Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus


Abstract
:1. Introduction
2. Results
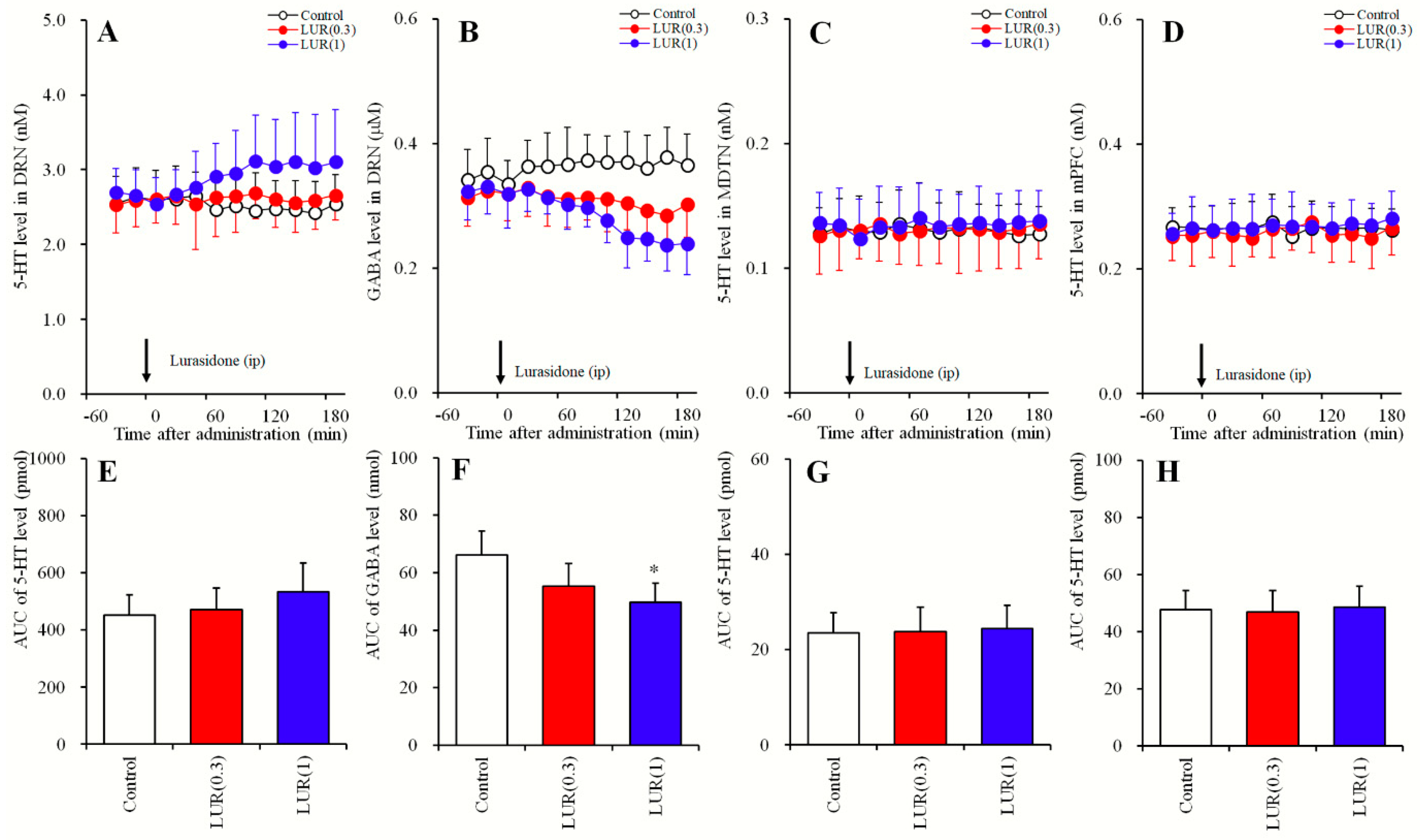
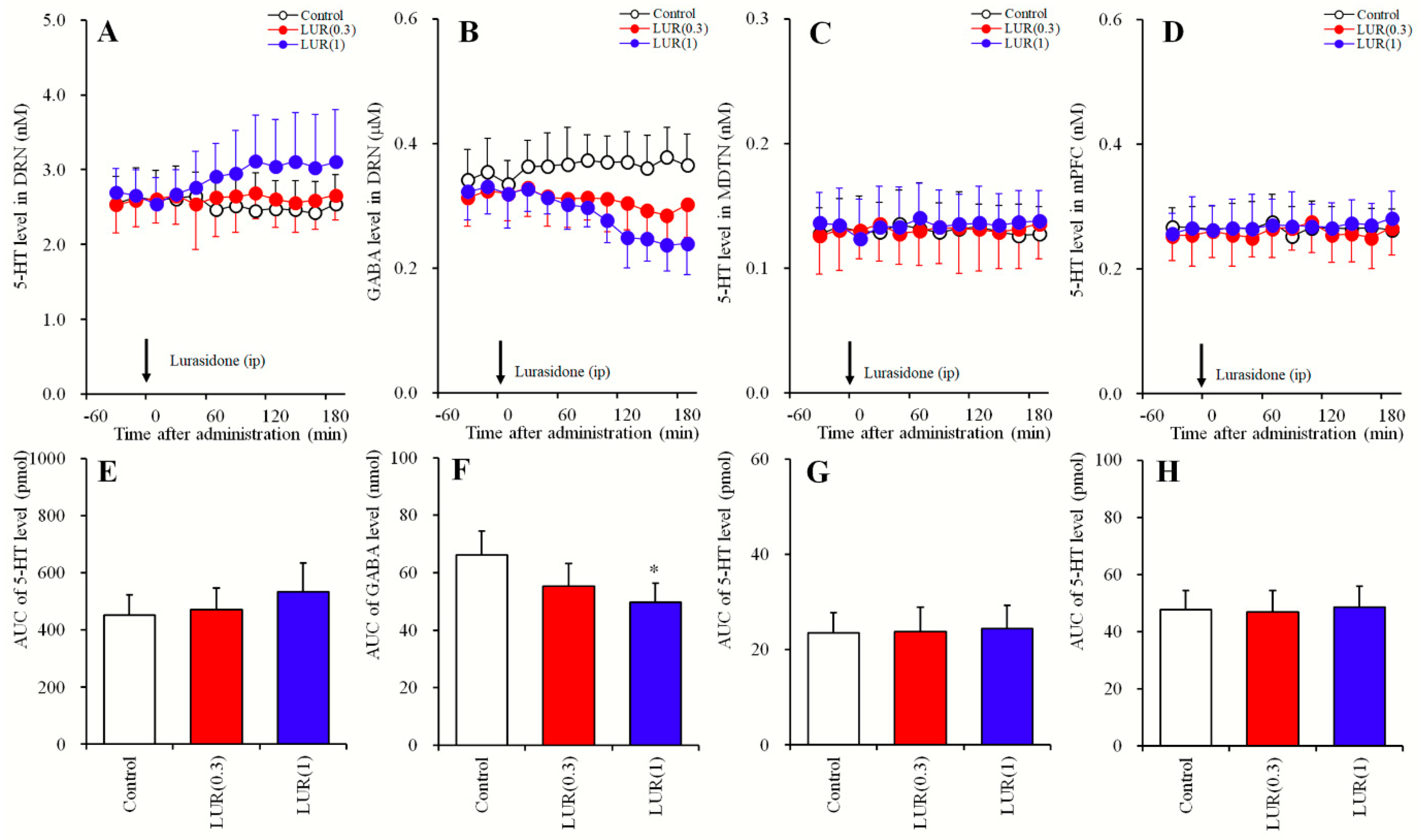
2.1. Dose-Dependent Effects of Systemic Lurasidone Administrations on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC, and GABA in the DRN (Study_1)
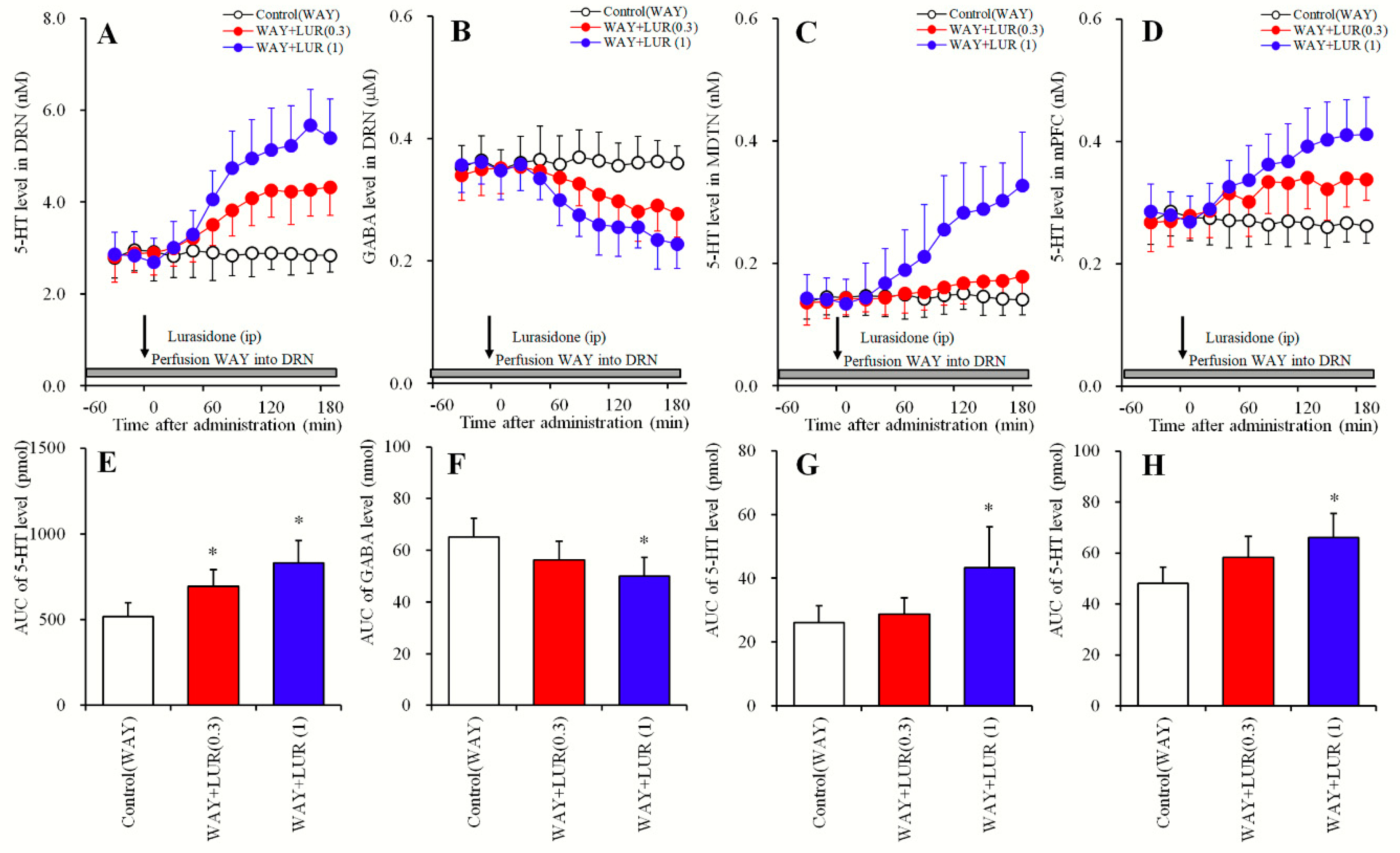
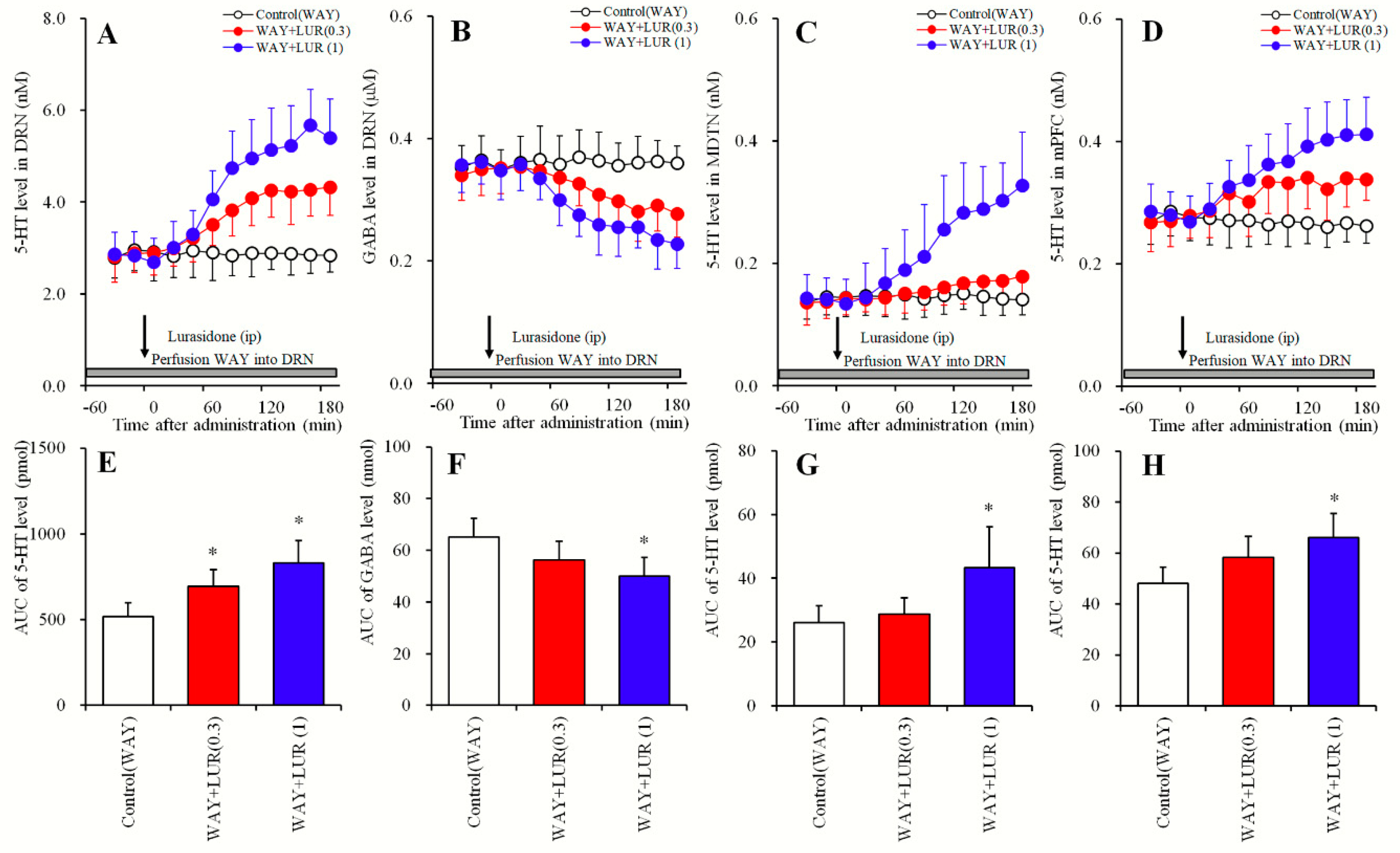
2.2. Dose-Dependent Effects of Systemic Lurasidone Administration on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC and GABA in the DRN Following Inhibition of 5-HT1AR in the DRN (Study_2)
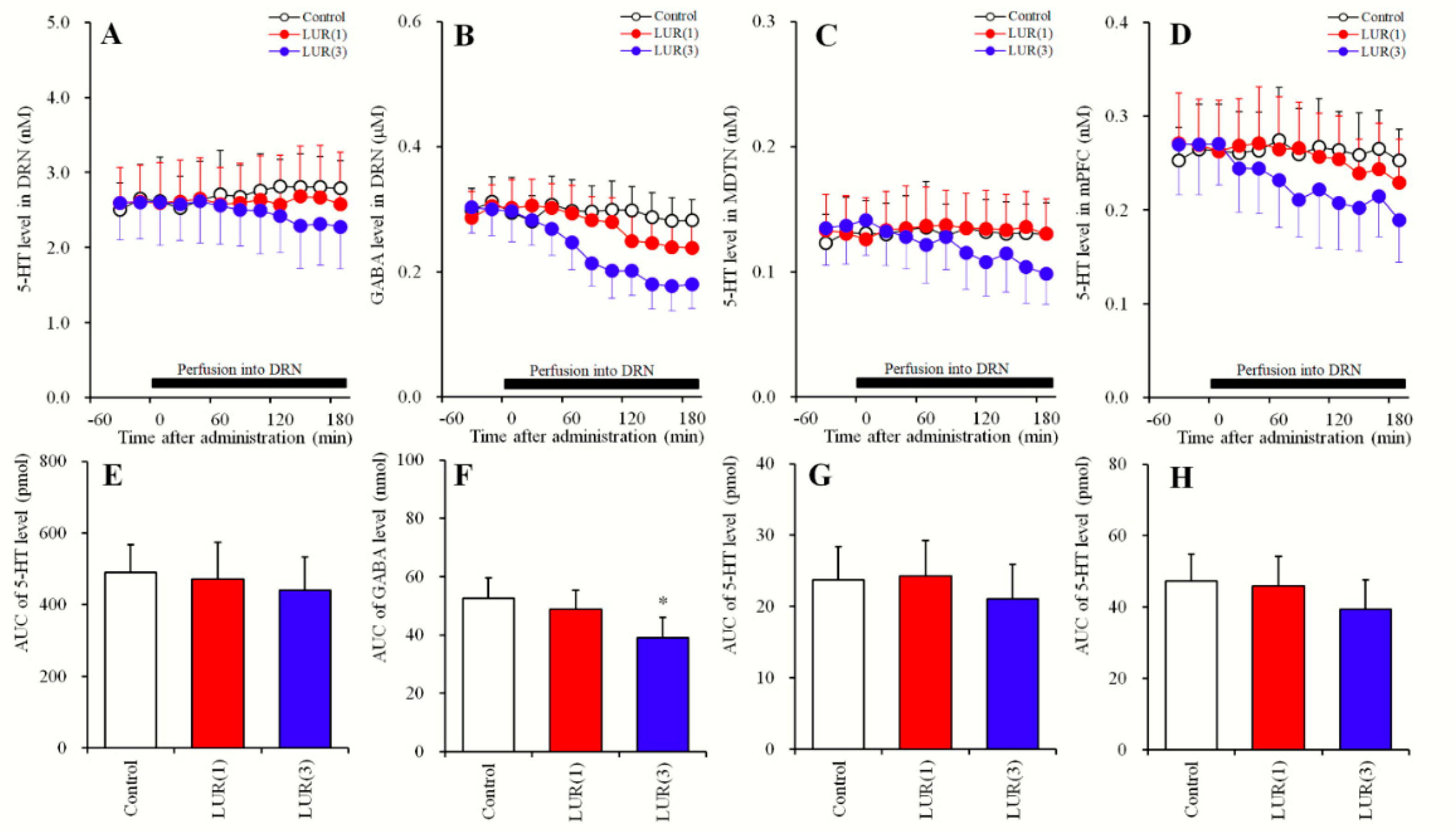
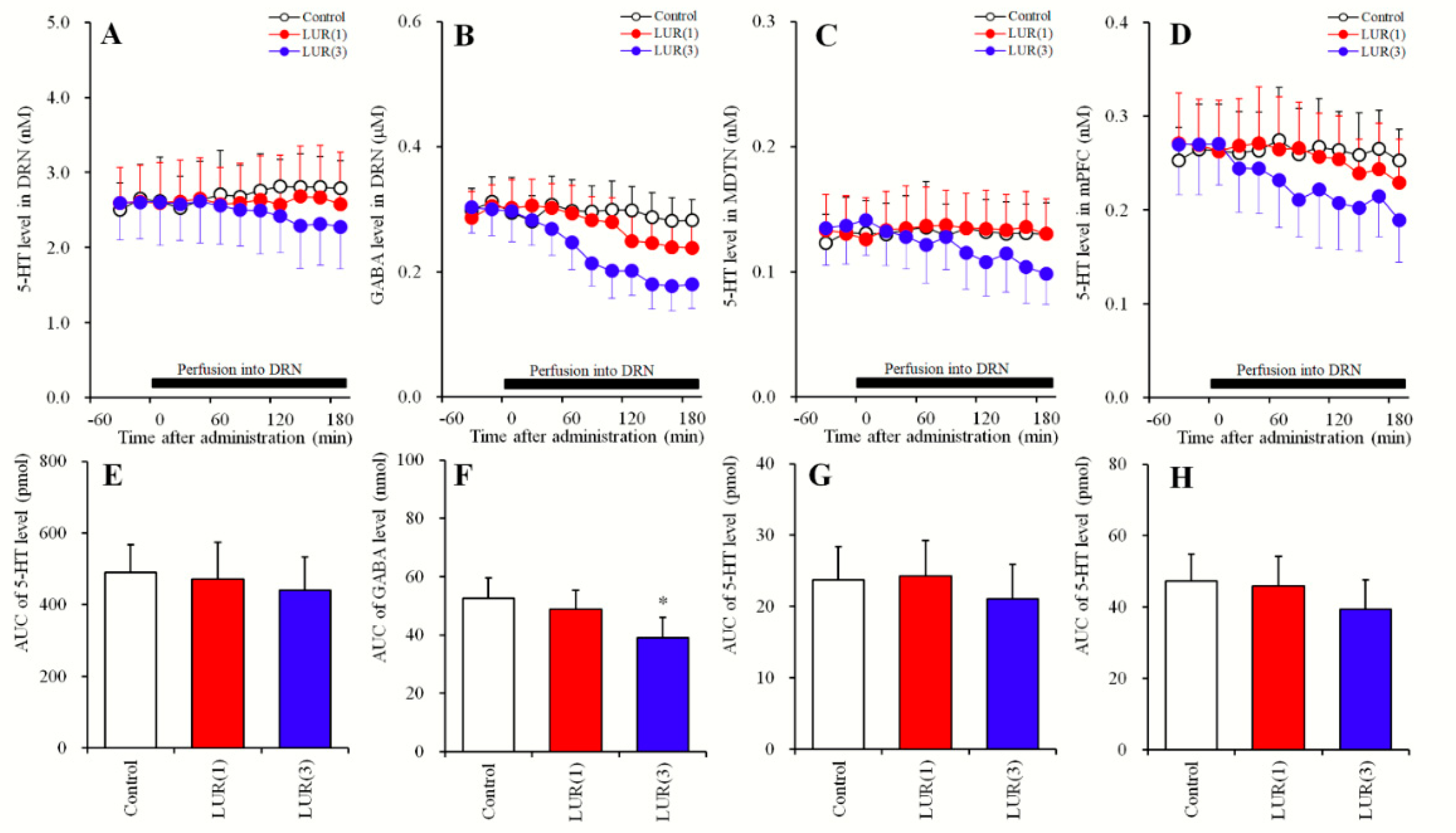
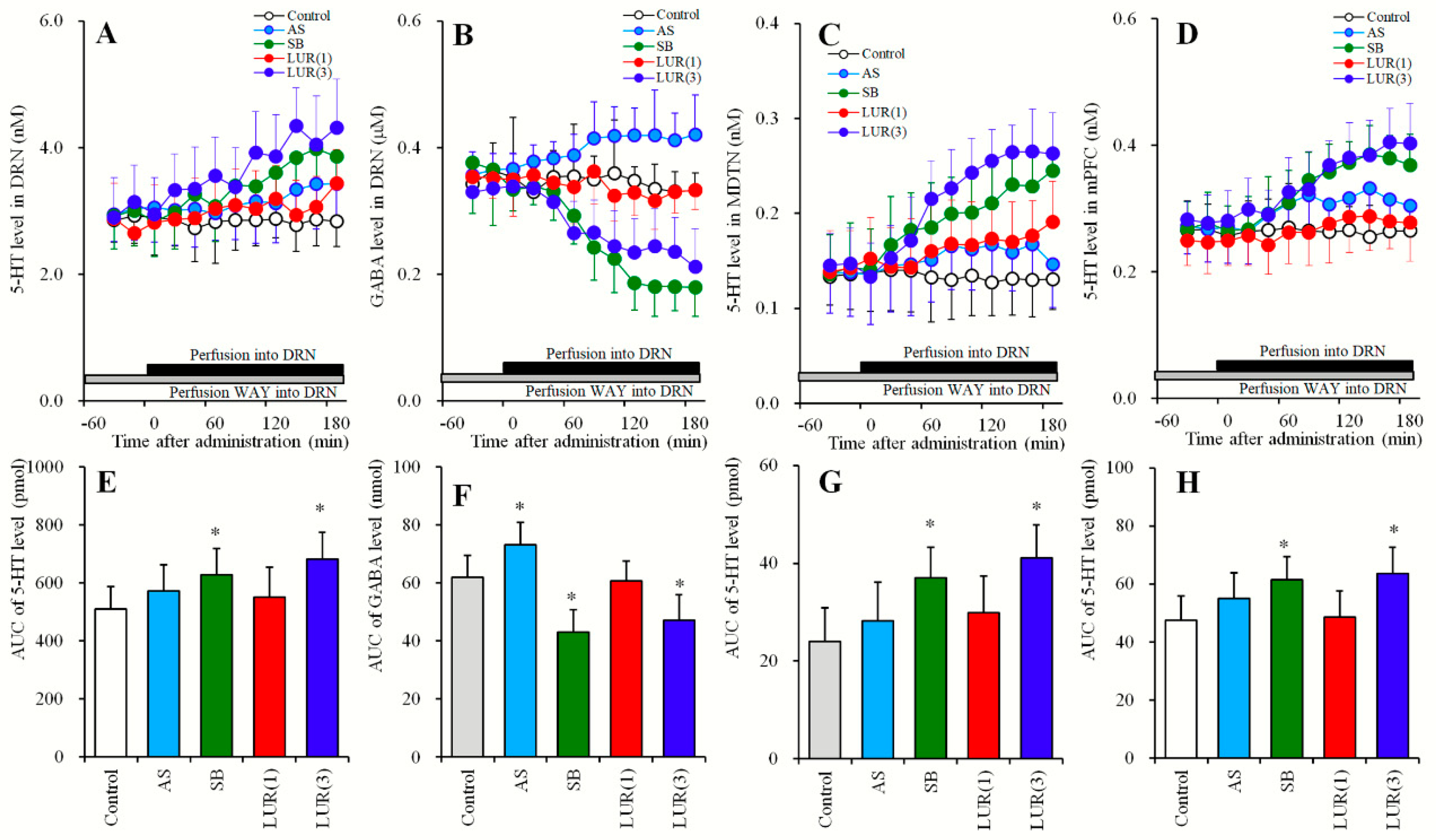
2.3. Concentration-Dependent Effects of Local Lurasidone Administrations into the DRN on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC and GABA in the DRN (Study_3)
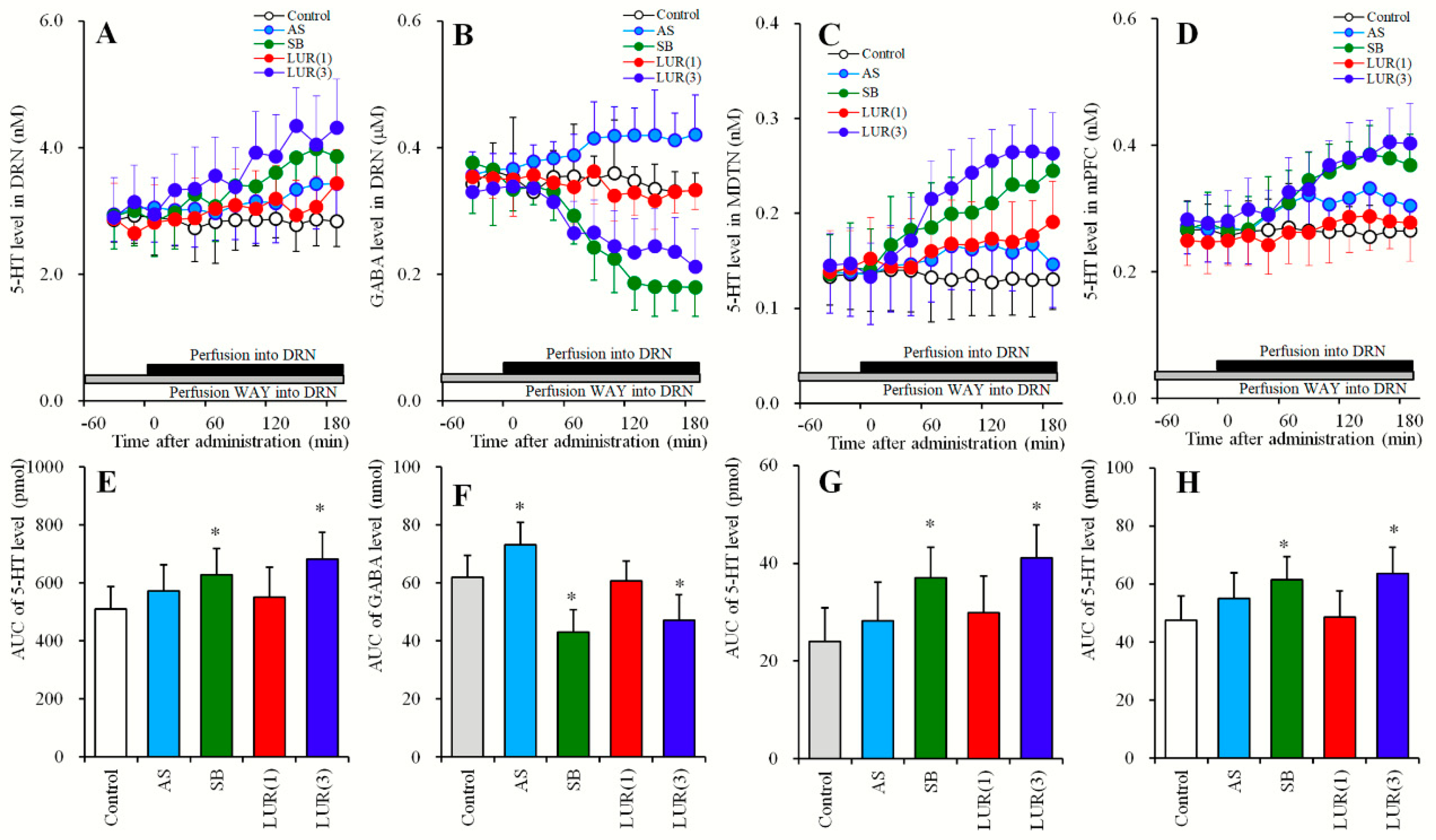
2.3.1. Concentration-Dependent Effects of Local Lurasidone Administrations into the DRN on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC and GABA in the DRN (Study_3-1)
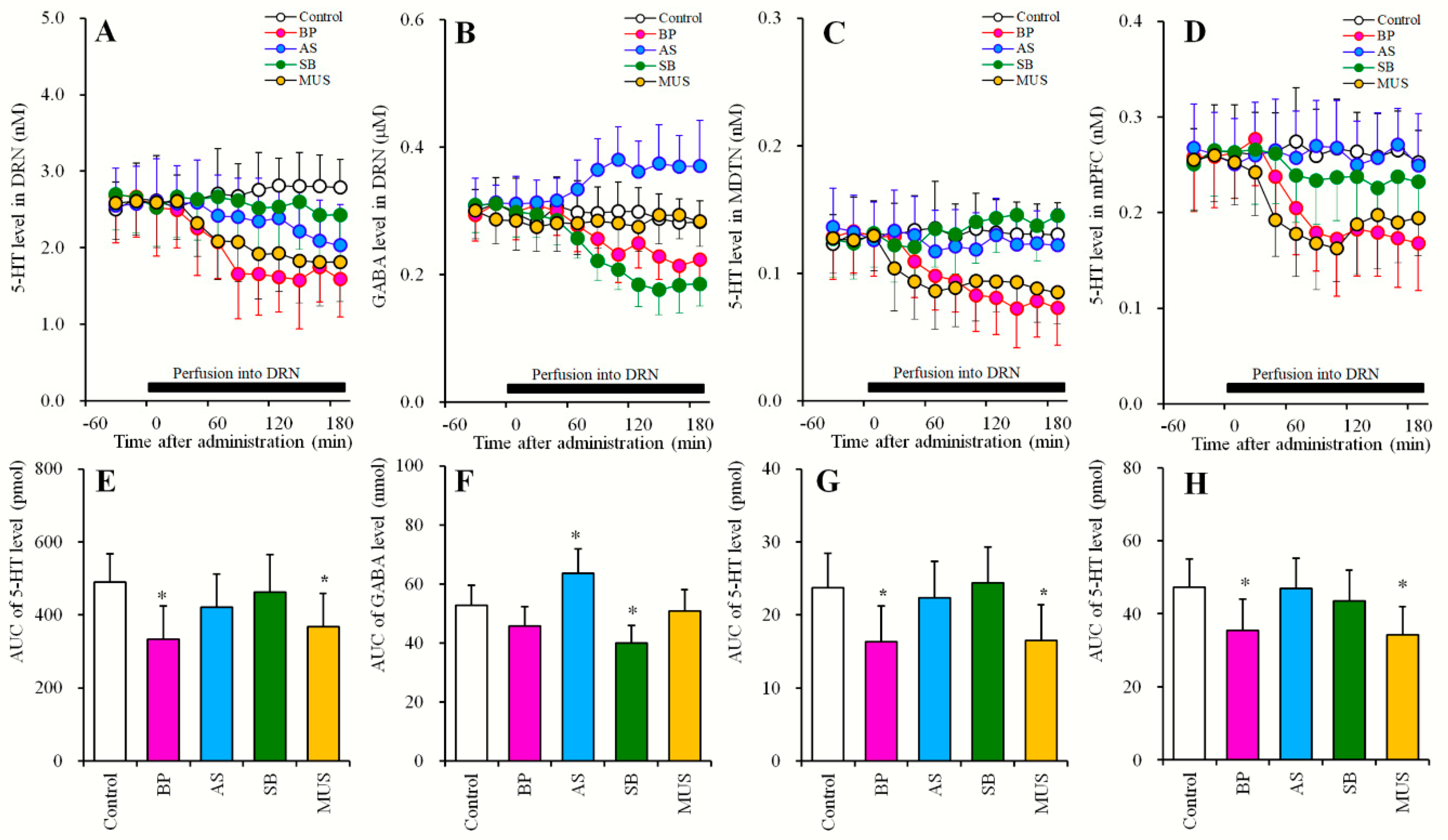
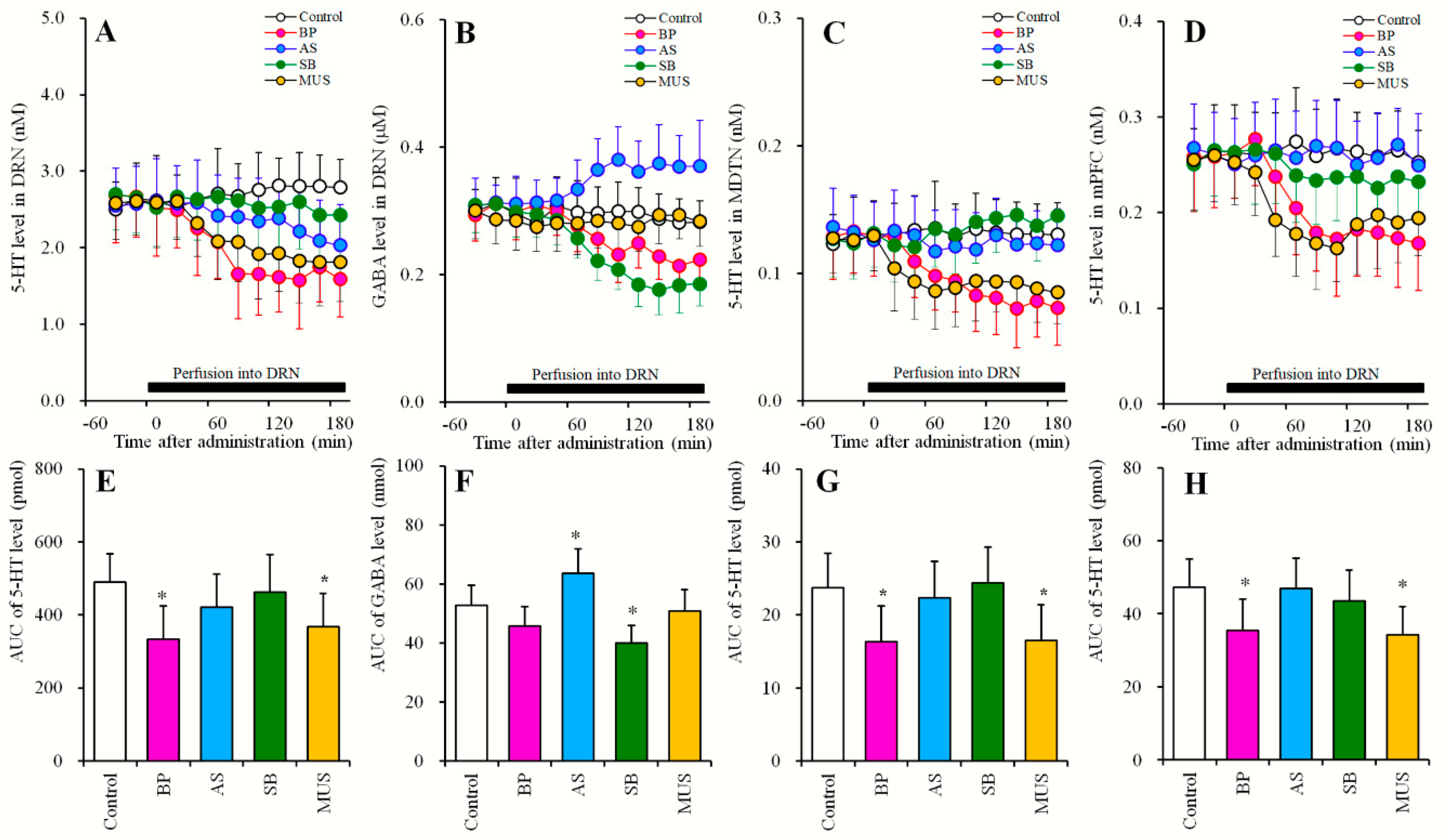
2.3.2. Effects of Local Administrations of BP554, AS19, SB269970 and Muscimol into the DRN on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC, and GABA in the DRN (Study_3-2)
2.4. Effects of Local Administrations of AS19, SB269970 and Lurasidone into the DRN on Extracellular Levels of 5-HT in the DRN, MDTN, mPFC, and GABA in the DRN Following Inhibition of 5-HT1AR in the DRN (Study_4)
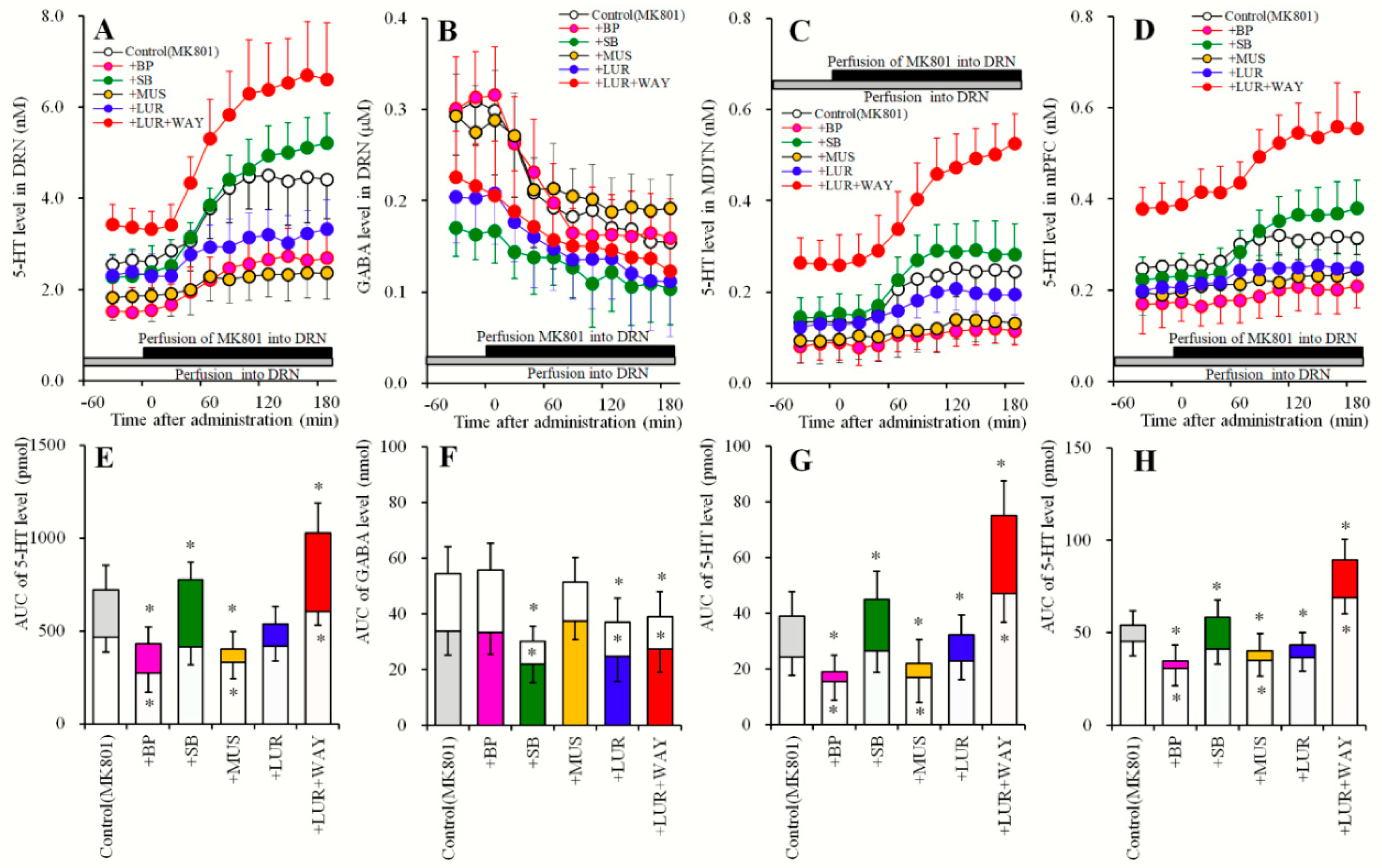
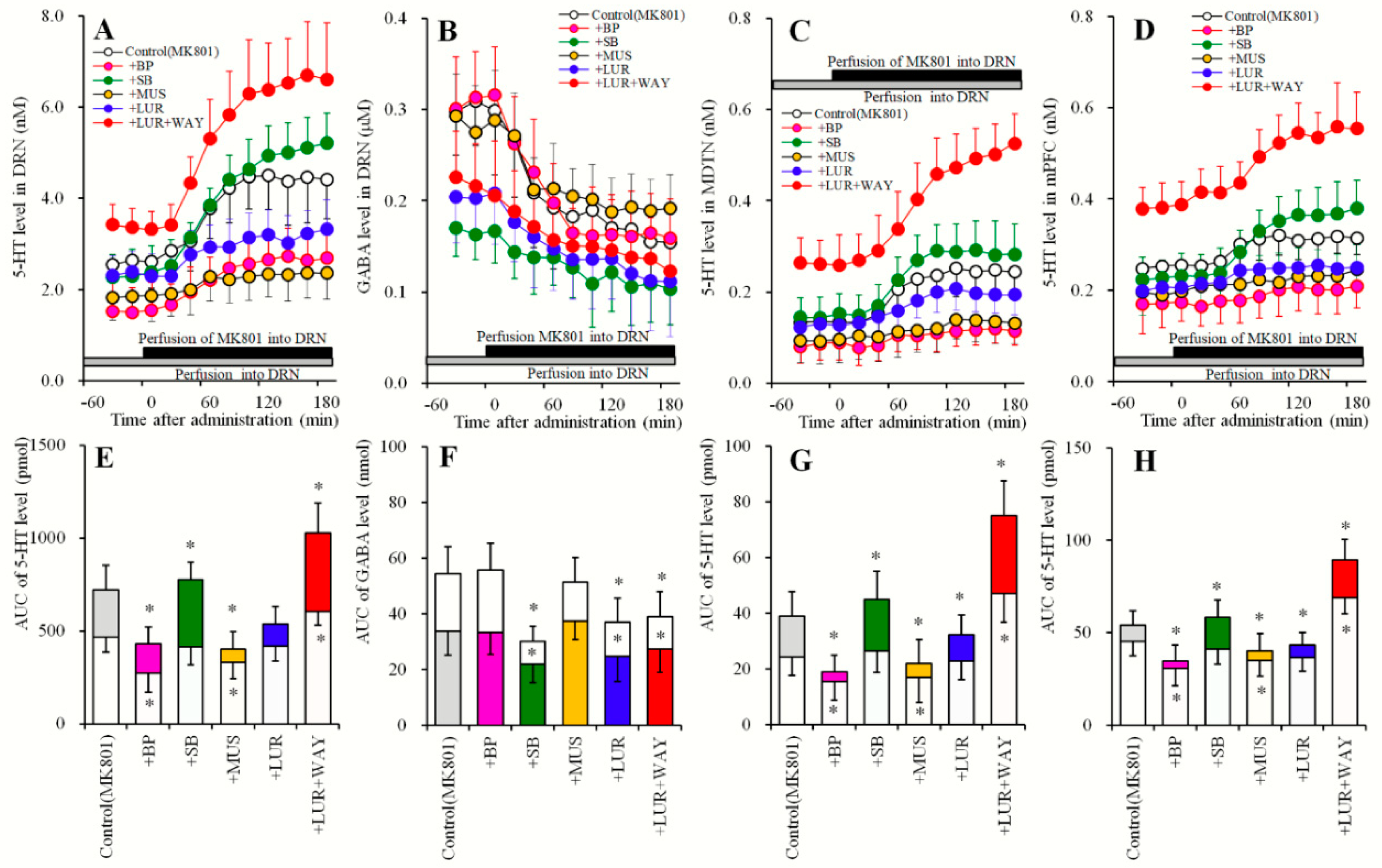
2.5. Effects of Local Administrations of BP554, SB269970, and Muscimol in the DRN on Changes in Extracellular Levels of 5-HT in the DRN, MDTN, mPFC and GABA in the DRN (Study_5)
