
Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions

 ,
,  and
and 
Abstract
1. Introduction

2. Results
2.1. Effect of Ag+ on AAC(2′)-Ia-Catalyzed Acetylation of Plazomicin
2.2. Effect of Ag+ on AAC(2′)-Ia-Mediated Resistance to Plazomicin
2.3. Bactericidal Effect
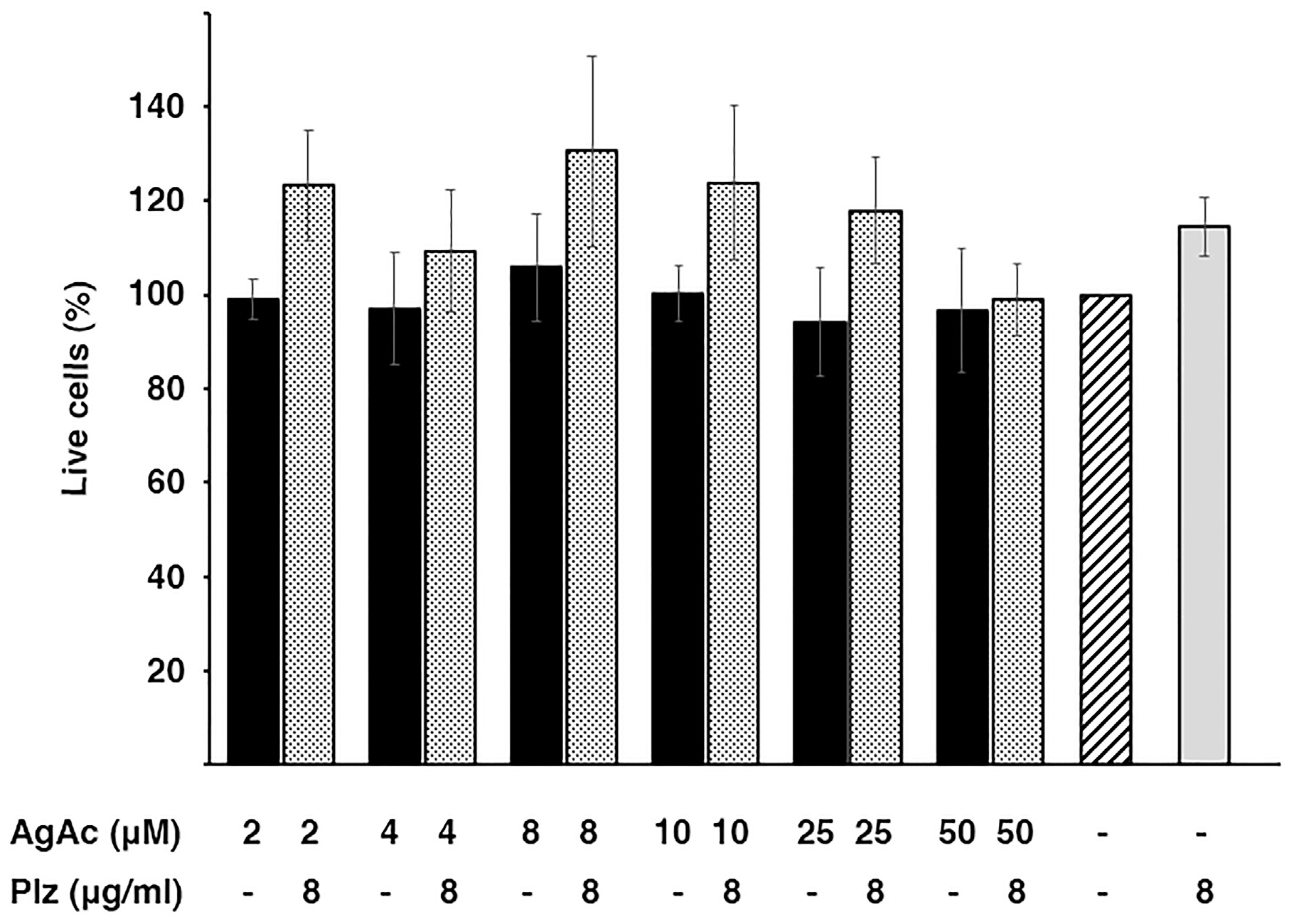
2.4. Cytotoxicity of the Mix Plazomicin/Silver Acetate
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
4.2. Bacterial Growth
4.3. MIC Determination
4.4. Time-Kill Assays
4.5. Acetyltransferase Assays
4.6. Cytotoxicity Assays
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boucher, H.W. Bad bugs, no drugs 2002–2020: Progress, challenges, and call to action. Trans. Am. Clin. Climatol. Assoc. 2020, 131, 65–71. [Google Scholar] [PubMed]
- Adler, A.; Friedman, N.D.; Marchaim, D. Multidrug-resistant Gram-negative bacilli: Infection control implications. Infect. Dis. Clin. N. Am. 2016, 30, 967–997. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Garau, J. Current and future perspectives in the treatment of multidrug-resistant Gram-negative infections. J. Antimicrob. Chemother. 2021, 76, iv23–iv37. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.; El Chakhtoura, N.G.; Yasmin, M.; Bonomo, R.A. Polymyxins: To combine or not to combine? Antibiotics 2019, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Machuca, I.; Gutierrez-Gutierrez, B.; Rivera-Espinar, F.; Cano, A.; Gracia-Ahufinger, I.; Guzman-Puche, J.; Marfil-Perez, E.; Perez-Nadales, E.; Caston, J.J.; Bonomo, R.A.; et al. External validation of the INCREMENT-CPE mortality score in a carbapenem-resistant Klebsiella pneumoniae bacteraemia cohort: The prognostic significance of colistin resistance. Int. J. Antimicrob. Agents 2019, 54, 442–448. [Google Scholar] [CrossRef]
- Rodriguez-Santiago, J.; Cornejo-Juarez, P.; Silva-Sanchez, J.; Garza-Ramos, U. Polymyxin resistance in Enterobacterales: Overview and epidemiology in the Americas. Int. J. Antimicrob. Agents 2021, 58, 106426. [Google Scholar] [CrossRef]
- Burillo, A.; Bouza, E. Controversies over the management of infections caused by Amp-C- and ESBL-producing Enterobacterales: What questions remain for future studies? Curr. Opin. Infect. Dis. 2022, 35, 575–582. [Google Scholar] [CrossRef]
- Sato, T.; Yamawaki, K. Cefiderocol: Discovery, chemistry, and in vivo profiles of a novel siderophore cephalosporin. Clin. Infect. Dis. 2019, 69, S538–S543. [Google Scholar] [CrossRef]
- Mezcord, V.; Wong, O.; Pasteran, F.; Corso, A.; Tolmasky, M.E.; Bonomo, R.A.; Ramirez, M.S. Role of beta-lactamase inhibitors on cefiderocol activity against carbapenem-resistant Acinetobacter species. Int. J. Antimicrob. Agents 2022, 61, 106700. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef]
