
Antimetastatic Activity of Apoptolidin A by Upregulation of N-Myc Downstream-Regulated Gene 1 Expression in Human Colorectal Cancer Cells

 ,
,
Abstract
1. Introduction
2. Results
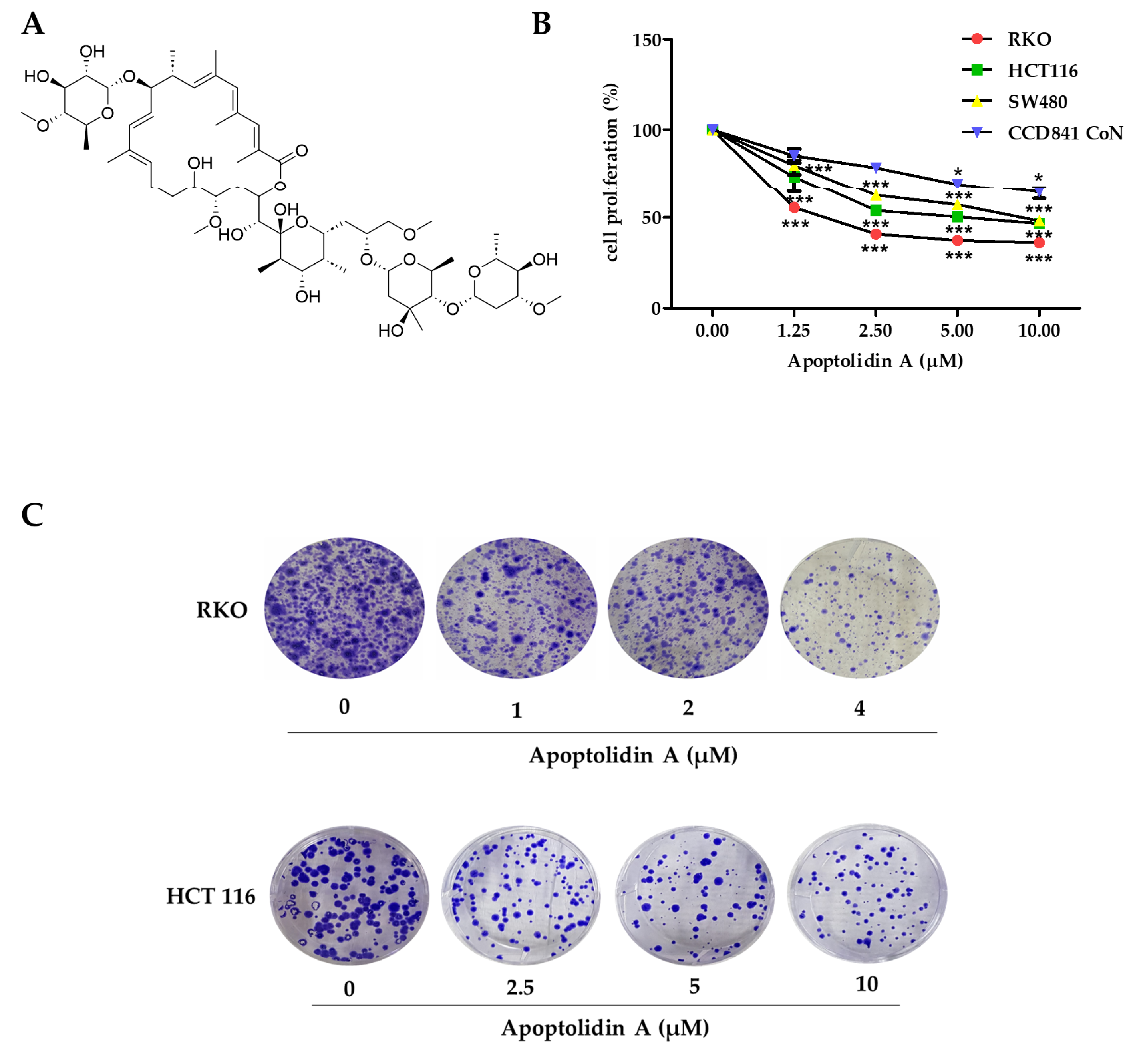
2.1. Growth Inhibition Activity of Apoptolidin A in Human Colorectal Cancer (CRC) Cells
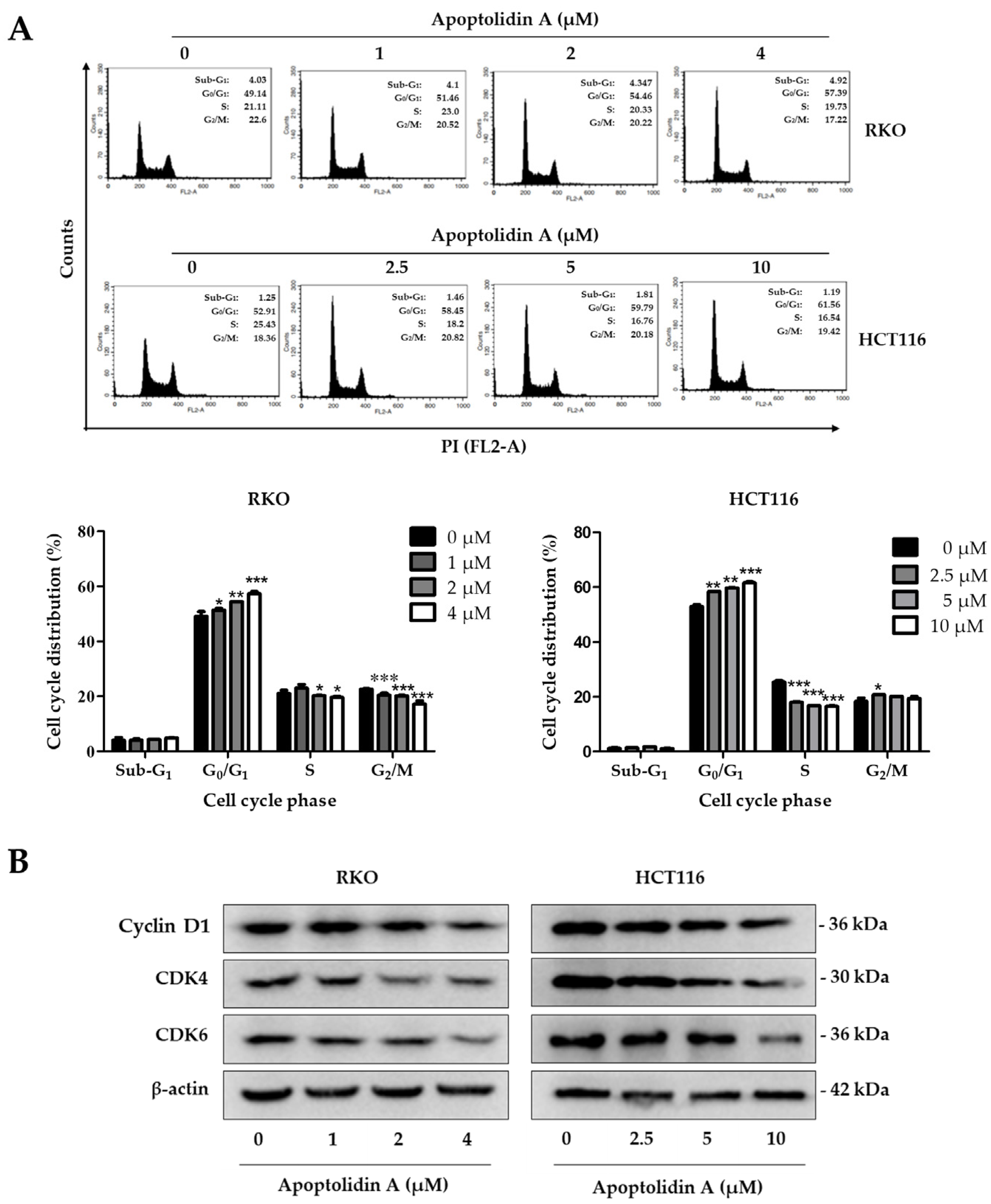
2.2. Effect of Apoptolidin A on Induction of Cell Cycle Arrest in CRC Cells
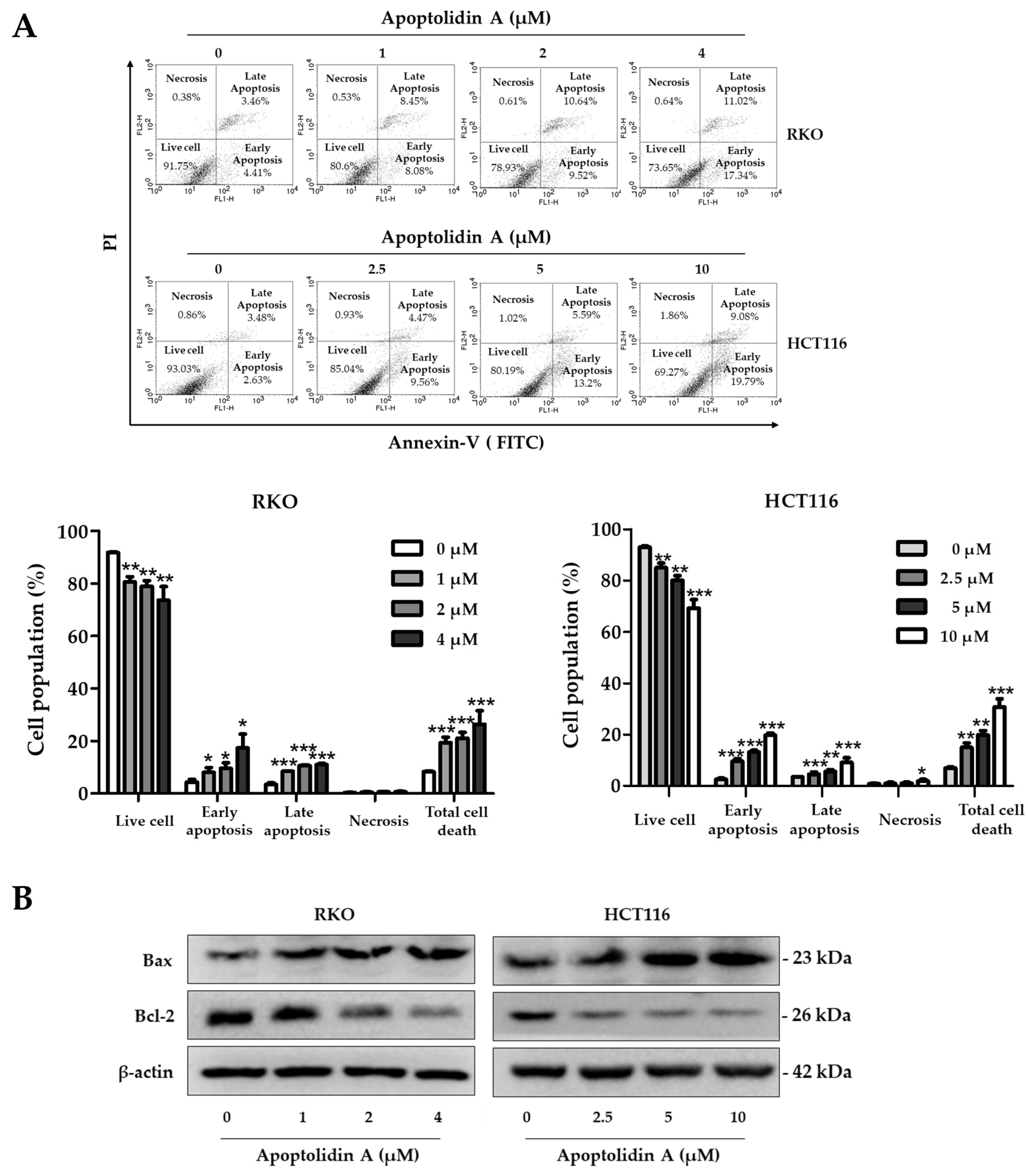
2.3. Effect of Apoptolidin A on Induction of Apoptosis in CRC Cells
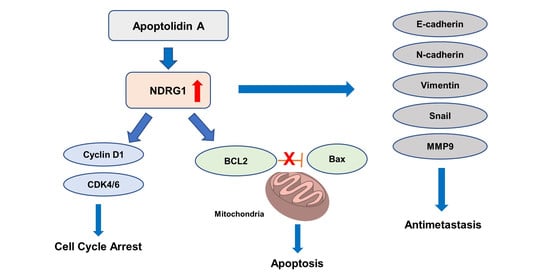
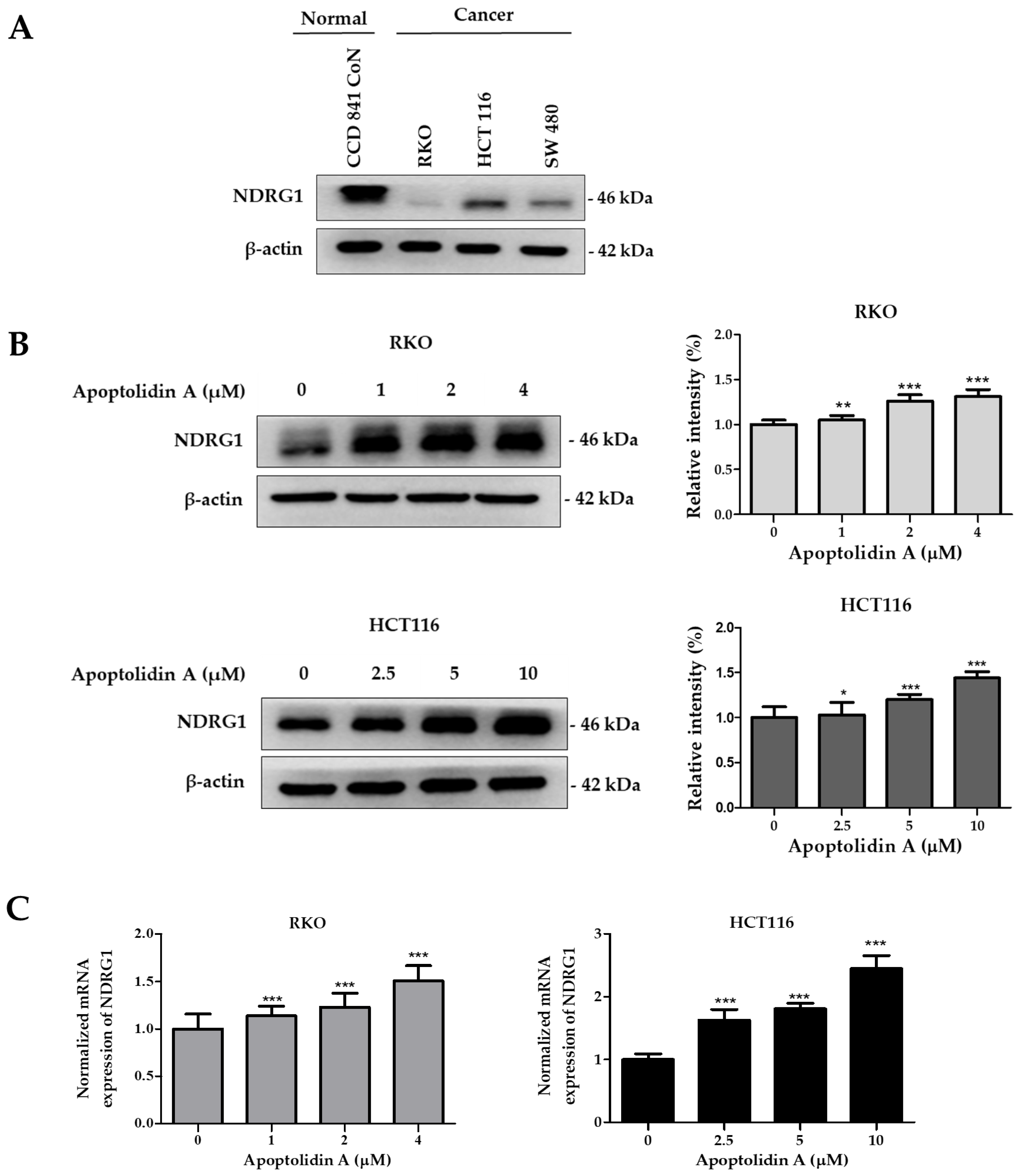
2.4. Effect of Apoptolidin A on Regulation of NDRG1 Expression in CRC Cells
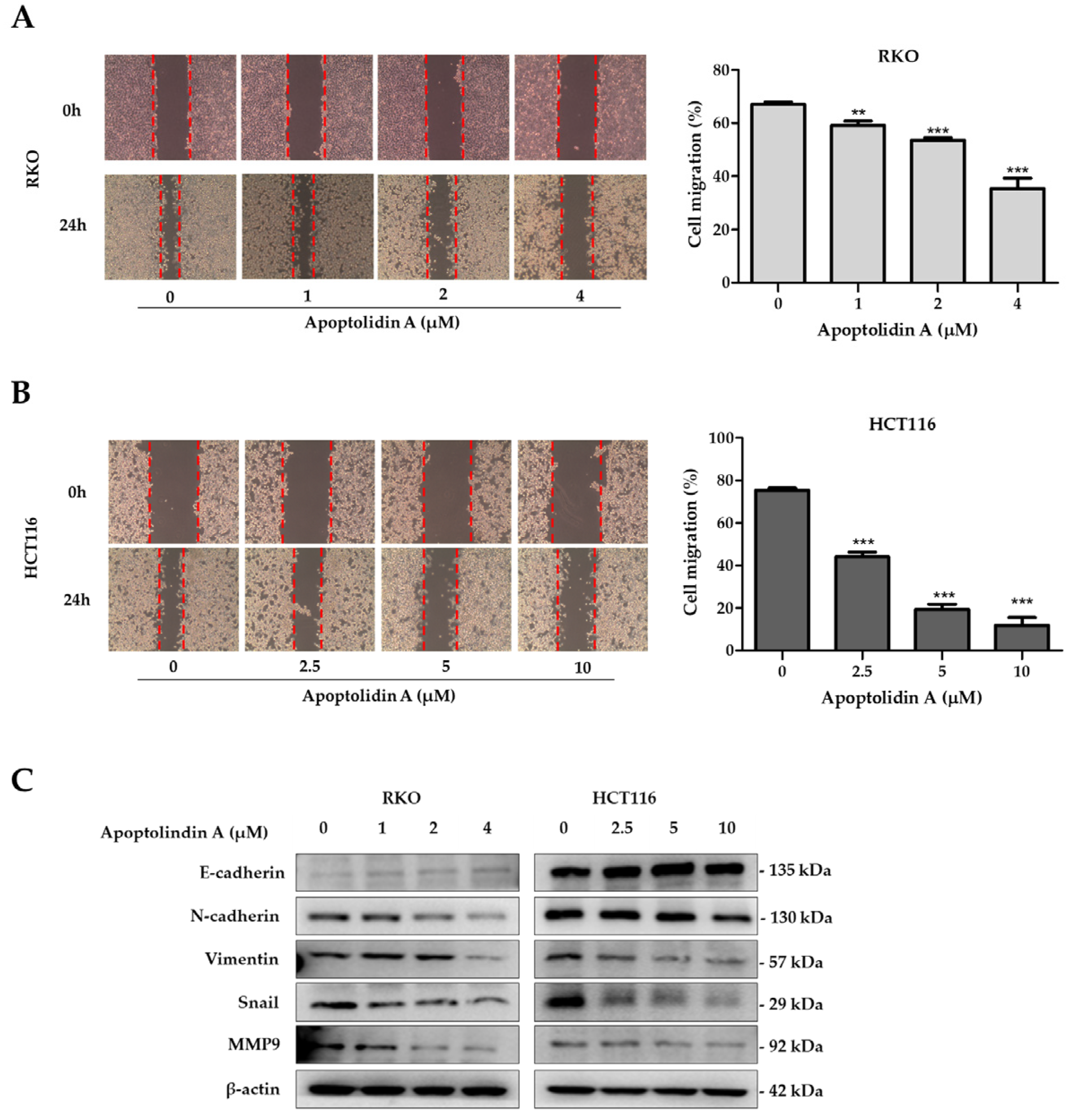
2.5. Effect of Apoptolidin A on Migration of CRC Cells
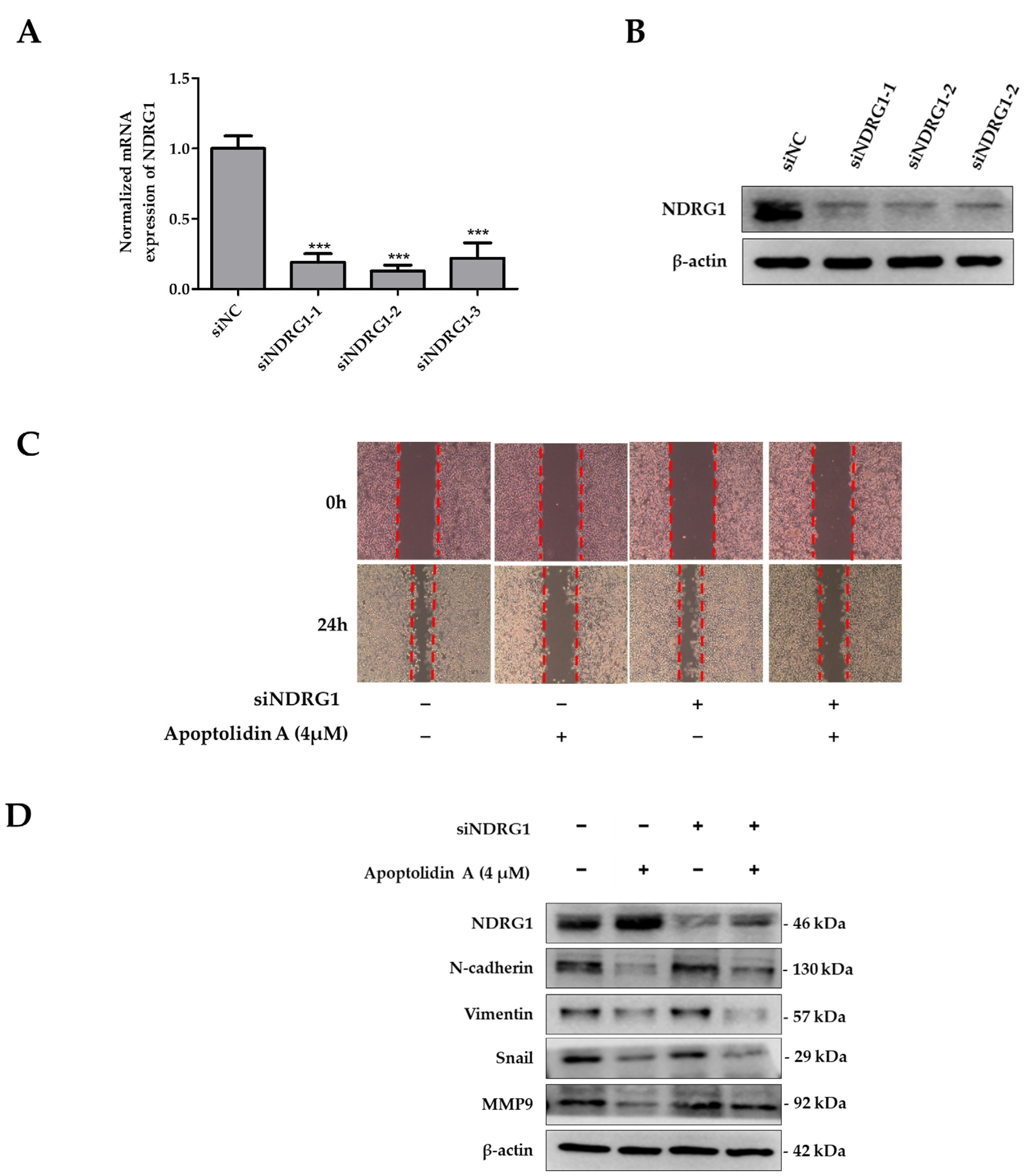
2.6. Effects of siNDRG1 on Cell Migration in Apoptolidin A-Treated CRC Cells
3. Discussion
4. Materials and Methods
4.1. General Reagents
4.2. Cell Culture
4.3. Cell Proliferation Assay
4.4. Gene Silencing by siRNA Transfection
4.5. Colony Formation
4.6. Immunoblotting
4.7. Cell Cycle Analysis
4.8. Apoptosis Analysis with Annexin V-Fluorescein Isothiocyanate (V-FITC)/PI Double Staining by Flow Cytometry
4.9. Wound Healing Assay
4.10. RNA Extraction and Real-Time PCR
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Weinberg, R.A. A Perspective on Cancer Cell Metastasis. Science 2011, 331, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, J.B.; Maggard, M.A.; Ko, C.Y. Colon cancer survival rates with the new American Joint Committee on Cancer sixth edition staging. J. Natl. Cancer Inst. 2004, 96, 1420–1425. [Google Scholar] [CrossRef]
- Ansa, B.E.; Coughlin, S.S.; Alema-Mensah, E.; Smith, S.A. Evaluation of Colorectal Cancer Incidence Trends in the United States (2000–2014). J. Clin. Med. 2018, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Berber, E. Role of thermal ablation in the management of colorectal liver metastasis. Hepatobiliary Surg. Nutr. 2020, 9, 49–58. [Google Scholar] [CrossRef]
