Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury




Abstract
1. An Overview on Spinal Cord Injury
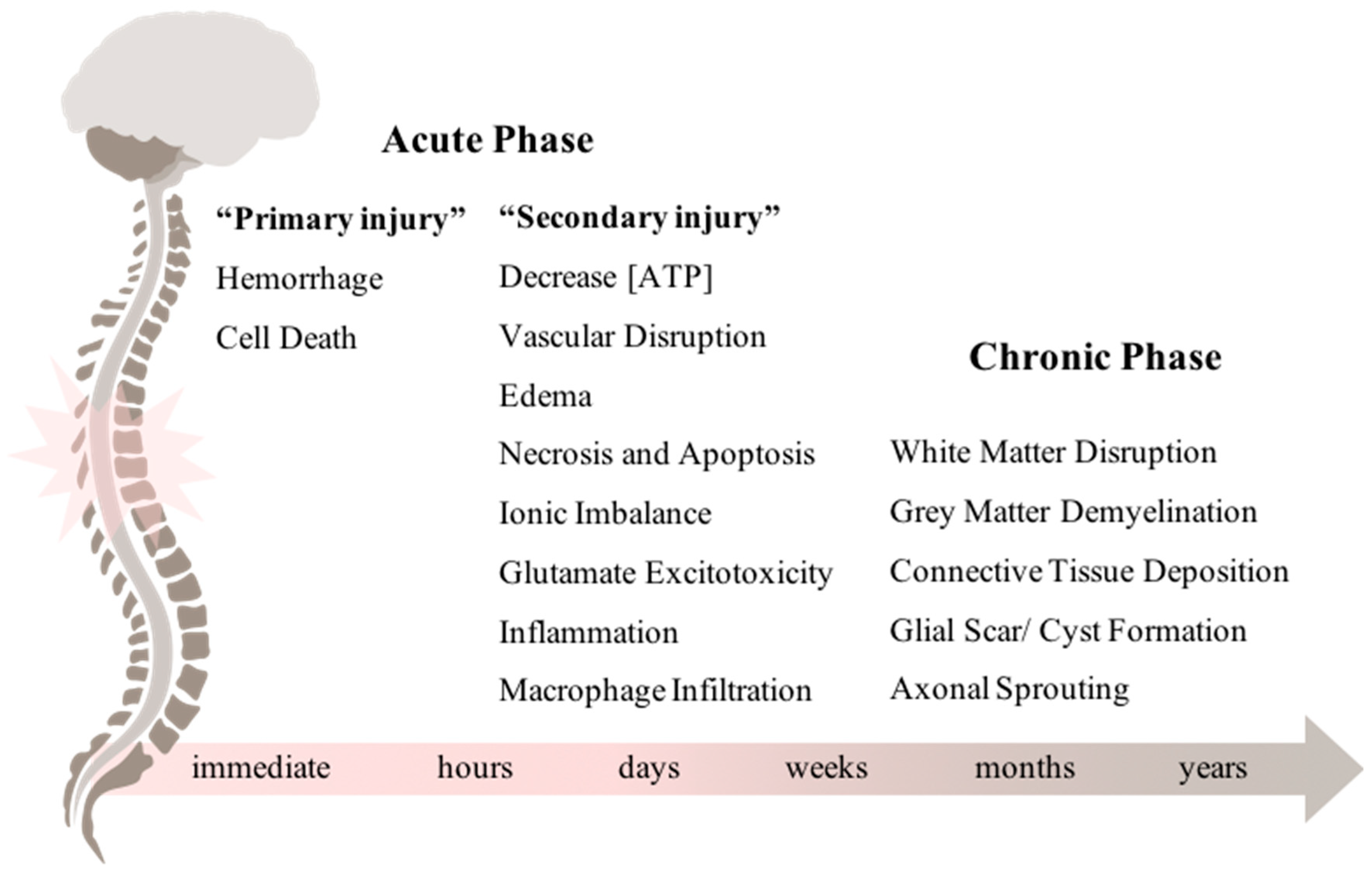
1.1. From Acute to Chronic Phase
1.2. Clinical Management After SCI
1.3. Novel Treatments for SCI
2. Stem Cells in SCI: Past, Present, and Future
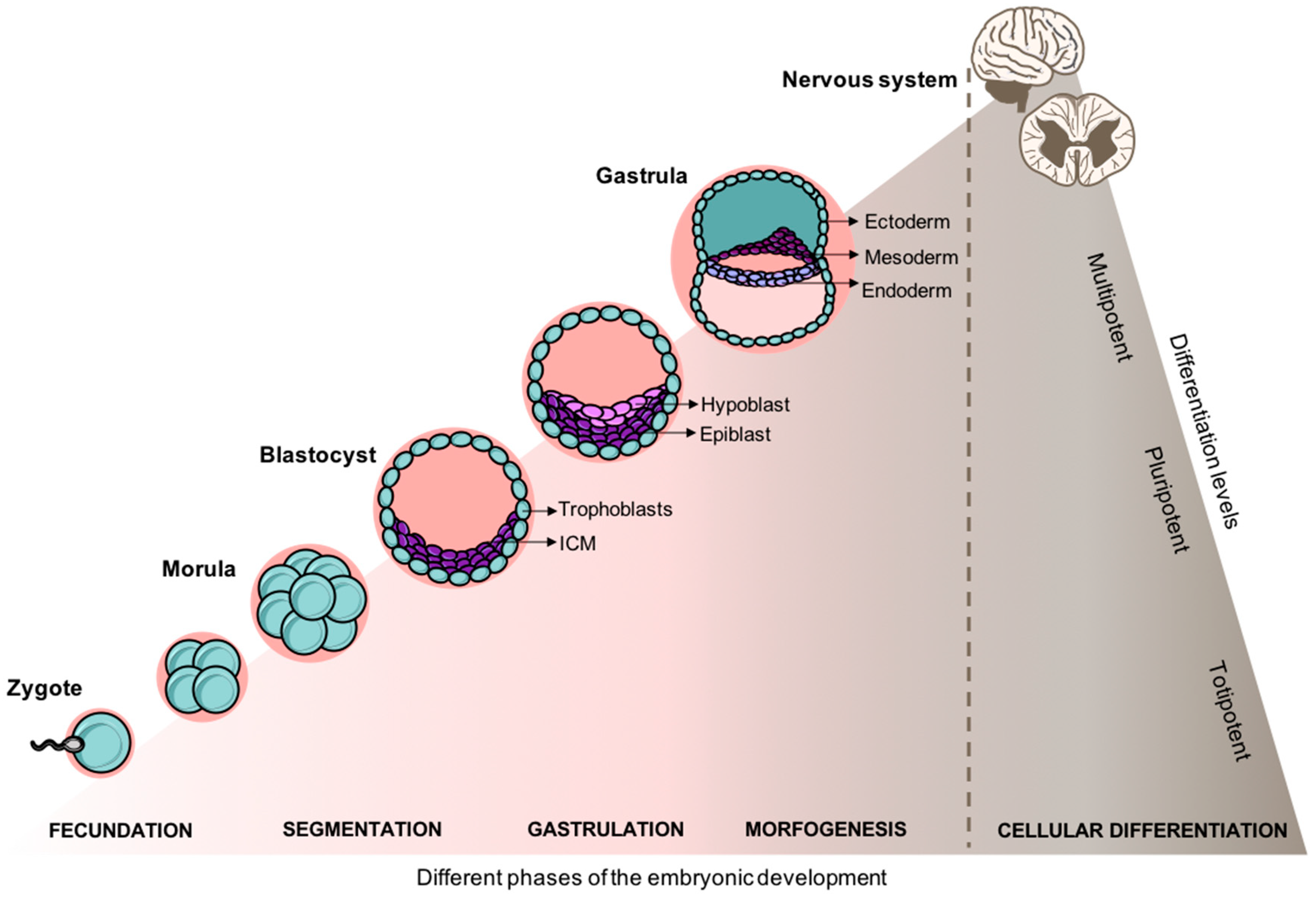
2.1. From the Embryo to A Structured Spinal Cord
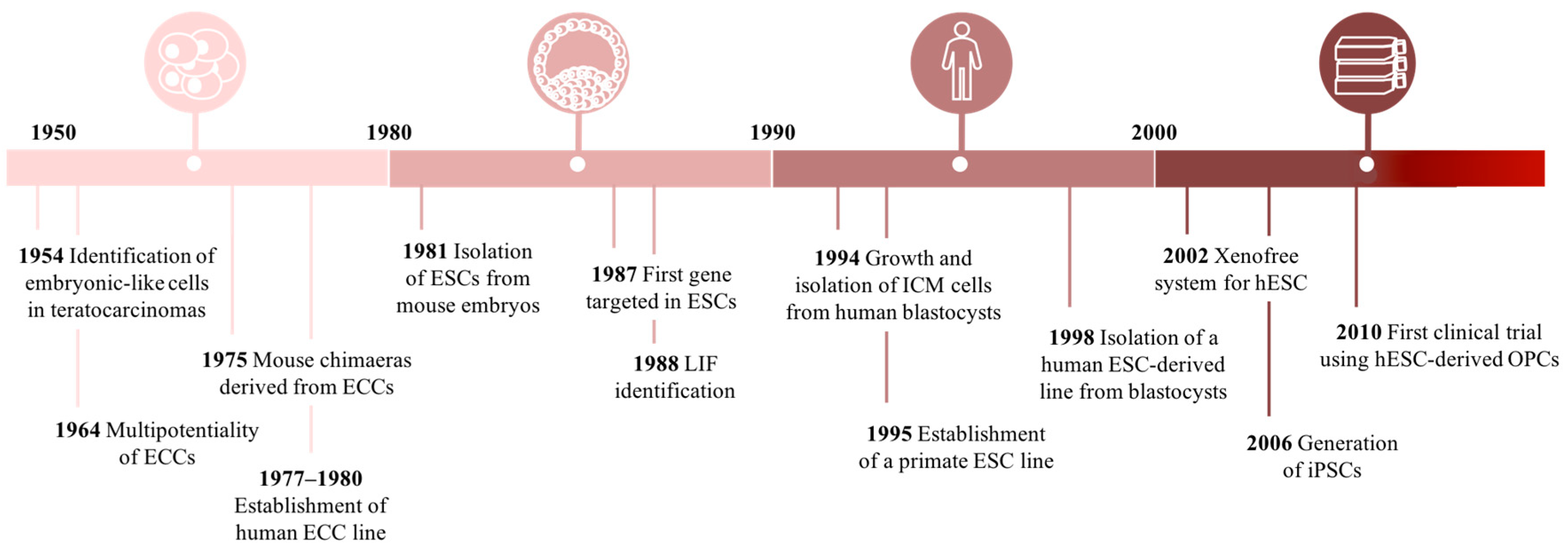
2.2. Historical Perspective of Cell-Based Research
2.2.1. Finding Embryonic Stem Cells
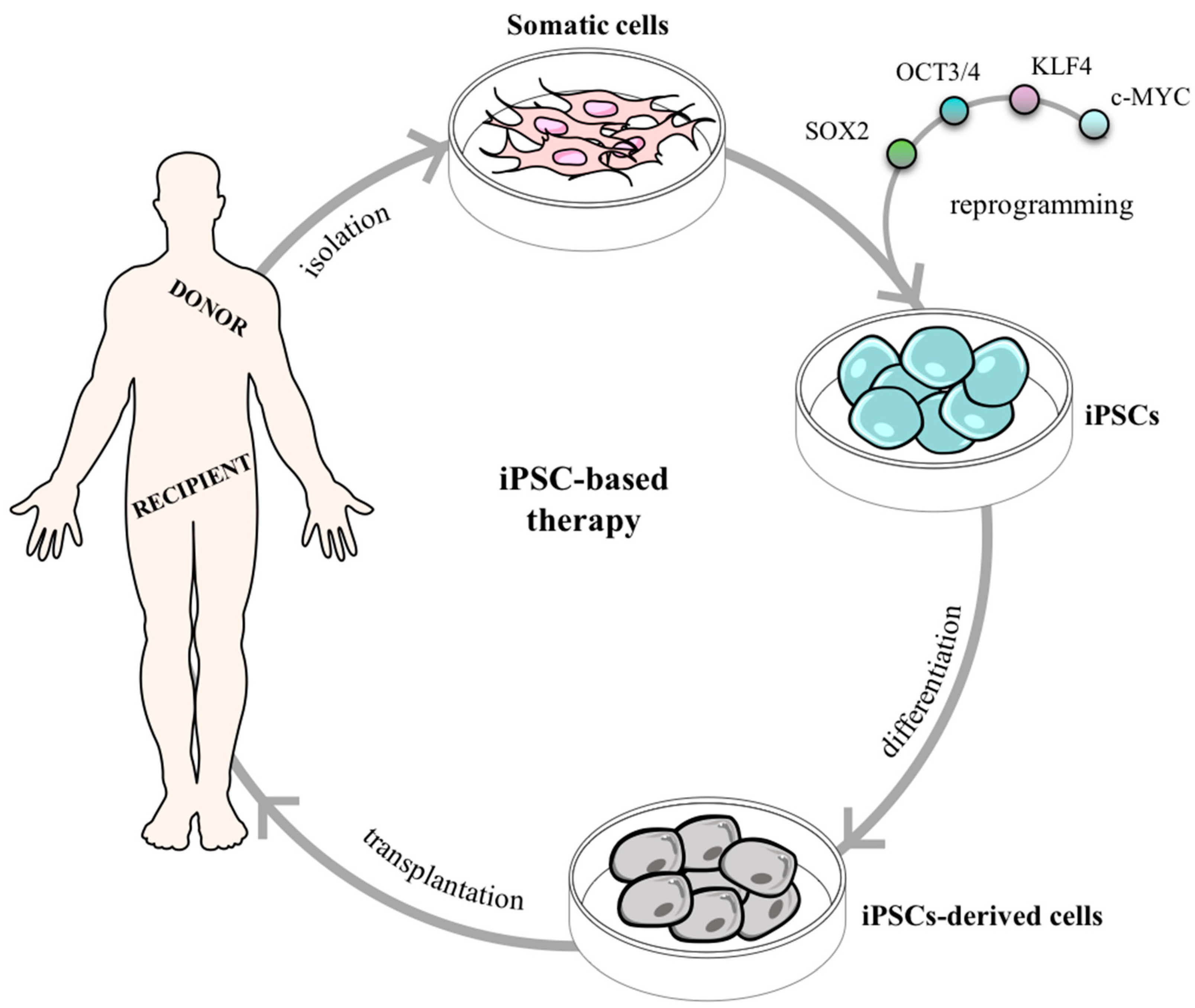
2.2.2. Searching for Pluripotency in Adult Tissues
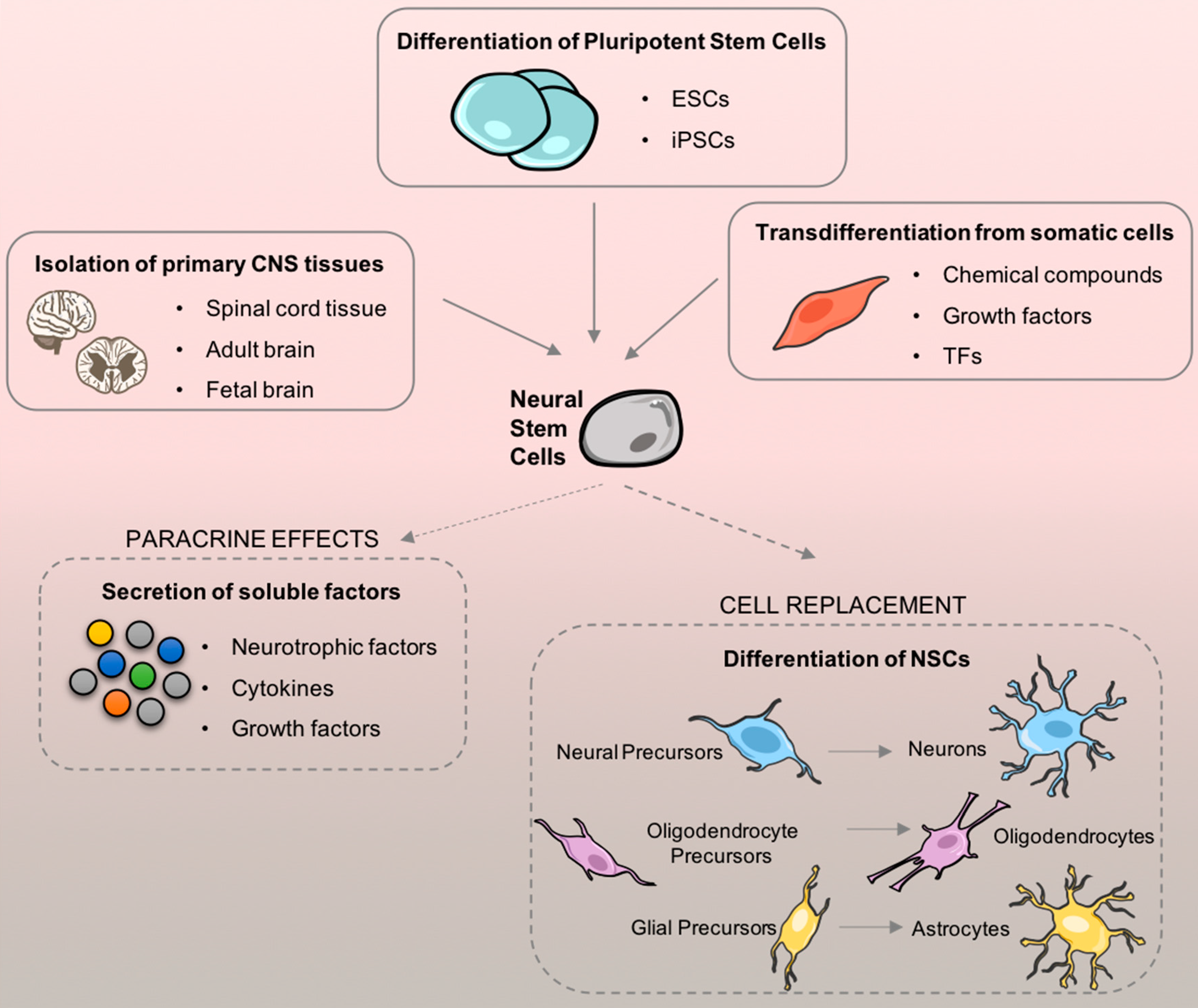
2.2.3. Neural Stem Cells
2.3. Advances on NSC-based Therapy for SCI
2.4. SCI Clinical Trials Based on NSCs
3. Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- James, S.L.; Theadom, A.; Ellenbogen, R.G.; Bannick, M.S.; Montjoy-Venning, W.; Lucchesi, L.R.; Abbasi, N.; Abdulkader, R.; Abraha, H.N.; Adsuar, J.C.; et al. Global, regional, and national burden of traumatic brain injury and spinal cord injury, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 56–87. [Google Scholar] [CrossRef]
- Karunakaran, K.D.; He, J.; Zhao, J.; Cui, J.-L.; Zang, Y.-F.; Zhang, Z.; Biswal, B.B. Differences in Cortical Gray Matter Atrophy of Paraplegia and Tetraplegia after Complete Spinal Cord Injury. J. Neurotrauma 2019. (In press) [CrossRef]
- McDonald, J.W.; Sadowsky, C. Spinal-cord injury. Lancet 2002, 359, 417–425. [Google Scholar] [CrossRef]
- Tator, C.H. Update on the Pathophysiology and Pathology of Acute Spinal Cord Injury. Brain Pathol. 1995, 5, 407–413. [Google Scholar] [CrossRef]
- Allen, A.R. Surgery of experimental lesion of spinal cord equivalent to crush injury of fracture dislocation of spinal column: A preliminary report. J. Am. Med. Assoc. 1911, LVII, 878. [Google Scholar] [CrossRef]
- Hagen, E.M. Acute complications of spinal cord injuries. World J. Orthop. 2015, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Benton, R.L.; Hagg, T. Vascular Pathology as a Potential Therapeutic Target in SCI. Transl. Stroke Res. 2011, 2, 556–574. [Google Scholar] [CrossRef]
- Li, X.-Q.; Fang, B.; Tan, W.-F.; Wang, Z.-L.; Sun, X.-J.; Zhang, Z.-L.; Ma, H. miR-320a affects spinal cord edema through negatively regulating aquaporin-1 of blood–spinal cord barrier during bimodal stage after ischemia reperfusion injury in rats. BMC Neurosci. 2016, 17, 10. [Google Scholar] [CrossRef]
- Phillips, A.A.; Krassioukov, A.V. Contemporary Cardiovascular Concerns after Spinal Cord Injury: Mechanisms, Maladaptations, and Management. J. Neurotrauma 2015, 32, 1927–1942. [Google Scholar] [CrossRef]
