
Natural Compounds as Non-Nucleoside Inhibitors of Zika Virus Polymerase through Integration of In Silico and In Vitro Approaches
 , , , ,
, , , ,  , , , , ,
, , , , ,  ,
, 
Abstract
1. Introduction
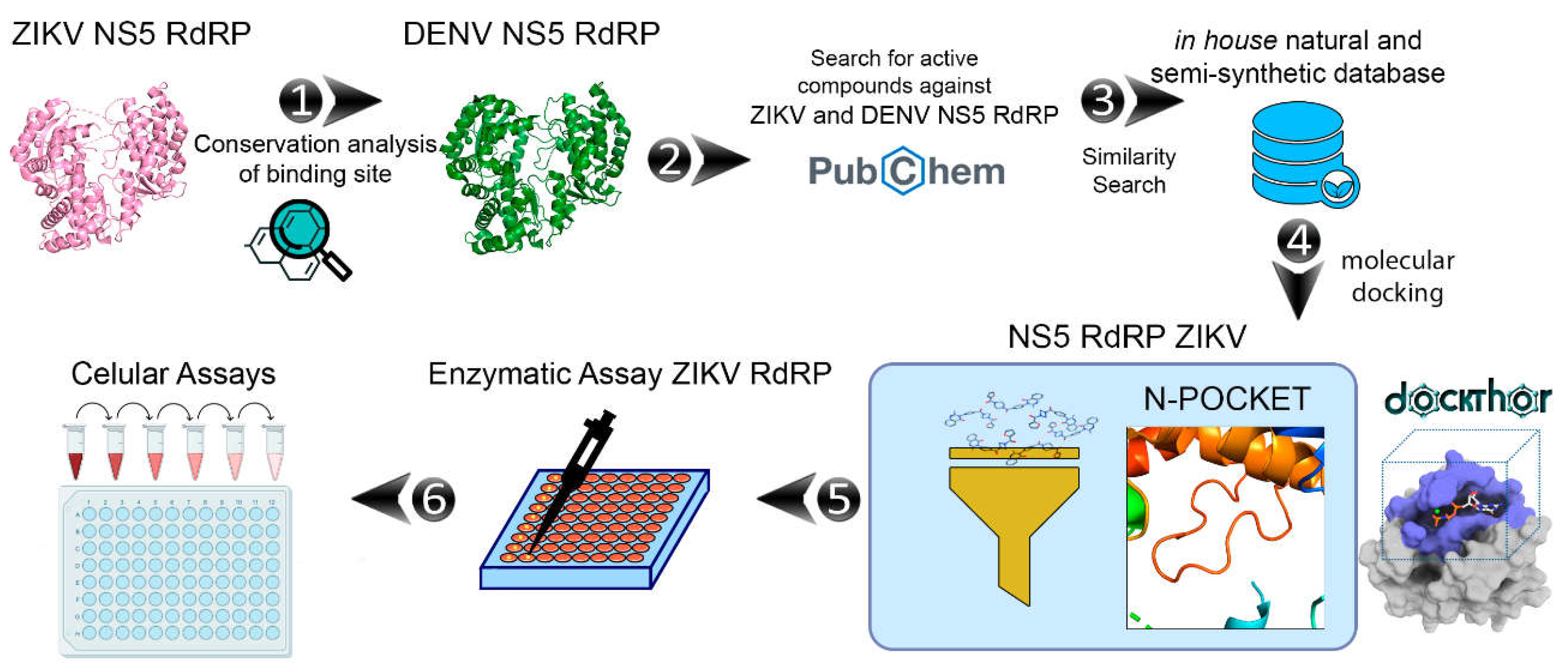
2. Results and Discussion
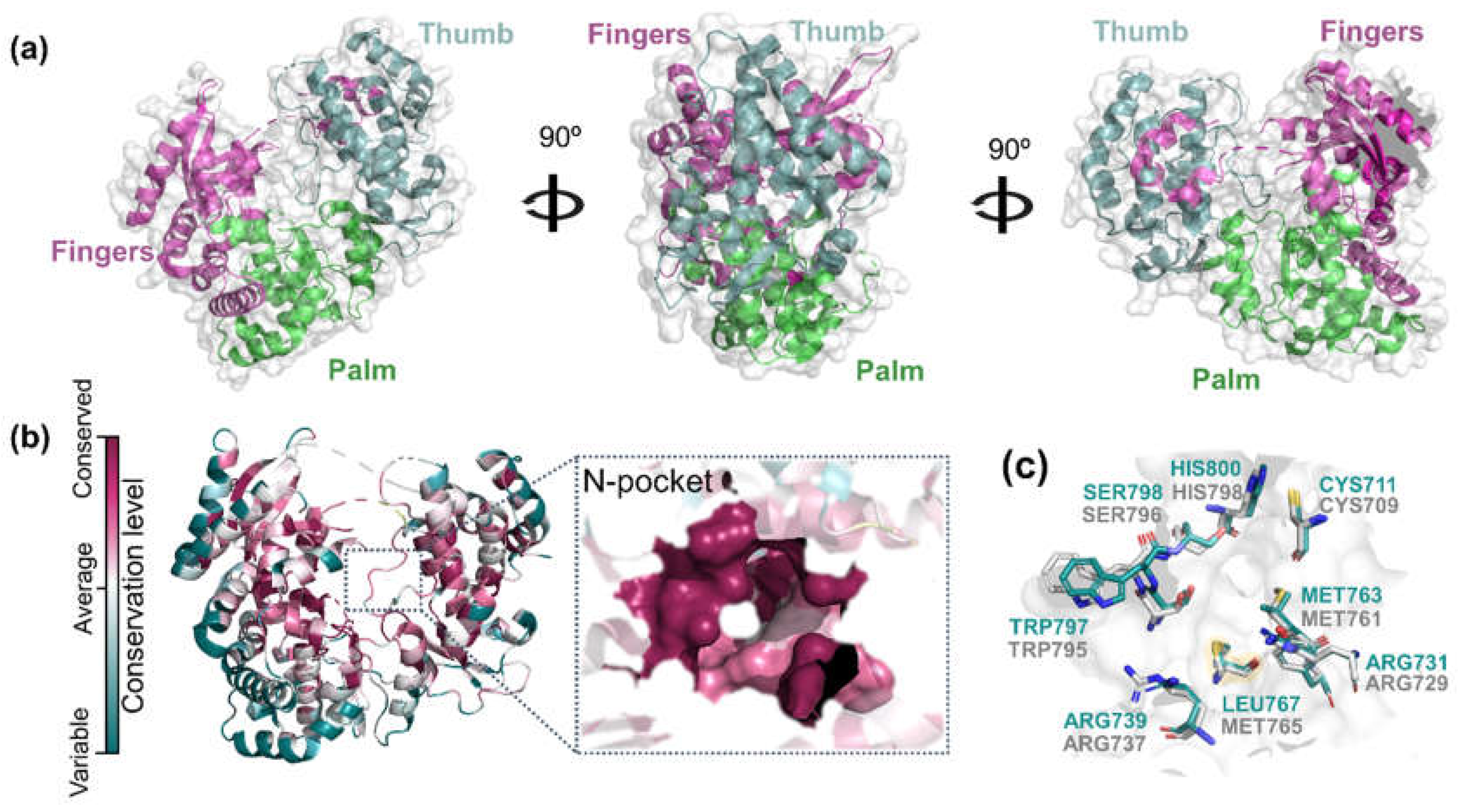
2.1. Binding Site Conservation Analysis
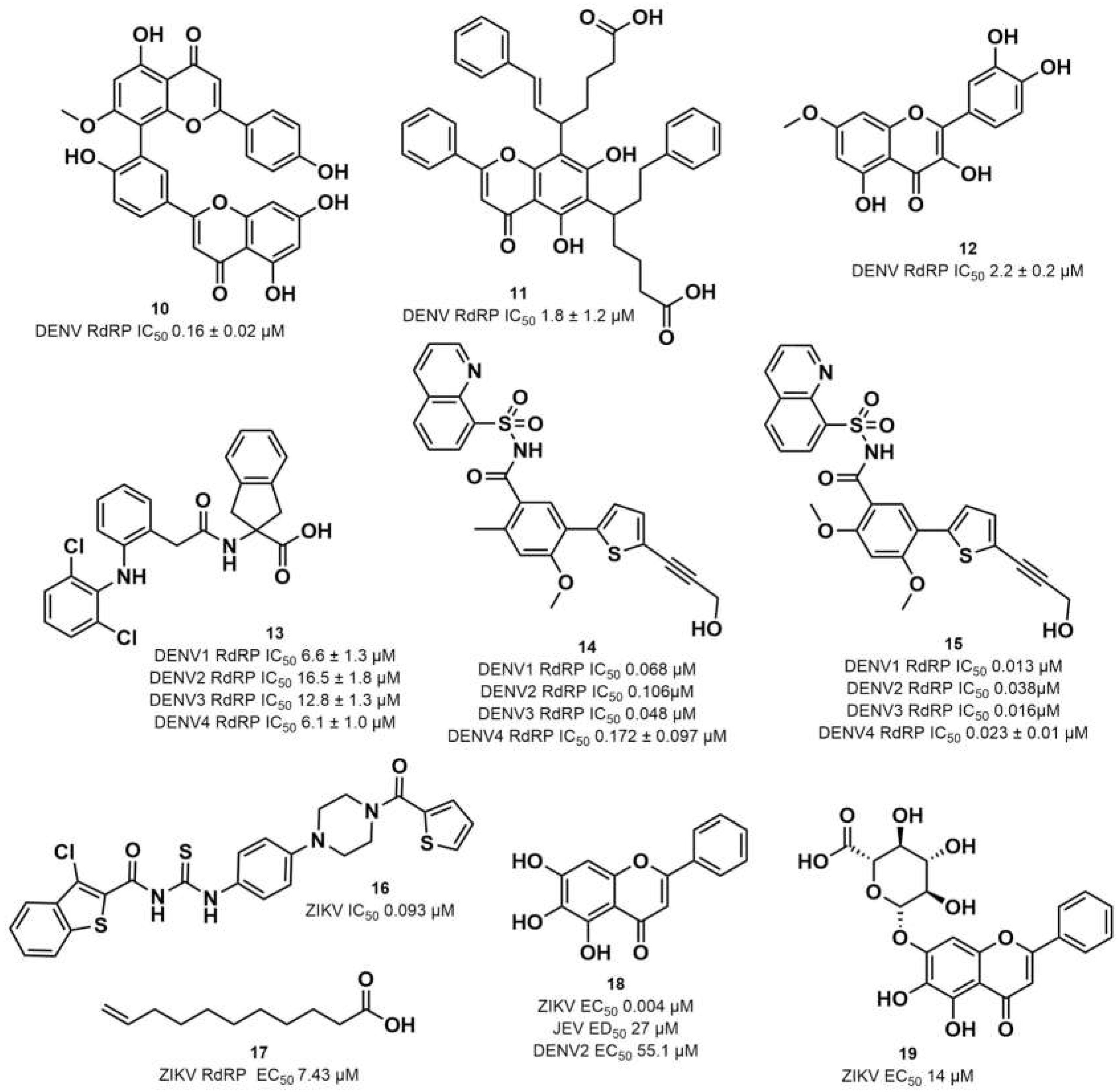
2.2. Chemical Space Analysis of RdRp Inhibitors
2.3. Docking Calculations at the ZIKV NS5 RdRp (N-Pocket)
2.4. Pedalitin and Quercetin Inhibits ZIKV RdRp Activity
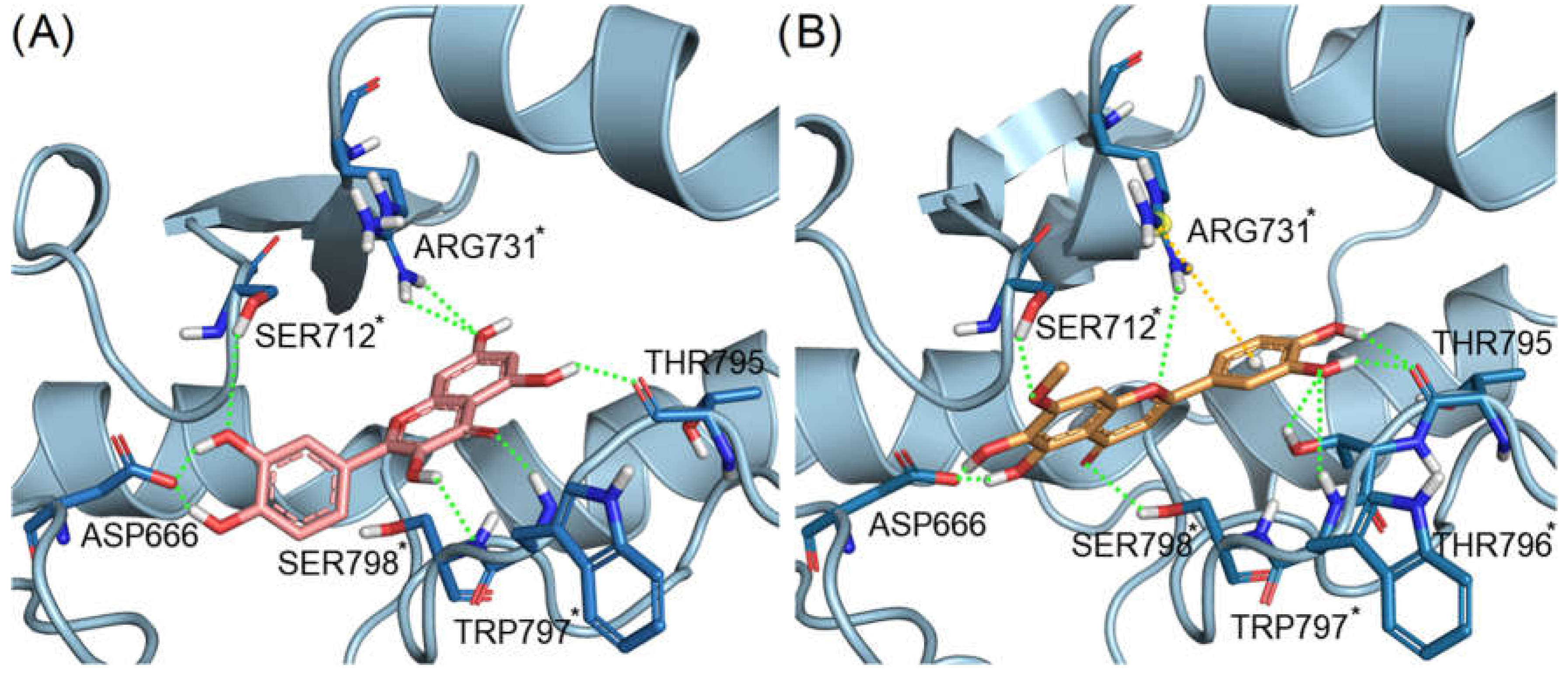
2.5. Pedalitin and Quercetin Binding Modes Predicted by Docking
2.6. Pedalitin and Quercetin Inhibits ZIKV Replication In Vitro
3. Materials and Methods
3.1. Computational
3.1.1. DENV and ZIKV NS5 RdRp Similarity Analysis
3.1.2. Collection of DENV RdRp Inhibitors
3.1.3. Chemical Space Analysis of RdRp Inhibitors
3.1.4. Protein and Ligand Preparation
3.1.5. Molecular Docking
3.2. Experimental
3.2.1. Quercetin and Pedalitin
3.2.2. Protein Cloning, Expression and Purification
3.2.3. NS5 RdRp Activity Assays
3.3. Cell Culture
3.4. Virus Rescue and Titration
3.5. Cell Viabillity
3.6. Antiviral Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- World Health Organization Zika Virus Outbreaks in the Americas. Relevé Épidémiologique Hebdomadaire/Section D’hygiène du Secrétariat de La Société des Nations = Weekly Epidemiological Record/Health Section of the Secretariat of the League of Nations; World Health Organization: Washington, DC, USA, 2015; Volume 90, pp. 609–610. [Google Scholar]
- Teixeira, G.A.; Dantas, D.N.A.; Carvalho, G.A.F.d.L.; da Silva, A.N.; Lira, A.L.B.d.C.; Enders, B.C. Analysis of the Concept of the Zika Virus Congenital Syndrome. Cienc. Saude Coletiva 2020, 25, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Dirlikov, E.; Major, C.G.; Mayshack, M.; Medina, N.; Matos, D.; Ryff, K.R.; Torres-Aponte, J.; Alkis, R.; Munoz-Jordan, J.; Colon-Sanchez, C.; et al. Guillain-Barré Syndrome during Ongoing Zika Virus Transmission—Puerto Rico, January 1–July 31, 2016. Morb. Mortal. Wkly. Rep. 2016, 65, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Regla-Nava, J.A.; Wang, Y.-T.; Fontes-Garfias, C.R.; Liu, Y.; Syed, T.; Susantono, M.; Gonzalez, A.; Viramontes, K.M.; Verma, S.K.; Kim, K.; et al. A Zika Virus Mutation Enhances Transmission Potential and Confers Escape from Protective Dengue Virus Immunity. Cell Rep. 2022, 39, 110655. [Google Scholar] [CrossRef] [PubMed]
- Mottin, M.; Borba, J.V.V.B.; Braga, R.C.; Torres, P.H.M.; Martini, M.C.; Proenca-Modena, J.L.; Judice, C.C.; Costa, F.T.M.; Ekins, S.; Perryman, A.L.; et al. The A–Z of Zika Drug Discovery. Drug Discov. Today 2018, 23, 1833–1847. [Google Scholar] [CrossRef] [PubMed]
- Malet, H.; Massé, N.; Selisko, B.; Romette, J.-L.; Alvarez, K.; Guillemot, J.C.; Tolou, H.; Yap, T.L.; Vasudevan, S.G.; Lescar, J.; et al. The Flavivirus Polymerase as a Target for Drug Discovery. Antiviral Res. 2008, 80, 23–35. [Google Scholar] [CrossRef]