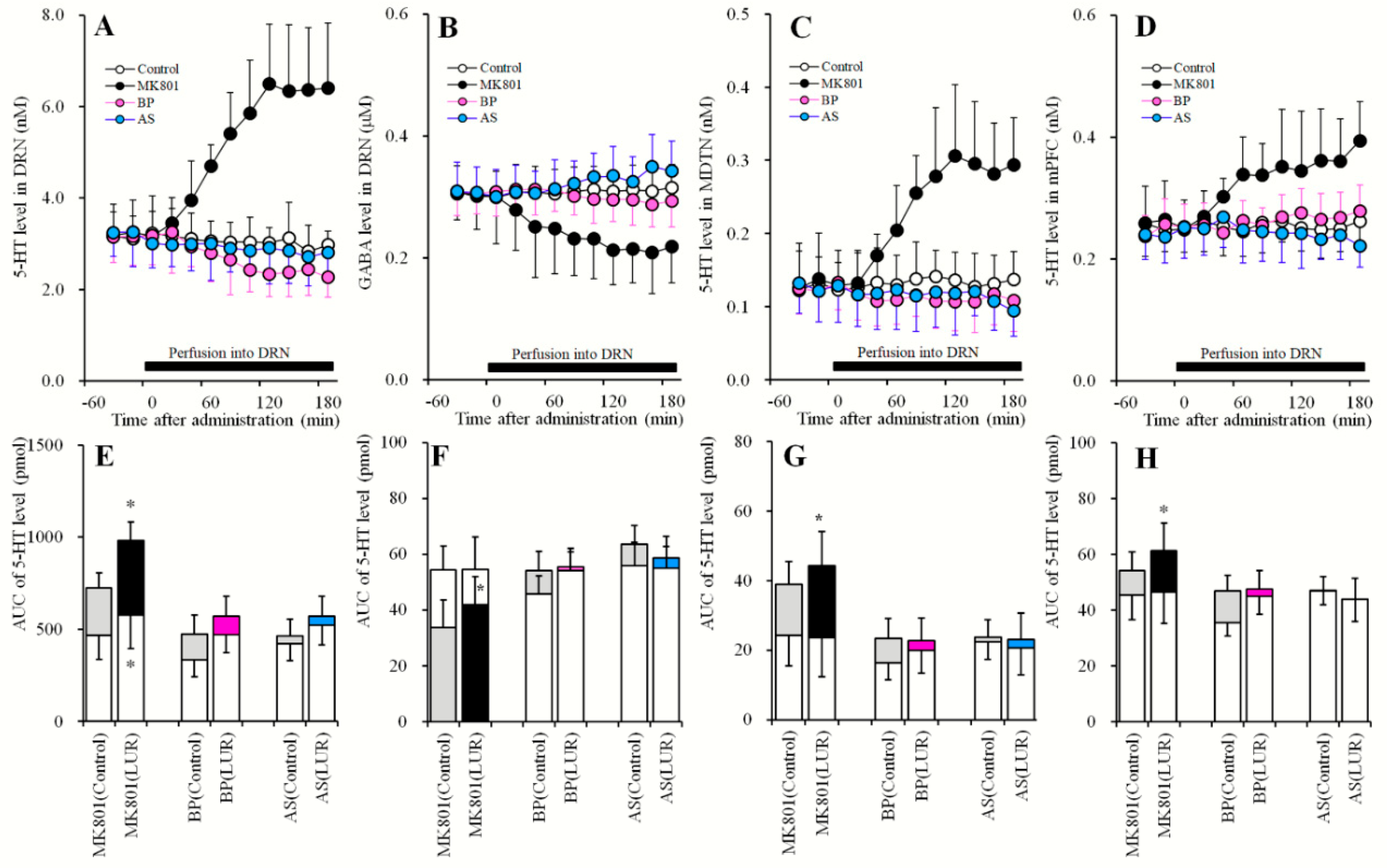
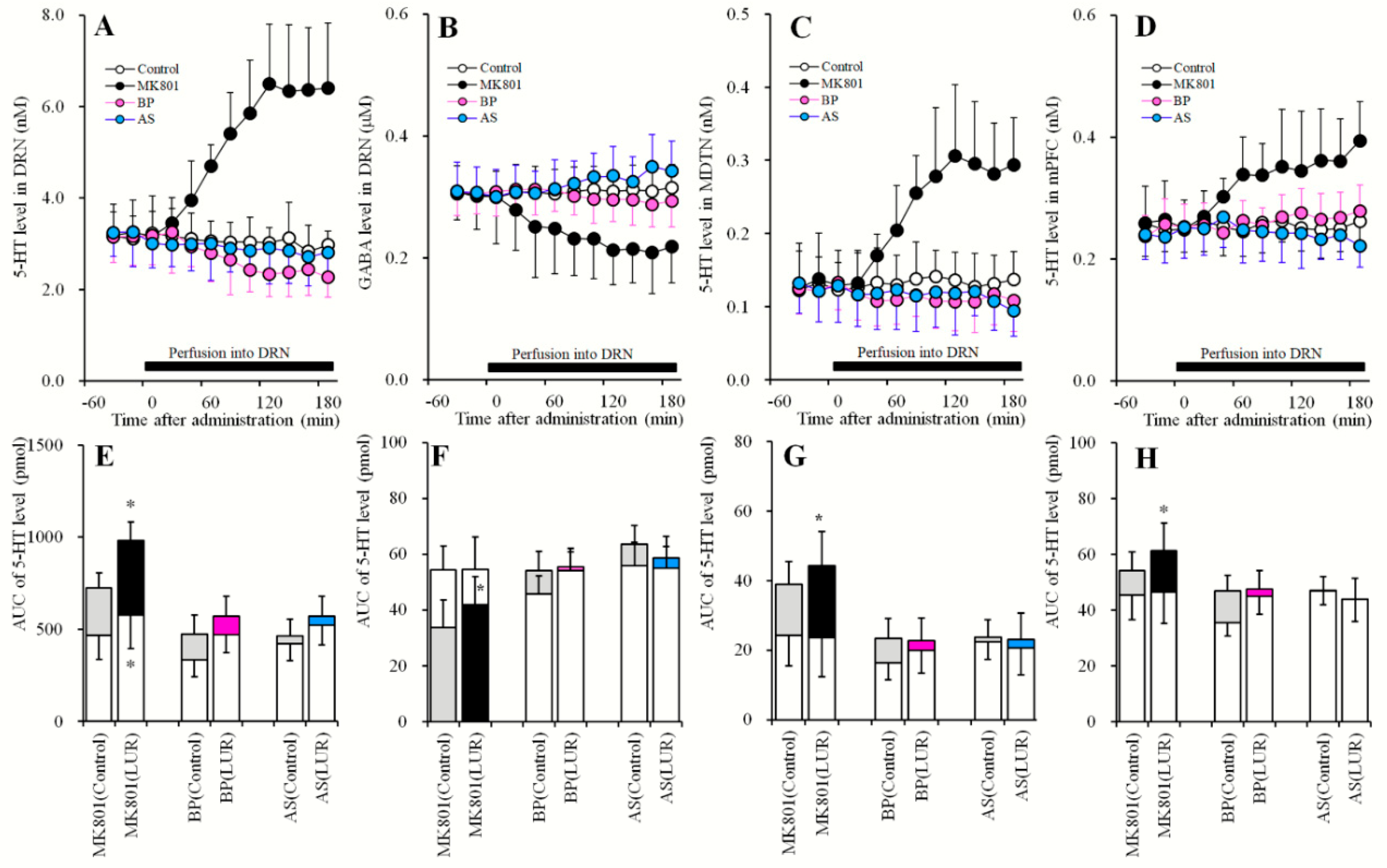
2.6. Effects of Local Administration of MK801, AS19 and BP554 in the DRN on 5-HT Release in the DRN, MDTN, mPFC, and GABA in the DRN After Sub-Chronic Administration of Lurasidone (Study_6)
3. Discussion
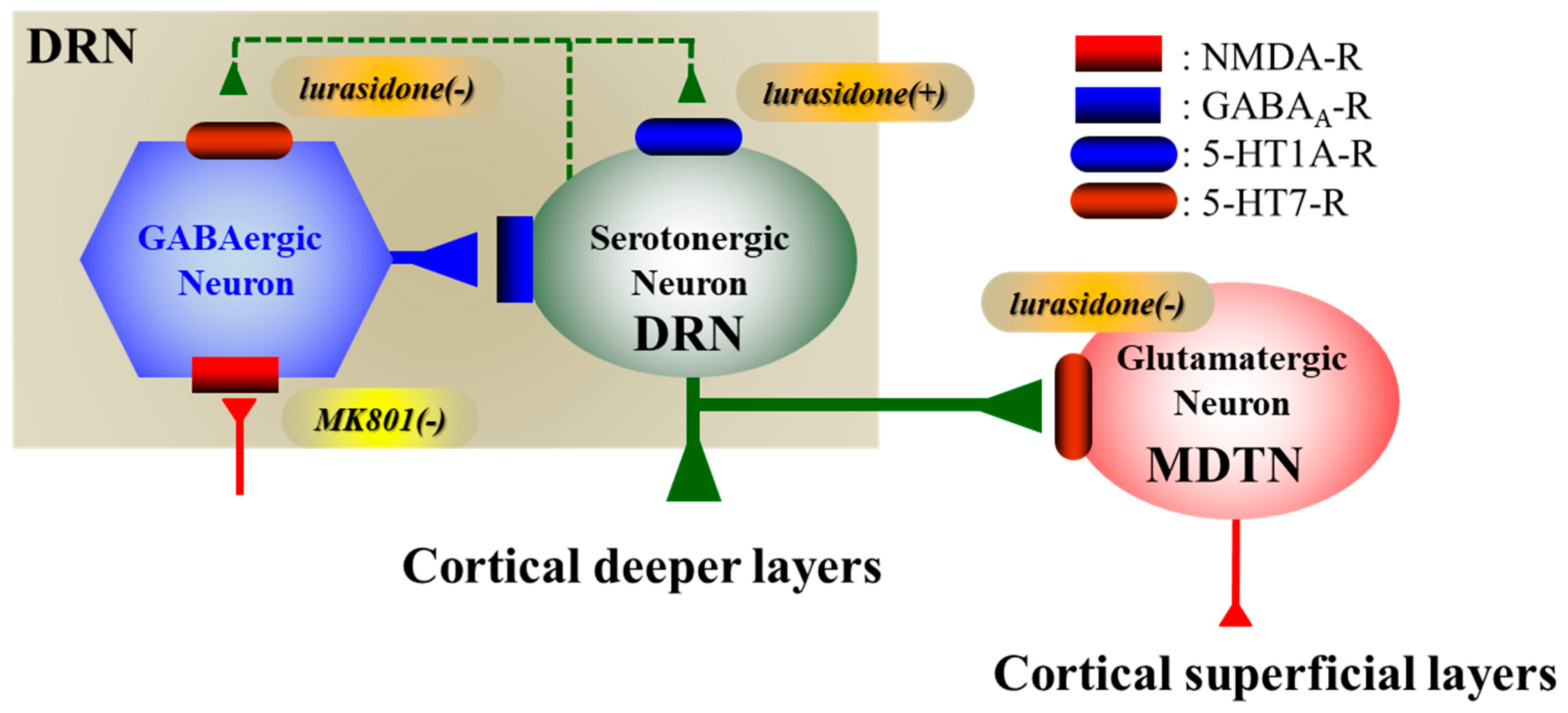
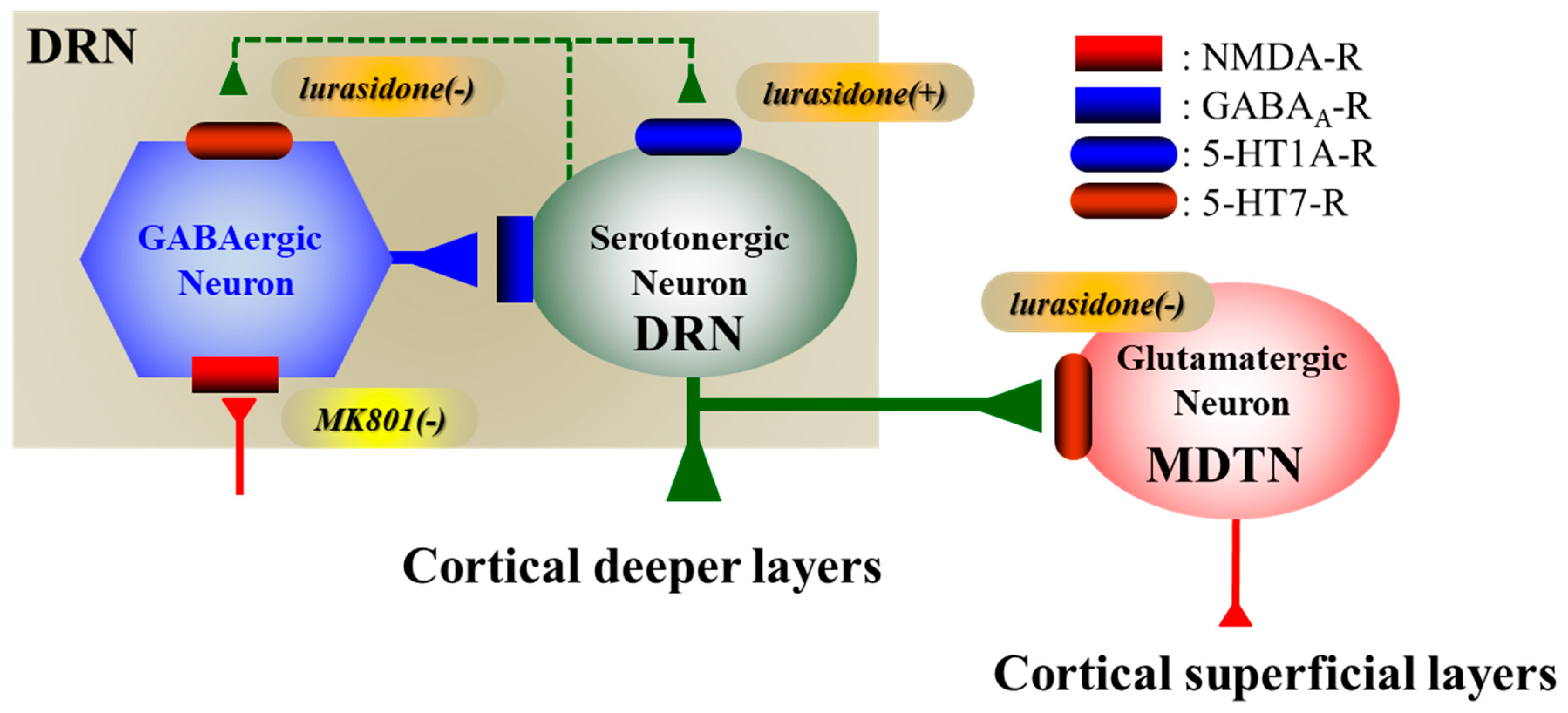
3.1. Regulatory Mechanisms of Serotonergic Transmission Associated with the DRN
3.2. 5-HT1AR-Assoicated Mechanisms of Lurasidone
3.3. 5-HT7R-Assoicated Mechanisms of Lurasidone
4. Materials and Methods
4.1. Preparation of the Microdialysis System
4.2. Determination of Extracellular GABA and 5-HT Levels
4.3. Data Analysis
4.4. Chemical Agents
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lisman, J.E.; Coyle, J.T.; Green, R.W.; Javitt, D.C.; Benes, F.M.; Heckers, S.; Grace, A.A. Circuit-based framework for understanding neurotransmitter and risk gene interactions in schizophrenia. Trends Neurosci. 2008, 31, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conn, P.J.; Lindsley, C.W.; Jones, C.K. Activation of metabotropic glutamate receptors as a novel approach for the treatment of schizophrenia. Trends Pharm. Sci. 2009, 30, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meltzer, H.Y. Serotonergic mechanisms as targets for existing and novel antipsychotics. In Handbook of Experimental Pharmacology; Springer: New York, NY, USA, 2012; pp. 87–124. [Google Scholar] [CrossRef]
- Svob Strac, D.; Pivac, N.; Muck-Seler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, S.; Ohoyama, K.; Hamaguchi, T.; Kashimoto, K.; Nakagawa, M.; Kanehara, S.; Suzuki, D.; Matsumoto, T.; Motomura, E.; Shiroyama, T.; et al. Effects of quetiapine on monoamine, GABA, and glutamate release in rat prefrontal cortex. Psychopharmacol. (Berl) 2009, 206, 243–258. [Google Scholar] [CrossRef]
