Synthetic Inhibitors of Snake Venom Enzymes: Thioesters Derived from 2-Sulfenyl Ethylacetate


Abstract
:1. Introduction
2. Results
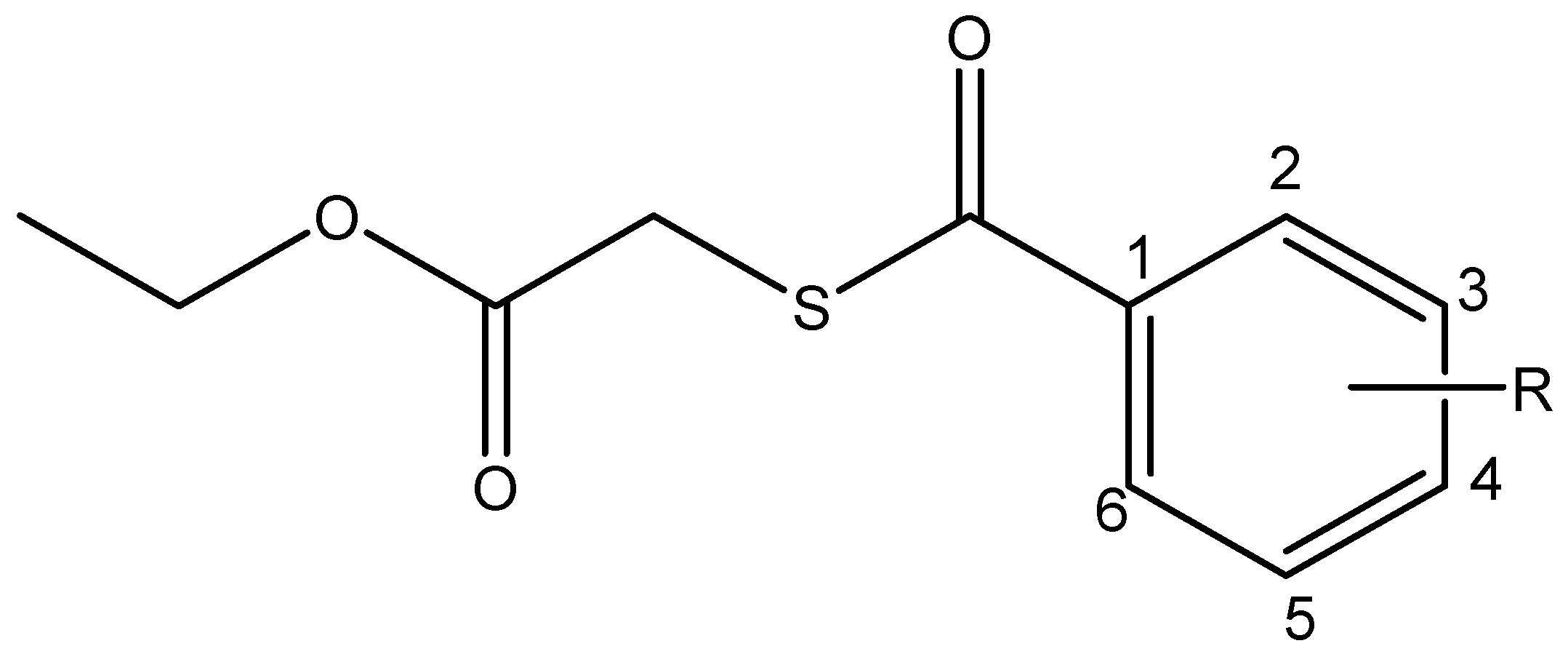
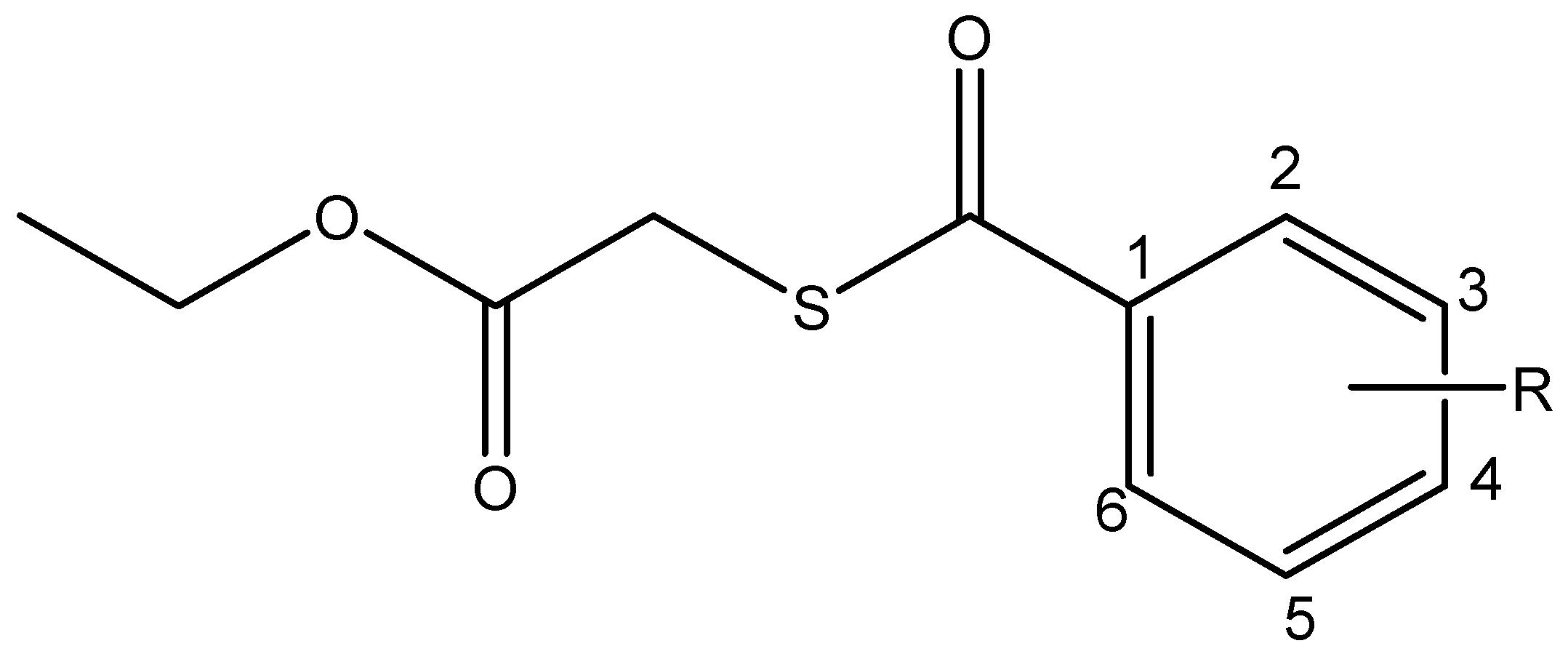
2.1. Synthesis and Spectroscopical Characterization
2.1.1. Ethyl 2-((4-chlorobenzoyl)thio)acetate (I)
2.1.2. Ethyl 2-((3-nitrobenzoyl)thio)acetate (II)
2.1.3. Ethyl 2-((4-nitrobenzoyl)thio)acetate (III)
2.2. Biological Activity
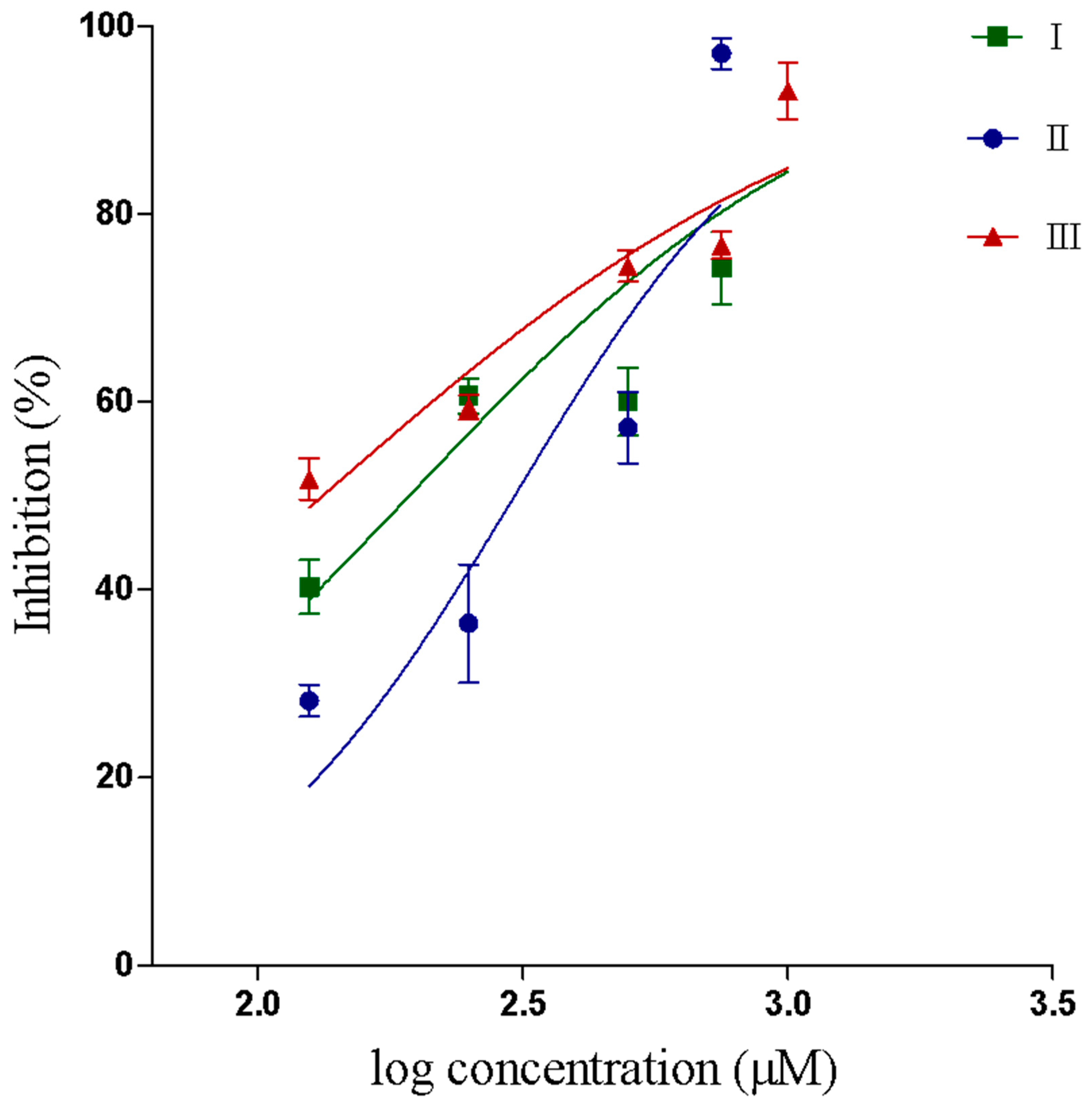
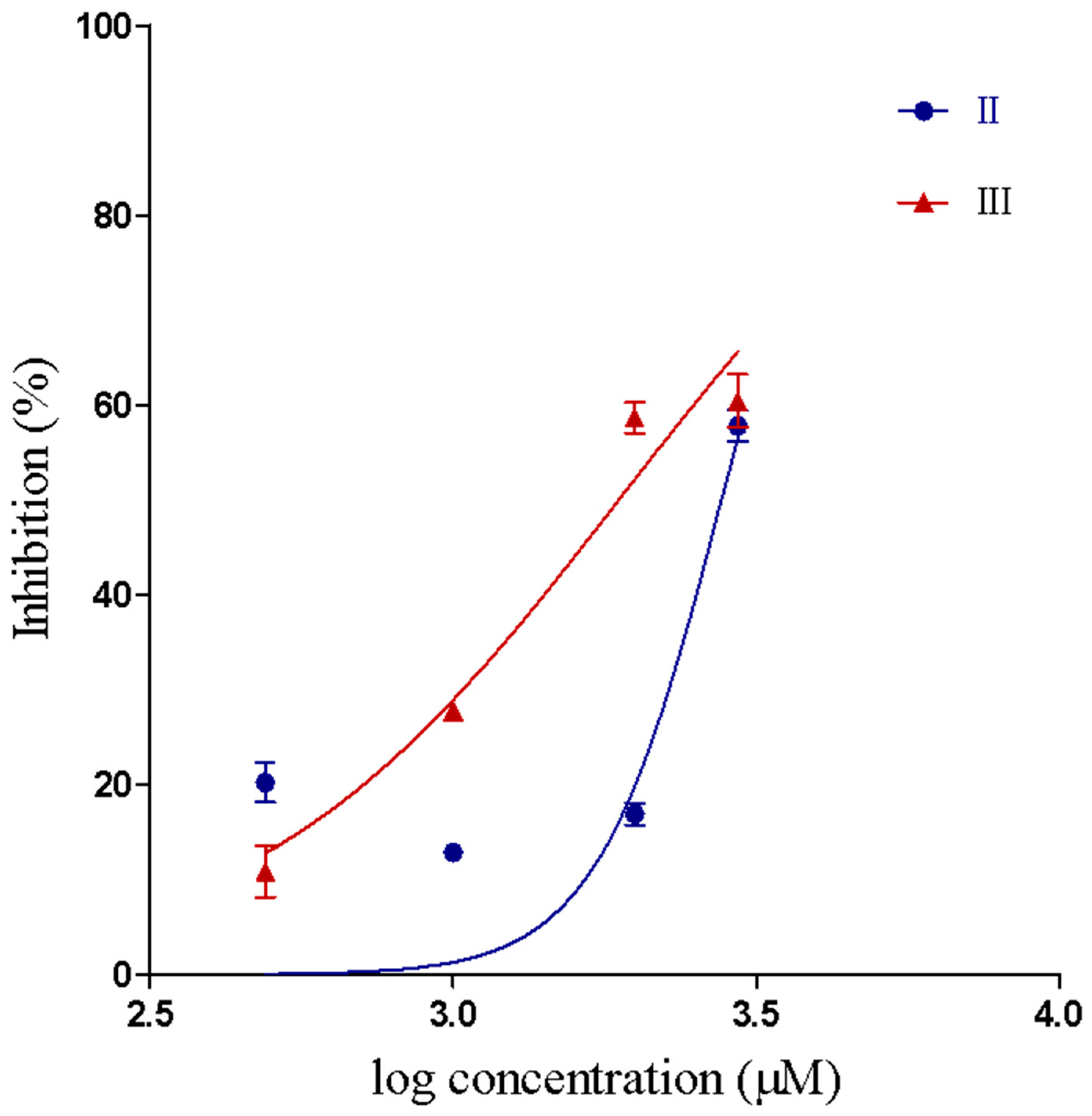
2.2.1. Inhibition of PLA2 activity
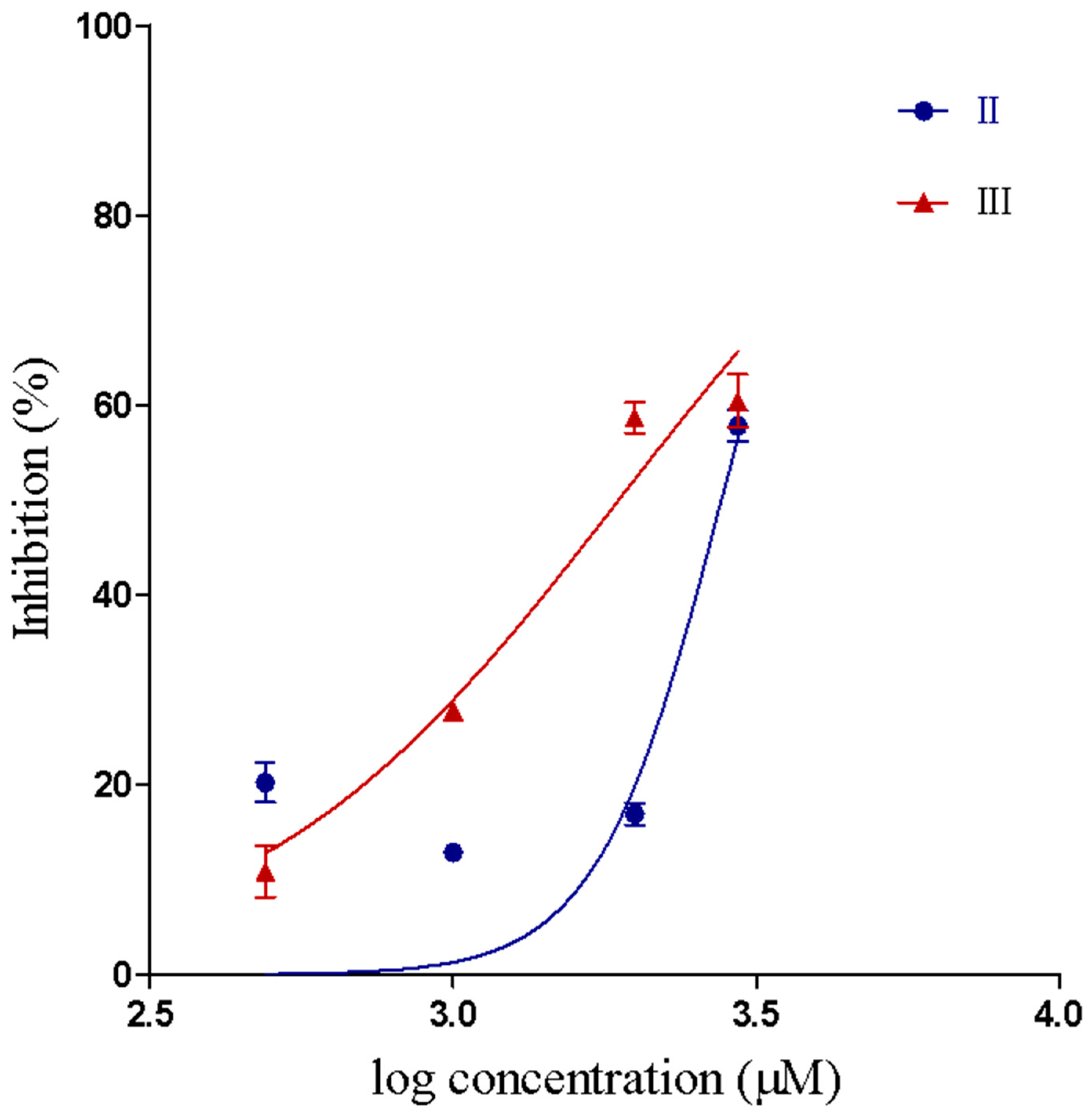
2.2.2. Inhibition of Proteolytic Activity
2.3. Computational Studies
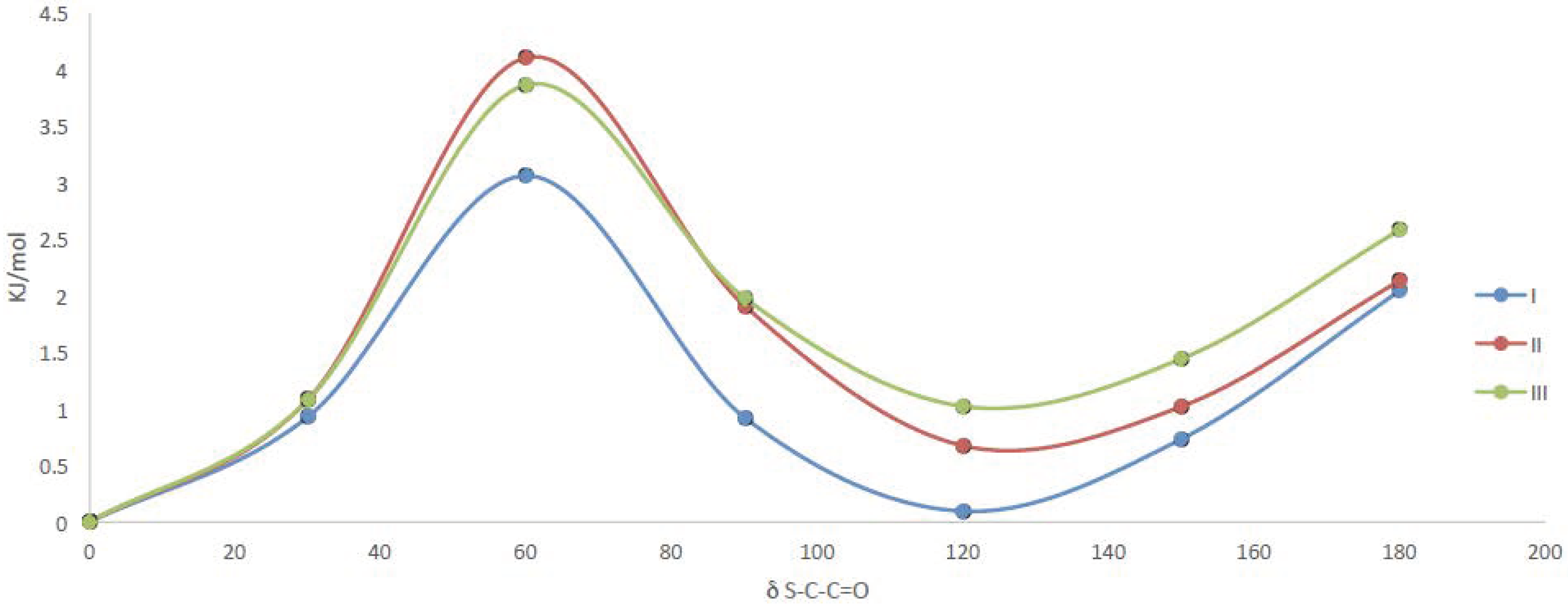
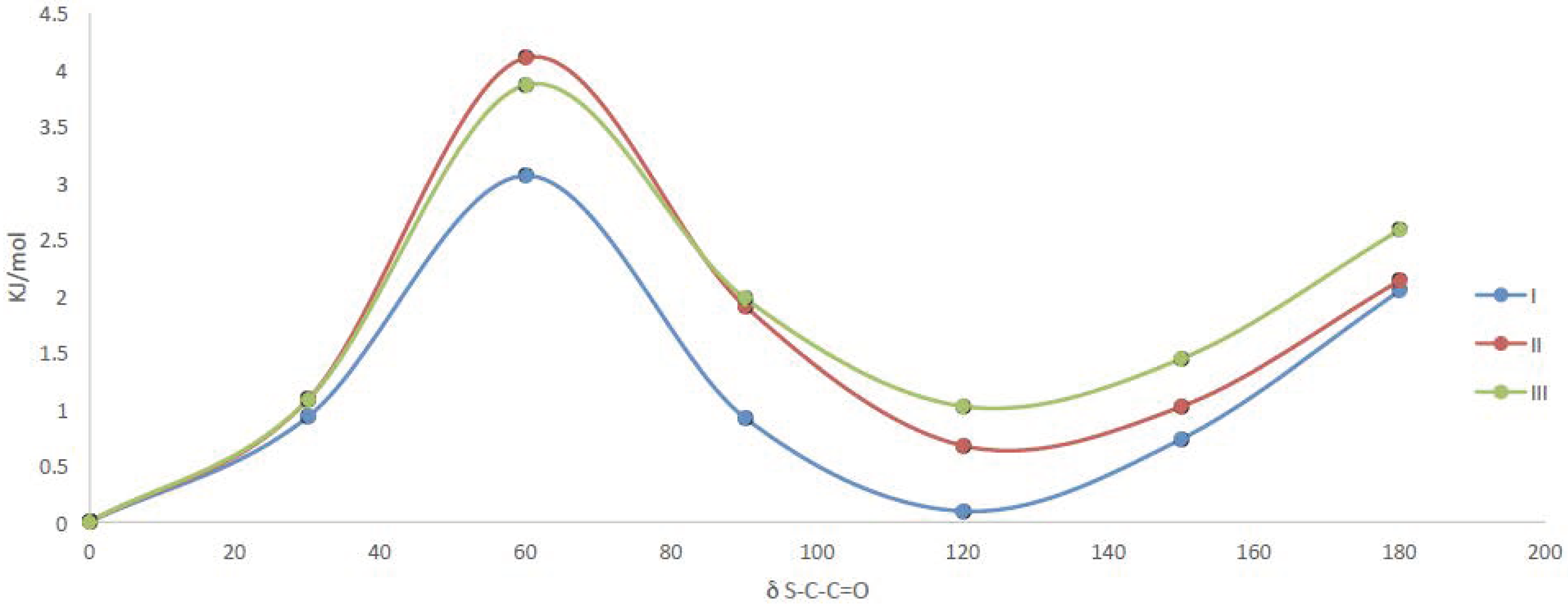
2.3.1. Quantum Chemical Calculations
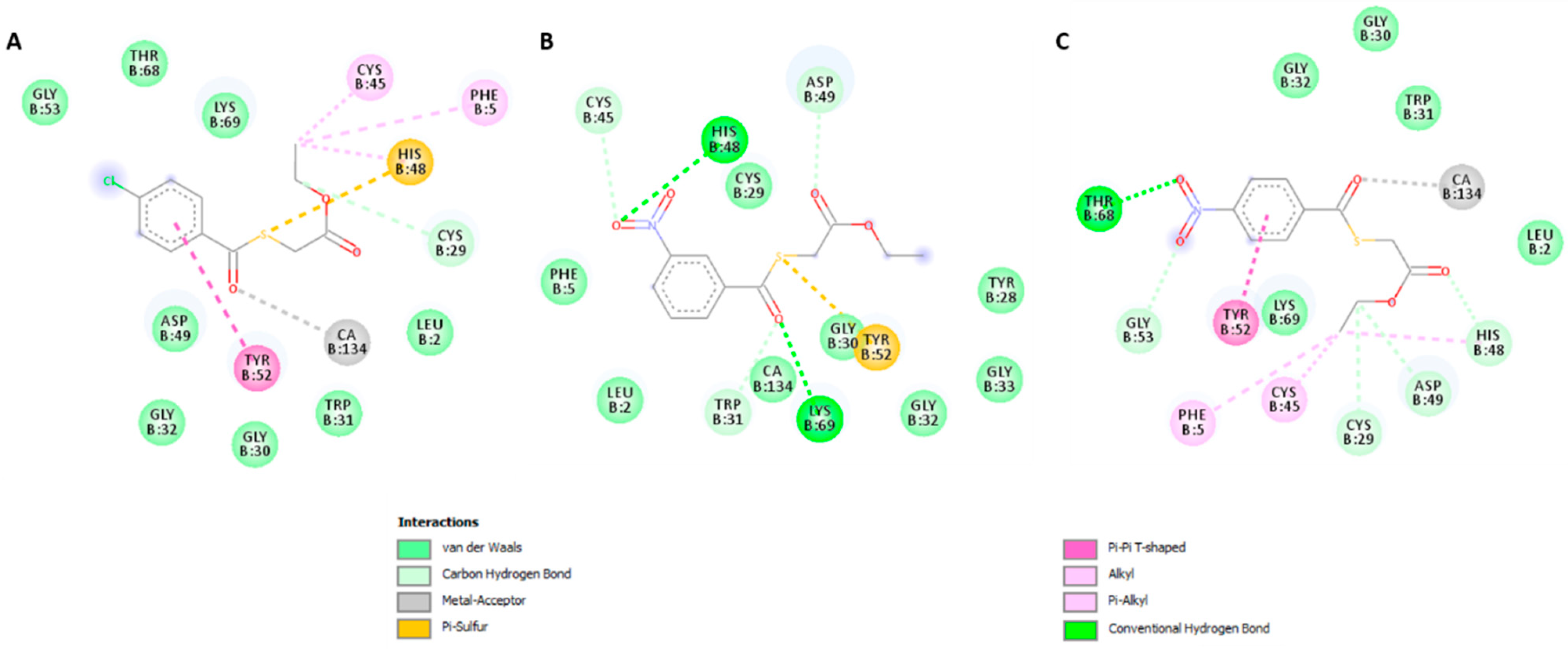
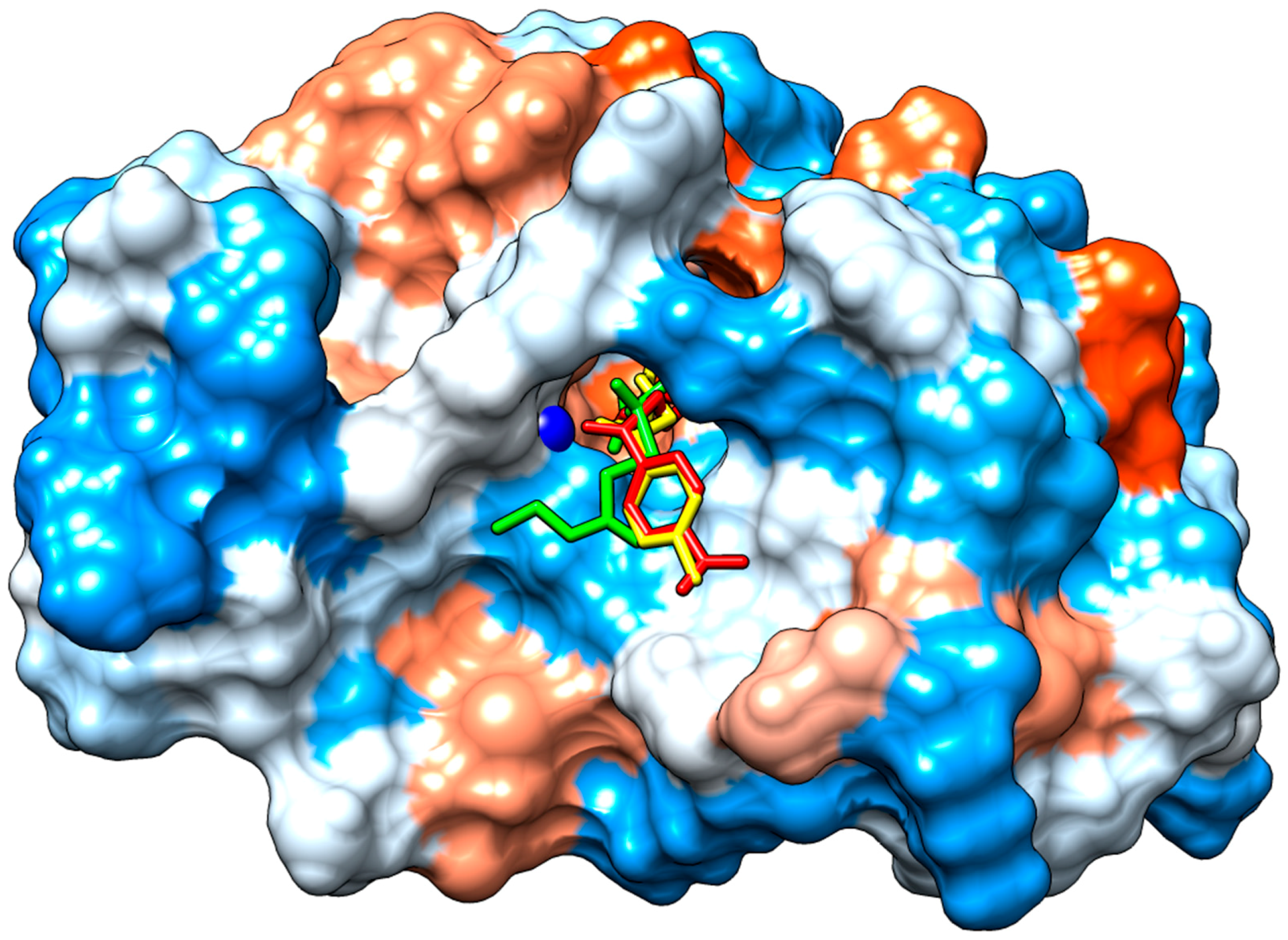
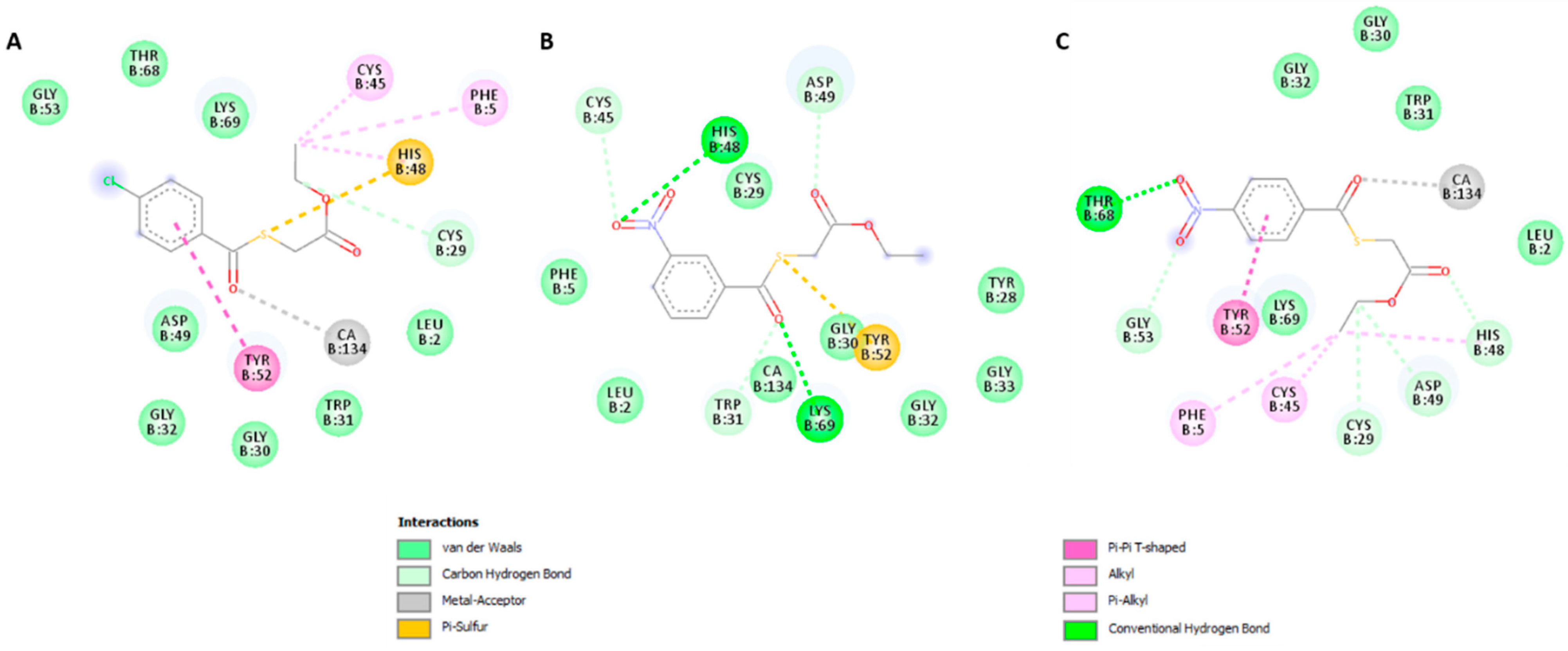
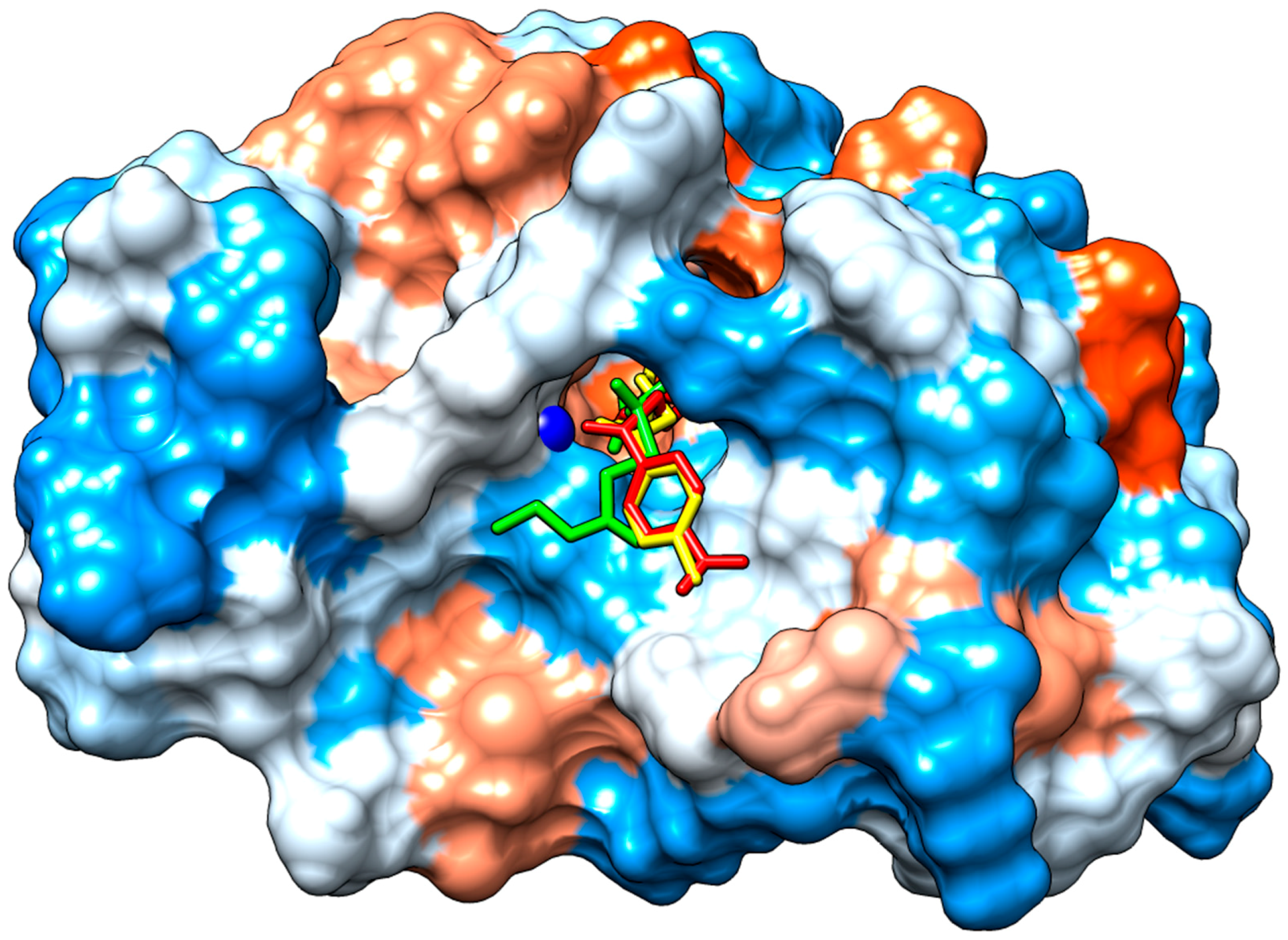
2.3.2. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. General
4.2. Syntheses
4.3. Spectroscopical Characterization
4.4. Toxins Isolation
4.5. Inhibition of PLA2 Activity Using 4-nitro-3-octanoyloxybenzoic acid (4N3OBA) as Monodispersed Substrate
4.6. Inhibition of Proteolytic Activity