- Noble, C.G.; Lim, S.P.; Chen, Y.-L.; Liew, C.W.; Yap, L.; Lescar, J.; Shi, P.-Y. Conformational Flexibility of the Dengue Virus RNA-Dependent RNA Polymerase Revealed by a Complex with an Inhibitor. J. Virol. 2013, 87, 5291–5295. [Google Scholar] [CrossRef]
- Lim, S.P.; Noble, C.G.; Seh, C.C.; Soh, T.S.; El Sahili, A.; Chan, G.K.Y.; Lescar, J.; Arora, R.; Benson, T.; Nilar, S.; et al. Potent Allosteric Dengue Virus NS5 Polymerase Inhibitors: Mechanism of Action and Resistance Profiling. PLoS Pathog. 2016, 12, e1005737. [Google Scholar] [CrossRef]
- Asselah, T. Sofosbuvir for the Treatment of Hepatitis C Virus. Expert Opin. Pharmacother. 2014, 15, 121–130. [Google Scholar] [CrossRef]
- Sacramento, C.Q.; de Melo, G.R.; de Freitas, C.S.; Rocha, N.; Hoelz, L.V.B.; Miranda, M.; Fintelman-Rodrigues, N.; Marttorelli, A.; Ferreira, A.C.; Barbosa-Lima, G.; et al. The Clinically Approved Antiviral Drug Sofosbuvir Inhibits Zika Virus Replication. Sci. Rep. 2017, 7, 40920. [Google Scholar] [CrossRef]
- Marshall, G.R. Computer-Aided Drug Design. Annu. Rev. Pharmacol. Toxicol. 1987, 27, 193–213. [Google Scholar] [CrossRef]
- Reynolds, C. Impact of Computational Structure-Based Methods on Drug Discovery. Curr. Pharm. Des. 2014, 20, 3380–3386. [Google Scholar] [CrossRef] [PubMed]
- Andrade, C.H.; Neves, B.J.; Melo-Filho, C.C.; Rodrigues, J.; Silva, D.C.; Braga, R.C.; Cravo, P.V.L. In Silico Chemogenomics Drug Repositioning Strategies for Neglected Tropical Diseases. Curr. Med. Chem. 2019, 26, 4355–4379. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazy, H.; Erez, E.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2010: Calculating Evolutionary Conservation in Sequence and Structure of Proteins and Nucleic Acids. Nucleic Acids Res. 2010, 38, W529–W533. [Google Scholar] [CrossRef] [PubMed]
- Celniker, G.; Nimrod, G.; Ashkenazy, H.; Glaser, F.; Martz, E.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf: Using Evolutionary Data to Raise Testable Hypotheses about Protein Function. Isr. J. Chem. 2013, 53, 199–206. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An Improved Methodology to Estimate and Visualize Evolutionary Conservation in Macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef]
- Noble, C.G.; Lim, S.P.; Arora, R.; Yokokawa, F.; Nilar, S.; Seh, C.C.; Wright, S.K.; Benson, T.E.; Smith, P.W.; Shi, P.-Y. A Conserved Pocket in the Dengue Virus Polymerase Identified through Fragment-Based Screening. J. Biol. Chem. 2016, 291, 8541–8548. [Google Scholar] [CrossRef]
- Duan, W.; Song, H.; Wang, H.; Chai, Y.; Su, C.; Qi, J.; Shi, Y.; Gao, G.F. The Crystal Structure of Zika Virus NS 5 Reveals Conserved Drug Targets. EMBO J. 2017, 36, 919–933. [Google Scholar] [CrossRef]
- Gharbi-Ayachi, A.; Santhanakrishnan, S.; Wong, Y.H.; Chan, K.W.K.; Tan, S.T.; Bates, R.W.; Vasudevan, S.G.; el Sahili, A.; Lescar, J. Non-Nucleoside Inhibitors of Zika Virus RNA-Dependent RNA Polymerase. J. Virol. 2020, 94, e00794-20. [Google Scholar] [CrossRef]
- Yin, Z.; Chen, Y.-L.; Kondreddi, R.R.; Chan, W.L.; Wang, G.; Ng, R.H.; Lim, J.Y.H.; Lee, W.Y.; Jeyaraj, D.A.; Niyomrattanakit, P.; et al. N-Sulfonylanthranilic Acid Derivatives as Allosteric Inhibitors of Dengue Viral RNA-Dependent RNA Polymerase. J. Med. Chem. 2009, 52, 7934–7937. [Google Scholar] [CrossRef]