- Yamamura, S.; Ohoyama, K.; Hamaguchi, T.; Nakagawa, M.; Suzuki, D.; Matsumoto, T.; Motomura, E.; Tanii, H.; Shiroyama, T.; Okada, M. Effects of zotepine on extracellular levels of monoamine, GABA and glutamate in rat prefrontal cortex. Br. J. Pharmacol. 2009, 157, 656–665. [Google Scholar] [CrossRef]
- Tanahashi, S.; Yamamura, S.; Nakagawa, M.; Motomura, E.; Okada, M. Clozapine, but not haloperidol, enhances glial D-serine and L-glutamate release in rat frontal cortex and primary cultured astrocytes. Br. J. Pharmacol. 2012, 165, 1543–1555. [Google Scholar] [CrossRef]
- Tanahashi, S.; Yamamura, S.; Nakagawa, M.; Motomura, E.; Okada, M. Dopamine D2 and serotonin 5-HT1A receptors mediate the actions of aripiprazole in mesocortical and mesoaccumbens transmission. Neuropharmacology 2012, 62, 765–774. [Google Scholar] [CrossRef]
- Fukuyama, K.; Hasegawa, T.; Okada, M. Cystine/Glutamate Antiporter and Aripiprazole Compensate NMDA Antagonist-Induced Dysfunction of Thalamocortical L-Glutamatergic Transmission. Int. J. Mol. Sci. 2018, 19, 3645. [Google Scholar] [CrossRef]
- Ohoyama, K.; Yamamura, S.; Hamaguchi, T.; Nakagawa, M.; Motomura, E.; Shiroyama, T.; Tanii, H.; Okada, M. Effect of novel atypical antipsychotic, blonanserin, on extracellular neurotransmitter level in rat prefrontal cortex. Eur. J. Pharmacol. 2011, 653, 47–57. [Google Scholar] [CrossRef]