4.7. Computational Studies
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chippaux, J. Snakebite envenomation turns again into a neglected tropical disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–2. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: http://www.webcitation.org/78DbPvhth (accessed on 8 May 2019).
- Instituto Nacional de Salud. Available online: http://www.webcitation.org/78DfIpikC (accessed on 8 May 2019).
- Cilibrasi, C.; Riva, G.; Romano, G.; Cadamuro, M.; Bazzoni, R.; Butta, V.; Paoletta, L.; Dalpra, L.; Strazzabosco, M.; Lavitrano, M.; et al. Resveratrol impairs glioma stem cells proliferation and motility by modulating the wnt signaling pathway. PLoS ONE 2017, 12, e0169854. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Six, D.A.; Dennis, E.A. The expanding superfamily of phospholipase A2 enzymes: Classification and characterization. Biochim. Biophys. Acta 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Squaiella-Baptistão, C.C.; Sant’Anna, O.A.; Marcelino, J.R.; Tambourgi, D.V. The history of antivenoms development: Beyond Calmette and Vital Brazil. Toxicon 2018, 150, 86–95. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Herrera, C.; Fernández, J.; Lomonte, B.; Fox, J.W. Unresolved issues in the understanding of the pathogenesis of local tissue damage induced by snake venoms. Toxicon 2018, 148, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lättig, J.; Böhl, M.; Fischer, P.; Tischer, S.; Tietböhl, C.; Menschikowski, M.; Gutzeit, H.O.; Metz, P.; Pisabarro, M.T. Mechanism of inhibition of human secretory phospholipase A2 by flavonoids: Rationale for lead design. J. Comput.-Aided Mol. Des. 2007, 21, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Henao Castañeda, I.C.; Pereañez, J.A.; Jios, J.L. Substituted thiobenzoic acid S-benzyl esters as potential inhibitors of a snake venom phospholipase A2: Synthesis, spectroscopic and computational studies. J. Mol. Struct. 2012, 1028, 7–12. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Doley, R.; Saikia, D. Isolation of a snake venom phospholipase A2 (PLA2) inhibitor (AIPLAI) from leaves of Azadirachta indica (Neem): Mechanism of PLA2 inhibition by AIPLAI in vitro condition. Toxicon 2008, 51, 1548–1553. [Google Scholar] [CrossRef]
- Pereañez, J.A.; Núñez, V.; Patiño, A.C. Inhibitory effects of bile acids on enzymatic and pharmacological activities of a snake venom phospholipase A2 from group IIA. Protein J. 2011, 30, 253–261. [Google Scholar] [CrossRef]