- Allard, P.-M.; Dau, E.T.H.; Eydoux, C.; Guillemot, J.-C.; Dumontet, V.; Poullain, C.; Canard, B.; Guéritte, F.; Litaudon, M. Alkylated Flavanones from the Bark of Cryptocarya Chartacea as Dengue Virus NS5 Polymerase Inhibitors. J. Nat. Prod. 2011, 74, 2446–2453. [Google Scholar] [CrossRef]
- Bourjot, M.; Leyssen, P.; Eydoux, C.; Guillemot, J.-C.; Canard, B.; Rasoanaivo, P.; Guéritte, F.; Litaudon, M. Flacourtosides A-F, Phenolic Glycosides Isolated from Flacourtia Ramontchi. J. Nat. Prod. 2012, 75, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Benmansour, F.; Eydoux, C.; Querat, G.; de Lamballerie, X.; Canard, B.; Alvarez, K.; Guillemot, J.-C.; Barral, K. Novel 2-Phenyl-5-[(E)-2-(Thiophen-2-Yl)Ethenyl]-1,3,4-Oxadiazole and 3-Phenyl-5-[(E)-2-(Thiophen-2-Yl)Ethenyl]-1,2,4-Oxadiazole Derivatives as Dengue Virus Inhibitors Targeting NS5 Polymerase. Eur. J. Med. Chem. 2016, 109, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Yokokawa, F.; Nilar, S.; Noble, C.G.; Lim, S.P.; Rao, R.; Tania, S.; Wang, G.; Lee, G.; Hunziker, J.; Karuna, R.; et al. Discovery of Potent Non-Nucleoside Inhibitors of Dengue Viral RNA-Dependent RNA Polymerase from a Fragment Hit Using Structure-Based Drug Design. J. Med. Chem. 2016, 59, 3935–3952. [Google Scholar] [CrossRef] [PubMed]
- Cannalire, R.; Tarantino, D.; Astolfi, A.; Barreca, M.L.; Sabatini, S.; Massari, S.; Tabarrini, O.; Milani, M.; Querat, G.; Mastrangelo, E.; et al. Functionalized 2,1-Benzothiazine 2,2-Dioxides as New Inhibitors of Dengue NS5 RNA-Dependent RNA Polymerase. Eur. J. Med. Chem. 2018, 143, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Lim, S.P.; Chen, Y.-L.; Hunziker, J.; Rao, R.; Gu, F.; Seh, C.C.; Ghafar, N.A.; Xu, H.; Chan, K.; et al. Structure-Activity Relationship of Uridine-Based Nucleoside Phosphoramidate Prodrugs for Inhibition of Dengue Virus RNA-Dependent RNA Polymerase. Bioorganic Med. Chem. Lett. 2018, 28, 2324–2327. [Google Scholar] [CrossRef]
- Cannalire, R.; Chan, K.W.K.; Burali, M.S.; Gwee, C.P.; Wang, S.; Astolfi, A.; Massari, S.; Sabatini, S.; Tabarrini, O.; Mastrangelo, E.; et al. Pyridobenzothiazolones Exert Potent Anti-Dengue Activity by Hampering Multiple Functions of NS5 Polymerase. ACS Med. Chem. Lett. 2020, 11, 773–782. [Google Scholar] [CrossRef]
- Peyrat, L.-A.; Eparvier, V.; Eydoux, C.; Guillemot, J.-C.; Litaudon, M.; Stien, D. Carneic Acids from an Endophytic Phomopsis sp. as Dengue Virus Polymerase Inhibitors. J. Nat. Prod. 2020, 83, 2330–2336. [Google Scholar] [CrossRef]
- Felicetti, T.; Burali, M.S.; Gwee, C.P.; Ki Chan, K.W.; Alonso, S.; Massari, S.; Sabatini, S.; Tabarrini, O.; Barreca, M.L.; Cecchetti, V.; et al. Sustainable, Three-Component, One-Pot Procedure to Obtain Active Anti-Flavivirus Agents. Eur. J. Med. Chem. 2021, 210, 112992. [Google Scholar] [CrossRef]
- Coulerie, P.; Nour, M.; Maciuk, A.; Eydoux, C.; Guillemot, J.-C.; Lebouvier, N.; Hnawia, E.; Leblanc, K.; Lewin, G.; Canard, B.; et al. Structure-Activity Relationship Study of Biflavonoids on the Dengue Virus Polymerase DENV-NS5 RdRp. Planta Med. 2013, 79, 1313–1318. [Google Scholar] [CrossRef]