- Okada, M.; Fukuyama, K.; Ueda, Y. Lurasidone inhibits NMDA/glutamate antagonist-induced functional abnormality of thalamocortical glutamatergic transmission via 5-HT7 receptor blockade. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Fukuyama, K.; Kato, R.; Murata, M.; Shiroyama, T.; Okada, M. Clozapine Normalizes a Glutamatergic Transmission Abnormality Induced by an Impaired NMDA Receptor in the Thalamocortical Pathway via the Activation of a Group III Metabotropic Glutamate Receptor. Biomolecules 2019, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, T.; Horisawa, T.; Tokuda, K.; Ishiyama, T.; Ogasa, M.; Tagashira, R.; Matsumoto, K.; Nishikawa, H.; Ueda, Y.; Toma, S.; et al. Pharmacological profile of lurasidone, a novel antipsychotic agent with potent 5-hydroxytryptamine 7 (5-HT7) and 5-HT1A receptor activity. J. Pharm. Exp. 2010, 334, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Pompili, M.; Verzura, C.; Trovini, G.; Buscajoni, A.; Falcone, G.; Naim, S.; Nardella, A.; Sorice, S.; Baldessarini, R.J.; Girardi, P. Lurasidone: Efficacy and safety in the treatment of psychotic and mood disorders. Expert Opin. Drug Saf. 2018, 17, 197–205. [Google Scholar] [CrossRef]
- Yatham, L.N.; Kennedy, S.H.; Parikh, S.V.; Schaffer, A.; Bond, D.J.; Frey, B.N.; Sharma, V.; Goldstein, B.I.; Rej, S.; Beaulieu, S.; et al. Canadian Network for Mood and Anxiety Treatments (CANMAT) and International Society for Bipolar Disorders (ISBD) 2018 guidelines for the management of patients with bipolar disorder. Bipolar Disord. 2018, 20, 97–170. [Google Scholar] [CrossRef]
- Bonaventure, P.; Kelly, L.; Aluisio, L.; Shelton, J.; Lord, B.; Galici, R.; Miller, K.; Atack, J.; Lovenberg, T.W.; Dugovic, C. Selective blockade of 5-hydroxytryptamine (5-HT)7 receptors enhances 5-HT transmission, antidepressant-like behavior, and rapid eye movement sleep suppression induced by citalopram in rodents. J. Pharm. Exp. 2007, 321, 690–698. [Google Scholar] [CrossRef]