- Gómez-Betancur, I.; Pereañez, J.A.; Patiño, A.C.; Benjumea, D. Inhibitory effect of pinostrobin from Renealmia alpinia, on the enzymatic and biological activities of a PLA2. Int. J. Biol. Macromol. 2016, 89, 35–42. [Google Scholar] [CrossRef]
- Pereañez, J.A.; Patiño, A.C.; Núñez, V.; Osorio, E. The biflavonoid morelloflavone inhibits the enzymatic and biological activities of a snake venom phospholipase A2. Chem.-Biol. Interact. 2014, 220, 94–101. [Google Scholar] [CrossRef]
- Henao Castañeda, I.C.; Della Védova, C.O.; Piro, O.E.; Metzler-Nolte, N.; Jios, J.L. Synthesis of two new thioesters bearing ferrocene: Vibrational characterization and ab initio calculations. X-ray crystal structure of S-(2-methoxyphenyl)ferrocenecarbothioate. Polyhedron 2010, 29, 827–832. [Google Scholar] [CrossRef]
- Castañeda, I.C.H.; Jios, J.L.; Piro, O.E.; Tobón, G.E.; Della Védova, C.O. Conformational and vibrational analysis of S-(2-methoxyphenyl)-4-substituted-benzenecarbothioates, using X-ray, infrared and Raman spectroscopy and theoretical calculations. J. Mol. Struct. 2007, 842, 46–54. [Google Scholar] [CrossRef]