- Piawah, S.; Venook, A.P. Targeted therapy for colorectal cancer metastases: A review of current methods of molecularly targeted therapy and the use of tumor biomarkers in the treatment of metastatic colorectal cancer. Cancer 2019, 125, 4139–4147. [Google Scholar] [CrossRef]
- Johdi, N.A.; Sukor, N.F. Colorectal Cancer Immunotherapy: Options and Strategies. Front. Immunol. 2020, 11, 1624. [Google Scholar] [CrossRef]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef]
- Busch, E.L.; McGraw, K.A.; Sandler, R.S. The potential for markers of epithelial-mesenchymal transition to improve colorectal cancer outcomes: A systematic review. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1164–1175. [Google Scholar] [CrossRef]
- Cao, H.; Xu, E.; Liu, H.; Wan, L.; Lai, M. Epithelial-mesenchymal transition in colorectal cancer metastasis: A system review. Pathol. Res. Pract. 2015, 211, 557–569. [Google Scholar] [CrossRef]
- Bates, R.C.; Mercurio, A.M. The epithelial-mesenchymal transition (EMT) and colorectal cancer progression. Cancer Biol. Ther. 2005, 4, 365–370. [Google Scholar] [CrossRef] [PubMed]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Dedhar, S.; Kalluri, R.; Thompson, E.W. The epithelial-mesenchymal transition: New insights in signaling, development, and disease. J. Cell Biol. 2006, 172, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Spaderna, S.; Schmalhofer, O.; Hlubek, F.; Berx, G.; Eger, A.; Merkel, S.; Jung, A.; Kirchner, T.; Brabletz, T. A transient, EMT-linked loss of basement membranes indicates metastasis and poor survival in colorectal cancer. Gastroenterology 2006, 131, 830–840. [Google Scholar] [CrossRef]