- Huang, M.; Kwon, S.; Rajagopal, L.; He, W.; Meltzer, H.Y. 5-HT1A parital agonism and 5-HT7 antagonism restore episodic memory in subchronic phencyclidine-treated mice: Role of brain glutamate, dopamine, acetylcholine and GABA. Psychopharmacol. (Berl) 2018, 235, 2795–2808. [Google Scholar] [CrossRef]
- Fukuyama, K.; Tanahashi, S.; Hamaguchi, T.; Nakagawa, M.; Shiroyama, T.; Motomura, E.; Okada, M. Differential mechanisms underlie the regulation of serotonergic transmission in the dorsal and median raphe nuclei by mirtazapine: A dual probe microdialysis study. Psychopharmacol. (Berl) 2013, 229, 617–626. [Google Scholar] [CrossRef]
- Gocho, Y.; Sakai, A.; Yanagawa, Y.; Suzuki, H.; Saitow, F. Electrophysiological and pharmacological properties of GABAergic cells in the dorsal raphe nucleus. J. Physiol. Sci. 2013, 63, 147–154. [Google Scholar] [CrossRef]
- Huang, M.; Kwon, S.; He, W.; Meltzer, H.Y. Neurochemical arguments for the use of dopamine D4 receptor stimulation to improve cognitive impairment associated with schizophrenia. Pharm. Biochem. Behav. 2017, 157, 16–23. [Google Scholar] [CrossRef]
- Bosker, F.J.; Folgering, J.H.; Gladkevich, A.V.; Schmidt, A.; van der Hart, M.C.; Sprouse, J.; den Boer, J.A.; Westerink, B.H.; Cremers, T.I. Antagonism of 5-HT(1A) receptors uncovers an excitatory effect of SSRIs on 5-HT neuronal activity, an action probably mediated by 5-HT(7) receptors. J. Neurochem. 2009, 108, 1126–1135. [Google Scholar] [CrossRef]
- Yamamura, S.; Abe, M.; Nakagawa, M.; Ochi, S.; Ueno, S.; Okada, M. Different actions for acute and chronic administration of mirtazapine on serotonergic transmission associated with raphe nuclei and their innervation cortical regions. Neuropharmacology 2011, 60, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.S.; Kim, S.J.; Lee, J.; Ha, D.J.; Baek, M.; Moon, H. Quantification of lurasidone, an atypical antipsychotic drug, in rat plasma with high-performance liquid chromatography with tandem mass spectrometry. Biomed. Chromatogr. 