- Warrell, D.A. Clinical Toxicology of Snakebite in Asia. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; CRC Press: New York, NY, USA, 1995. [Google Scholar]
- Escalante, T.; Rucavado, A.; Fox, J.W.; María, J.; Picado, I.C.; De Microbiología, F.; Rica, U.D.C.; Rica, C. Key events in microvascular damage induced by snake venom. J. Proteom. 2011, 74, 1781–1794. [Google Scholar] [CrossRef]
- Angulo, Y.; Lomonte, B.; Rica, U.D.C.; Jose, S.; Rica, C.; De Medicina, E.; Rica, U.D.C.; Jose, S.; Rica, C. Toxicon Biochemistry and toxicology of toxins purified from the venom of the snake Bothrops asper. Toxicon 2009, 54, 949–957. [Google Scholar] [CrossRef]
- Rrez, Â.; Leo, G.; Rojas, G.; Lomonte, B.; Rucavado, A.; Chaves, F.; Rojas, G.; Lomonte, B.; Rucavado, A. nNeutralization of local tissue damage induced by Bothrops asper (terciopelo) snake venom. Toxicon 1998, 36, 1529–1538. [Google Scholar]
- Berg, O.G.; Gelb, M.H. Interfacial Enzymology: The Secreted Phospholipase A2-Paradigm. Chem. Rev. 2001, 101, 2613–2654. [Google Scholar] [CrossRef]
- Emmanuel, A.; Okafor, I.; Fortunatus, E.C.; Colin, L.; Jonathan, I.F.; Andrew, N.C.; Ferdinand, C.C. The Role of Calcium on the Active Site of Snake Venom Phospholipase A2: Molecular Dynamics Simulations. Comput. Biol. Bioinform. 2016, 4, 10–14. [Google Scholar] [CrossRef]