- Voon, D.C.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. The EMT spectrum and therapeutic opportunities. Mol. Oncol. 2017, 11, 878–891. [Google Scholar] [CrossRef]
- Kohrmann, A.; Kammerer, U.; Kapp, M.; Dietl, J.; Anacker, J. Expression of matrix metalloproteinases (MMPs) in primary human breast cancer and breast cancer cell lines: New findings and review of the literature. BMC Cancer 2009, 9, 188. [Google Scholar] [CrossRef]
- Brown, G.T.; Murray, G.I. Current mechanistic insights into the roles of matrix metalloproteinases in tumour invasion and metastasis. J. Pathol. 2015, 237, 273–281. [Google Scholar] [CrossRef]
- Chang, X.; Xu, X.; Xue, X.; Ma, J.; Li, Z.; Deng, P.; Chen, J.; Zhang, S.; Zhi, Y.; Dai, D. NDRG1 Controls Gastric Cancer Migration and Invasion through Regulating MMP-9. Pathol. Oncol. Res. 2016, 22, 789–796. [Google Scholar] [CrossRef]
- Hu, Z.Y.; Xie, W.B.; Yang, F.; Xiao, L.W.; Wang, X.Y.; Chen, S.Y.; Li, Z.G. NDRG1 attenuates epithelial-mesenchymal transition of nasopharyngeal cancer cells via blocking Smad2 signaling. Biochim. Biophys. Acta 2015, 1852, 1876–1886. [Google Scholar] [CrossRef]
- Mi, L.; Zhu, F.; Yang, X.; Lu, J.; Zheng, Y.; Zhao, Q.; Wen, X.; Lu, A.; Wang, M.; Zheng, M.; et al. The metastatic suppressor NDRG1 inhibits EMT, migration and invasion through interaction and promotion of caveolin-1 ubiquitylation in human colorectal cancer cells. Oncogene 2017, 36, 4323–4335. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.Y.; Chun, Y.S.; Shin, H.W.; Park, J.W. Potential role of the N-MYC downstream-regulated gene family in reprogramming cancer metabolism under hypoxia. Oncotarget 2016, 7, 57442–57451. [Google Scholar] [CrossRef]
- Wang, J.; Cai, J.; Li, Z.; Hu, S.; Yu, L.; Xiao, L.; Wang, Z. Expression and biological function of N-myc down-regulated gene 1 in human cervical cancer. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 30, 771–776. [Google Scholar] [CrossRef]