- Visavadiya, N.P.; Patel, S.P.; VanRooyen, J.L.; Sullivan, P.G.; Rabchevsky, A.G. Cellular and subcellular oxidative stress parameters following severe spinal cord injury. Redox Biol. 2016, 8, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.S.; Hulsebosch, C.E. GABA and central neuropathic pain following spinal cord injury. Neuropharmacology 2011, 60, 799–808. [Google Scholar] [CrossRef]
- Park, E.; Velumian, A.A.; Fehlings, M.G. The Role of Excitotoxicity in Secondary Mechanisms of Spinal Cord Injury: A Review with an Emphasis on the Implications for White Matter Degeneration. J. Neurotrauma 2004, 21, 754–774. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Kroner, A.; Greenhalgh, A.D.; Zarruk, J.G.; López-Vales, R. Myeloid cell responses after spinal cord injury. J. Neuroimmunol. 2018, 321, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Gensel, J.C.; Zhang, B. Macrophage activation and its role in repair and pathology after spinal cord injury. Brain Res. 2015, 1619, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, S.; Salgado, A.; Silva, N. Immunomodulation as a neuroprotective strategy after spinal cord injury. Neural Regen. Res. 2018, 13, 423. [Google Scholar] [PubMed]
- Lim, S.-W.; Shiue, Y.-L.; Ho, C.-H.; Yu, S.-C.; Kao, P.-H.; Wang, J.-J.; Kuo, J.-R. Anxiety and Depression in Patients with Traumatic Spinal Cord Injury: A Nationwide Population-Based Cohort Study. PLoS ONE 2017, 12, e0169623. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Bouchard, S.; Peters, K.; Woller, S.A.; Madahian, B.; Faghihi, U.; Patel, S.; Bake, S.; Hook, M.A. Inflammation is increased with anxiety- and depression-like signs in a rat model of spinal cord injury. Brain Behav. Immun. 2016, 51, 176–195. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.A.; Burda, J.E.; Ren, Y.; Ao, Y.; O’Shea, T.M.; Kawaguchi, R.; Coppola, G.; Khakh, B.S.; Deming, T.J.; Sofroniew, M.V. Astrocyte scar formation aids central nervous system axon regeneration. Nature 2016, 532, 195–200. [Google Scholar] [CrossRef]
- Dias, D.O.; Kim, H.; Holl, D.; Werne Solnestam, B.; Lundeberg, J.; Carlén, M.; Göritz, C.; Frisén, J. Reducing Pericyte-Derived Scarring Promotes Recovery after Spinal Cord Injury. Cell 2018, 173, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Bertolotto, A.; Levi, G. The proteoglycan chondroitin sulfate is present in a subpopulation of cultured astrocytes and in their precursors. Dev. Biol. 1987, 123, 282–285. [Google Scholar] [CrossRef]
- Hackett, A.R.; Lee, J.K. Understanding the NG2 Glial Scar after Spinal Cord Injury. Front. Neurol. 2016, 7, 199. [Google Scholar] [CrossRef]
- Feng, N.; Hao, G.; Yang, F.; Qu, F.; Zheng, H.; Liang, S.; Jin, Y. Transplantation of mesenchymal stem cells promotes the functional recovery of the central nervous system following cerebral ischemia by inhibiting myelin-associated inhibitor expression and neural apoptosis. Exp. Ther. Med. 2016, 11, 1595–1600. [Google Scholar] [CrossRef]