- Coulerie, P.; Eydoux, C.; Hnawia, E.; Stuhl, L.; Maciuk, A.; Lebouvier, N.; Canard, B.; Figadère, B.; Guillemot, J.-C.; Nour, M. Biflavonoids of Dacrydium Balansae with Potent Inhibitory Activity on Dengue 2 NS5 Polymerase. Planta Med. 2012, 78, 672–677. [Google Scholar] [CrossRef]
- Bourjot, M.; Leyssen, P.; Eydoux, C.; Guillemot, J.-C.; Canard, B.; Rasoanaivo, P.; Guéritte, F.; Litaudon, M. Chemical Constituents of Anacolosa Pervilleana and Their Antiviral Activities. Fitoterapia 2012, 83, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Manvar, D.; Küçükgüzel, İ.; Erensoy, G.; Tatar, E.; Deryabaşoğulları, G.; Reddy, H.; Talele, T.T.; Cevik, O.; Kaushik-Basu, N. Discovery of Conjugated Thiazolidinone-Thiadiazole Scaffold as Anti-Dengue Virus Polymerase Inhibitors. Biochem. Biophys. Res. Commun. 2016, 469, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Niyomrattanakit, P.; Chen, Y.-L.; Dong, H.; Yin, Z.; Qing, M.; Glickman, J.F.; Lin, K.; Mueller, D.; Voshol, H.; Lim, J.Y.H.; et al. Inhibition of Dengue Virus Polymerase by Blocking of the RNA Tunnel. J. Virol. 2010, 84, 5678–5686. [Google Scholar] [CrossRef] [PubMed]
- Venkatesham, A.; Saudi, M.; Kaptein, S.; Neyts, J.; Rozenski, J.; Froeyen, M.; Van Aerschot, A. Aminopurine and Aminoquinazoline Scaffolds for Development of Potential Dengue Virus Inhibitors. Eur. J. Med. Chem. 2017, 126, 101–109. [Google Scholar] [CrossRef]
- Kaushik, S.; Dar, L.; Kaushik, S.; Yadav, J.P. Identification and Characterization of New Potent Inhibitors of Dengue Virus NS5 Proteinase from Andrographis Paniculata Supercritical Extracts on in Animal Cell Culture and in Silico Approaches. J. Ethnopharmacol. 2021, 267, 113541. [Google Scholar] [CrossRef]
- Mottin, M.; Caesar, L.K.; Brodsky, D.; Mesquita, N.C.M.R.; de Oliveira, K.Z.; Noske, G.D.; Sousa, B.K.P.; Ramos, P.R.P.S.; Jarmer, H.; Loh, B.; et al. Chalcones from Angelica Keiskei (Ashitaba) Inhibit Key Zika Virus Replication Proteins. Bioorganic Chem. 2022, 120, 105649. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, H.; Song, W.; Si, S.; Han, Y.; Jiang, J. Identification and Characterization of Zika Virus NS5 RNA-Dependent RNA Polymerase Inhibitors. Int. J. Antimicrob. Agents 2019, 54, 502–506. [Google Scholar] [CrossRef]
- Yang, S.; Xu, M.; Lee, E.M.; Gorshkov, K.; Shiryaev, S.A.; He, S.; Sun, W.; Cheng, Y.-S.; Hu, X.; Tharappel, A.M.; et al. Emetine Inhibits Zika and Ebola Virus Infections through Two Molecular Mechanisms: Inhibiting Viral Replication and Decreasing Viral Entry. Cell Discov. 2018, 4, 31. [Google Scholar] [CrossRef]
- Bernatchez, J.A.; Tran, L.T.; Li, J.; Luan, Y.; Siqueira-Neto, J.L.; Li, R. Drugs for the Treatment of Zika Virus Infection. J. Med. Chem. 2020, 63, 470–489. [Google Scholar] [CrossRef]
- Vincetti, P.; Kaptein, S.J.F.; Costantino, G.; Neyts, J.; Radi, M. Scaffold Morphing Approach To Expand the Toolbox of Broad-Spectrum Antivirals Blocking Dengue/Zika Replication. ACS Med. Chem. Lett. 2019, 10, 558–563. [Google Scholar] [CrossRef]