- Song, Y.; Lv, L.; Du, J.; Yue, L.; Cao, L. Correlation of N-myc downstream-regulated gene 1 subcellular localization and lymph node metastases of colorectal neoplasms. Biochem. Biophys. Res. Commun. 2013, 439, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Liu, W.; Menezes, S.; Yue, F.; Zheng, M.; Kovacevic, Z.; Richardson, D.R. The metastasis suppressor NDRG1 modulates the phosphorylation and nuclear translocation of beta-catenin through mechanisms involving FRAT1 and PAK4. J. Cell Sci. 2014, 127, 3116–3130. [Google Scholar]
- Wang, B.; Li, J.; Ye, Z.; Li, Z.; Wu, X. N-myc downstream regulated gene 1 acts as a tumor suppressor in ovarian cancer. Oncol. Rep. 2014, 31, 2279–2285. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Pai, S.K.; Gross, S.C.; Hirota, S.; Hosobe, S.; Miura, K.; Saito, K.; Commes, T.; Hayashi, S.; Watabe, M.; et al. The Drg-1 gene suppresses tumor metastasis in prostate cancer. Cancer Res. 2003, 63, 1731–1736. [Google Scholar] [PubMed]
- Li, Q.; Chen, H. Transcriptional silencing of N-Myc downstream-regulated gene 1 (NDRG1) in metastatic colon cancer cell line SW620. Clin. Exp. Metastasis 2011, 28, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Iiizumi-Gairani, M.; Okuda, H.; Kobayashi, A.; Watabe, M.; Pai, S.K.; Pandey, P.R.; Xing, F.; Fukuda, K.; Modur, V.; et al. KAI1 gene is engaged in NDRG1 gene-mediated metastasis suppression through the ATF3-NFkappaB complex in human prostate cancer. J. Biol. Chem. 2011, 286, 18949–18959. [Google Scholar] [CrossRef]
- Zhao, G.; Chen, J.; Deng, Y.; Gao, F.; Zhu, J.; Feng, Z.; Lv, X.; Zhao, Z. Identification of NDRG1-regulated genes associated with invasive potential in cervical and ovarian cancer cells. Biochem. Biophys. Res. Commun. 2011, 408, 154–159. [Google Scholar] [CrossRef]