- Kucher, K.; Johns, D.; Maier, D.; Abel, R.; Badke, A.; Baron, H.; Thietje, R.; Casha, S.; Meindl, R.; Gomez-Mancilla, B.; et al. First-in-Man Intrathecal Application of Neurite Growth-Promoting Anti-Nogo-A Antibodies in Acute Spinal Cord Injury. Neurorehabil. Neural Repair 2018, 32, 578–589. [Google Scholar] [CrossRef]
- Dyck, S.M.; Karimi-Abdolrezaee, S. Chondroitin sulfate proteoglycans: Key modulators in the developing and pathologic central nervous system. Exp. Neurol. 2015, 269, 169–187. [Google Scholar] [CrossRef]
- Andrews, P.W. From teratocarcinomas to embryonic stem cells. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2002, 357, 405–417. [Google Scholar] [CrossRef]
- Lang, B.T.; Cregg, J.M.; DePaul, M.A.; Tran, A.P.; Xu, K.; Dyck, S.M.; Madalena, K.M.; Brown, B.P.; Weng, Y.-L.; Li, S.; et al. Modulation of the proteoglycan receptor PTPσ promotes recovery after spinal cord injury. Nature 2015, 518, 404–408. [Google Scholar] [CrossRef]
- Suzuki, H.; Ahuja, C.S.; Salewski, R.P.; Li, L.; Satkunendrarajah, K.; Nagoshi, N.; Shibata, S.; Fehlings, M.G. Neural stem cell mediated recovery is enhanced by Chondroitinase ABC pretreatment in chronic cervical spinal cord injury. PLoS ONE 2017, 12, e0182339. [Google Scholar] [CrossRef]
- Oni-Orisan, A.; Kaushal, M.; Li, W.; Leschke, J.; Ward, B.D.; Vedantam, A.; Kalinosky, B.; Budde, M.D.; Schmit, B.D.; Li, S.-J.; et al. Alterations in Cortical Sensorimotor Connectivity following Complete Cervical Spinal Cord Injury: A Prospective Resting-State fMRI Study. PLoS ONE 2016, 11, e0150351. [Google Scholar] [CrossRef]
- Theodore, N.; Hadley, M.N.; Aarabi, B.; Dhall, S.S.; Gelb, D.E.; Hurlbert, R.J.; Rozzelle, C.J.; Ryken, T.C.; Walters, B.C. Prehospital Cervical Spinal Immobilization After Trauma. Neurosurgery 2013, 72, 22–34. [Google Scholar] [CrossRef]
- Li, Y.; Walker, C.L.; Zhang, Y.P.; Shields, C.B.; Xu, X.-M. Surgical decompression in acute spinal cord injury: A review of clinical evidence, animal model studies, and potential future directions of investigation. Front. Biol. 2014, 9, 127–136. [Google Scholar] [CrossRef]
- Bartholdi, D.; Schwab, M.E. Methylprednisolone inhibits early inflammatory processes but not ischemic cell death after experimental spinal cord lesion in the rat. Brain Res. 1995, 672, 177–186. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Schroeder, G.D.; Vaccaro, A.R.; Fehlings, M.G. Spinal Cord Injury—What Are the Controversies? J. Orthop. Trauma 2017, 31, S7–S13. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.C.; Daugherty, M.C.; Moody, S.M.; Falcone, R.A.; Bierbrauer, K.S.; Geis, G.L. Lessons learned from administration of high-dose methylprednisolone sodium succinate for acute pediatric spinal cord injuries. J. Neurosurg. Pediatr. 2017, 20, 567–574. [Google Scholar] [CrossRef]
