 4.30 | 42 | [151] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, R.; Campos, J.; Serra, M.; Fidalgo, J.; Almeida, H.; Casas, A.; Toubarro, D.; Barros, A.I.R.N.A. Exploring the Benefits of Phycocyanin: From Spirulina Cultivation to Its Widespread Applications. Pharmaceuticals 2023, 16, 592. https://doi.org/10.3390/ph16040592
Fernandes R, Campos J, Serra M, Fidalgo J, Almeida H, Casas A, Toubarro D, Barros AIRNA. Exploring the Benefits of Phycocyanin: From Spirulina Cultivation to Its Widespread Applications. Pharmaceuticals. 2023; 16(4):592. https://doi.org/10.3390/ph16040592
Chicago/Turabian StyleFernandes, Raquel, Joana Campos, Mónica Serra, Javier Fidalgo, Hugo Almeida, Ana Casas, Duarte Toubarro, and Ana I. R. N. A. Barros. 2023. "Exploring the Benefits of Phycocyanin: From Spirulina Cultivation to Its Widespread Applications" Pharmaceuticals 16, no. 4: 592. https://doi.org/10.3390/ph16040592
APA StyleFernandes, R., Campos, J., Serra, M., Fidalgo, J., Almeida, H., Casas, A., Toubarro, D., & Barros, A. I. R. N. A. (2023). Exploring the Benefits of Phycocyanin: From Spirulina Cultivation to Its Widespread Applications. Pharmaceuticals, 16(4), 592. https://doi.org/10.3390/ph16040592