- Shah, M.A.; Kemeny, N.; Hummer, A.; Drobnjak, M.; Motwani, M.; Cordon-Cardo, C.; Gonen, M.; Schwartz, G.K. Drg1 expression in 131 colorectal liver metastases: Correlation with clinical variables and patient outcomes. Clin. Cancer Res. 2005, 11, 3296–3302. [Google Scholar] [CrossRef]
- Strzelczyk, B.; Szulc, A.; Rzepko, R.; Kitowska, A.; Skokowski, J.; Szutowicz, A.; Pawelczyk, T. Identification of high-risk stage II colorectal tumors by combined analysis of the NDRG1 gene expression and the depth of tumor invasion. Ann. Surg. Oncol. 2009, 16, 1287–1294. [Google Scholar] [CrossRef]
- Kim, J.W.; Adachi, H.; Shin-ya, K.; Hayakawa, Y.; Seto, H. Apoptolidin, a new apoptosis inducer in transformed cells from Nocardiopsis sp. J. Antibiot. 1997, 50, 628–630. [Google Scholar] [CrossRef]
- Oh, S.H.; Park, J.; Moon, D.H.; Hyun, G.H.; Jang, Y.-J.; Hwang, S.; An, J.S.; Du, Y.E.; Kwon, Y.; Kang, Y.P.; et al. Apoptolidin I: A new glycosylated macrocyclic lactone from wasp gut bacterium, Amycolatopsis sp. Tetrahedron 2022, 127, 133091. [Google Scholar] [CrossRef]
- Serrill, J.D.; Tan, M.; Fotso, S.; Sikorska, J.; Kasanah, N.; Hau, A.M.; McPhail, K.L.; Santosa, D.A.; Zabriskie, T.M.; Mahmud, T.; et al. Apoptolidins A and C activate AMPK in metabolically sensitive cell types and are mechanistically distinct from oligomycin A. Biochem. Pharmacol. 2015, 93, 251–265. [Google Scholar] [CrossRef]
- Wender, P.A.; Jankowski, O.D.; Longcore, K.; Tabet, E.A.; Seto, H.; Tomikawa, T. Correlation of F0F1-ATPase Inhibition and Antiproliferative Activity of Apoptolidin Analogues. Org. Lett. 2006, 8, 589–592. [Google Scholar] [CrossRef] [PubMed]
- Pucci, B.; Kasten, M.; Giordano, A. Cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Prenek, L.; Boldizsar, F.; Kugyelka, R.; Ugor, E.; Berta, G.; Nemeth, P.; Berki, T. The regulation of the mitochondrial apoptotic pathway by glucocorticoid receptor in collaboration with Bcl-2 family proteins in developing T cells. Apoptosis 2017, 22, 239–253. [Google Scholar] [CrossRef]