- Shultz, R.; Zhong, Y. Minocycline targets multiple secondary injury mechanisms in traumatic spinal cord injury. Neural Regen. Res. 2017, 12, 702. [Google Scholar] [PubMed]
- Casha, S.; Zygun, D.; McGowan, M.D.; Bains, I.; Yong, V.W.; John Hurlbert, R. Results of a phase II placebo-controlled randomized trial of minocycline in acute spinal cord injury. Brain 2012, 135, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Nagoshi, N.; Nakashima, H.; Fehlings, M. Riluzole as a Neuroprotective Drug for Spinal Cord Injury: From Bench to Bedside. Molecules 2015, 20, 7775–7789. [Google Scholar] [CrossRef]
- Vasconcelos, N.L.; Gomes, E.D.; Oliveira, E.P.; Silva, C.J.; Lima, R.; Sousa, N.; Salgado, A.J.; Silva, N.A. Combining neuroprotective agents: Effect of riluzole and magnesium in a rat model of thoracic spinal cord injury. Spine J. 2016, 16, 1015–1024. [Google Scholar] [CrossRef]
- Chow, D.S.L.; Teng, Y.; Toups, E.G.; Aarabi, B.; Harrop, J.S.; Shaffrey, C.I.; Johnson, M.M.; Boakye, M.; Frankowski, R.F.; Fehlings, M.G.; et al. Pharmacology of riluzole in acute spinal cord injury. J. Neurosurg. Spine 2012, 17, 129–140. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Nakashima, H.; Nagoshi, N.; Chow, D.S.L.; Grossman, R.G.; Kopjar, B. Rationale, design and critical end points for the Riluzole in Acute Spinal Cord Injury Study (RISCIS): A randomized, double-blinded, placebo-controlled parallel multi-center trial. Spinal Cord 2016, 54, 8–15. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.; Li, J.; Li, X.; Xiao, J. Fibroblast growth factors in the management of spinal cord injury. J. Cell. Mol. Med. 2018, 22, 25–37. [Google Scholar] [CrossRef]
- Nordblom, J.; Persson, J.K.E.; Brismar, H.; Sjodahl, J.; Josephson, A.; Frostell, A.; Thams, S.; Brundin, L.; Svensson, M.; Mattsson, P. FGF1 containing biodegradable device with peripheral nerve grafts induces corticospinal tract regeneration and motor evoked potentials after spinal cord resection. Restor. Neurol. Neurosci. 2012, 30, 91–102. [Google Scholar]
- Nishio, Y.; Koda, M.; Kamada, T.; Someya, Y.; Kadota, R.; Mannoji, C.; Miyashita, T.; Okada, S.; Okawa, A.; Moriya, H.; et al. Granulocyte Colony-Stimulating Factor Attenuates Neuronal Death and Promotes Functional Recovery After Spinal Cord Injury in Mice. J. Neuropathol. Exp. Neurol. 2007, 66, 724–731. [Google Scholar] [CrossRef]