- Hinton, G.; Roweis, S. Stochastic Neighbor Embedding. Adv. Neural Inf. Process. Syst. 2003. [Google Scholar]
- Hopkins, A.L.; Groom, C.R.; Alex, A. Ligand Efficiency: A Useful Metric for Lead Selection. Drug Discov. Today 2004, 9, 430–431. [Google Scholar] [CrossRef]
- Kenny, P.W. The Nature of Ligand Efficiency. J. Cheminform. 2019, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Abad-Zapatero, C. Ligand Efficiency Indices for Effective Drug Discovery. Expert Opin. Drug Discov. 2007, 2, 469–488. [Google Scholar] [CrossRef]
- Hevener, K.E.; Pesavento, R.; Ren, J.; Lee, H.; Ratia, K.; Johnson, M.E. Hit-to-Lead: Hit Validation and Assessment. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2018; pp. 265–309. [Google Scholar]
- Neves, B.J.; Mottin, M.; Moreira-Filho, J.T.; De, B.K.; Sousa, P.; Mendonça, S.S.; Andrade, C.H. Best Practices for Docking-Based Virtual Screening. In Molecular Docking for Computer-Aided Drug Design; Elsevier: Amsterdam, The Netherlands, 2021; pp. 75–98. ISBN 9780128223123. [Google Scholar]
- Mottin, M.; de Paula Sousa, B.K.; de Moraes Roso Mesquita, N.C.; de Oliveira, K.I.Z.; Noske, G.D.; Sartori, G.R.; de Oliveira Albuquerque, A.; Urbina, F.; Puhl, A.C.; Moreira-Filho, J.T.; et al. Discovery of New Zika Protease and Polymerase Inhibitors through the Open Science Collaboration Project OpenZika. J. Chem. Inf. Model. 2022. [Google Scholar] [CrossRef]
- Nascimento, I.J.d.S.; Santos-Júnior, P.F.d.S.; de Aquino, T.M.; de Araújo-Júnior, J.X.; da Silva-Júnior, E.F. Insights on Dengue and Zika NS5 RNA-Dependent RNA Polymerase (RdRp) Inhibitors. Eur. J. Med. Chem. 2021, 224, 113698. [Google Scholar] [CrossRef]
- Campos, G.R.F.; Bittar, C.; Jardim, A.C.G.; Shimizu, J.F.; Batista, M.N.; Paganini, E.R.; de Assis, L.R.; Bartlett, C.; Harris, M.; Bolzani, V.d.S.; et al. Hepatitis C Virus in Vitro Replication Is Efficiently Inhibited by Acridone Fac4. J. Gen. Virol. 2017, 98, 1693–1701. [Google Scholar] [CrossRef]
- Sasvari, Z.; Bach, S.; Blondel, M.; Nagy, P.D. Inhibition of RNA Recruitment and Replication of an RNA Virus by Acridine Derivatives with Known Anti-Prion Activities. PLoS ONE 2009, 4, e7376. [Google Scholar] [CrossRef]
- Sepúlveda, C.S.; Fascio, M.L.; Mazzucco, M.B.; Palacios, M.L.D.; Pellón, R.F.; García, C.C.; D’Accorso, N.B.; Damonte, E.B. Synthesis and Evaluation of N -Substituted Acridones as Antiviral Agents against Haemorrhagic Fever Viruses. Antivir. Chem. Chemother. 2008, 19, 41–47. [Google Scholar] [CrossRef]
- Arora, R.; Liew, C.W.; Soh, T.S.; Otoo, D.A.; Seh, C.C.; Yue, K.; Nilar, S.; Wang, G.; Yokokawa, F.; Noble, C.G.; et al. Two RNA Tunnel Inhibitors Bind in Highly Conserved Sites in Dengue Virus NS5 Polymerase: Structural and Functional Studies. J. Virol. 2020, 94, e01130-20. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Yokokawa, F.; Shi, P.-Y. The Search for Nucleoside/Nucleotide Analog Inhibitors of Dengue Virus. Antivir. Res. 2015, 122, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Sofia, M.J.; Chang, W.; Furman, P.A.; Mosley, R.T.; Ross, B.S. Nucleoside, Nucleotide, and Non-Nucleoside Inhibitors of Hepatitis C Virus NS5B RNA-Dependent RNA-Polymerase. J. Med. Chem. 2012, 55, 2481–2531. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-T.; Colby-Germinario, S.P.; Hassounah, S.A.; Fogarty, C.; Osman, N.; Palanisamy, N.; Han, Y.; Oliveira, M.; Quan, Y.; Wainberg, M.A. Evaluation of Sofosbuvir (β-D-2′-Deoxy-2′-α-Fluoro-2′-β-C-Methyluridine) as an Inhibitor of Dengue Virus Replication #. Sci. Rep. 2017, 7, 6345. [Google Scholar] [CrossRef] [PubMed]
- Pattnaik, A.; Palermo, N.; Sahoo, B.R.; Yuan, Z.; Hu, D.; Annamalai, A.S.; Vu, H.L.X.; Correas, I.; Prathipati, P.K.; Destache, C.J.; et al. Discovery of a Non-Nucleoside RNA Polymerase Inhibitor for Blocking Zika Virus Replication through in Silico Screening. Antivir. Res. 2018, 151, 78–86. [Google Scholar] [CrossRef]
- Munafò, F.; Donati, E.; Brindani, N.; Ottonello, G.; Armirotti, A.; De Vivo, M. Quercetin and Luteolin Are Single-Digit Micromolar Inhibitors of the SARS-CoV-2 RNA-Dependent RNA Polymerase. Sci. Rep. 2022, 12, 10571. [Google Scholar] [CrossRef]
- Oo, A.; Teoh, B.T.; Sam, S.S.; Bakar, S.A.; Zandi, K. Baicalein and Baicalin as Zika Virus Inhibitors. Arch. Virol. 2019, 164, 585–593. [Google Scholar] [CrossRef]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral Activities of Flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef]
- Moghaddam, E.; Teoh, B.-T.; Sam, S.-S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a Metabolite of Baicalein with Antiviral Activity against Dengue Virus. Sci. Rep. 2015, 4, 5452. [Google Scholar] [CrossRef]
- Fong, Y.D.; Chu, J.J.H. Natural Products as Zika Antivirals. Med. Res. Rev. 2022. [Google Scholar] [CrossRef]
- Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; Mustafa, M.R.; AbuBakar, S. Antiviral Activity of Four Types of Bioflavonoid against Dengue Virus Type-2. Virol. J. 2011, 8, 560. [Google Scholar] [CrossRef]
- Hoenen, T.; Groseth, A. Virus–Host Cell Interactions. Cells 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Amsden, H.; Kourko, O.; Roth, M.; Gee, K. Antiviral Activities of Interleukin-27: A Partner for Interferons? Front. Immunol. 2022, 13, 902853. [Google Scholar] [CrossRef] [PubMed]
- Iannello, A.; Debbeche, O.; Martin, E.; Attalah, L.H.; Samarani, S.; Ahmad, A. Viral Strategies for Evading Antiviral Cellular Immune Responses of the Host. J. Leukoc. Biol. 2006, 79, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Lubick, K.J.; Robertson, S.J.; McNally, K.L.; Freedman, B.A.; Rasmussen, A.L.; Taylor, R.T.; Walts, A.D.; Tsuruda, S.; Sakai, M.; Ishizuka, M.; et al. Flavivirus Antagonism of Type I Interferon Signaling Reveals Prolidase as a Regulator of IFNAR1 Surface Expression. Cell Host Microbe 2015, 18, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Laurent-Rolle, M.; Morrison, J.; Rajsbaum, R.; Macleod, J.M.L.; Pisanelli, G.; Pham, A.; Ayllon, J.; Miorin, L.; Martínez-Romero, C.; TenOever, B.R.; et al. The Interferon Signaling Antagonist Function of Yellow Fever Virus NS5 Protein Is Activated by Type I Interferon. Cell Host Microbe 2014, 16, 314–327. [Google Scholar] [CrossRef]