- Labby, K.J.; Garneau-Tsodikova, S. Strategies to overcome the action of aminoglycoside-modifying enzymes for treating resistant bacterial infections. Future Med. Chem. 2013, 5, 1285–1309. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Nikolaidis, N.; Tolmasky, M.E. Rise and dissemination of aminoglycoside resistance: The aac(6′)-Ib paradigm. Front. Microbiol. 2013, 4, 121. [Google Scholar] [CrossRef] [PubMed]
- Tolmasky, M.E. Strategies to Prolong the Useful Life of Existing Antibiotics and Help Overcoming the Antibiotic Resistance Crisis. In Frontiers in Clinical Drug Research-Anti Infectives; Atta-ur-Rhaman, Ed.; Bentham Books: Sharjah, United Arab Emirates, 2017; Volume 1, pp. 1–27. [Google Scholar]
- Kondo, S.; Hotta, K. Semisynthetic aminoglycoside antibiotics: Development and enzymatic modifications. J. Infect. Chemother. 1999, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Umemura, E.; Sakamoto, Y.; Takahashi, Y.; Miyake, T. Synthesis and antibacterial activity of 1-N-[(S)-omega-amino-2-hydroxyalkyl] derivatives of dibekacin, 5-deoxydibekacin, 3′-deoxykanamycin A and gentamicin B. J. Antibiot. 2015, 68, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, T.; Sati, G.C.; Kondasinghe, N.; Pirrone, M.G.; Kato, T.; Waduge, P.; Kumar, H.S.; Sanchon, A.C.; Dobosz-Bartoszek, M.; Shcherbakov, D.; et al. Design, multigram synthesis, and in vitro and in vivo evaluation of propylamycin: A semisynthetic 4,5-deoxystreptamine class aminoglycoside for the treatment of drug-resistant Enterobacteriaceae and other Gram-negative pathogens. J. Am. Chem. Soc. 2019, 141, 5051–5061. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Tolmasky, M.E. Amikacin: Uses, resistance, and prospects for inhibition. Molecules 2017, 22, 2267. [Google Scholar] [CrossRef]
- Serio, A.W.; Keepers, T.; Andrews, L.; Krause, K.M. Aminoglycoside revival: Review of a historically important class of antimicrobials undergoing rejuvenation. EcoSal Plus 2018, 8, 1. [Google Scholar] [CrossRef]
- Umezawa, H.; Umezawa, S.; Tsuchiya, T.; Okazaki, Y. 3′,4′-dideoxy-kanamycin B active against kanamycin-resistant Escherichia coli and Pseudomonas aeruginosa. J. Antibiot. 1971, 24, 485–487. [Google Scholar] [CrossRef]
- Guay, D.R. Netilmicin (Netromycin, Schering-Plough). Drug Intell. Clin. Pharm. 1983, 17, 83–91. [Google Scholar]
- Kawaguchi, H.; Naito, T.; Nakagawa, S.; Fujisawa, K.I. BB-K 8, a new semisynthetic aminoglycoside antibiotic. J. Antibiot. 1972, 25, 695–708. [Google Scholar] [CrossRef]
- Hotta, K.; Kondo, S. Kanamycin and its derivative, arbekacin: Significance and impact. J. Antibiot. 2018, 71, 417–424. [Google Scholar] [CrossRef]