- Kovacevic, Z.; Richardson, D.R. The metastasis suppressor, Ndrg-1: A new ally in the fight against cancer. Carcinogenesis 2006, 27, 2355–2366. [Google Scholar] [CrossRef]
- Hood, J.D.; Cheresh, D.A. Role of integrins in cell invasion and migration. Nat. Rev. Cancer 2002, 2, 91–100. [Google Scholar] [CrossRef]
- Banyard, J.; Bielenberg, D.R. The role of EMT and MET in cancer dissemination. Connect. Tissue Res. 2015, 56, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Celia-Terrassa, T.; Bastian, C.; Liu, D.D.; Ell, B.; Aiello, N.M.; Wei, Y.; Zamalloa, J.; Blanco, A.M.; Hang, X.; Kunisky, D.; et al. Hysteresis control of epithelial-mesenchymal transition dynamics conveys a distinct program with enhanced metastatic ability. Nat. Commun. 2018, 9, 5005. [Google Scholar] [CrossRef]
- Fidler, I.J.; Kripke, M.L. The challenge of targeting metastasis. Cancer Metastasis Rev. 2015, 34, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Gerhards, N.M.; Rottenberg, S. New tools for old drugs: Functional genetic screens to optimize current chemotherapy. Drug Resist. Updates 2018, 36, 30–46. [Google Scholar] [CrossRef]
- Wu, X.; Song, M.; Qiu, P.; Li, F.; Wang, M.; Zheng, J.; Wang, Q.; Xu, F.; Xiao, H. A metabolite of nobiletin, 4′-demethylnobiletin and atorvastatin synergistically inhibits human colon cancer cell growth by inducing G0/G1 cell cycle arrest and apoptosis. Food Funct. 2018, 9, 87–95. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, X.; Alt, E.; Bai, X.; Huang, S.; Xu, Z.; Lynch, P.M.; Moyer, M.P.; Wen, X.F.; Wu, X. Chemoprevention of colorectal cancer by targeting APC-deficient cells for apoptosis. Nature 2010, 464, 1058–1061. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lin, H.; Wang, D.; Li, Q.; Luo, H.; Li, G.; Chen, X.; Li, Y.; Chen, P.; Zhai, B.; et al. PCDH17 increases the sensitivity of colorectal cancer to 5-fluorouracil treatment by inducing apoptosis and autophagic cell death. Signal Transduct. Target Ther. 2019, 4, 53. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Leibowitz, B.; Yu, J. Mitochondrial signaling in cell death via the Bcl-2 family. Cancer Biol. Ther. 2010, 9, 417–422. [Google Scholar] [CrossRef]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran Biomed. J. 2015, 19, 69–75. [Google Scholar]
- Li, Y.; Pan, P.; Qiao, P.; Liu, R. Downregulation of N-myc downstream regulated gene 1 caused by the methylation of CpG islands of NDRG1 promoter promotes proliferation and invasion of prostate cancer cells. Int. J. Oncol. 2015, 47, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Li, E.Y.; Huang, W.Y.; Chang, Y.C.; Tsai, M.H.; Chuang, E.Y.; Kuok, Q.Y.; Bai, S.T.; Chao, L.Y.; Sher, Y.P.; Lai, L.C. Aryl Hydrocarbon Receptor Activates NDRG1 Transcription under Hypoxia in Breast Cancer Cells. Sci. Rep. 2016, 6, 20808. [Google Scholar] [CrossRef]
- Tian, L.; Shen, D.; Li, X.; Shan, X.; Wang, X.; Yan, Q.; Liu, J. Ginsenoside Rg3 inhibits epithelial-mesenchymal transition (EMT) and invasion of lung cancer by down-regulating FUT4. Oncotarget 2016, 7, 1619–1632. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.H.; Shim, H.S.; Rha, S.Y.; Kang, S.H.; Hong, S.H.; Choi, Y.D.; Hong, S.J.; Cho, S.H. Increased expression of matrix metalloproteinase 9 correlates with poor prognostic variables in renal cell carcinoma. Eur. Urol. 2003, 44, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.L.; Chu, Y.L.; Ho, C.T.; Chung, J.G.; Lai, C.I.; Su, Y.C.; Kuo, Y.H.; Sheen, L.Y. Antcin K, an Active Triterpenoid from the Fruiting Bodies of Basswood-Cultivated Antrodia cinnamomea, Inhibits Metastasis via Suppression of Integrin-Mediated Adhesion, Migration, and Invasion in Human Hepatoma Cells. J. Agric. Food Chem. 2015, 63, 4561–4569. [Google Scholar] [CrossRef]