- Kamiya, K.; Koda, M.; Furuya, T.; Kato, K.; Takahashi, H.; Sakuma, T.; Inada, T.; Ota, M.; Maki, S.; Okawa, A.; et al. Neuroprotective therapy with granulocyte colony-stimulating factor in acute spinal cord injury: A comparison with high-dose methylprednisolone as a historical control. Eur. Spine J. 2015, 24, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Yamazaki, M.; Okawa, A.; Sakuma, T.; Kato, K.; Hashimoto, M.; Hayashi, K.; Furuya, T.; Fujiyoshi, T.; Kawabe, J.; et al. Neuroprotective therapy using granulocyte colony-stimulating factor for acute spinal cord injury: A phase I/IIa clinical trial. Eur. Spine J. 2012, 21, 2580–2587. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Gensel, J.C.; Ankeny, D.P.; Alexander, J.K.; Donnelly, D.J.; Popovich, P.G. Identification of two distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in the injured mouse spinal cord. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 13435–13444. [Google Scholar] [CrossRef] [PubMed]
- Francos-Quijorna, I.; Amo-Aparicio, J.; Martinez-Muriana, A.; López-Vales, R. IL-4 drives microglia and macrophages toward a phenotype conducive for tissue repair and functional recovery after spinal cord injury. Glia 2016, 64, 2079–2092. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.; Monteiro, S.; Lopes, J.; Barradas, P.; Vasconcelos, N.; Gomes, E.; Assunção-Silva, R.; Teixeira, F.; Morais, M.; Sousa, N.; et al. Systemic Interleukin-4 Administration after Spinal Cord Injury Modulates Inflammation and Promotes Neuroprotection. Pharmaceuticals 2017, 10, 83. [Google Scholar] [CrossRef]
- Anderson, K.D.; Guest, J.D.; Dietrich, W.D.; Bartlett Bunge, M.; Curiel, R.; Dididze, M.; Green, B.A.; Khan, A.; Pearse, D.D.; Saraf-Lavi, E.; et al. Safety of Autologous Human Schwann Cell Transplantation in Subacute Thoracic Spinal Cord Injury. J. Neurotrauma 2017, 34, 2950–2963. [Google Scholar] [CrossRef]
- Gomes, E.D.; Mendes, S.S.; Assunção-Silva, R.C.; Teixeira, F.G.; Pires, A.O.; Anjo, S.I.; Manadas, B.; Leite-Almeida, H.; Gimble, J.M.; Sousa, N.; et al. Co-Transplantation of Adipose Tissue-Derived Stromal Cells and Olfactory Ensheathing Cells for Spinal Cord Injury Repair: Co-transplantation of ASCs and OECs for SCI Repair. Stem Cells 2018, 36, 696–708. [Google Scholar] [CrossRef]
- May, Z.; Kumar, R.; Fuehrmann, T.; Tam, R.; Vulic, K.; Forero, J.; Osma, A.L.; Fenrich, K.; Assinck, P.; Lee, M.J.; et al. Adult skin-derived precursor Schwann cell grafts form growths in the injured spinal cord of Fischer rats. Biomed. Mater. 2018, 13, 034101. [Google Scholar] [CrossRef]
- Gomes, E.D.; Mendes, S.S.; Leite-Almeida, H.; Gimble, J.M.; Tam, R.Y.; Shoichet, M.S.; Sousa, N.; Silva, N.A.; Salgado, A.J. Combination of a peptide-modified gellan gum hydrogel with cell therapy in a lumbar spinal cord injury animal model. Biomaterials 2016, 105, 38–51. [Google Scholar] [CrossRef]