2011, 25, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Javitt, D.C. Negative schizophrenic symptomatology and the PCP (phencyclidine) model of schizophrenia. Hillside J. Clin. Psychiatry 1987, 9, 12–35. [Google Scholar] [PubMed]
- Lieberman, J.A.; Bymaster, F.P.; Meltzer, H.Y.; Deutch, A.Y.; Duncan, G.E.; Marx, C.E.; Aprille, J.R.; Dwyer, D.S.; Li, X.M.; Mahadik, S.P.; et al. Antipsychotic drugs: Comparison in animal models of efficacy, neurotransmitter regulation, and neuroprotection. Pharm. Rev. 2008, 60, 358–403. [Google Scholar] [CrossRef]
- Lavoie, B.; Parent, A. Serotoninergic innervation of the thalamus in the primate: An immunohistochemical study. J. Comp. Neurol. 1991, 312, 1–18. [Google Scholar] [CrossRef]
- Imai, H.; Steindler, D.A.; Kitai, S.T. The organization of divergent axonal projections from the midbrain raphe nuclei in the rat. J. Comp. Neurol. 1986, 243, 363–380. [Google Scholar] [CrossRef]
- Sharp, T.; Boothman, L.; Raley, J.; Queree, P. Important messages in the ‘post’: Recent discoveries in 5-HT neurone feedback control. Trends Pharm. Sci. 2007, 28, 629–636. [Google Scholar] [CrossRef]
- Chemel, B.R.; Roth, B.L.; Armbruster, B.; Watts, V.J.; Nichols, D.E. WAY-100635 is a potent dopamine D4 receptor agonist. Psychopharmacol. (Berl) 2006, 188, 244–251. [Google Scholar] [CrossRef]
- Murai, T.; Nakako, T.; Ikeda, K.; Ikejiri, M.; Ishiyama, T.; Taiji, M. Lack of dopamine D4 receptor affinity contributes to the procognitive effect of lurasidone. Behav. Brain Res. 2014, 261, 26–30. [Google Scholar] [CrossRef]