- Salvador, G.; dos Santos, J.; Lomonte, B.; Fontes, M.R.M. Biochimie: Crystal structure of a phospholipase A2 from Bothrops asper venom: Insights into a new putative “myotoxic cluster”. Biochimie 2017, 133, 95–102. [Google Scholar] [CrossRef]
- Cardoso, F.F.; Borges, R.J.; Dreyer, T.R.; Salvador, G.H.M.; Cavalcante, W.L.G.; Pai, M.D.; Gallacci, M.; Fontes, M.R.M. Biochim Biophys Acta Gen Subj: Structural basis of phospholipase A2-like myotoxin inhibition by chicoric acid, a novel potent inhibitor of ophidian toxins. Biochim. Biophys. Acta 2018, 1862, 2728–2737. [Google Scholar] [CrossRef] [PubMed]
- Nu, V.; Marangoni, S.; Ponce-soto, L.A. Biochemical and biological characterization of a PLA 2 from crotoxin complex of Crotalus durissus cumanensis. Toxicon 2009, 53, 534–542. [Google Scholar]
- Patiño, A.C.; Pereañez, J.A.; Núñez, V.; Benjumea, D.M.; Fernandez, M.; Rucavado, A.; Sanz, L.; Calvete, J.J. Isolation and biological characterization of Batx-I, a weak hemorrhagic and fi brinogenolytic PI metalloproteinase from Colombian Bothrops atrox venom. Toxicon 2010, 56, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Preciado, L.M.; Rey-Suárez, P.; Henao, I.C.; Pereañez, J.A. Betulinic, oleanolic and ursolic acids inhibit the enzymatic and biological effects induced by a P-I snake venom metalloproteinase. Chem.