- Kawabata, S.; Takano, M.; Numasawa-Kuroiwa, Y.; Itakura, G.; Kobayashi, Y.; Nishiyama, Y.; Sugai, K.; Nishimura, S.; Iwai, H.; Isoda, M.; et al. Grafted Human iPS Cell-Derived Oligodendrocyte Precursor Cells Contribute to Robust Remyelination of Demyelinated Axons after Spinal Cord Injury. Stem Cell Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Salewski, R.P.; Mitchell, R.A.; Li, L.; Shen, C.; Milekovskaia, M.; Nagy, A.; Fehlings, M.G. Transplantation of Induced Pluripotent Stem Cell-Derived Neural Stem Cells Mediate Functional Recovery Following Thoracic Spinal Cord Injury Through Remyelination of Axons: Myelination Following SCI With iPS-dNSC Treatment. Stem Cell. Transl. Med. 2015, 4, 743–754. [Google Scholar] [CrossRef]
- Tam, P.P.L.; Rossant, J. Mouse embryonic chimeras: Tools for studying mammalian development. Development 2003, 130, 6155–6163. [Google Scholar] [CrossRef]
- Arkell Ruth, M.; Tam Patrick, P.L. Initiating head development in mouse embryos: Integrating signalling and transcriptional activity. Open Biol. 2012, 2, 120030. [Google Scholar] [CrossRef]
- Takata, N.; Sakakura, E.; Eiraku, M.; Kasukawa, T.; Sasai, Y. Self-patterning of rostral-caudal neuroectoderm requires dual role of Fgf signaling for localized Wnt antagonism. Nat. Commun. 2017, 8, 1339. [Google Scholar] [CrossRef]
- Purves, D.; Augustine, G.J.; Fitzpatrick, D.; Katz, L.C.; LaMantia, A.-S.; McNamara, J.O.; Williams, S.M. The Initial Formation of the Nervous System: Gastrulation and Neurulation, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2001. [Google Scholar]
- Gouti, M.; Tsakiridis, A.; Wymeersch, F.J.; Huang, Y.; Kleinjung, J.; Wilson, V.; Briscoe, J. In Vitro Generation of Neuromesodermal Progenitors Reveals Distinct Roles for Wnt Signalling in the Specification of Spinal Cord and Paraxial Mesoderm Identity. PLoS Biol. 2014, 12, e1001937. [Google Scholar] [CrossRef]
- Henrique, D.; Abranches, E.; Verrier, L.; Storey, K.G. Neuromesodermal progenitors and the making of the spinal cord. Development 2015, 142, 2864–2875. [Google Scholar] [CrossRef]
- Javali, A.; Misra, A.; Leonavicius, K.; Acharyya, D.; Vyas, B.; Sambasivan, R. Co-expression of Tbx6 and Sox2 identifies a novel transient neuromesoderm progenitor cell state. Dev. Camb. Engl. 2017, 144, 4522–4529. [Google Scholar] [CrossRef]
- Lippmann, E.S.; Williams, C.E.; Ruhl, D.A.; Estevez-Silva, M.C.; Chapman, E.R.; Coon, J.J.; Ashton, R.S. Deterministic HOX Patterning in Human Pluripotent Stem Cell-Derived Neuroectoderm. Stem Cell Rep. 2015, 4, 632–644. [Google Scholar] [CrossRef]
- Mazzoni, E.O.; Mahony, S.; Peljto, M.; Patel, T.; Thornton, S.R.; McCuine, S.; Reeder, C.; Boyer, L.A.; Young, R.A.; Gifford, D.K.; et al. Saltatory remodeling of Hox chromatin in response to rostrocaudal patterning signals. Nat. Neurosci. 2013, 16, 1191–1198. [Google Scholar] [CrossRef]