- Ishiyama, T.; Tokuda, K.; Ishibashi, T.; Ito, A.; Toma, S.; Ohno, Y. Lurasidone (SM-13496), a novel atypical antipsychotic drug, reverses MK-801-induced impairment of learning and memory in the rat passive-avoidance test. Eur. J. Pharmacol. 2007, 572, 160–170. [Google Scholar] [CrossRef]
- Richardson-Jones, J.W.; Craige, C.P.; Guiard, B.P.; Stephen, A.; Metzger, K.L.; Kung, H.F.; Gardier, A.M.; Dranovsky, A.; David, D.J.; Beck, S.G.; et al. 5-HT1A autoreceptor levels determine vulnerability to stress and response to antidepressants. Neuron 2010, 65, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, S. (-)-Pindolol, but not buspirone, potentiates the citalopram-induced rise in extracellular 5-hydroxytryptamine. Eur. J. Pharmacol. 1996, 303, 183–186. [Google Scholar] [CrossRef]
- Assie, M.B.; Lomenech, H.; Ravailhe, V.; Faucillon, V.; Newman-Tancredi, A. Rapid desensitization of somatodendritic 5-HT1A receptors by chronic administration of the high-efficacy 5-HT1A agonist, F13714: A microdialysis study in the rat. Br. J. Pharmacol. 2006, 149, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Riad, M.; Garcia, S.; Watkins, K.C.; Jodoin, N.; Doucet, E.; Langlois, X.; el Mestikawy, S.; Hamon, M.; Descarries, L. Somatodendritic localization of 5-HT1A and preterminal axonal localization of 5-HT1B serotonin receptors in adult rat brain. J. Comp. Neurol. 2000, 417, 181–194. [Google Scholar] [CrossRef]
- US Food and Drug Administration. FDA Approves New Nasal Spray Medication for Treatment-Resistant Depression; Available Only at a Certified Doctor’s Office or Clinic. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-new-nasal-spray- medication-treatment-resistant-depression-available-only-certified (accessed on 5 March 2019).