- Barr, W.H.; Colucci, R.; Radwanski, E.; Zampaglione, N.; Cutler, D.; Lin, C.C.; Elliott, M.; Affrime, M.B. Pharmacokinetics of isepamicin. J. Chemother. 1995, 7 (Suppl. S2), 53–61. [Google Scholar]
- Woloj, M.; Tolmasky, M.E.; Roberts, M.C.; Crosa, J.H. Plasmid-encoded amikacin resistance in multiresistant strains of Klebsiella pneumoniae isolated from neonates with meningitis. Antimicrob. Agents Chemother. 1986, 29, 315–319. [Google Scholar] [CrossRef]
- Tolmasky, M.E. Aminoglycoside-modifying enzymes: Characteristics, localization, and dissemination. In Enzyme-Mediated Resistance to Antibiotics: Mechanisms, Dissemination, and Prospects for Inhibition; Bonomo, R.A., Tolmasky, M.E., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 35–52. [Google Scholar]
- Aggen, J.B.; Armstrong, E.S.; Goldblum, A.A.; Dozzo, P.; Linsell, M.S.; Gliedt, M.J.; Hildebrandt, D.J.; Feeney, L.A.; Kubo, A.; Matias, R.D.; et al. Synthesis and spectrum of the neoglycoside ACHN-490. Antimicrob. Agents Chemother. 2010, 54, 4636–4642. [Google Scholar] [CrossRef]
- Endimiani, A.; Hujer, K.M.; Hujer, A.M.; Armstrong, E.S.; Choudhary, Y.; Aggen, J.B.; Bonomo, R.A. ACHN-490, a neoglycoside with potent in vitro activity against multidrug-resistant Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 2009, 53, 4504–4507. [Google Scholar] [CrossRef]
- Cox, G.; Ejim, L.; Stogios, P.J.; Koteva, K.; Bordeleau, E.; Evdokimova, E.; Sieron, A.O.; Savchenko, A.; Serio, A.W.; Krause, K.M.; et al. Plazomicin retains antibiotic activity against most aminoglycoside modifying enzymes. ACS Infect. Dis. 2018, 4, 980–987. [Google Scholar] [CrossRef]
- Saravolatz, L.D.; Stein, G.E. Plazomicin: A new aminoglycoside. Clin. Infect. Dis. 2020, 70, 704–709. [Google Scholar] [CrossRef]
- Lin, J.; Nishino, K.; Roberts, M.C.; Tolmasky, M.; Aminov, R.I.; Zhang, L. Mechanisms of antibiotic resistance. Front. Microbiol. 2015, 6, 34. [Google Scholar] [CrossRef]
- Clark, J.A.; Burgess, D.S. Plazomicin: A new aminoglycoside in the fight against antimicrobial resistance. Ther. Adv. Infect. Dis. 2020, 7, 2049936120952604. [Google Scholar] [CrossRef]
- Tang, H.J.; Lai, C.C. Plazomicin-associated nephrotoxicity. Clin. Infect. Dis. 2020, 71, 1130–1131. [Google Scholar] [CrossRef]
- Alfieri, A.; Di Franco, S.; Donatiello, V.; Maffei, V.; Fittipaldi, C.; Fiore, M.; Coppolino, F.; Sansone, P.; Pace, M.C.; Passavanti, M.B. Plazomicin against multidrug-resistant bacteria: A scoping review. Life 2022, 12, 1949. [Google Scholar] [CrossRef] [PubMed]
- Macinga, D.R.; Rather, P.N. The chromosomal 2′-N-acetyltransferase of Providencia stuartii: Physiological functions and genetic regulation. Front. Biosci. 1999, 4, D132–D140. [Google Scholar] [CrossRef]