-Biol. Interact. 2018, 279, 219–226. [Google Scholar] [CrossRef]
- Dennington, R.; Keith, T.; Millam, J. Gauss View, Version 5; Semichem Inc.: Shawnee Mission, KS, USA, 2009. [Google Scholar]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R. Gaussian 09, Revision, B.01; Gaussian, Inc.: Wallingford, CT, USA, 2010. [Google Scholar]
- Trott, O.; Olson, A.J. Software News and Update AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar]
- Molinspiration. Available online: https://www.molinspiration.com/cgi-bin/properties (accessed on 21 November 2018).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | R3 | R4 | MW a | nON a | nOHNH a | LogP(calc) a |
|---|---|---|---|---|---|---|
| I | H | Cl | 258.73 | 3 | 0 | 3.23 |
| II | NO2 | H | 269.28 | 6 | 0 | 2.48 |
| III | H | NO2 | 269.28 | 6 | 0 | 2.51 |
| Compound | IC50 Value for Inhibition of PLA2 Activity (μM) | 95% Confidence Interval (μM) | IC50 Value for Inhibition of Proteolytic Activity (μM) | 95% Confidence Interval (μM) |
|---|---|---|---|---|
| I | 193.2 a | 133.5–279.5 | - | - |
| II | 305.4 b | 230.5–404.5 | 2774 | 2471–3115 |