- Fu, Y.; Wei, J.; Dai, X.; Ye, Y. Increased NDRG1 expression attenuate trophoblast invasion through ERK/MMP-9 pathway in preeclampsia. Placenta 2017, 51, 76–81. [Google Scholar] [CrossRef]
- Kyaw, K.Z.; Byun, W.S.; Shin, Y.H.; Huynh, T.H.; Lee, J.Y.; Bae, E.S.; Park, H.J.; Oh, D.C.; Lee, S.K. Antitumor Activity of Piceamycin by Upregulation of N-Myc Downstream-Regulated Gene 1 in Human Colorectal Cancer Cells. J. Nat. Prod. 2022, 85, 2817–2827. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | Apoptolidin A (IC50, μM) | Etoposide 1 (IC50, μM) |
|---|---|---|
| RKO HCT116 | 1.45 4.30 | 1.90 0.78 |
| SW480 CCD841 CoN | 8.57 >10 | 0.90 7.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyaw, K.Z.; Park, J.; Oh, S.H.; Lee, J.Y.; Bae, E.S.; Park, H.J.; Oh, D.-C.; Lee, S.K. Antimetastatic Activity of Apoptolidin A by Upregulation of N-Myc Downstream-Regulated Gene 1 Expression in Human Colorectal Cancer Cells. Pharmaceuticals 2023, 16, 491. https://doi.org/10.3390/ph16040491
Kyaw KZ, Park J, Oh SH, Lee JY, Bae ES, Park HJ, Oh D-C, Lee SK. Antimetastatic Activity of Apoptolidin A by Upregulation of N-Myc Downstream-Regulated Gene 1 Expression in Human Colorectal Cancer Cells. Pharmaceuticals. 2023; 16(4):491. https://doi.org/10.3390/ph16040491
Chicago/Turabian StyleKyaw, Kay Zin, Jiyoon Park, Seung Ho Oh, Ji Yun Lee, Eun Seo Bae, Hyen Joo Park, Dong-Chan Oh, and Sang Kook Lee. 2023. "Antimetastatic Activity of Apoptolidin A by Upregulation of N-Myc Downstream-Regulated Gene 1 Expression in Human Colorectal Cancer Cells" Pharmaceuticals 16, no. 4: 491. https://doi.org/10.3390/ph16040491
APA StyleKyaw, K. Z., Park, J., Oh, S. H., Lee, J. Y., Bae, E. S., Park, H. J., Oh, D.-C., & Lee, S. K. (2023). Antimetastatic Activity of Apoptolidin A by Upregulation of N-Myc Downstream-Regulated Gene 1 Expression in Human Colorectal Cancer Cells. Pharmaceuticals, 16(4), 491. https://doi.org/10.3390/ph16040491