- Bassenden, A.V.; Dumalo, L.; Park, J.; Blanchet, J.; Maiti, K.; Arya, D.P.; Berghuis, A.M. Structural and phylogenetic analyses of resistance to next-generation aminoglycosides conferred by AAC(2′) enzymes. Sci. Rep. 2021, 11, 11614. [Google Scholar] [CrossRef]
- Golkar, T.; Bassenden, A.V.; Maiti, K.; Arya, D.P.; Schmeing, T.M.; Berghuis, A.M. Structural basis for plazomicin antibiotic action and resistance. Commun. Biol. 2021, 4, 729. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.L.; Tran, T.; Alam, J.Y.; Herron, S.R.; Ramirez, M.S.; Tolmasky, M.E. Inhibition of aminoglycoside 6′-N-acetyltransferase type Ib by zinc: Reversal of amikacin resistance in Acinetobacter baumannii and Escherichia coli by a zinc ionophore. Antimicrob. Agents Chemother. 2014, 58, 4238–4241. [Google Scholar] [CrossRef]
- Chiem, K.; Fuentes, B.A.; Lin, D.L.; Tran, T.; Jackson, A.; Ramirez, M.S.; Tolmasky, M.E. Inhibition of aminoglycoside 6′-N-acetyltransferase type Ib-mediated amikacin resistance in Klebsiella pneumoniae by zinc and copper pyrithione. Antimicrob. Agents Chemother. 2015, 59, 5851–5853. [Google Scholar] [CrossRef] [PubMed]
- Chiem, K.; Hue, F.; Magallon, J.; Tolmasky, M.E. Inhibition of aminoglycoside 6′-N-acetyltransferase type Ib-mediated amikacin resistance by zinc complexed with clioquinol, an ionophore active against tumors and neurodegenerative diseases. Int. J. Antimicrob. Agents 2018, 51, 271–273. [Google Scholar] [CrossRef]
- Magallon, J.; Chiem, K.; Tran, T.; Ramirez, M.S.; Jimenez, V.; Tolmasky, M.E. Restoration of susceptibility to amikacin by 8-hydroxyquinoline analogs complexed to zinc. PLoS ONE 2019, 14, e0217602. [Google Scholar] [CrossRef]
- Magallon, J.; Vu, P.; Reeves, C.; Kwan, S.; Phan, K.; Oakley-Havens, C.; Ramirez, M.S.; Tolmasky, M.E. Amikacin in combination with zinc pyrithione prevents growth of a carbapenem-resistant/multidrug-resistant Klebsiella pneumoniae isolate. Int. J. Antimicrob. Agents 2021, 58, 106442. [Google Scholar] [CrossRef]
- Reeves, C.M.; Magallon, J.; Rocha, K.; Tran, T.; Phan, K.; Vu, P.; Yi, Y.; Oakley-Havens, C.L.; Cedano, J.; Jimenez, V.; et al. Aminoglycoside 6′-N-acetyltransferase type Ib [AAC(6′)-Ib]-mediated aminoglycoside resistance: Phenotypic conversion to susceptibility by silver ions. Antibiotics 2020, 10, 29. [Google Scholar] [CrossRef]
- Li, Y.; Green, K.D.; Johnson, B.R.; Garneau-Tsodikova, S. Inhibition of aminoglycoside acetyltransferase resistance enzymes by metal salts. Antimicrob. Agents Chemother. 2015, 59, 4148–4156. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Iinuma, K.; Yamamoto, H.; Maeda, K.; Umezawa, H. Syntheses of 1-n-(S)-4-amino-2-hydroxybutyryl)-kanamycin B and -3′, 4′-dideoxykanamycin B active against kanamycin-resistant bacteria. J. Antibiot. 1973, 26, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.N. Isepamicin (SCH 21420, 1-N-HAPA gentamicin B): Microbiological characteristics including antimicrobial potency of spectrum of activity. J. Chemother. 1995, 7 (Suppl. S2), 7–16. [Google Scholar] [PubMed]