- De Maio, F.A.; Risso, G.; Iglesias, N.G.; Shah, P.; Pozzi, B.; Gebhard, L.G.; Mammi, P.; Mancini, E.; Yanovsky, M.J.; Andino, R.; et al. The Dengue Virus NS5 Protein Intrudes in the Cellular Spliceosome and Modulates Splicing. PLoS Pathog. 2016, 12, e1005841. [Google Scholar] [CrossRef]
- Shugar, D. Viral and Host-Cell Protein KinasesEnticing Antiviral Targets and Relevance of Nucleoside, and Viral Thymidine, Kinases. Pharmacol. Ther. 1999, 82, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Suzek, B.E.; Wang, Y.; Huang, H.; McGarvey, P.B.; Wu, C.H. UniRef Clusters: A Comprehensive and Scalable Alternative for Improving Sequence Similarity Searches. Bioinformatics 2015, 31, 926–932. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Kuma, K.; Miyata, T.; Toh, H. Improvement in the Accuracy of Multiple Sequence Alignment Program MAFFT. Genome Inform. 2005, 16, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Pupko, T.; Bell, R.E.; Mayrose, I.; Glaser, F.; Ben-Tal, N. Rate4Site: An Algorithmic Tool for the Identification of Functional Regions in Proteins by Surface Mapping of Evolutionary Determinants within Their Homologues. Bioinformatics 2002, 18, S71–S77. [Google Scholar] [CrossRef]
- Mayrose, I. Comparison of Site-Specific Rate-Inference Methods for Protein Sequences: Empirical Bayesian Methods Are Superior. Mol. Biol. Evol. 2004, 21, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System, Version 1.8; Schrödinger, LCC: New York, NY, USA, 2015.
- Bateman, A. UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, J.; Suzek, T.O.; Zhang, J.; Wang, J.; Zhou, Z.; Han, L.; Karapetyan, K.; Dracheva, S.; Shoemaker, B.A.; et al. PubChem’s BioAssay Database. Nucleic Acids Res. 2012, 40, D400-12. [Google Scholar] [CrossRef] [PubMed]
- Bolton, E.E.; Wang, Y.; Thiessen, P.A.; Bryant, S.H. PubChem: Integrated Platform of Small Molecules and Biological Activities. In Annual Reports in Computational Chemistry Volume 4; American Chemical Society: Washington, DC, USA, 2008; Volume 4, pp. 217–241. [Google Scholar]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound Databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New Data Content and Improved Web Interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Landrum, G. Open-Source Cheminformatics. Available online: Https://Www.Rdkit.Org (accessed on 10 August 2022).
- Schrödinger Release 2015-2: Protein Preparation Wizard; Schrödinger, LLC: New York, NY, USA, 2015.
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and Ligand Preparation: Parameters, Protocols, and Influence on Virtual Screening Enrichments. J. Comput. Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Shelley, J.C.; Cholleti, A.; Frye, L.L.; Greenwood, J.R.; Timlin, M.R.; Uchimaya, M. Epik: A Software Program for PK a Prediction and Protonation State Generation for Drug-like Molecules. J. Comput. Aided Mol. Des. 2007, 21, 681–691. [Google Scholar] [CrossRef]
- Schrödinger Release 2015-2: Epik; Schrödinger, LLC: New York, NY, USA, 2015.
- Schrödinger Release 2015-2: LigPrep; Schrödinger, LLC: New York, NY, USA, 2015.