- Philippidou, P.; Dasen, J.S. Hox Genes: Choreographers in Neural Development, Architects of Circuit Organization. Neuron 2013, 80, 12–34. [Google Scholar] [CrossRef]
- Stevens, L.C.; Little, C.C. Spontaneous testicular teratomas in an inbred strain of mice. Proc. Natl. Acad. Sci. USA 1954, 40, 1080–1087. [Google Scholar] [CrossRef]
- Kleinsmith, L.J.; Pierce, G.B. Multipotentiality of single embryonal carcinoma cells. Cancer Res. 1964, 24, 1544–1551. [Google Scholar]
- Brinster, R.L. The effect of cells transferred into the mouse blastocyst on subsequent development. J. Exp. Med. 1974, 140, 1049–1056. [Google Scholar] [CrossRef]
- Mintz, B.; Illmensee, K. Normal genetically mosaic mice produced from malignant teratocarcinoma cells. Proc. Natl. Acad. Sci. USA 1975, 72, 3585–3589. [Google Scholar] [CrossRef]
- Martin, G.R.; Evans, M.J. Differentiation of clonal lines of teratocarcinoma cells: Formation of embryoid bodies in vitro. Proc. Natl. Acad. Sci. USA 1975, 72, 1441–1445. [Google Scholar] [CrossRef]
- Solter, D. From teratocarcinomas to embryonic stem cells and beyond: A history of embryonic stem cell research. Nat. Rev. Genet. 2006, 7, 319–327. [Google Scholar] [CrossRef]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154. [Google Scholar] [CrossRef]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef]
- Thompson, S.; Stern, P.L.; Webb, M.; Walsh, F.S.; Engstrom, W.; Evans, E.P.; Graham, C.F. Cloned human teratoma cells differentiate into neuron-like cells and other cell types in retinoic acid. J. Cell Sci. 1984, 72, 37–64. [Google Scholar]
- Matsui, Y.; Zsebo, K.; Hogan, B.L. Derivation of pluripotential embryonic stem cells from murine primordial germ cells in culture. Cell 1992, 70, 841–847. [Google Scholar] [CrossRef]
- Bongso, A.; Fong, C.Y.; Ng, S.C.; Ratnam, S. Isolation and culture of inner cell mass cells from human blastocysts. Hum. Reprod. Oxf. Engl. 1994, 9, 2110–2117. [Google Scholar] [CrossRef]
- Shamblott, M.J.; Axelman, J.; Wang, S.; Bugg, E.M.; Littlefield, J.W.; Donovan, P.J.; Blumenthal, P.D.; Huggins, G.R.; Gearhart, J.D. Derivation of pluripotent stem cells from cultured human primordial germ cells. Dev. Biol. 1998, 95, 13726–13731. [Google Scholar] [CrossRef]
- Thomson, J.A. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Richards, M.; Fong, C.-Y.; Chan, W.-K.; Wong, P.-C.; Bongso, A. Human feeders support prolonged undifferentiated growth of human inner cell masses and embryonic stem cells. Nat. Biotechnol. 2002, 20, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Lebkowski, J. GRNOPC1: The world’s first embryonic stem cell-derived therapy. Regen. Med. 2011, 6, 11–13. [Google Scholar] [CrossRef]