- Ding, X.; Baca-DeLancey, R.R.; Rather, P.N. Role of SspA in the density-dependent expression of the transcriptional activator AarP in Providencia stuartii. FEMS Microbiol. Lett. 2001, 196, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Thwaites, M.; Hall, D.; Shinabarger, D.; Serio, A.W.; Krause, K.M.; Marra, A.; Pillar, C. Evaluation of the bactericidal activity of plazomicin and comparators against multidrug-resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, 8. [Google Scholar] [CrossRef]
- Fauci, A.S. Emerging and reemerging infectious diseases: The perpetual challenge. Acad. Med. 2005, 80, 1079–1085. [Google Scholar] [CrossRef]
- Anil, A.; Banerjee, A. Pneumococcal encounter with the blood-brain barrier endothelium. Front. Cell Infect. Microbiol. 2020, 10, 590682. [Google Scholar] [CrossRef]
- Seale, M.; Lee, W.K.; Daffy, J.; Tan, Y.; Trost, N. Fulminant endogenous Klebsiella pneumoniae endophthalmitis: Imaging findings. Emerg. Radiol. 2007, 13, 209–212. [Google Scholar] [CrossRef]
- Sprenger, M.; Fukuda, K. Antimicrobial resistance. New mechanisms, new worries. Science 2016, 351, 1263–1264. [Google Scholar] [CrossRef]
- Goldstein, E. Rise in the prevalence of resistance to extended-spectrum cephalosporins in the USA, nursing homes and antibiotic prescribing in outpatient and inpatient settings. J. Antimicrob. Chemother. 2021, 76, 2745–2747. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar]
- Pierce, G.N.; Resch, C.; Mourin, M.; Dibrov, P.; Dibrov, E.; Ravandi, A. Bacteria and the growing threat of multidrug resistance for invasive cardiac interventions. Rev. Cardiovasc. Med. 2022, 23, 15. [Google Scholar] [CrossRef] [PubMed]
- Teillant, A.; Gandra, S.; Barter, D.; Morgan, D.J.; Laxminarayan, R. Potential burden of antibiotic resistance on surgery and cancer chemotherapy antibiotic prophylaxis in the USA: A literature review and modelling study. Lancet Infect. Dis. 2015, 15, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Nanayakkara, A.K.; Boucher, H.W.; Fowler, V.G., Jr.; Jezek, A.; Outterson, K.; Greenberg, D.E. Antibiotic resistance in the patient with cancer: Escalating challenges and paths forward. CA Cancer J. Clin. 2021, 71, 488–504. [Google Scholar] [CrossRef]
- WHO. Ten Threats to Global Health in 2019. Available online: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019 (accessed on 16 December 2022).