- Santos, K.B.; Guedes, I.A.; Karl, A.L.M.; Dardenne, L.E. Highly Flexible Ligand Docking: Benchmarking of the DockThor Program on the LEADS-PEP Protein–Peptide Data Set. J. Chem. Inf. Model. 2020, 60, 667–683. [Google Scholar] [CrossRef]
- Guedes, I.A.; Barreto, A.M.S.; Marinho, D.; Krempser, E.; Kuenemann, M.A.; Sperandio, O.; Dardenne, L.E.; Miteva, M.A. New Machine Learning and Physics-Based Scoring Functions for Drug Discovery. Sci. Rep. 2021, 11, 3198. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully Automated Protein–Ligand Interaction Profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.S.; Mottin, M.; de Assis, L.R.; Mesquita, N.C.d.M.R.; Sousa, B.K.d.P.; Coimbra, L.D.; Santos, K.B.; Zorn, K.M.; Guido, R.V.C.; Ekins, S.; et al. Flavonoids from Pterogyne Nitens as Zika Virus NS2B-NS3 Protease Inhibitors. Bioorganic Chem. 2021, 109, 104719. [Google Scholar] [CrossRef] [PubMed]
- Godoy, A.S.; Lima, G.M.A.; Oliveira, K.I.Z.; Torres, N.U.; Maluf, F.V.; Guido, R.V.C.; Oliva, G. Crystal Structure of Zika Virus NS5 RNA-Dependent RNA Polymerase. Nat. Commun. 2017, 8, 14764. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.S.; Noske, G.D.; Gawriljuk, V.O.; de Oliveira, K.I.Z.; Godoy, A.S.; Mesquita, N.C.M.R.; Oliva, G. High-Throughput Antiviral Assays to Screen for Inhibitors of Zika Virus Replication. J. Vis. Exp. Jove 2021, e62422. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, R.S.; de Godoy, A.S.; Santos, I.A.; Noske, G.D.; de Oliveira, K.I.Z.; Gawriljuk, V.O.; Gomes Jardim, A.C.; Oliva, G. Discovery of an Imidazonaphthyridine and a Riminophenazine as Potent Anti-Zika Virus Agents through a Replicon-Based High-Throughput Screening. Virus Res. 2021, 299, 198388. [Google Scholar] [CrossRef] [PubMed]
- GraphPad Prism, Version 8.0.0 for Windows; GraphPad Software: San Diego, CA, USA. Available online: www.graphpad.com (accessed on 25 March 2021).
- Cugola, F.R.; Fernandes, I.R.; Russo, F.B.; Freitas, B.C.; Dias, J.L.M.; Guimarães, K.P.; Benazzato, C.; Almeida, N.; Pignatari, G.C.; Romero, S.; et al. The Brazilian Zika Virus Strain Causes Birth Defects in Experimental Models. Nature 2016, 534, 267–271. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Structure | Chemical Class | Docking Score (Kcal·mol−1) | LE * (Kcal·mol−1·Non-Hydrogen Atom−1) |
|---|---|---|---|---|
| 1 |  | Anthranilic acid derivative | −8.12 | 0.43 |
| 2 |  | Anthranilic acid derivative | −8.43 | 0.38 |
| 3 |  | Acridone | −8.68 | 0.38 |
| 4 |  | Acridone | −8.30 | 0.35 |
| 5 |  | Acridone | −8.72 | 0.35 |
| 6 Chrysin |  | Flavonoid | −8.14 | 0.43 |
| 7 Sorbifolin |  | Flavonoid | −8.36 | 0.38 |
| 8 Pedalitin |  | Flavonoid | −7.94 | 0.35 |
| 9 Quercetin |  | Flavonoid | −7.74 | 0.35 |
| Compound | Docking Score (Kcal·mol−1) | IC50 ZIKV RdRp (µM) | EC50 ZIKV (µM) | CC50 (µM) | SI * |
|---|---|---|---|---|---|
| pedalitin | −7.93 | 4.1 ± 0.3 | 19.28 | 83.66 | 4.34 |
| quercetin | −7.74 | 0.5 ± 0.1 | 17.74 | 35.99 | 2.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, P.R.P.d.S.; Mottin, M.; Lima, C.S.; Assis, L.R.; de Oliveira, K.Z.; Mesquita, N.C.d.M.R.; Cassani, N.M.; Santos, I.A.; Borba, J.V.V.B.; Fiaia Costa, V.A.; et al. Natural Compounds as Non-Nucleoside Inhibitors of Zika Virus Polymerase through Integration of In Silico and In Vitro Approaches. Pharmaceuticals 2022, 15, 1493. https://doi.org/10.3390/ph15121493
Ramos PRPdS, Mottin M, Lima CS, Assis LR, de Oliveira KZ, Mesquita NCdMR, Cassani NM, Santos IA, Borba JVVB, Fiaia Costa VA, et al. Natural Compounds as Non-Nucleoside Inhibitors of Zika Virus Polymerase through Integration of In Silico and In Vitro Approaches. Pharmaceuticals. 2022; 15(12):1493. https://doi.org/10.3390/ph15121493
Chicago/Turabian StyleRamos, Paulo Ricardo Pimenta da Silva, Melina Mottin, Caroline Sprengel Lima, Letícia R. Assis, Ketllyn Zagato de Oliveira, Nathalya Cristina de Moraes Roso Mesquita, Natasha Marques Cassani, Igor Andrade Santos, Joyce Villa Verde Bastos Borba, Vinícius Alexandre Fiaia Costa, and et al. 2022. "Natural Compounds as Non-Nucleoside Inhibitors of Zika Virus Polymerase through Integration of In Silico and In Vitro Approaches" Pharmaceuticals 15, no. 12: 1493. https://doi.org/10.3390/ph15121493
APA StyleRamos, P. R. P. d. S., Mottin, M., Lima, C. S., Assis, L. R., de Oliveira, K. Z., Mesquita, N. C. d. M. R., Cassani, N. M., Santos, I. A., Borba, J. V. V. B., Fiaia Costa, V. A., Neves, B. J., Guido, R. V. C., Oliva, G., Jardim, A. C. G., Regasini, L. O., & Andrade, C. H. (2022). Natural Compounds as Non-Nucleoside Inhibitors of Zika Virus Polymerase through Integration of In Silico and In Vitro Approaches. Pharmaceuticals, 15(12), 1493. https://doi.org/10.3390/ph15121493