- Leon, A.C.; Fiedorowicz, J.G.; Solomon, D.A.; Li, C.; Coryell, W.H.; Endicott, J.; Fawcett, J.; Keller, M.B. Risk of suicidal behavior with antidepressants in bipolar and unipolar disorders. J. Clin. Psychiatry 2014, 75, 720–727. [Google Scholar] [CrossRef]
- Aan Het Rot, M.; Zarate, C.A., Jr.; Charney, D.S.; Mathew, S.J. Ketamine for depression: Where do we go from here? Biol. Psychiatry 2012, 72, 537–547. [Google Scholar] [CrossRef]
- DiazGranados, N.; Ibrahim, L.A.; Brutsche, N.E.; Ameli, R.; Henter, I.D.; Luckenbaugh, D.A.; Machado-Vieira, R.; Zarate, C.A., Jr. Rapid resolution of suicidal ideation after a single infusion of an N-methyl-D-aspartate antagonist in patients with treatment-resistant major depressive disorder. J. Clin. Psychiatry 2010, 71, 1605–1611. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238. [Google Scholar] [CrossRef]
- Hashimoto, K. Rapid-acting antidepressant ketamine, its metabolites and other candidates: A historical overview and future perspective. Psychiatry Clin. Neurosci. 2019. [Google Scholar] [CrossRef]
- Okada, M.; Fukuyama, K.; Kawano, Y.; Shiroyama, T.; Ueda, Y. Memantine protects thalamocortical hyper-glutamatergic transmission induced by NMDA receptor antagonism via activation of system xc−. Pharm. Res. Perspect. 2019, 7, e00457. [Google Scholar] [CrossRef]
- Nakano, T.; Hasegawa, T.; Suzuki, D.; Motomura, E.; Okada, M. Amantadine Combines Astroglial System Xc(-) Activation with Glutamate/NMDA Receptor Inhibition. Biomolecules 2019, 9, 191. [Google Scholar] [CrossRef]
- Andressen, K.W.; Manfra, O.; Brevik, C.H.; Ulsund, A.H.; Vanhoenacker, P.; Levy, F.O.; Krobert, K.A. The atypical antipsychotics clozapine and olanzapine promote down-regulation and display functional selectivity at human 5-HT7 receptors. Br. J. Pharmacol. 2015, 172, 3846–3860. [Google Scholar] [CrossRef]
- Okada, M.; Fukuyama, K.; Kawano, Y.; Shiroyama, T.; Suzuki, D.; Ueda, Y. Effects of Acute and Sub-chronic Administrations of Guanfacine on Catecholaminergic Transmissions in the Orbitofrontal Cortex. Neuropharmacology 2019. [Google Scholar] [CrossRef]
- Vertes, R.P.; Linley, S.B.; Hoover, W.B. Limbic circuitry of the midline thalamus. Neurosci Biobehav Rev. 2015, 54, 89–107. [Google Scholar] [CrossRef] [Green Version]
- McGrath, J.C.; Drummond, G.B.; McLachlan, E.M.; Kilkenny, C.; Wainwright, C.L. Guidelines for reporting experiments involving animals: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1573–1576. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain: In Stereotoxic Coordinates, 6th ed.; Academic Press: San Dieg, CA, USA, 2007. [Google Scholar]
- Okada, M.; Yoshida, S.; Zhu, G.; Hirose, S.; Kaneko, S. Biphasic actions of topiramate on monoamine exocytosis associated with both soluble N-ethylmaleimide-sensitive factor attachment protein receptors and Ca(2+)-induced Ca(2+)-releasing systems. Neuroscience 2005, 134, 233–246. [Google Scholar] [CrossRef]
- Yamamura, S.; Hoshikawa, M.; Dai, K.; Saito, H.; Suzuki, N.; Niwa, O.; Okada, M. ONO-2506 inhibits spike-wave discharges in a genetic animal model without affecting traditional convulsive tests via gliotransmission regulation. Br. J. Pharmacol. 2013, 168, 1088–1100. [Google Scholar] [CrossRef]
- Fukuyama, K.; Tanahashi, S.; Hoshikawa, M.; Shinagawa, R.; Okada, M. Zonisamide regulates basal ganglia transmission via astroglial kynurenine pathway. Neuropharmacology 2014, 76, 137–145. [Google Scholar] [CrossRef]
- Okada, M.; Hirano, T.; Mizuno, K.; Kawata, Y.; Wada, K.; Murakami, T.; Tasaki, H.; Kaneko, S. Effects of carbamazepine on hippocampal serotonergic system. Epilepsy Res. 1998, 31, 187–198. [Google Scholar] [CrossRef]
- Kawata, Y.; Okada, M.; Murakami, T.; Kamata, A.; Zhu, G.; Kaneko, S. Pharmacological discrimination between effects of carbamazepine on hippocampal basal, Ca(2+)- and K(+)-evoked serotonin release. Br. J. Pharmacol. 2001, 133, 557–567. [Google Scholar] [CrossRef]
- Tanahashi, S.; Yamamura, S.; Nakagawa, M.; Motomura, E.; Okada, M. Effect of lamotrigine and carbamazepine on corticotropin-releasing factor-associated serotonergic transmission in rat dorsal raphe nucleus. Psychopharmacol. (Berl) 2012, 220, 599–610. [Google Scholar] [CrossRef]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental design and analysis and their reporting II: Updated and simplified guidance for authors and peer reviewers. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Administration Root (Target Agent) | TARGET AGENT | Pre-Treatment Roots | Pre-Treatment Agents | Reference | |
|---|---|---|---|---|---|
| Study_1 | Acute systemic (i.p.) | LUR (0.3 and 1 mg/kg) | - | - | [11,17,20] |
| Study_2 | Acute systemic (i.p.) | LUR (0.3 and 1 mg/kg) | Local (DRN) | WAY (10 μM) | [11,17,20] |
| Study_3 | Local (DRN) | LUR (1 and 3 μM) BP (50 μM) AS (1 μM) SB (10 μM) Muscimol (1 μM) | - | - | [11] [8] [21] [11] [6] |
| Study_4 | Local (DRN) | LUR (1 and 3 μM) AS (1 μM) SB (10 μM) | Local (DRN) | WAY (10 μM) | [11,22] [21] [11] |
| Study_5 | Local (DRN) | MK801 (5 μM) | Local (DRN) | LUR (3 μM) LUR (3 μM) + WAY (10 μM) BP (50 μM) SB (10 μM) Muscimol (1 μM) | [11,12] [11] [8] [11] [6] |
| Study_6 | Local (DRN) | MK801 (5 μM) BP (50 μM) AS (1 μM) | Chronic systemic (s.c.) | LUR (3mg/kg/day) for 7 days | [12] [17] [20] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, M.; Fukuyama, K.; Okubo, R.; Shiroyama, T.; Ueda, Y. Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus. Pharmaceuticals 2019, 12, 149. https://doi.org/10.3390/ph12040149
Okada M, Fukuyama K, Okubo R, Shiroyama T, Ueda Y. Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus. Pharmaceuticals. 2019; 12(4):149. https://doi.org/10.3390/ph12040149
Chicago/Turabian StyleOkada, Motohiro, Kouji Fukuyama, Ruri Okubo, Takashi Shiroyama, and Yuto Ueda. 2019. "Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus" Pharmaceuticals 12, no. 4: 149. https://doi.org/10.3390/ph12040149
APA StyleOkada, M., Fukuyama, K., Okubo, R., Shiroyama, T., & Ueda, Y. (2019). Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus. Pharmaceuticals, 12(4), 149. https://doi.org/10.3390/ph12040149