- Bush, K.; Bradford, P.A. Interplay between beta-lactamases and new beta-lactamase inhibitors. Nat. Rev. Microbiol. 2019, 17, 295–306. [Google Scholar] [CrossRef]
- Bohlmann, L.; De Oliveira, D.M.P.; El-Deeb, I.M.; Brazel, E.B.; Harbison-Price, N.; Ong, C.Y.; Rivera-Hernandez, T.; Ferguson, S.A.; Cork, A.J.; Phan, M.D.; et al. Chemical synergy between ionophore PBT2 and zinc reverses antibiotic resistance. mBio 2018, 9, 6. [Google Scholar] [CrossRef]
- Rather, P.N.; Orosz, E.; Shaw, K.J.; Hare, R.; Miller, G. Characterization and transcriptional regulation of the 2′-N-acetyltransferase gene from Providencia stuartii. J. Bacteriol. 1993, 175, 6492–6498. [Google Scholar] [CrossRef]
- Herisse, M.; Duverger, Y.; Martin-Verstraete, I.; Barras, F.; Ezraty, B. Silver potentiates aminoglycoside toxicity by enhancing their uptake. Mol. Microbiol. 2017, 105, 115–126. [Google Scholar] [CrossRef]
- Morones-Ramirez, J.R.; Winkler, J.A.; Spina, C.S.; Collins, J.J. Silver enhances antibiotic activity against Gram-negative bacteria. Sci. Transl. Med. 2013, 5, 190ra181. [Google Scholar] [CrossRef]
- Barras, F.; Aussel, L.; Ezraty, B. Silver and antibiotic, new facts to an old story. Antibiotics 2018, 7, 79. [Google Scholar] [CrossRef]
- Sutcliffe, J.G. Nucleotide sequence of the ampicillin resistance gene of Escherichia coli plasmid pBR322. Proc. Natl. Acad. Sci. USA 1978, 75, 3737–3741. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.J.; Dowding, J.E. Aminoglycoside-modifying enzymes. Methods Enzymol. 1975, 43, 611–628. [Google Scholar] [PubMed]
- Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 1977, 36, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.; Chiem, K.; Jani, S.; Arivett, B.A.; Lin, D.L.; Lad, R.; Jimenez, V.; Farone, M.B.; Debevec, G.; Santos, R.; et al. Identification of a small molecule inhibitor of the aminoglycoside 6′-N-acetyltransferase type Ib [AAC(6′)-Ib] using mixture-based combinatorial libraries. Int. J. Antimicrob. Agents 2018, 51, 752–761. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plazomicin (μg/mL) | Silver Acetate (μM) OD600 | Sodium Acetate (μM) OD600 | ||||
|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 4 | 0 | 8 | |
| 0 | 3.21 ± 0.02 | 3.09 ± 0.01 | 3.09 ± 0.02 | 3.18 ± 0.11 | 3.04 ± 0.06 | 3.12 ± 0.08 |
| 4 | 3.05 ± 0.08 | 2.86 ± 0.11 | 1.95 ± 0.01 | 0.12 ± 0.01 | 1.39 ± 0.03 | 1.34 ± 0.06 |
| 8 | 1.21 ± 0.01 | 1.26 ± 0.01 | 0.10 ± 0.03 | 0.02 ± 0 | 1.13 ± 0.02 | 1.08 ± 0.04 |
| Plazomicin (μg/mL) | Silver Acetate (μM) OD600 | |||
|---|---|---|---|---|
| 0 | 1 | 2 | 4 | |
| 0 | 3.54 ± 0.01 | 3.48 ± 0.02 | 3.50 ± 0.07 | 3.35 ± 0.10 |
| 4 | 1.71 ± 0.02 | 1.66 ± 0.09 | 0.95 ± 0.09 | 0.09 ± 0.07 |
| 8 | 0.68 ± 0.06 | 0.24 ± 0.02 | 0.14 ± 0.06 | 0.03 ± 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngo, D.; Magaña, A.J.; Tran, T.; Sklenicka, J.; Phan, K.; Eykholt, B.; Jimenez, V.; Ramirez, M.S.; Tolmasky, M.E. Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions. Pharmaceuticals 2023, 16, 236. https://doi.org/10.3390/ph16020236
Ngo D, Magaña AJ, Tran T, Sklenicka J, Phan K, Eykholt B, Jimenez V, Ramirez MS, Tolmasky ME. Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions. Pharmaceuticals. 2023; 16(2):236. https://doi.org/10.3390/ph16020236
Chicago/Turabian StyleNgo, David, Angel J. Magaña, Tung Tran, Jan Sklenicka, Kimberly Phan, Brian Eykholt, Verónica Jimenez, María S. Ramirez, and Marcelo E. Tolmasky. 2023. "Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions" Pharmaceuticals 16, no. 2: 236. https://doi.org/10.3390/ph16020236
APA StyleNgo, D., Magaña, A. J., Tran, T., Sklenicka, J., Phan, K., Eykholt, B., Jimenez, V., Ramirez, M. S., & Tolmasky, M. E. (2023). Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions. Pharmaceuticals, 16(2), 236. https://doi.org/10.3390/ph16020236