| III | 132.7 a | 98.8–178.3 | 1879 | 1679–2102 |
| Compound | 0° | 120° | ||
|---|---|---|---|---|
| δC-S-C=O | δC-O-C=O | δC-S-C=O | δC-O-C=O | |
| I | 5.0° | −0.8° | 5.4° | −0.5° |
| II | 4.7° | −0.5° | 5.8° | 0.0° |
| III | 4.9° | −0.5° | 5.8° | 0.3° |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henao Castañeda, I.C.; Pereañez, J.A.; Preciado, L.M. Synthetic Inhibitors of Snake Venom Enzymes: Thioesters Derived from 2-Sulfenyl Ethylacetate. Pharmaceuticals 2019, 12, 80. https://doi.org/10.3390/ph12020080
Henao Castañeda IC, Pereañez JA, Preciado LM. Synthetic Inhibitors of Snake Venom Enzymes: Thioesters Derived from 2-Sulfenyl Ethylacetate. Pharmaceuticals. 2019; 12(2):80. https://doi.org/10.3390/ph12020080
Chicago/Turabian StyleHenao Castañeda, Isabel C., Jaime A. Pereañez, and Lina M. Preciado. 2019. "Synthetic Inhibitors of Snake Venom Enzymes: Thioesters Derived from 2-Sulfenyl Ethylacetate" Pharmaceuticals 12, no. 2: 80. https://doi.org/10.3390/ph12020080
APA StyleHenao Castañeda, I. C., Pereañez, J. A., & Preciado, L. M. (2019). Synthetic Inhibitors of Snake Venom Enzymes: Thioesters Derived from 2-Sulfenyl Ethylacetate. Pharmaceuticals, 12(2), 80. https://doi.org/10.3390/ph12020080