- Okamura, R.M.; Lebkowski, J.; Au, M.; Priest, C.A.; Denham, J.; Majumdar, A.S. Immunological properties of human embryonic stem cell-derived oligodendrocyte progenitor cells. J. Neuroimmunol. 2007, 192, 134–144. [Google Scholar] [CrossRef]
- Manley, N.C.; Priest, C.A.; Denham, J.; Wirth, E.D.; Lebkowski, J.S. Human Embryonic Stem Cell-Derived Oligodendrocyte Progenitor Cells: Preclinical Efficacy and Safety in Cervical Spinal Cord Injury: hESC-Derived OPCs for Cervical Spinal Cord Injury. Stem Cells Transl. Med. 2017, 6, 1917–1929. [Google Scholar] [CrossRef]
- Pera, M.F.; Reubinoff, B.; Trounson, A. Human embryonic stem cells. J. Cell Sci. 2000, 113, 5–10. [Google Scholar]
- Meinhardt, A.; Eberle, D.; Tazaki, A.; Ranga, A.; Niesche, M.; Wilsch-Bräuninger, M.; Stec, A.; Schackert, G.; Lutolf, M.; Tanaka, E.M. 3D Reconstitution of the Patterned Neural Tube from Embryonic Stem Cells. Stem Cell Rep. 2014, 3, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.-A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Ao, H.; Liu, S.; Xu, Y.; McDonald, J.W.; Belegu, V. Spinal cord organogenesis model reveals role of Flk1+ cells in self-organization of neural progenitor cells into complex spinal cord tissue. Stem Cell Res. 2018, 33, 156–165. [Google Scholar] [CrossRef]
- Gurdon, J.B. The Developmental Capacity of Nuclei taken from Intestinal Epithelium Cells of Feeding Tadpoles. J. Embryol. Exp. Morphol. 1962, 10, 622–640. [Google Scholar] [PubMed]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H.S. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810. [Google Scholar] [CrossRef] [PubMed]
- Fulka, J.; Langerova, A.; Loi, P.; Ptak, G.; Albertini, D.; Fulka, H. The ups and downs of somatic cell nucleus transfer (SCNT) in humans. J. Assist. Reprod. Genet. 2013, 30, 1055–1058. [Google Scholar] [CrossRef][Green Version]
- Sagi, I.; Chia, G.; Golan-Lev, T.; Peretz, M.; Weissbein, U.; Sui, L.; Sauer, M.V.; Yanuka, O.; Egli, D.; Benvenisty, N. Derivation and differentiation of haploid human embryonic stem cells. Nature 2016, 532, 107–111. [Google Scholar] [CrossRef]
- Malinowski, A.R.; Fisher, A.G. Reprogramming of Somatic Cells Towards Pluripotency by Cell Fusion. In Polycomb Group Proteins; Lanzuolo, C., Bodega, B., Eds.; Springer: New York, NY, USA, 2016; Volume 1480, pp. 289–299. ISBN 978-1-4939-6378-2. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Conrad, S.; Renninger, M.; Hennenlotter, J.; Wiesner, T.; Just, L.; Bonin, M.; Aicher, W.; Bühring, H.-J.; Mattheus, U.; Mack, A.; et al. Generation of pluripotent stem cells from adult human testis. Nature 2008, 456, 344–349. [Google Scholar] [CrossRef]
- Müller, R.; Lengerke, C. Patient-specific pluripotent stem cells: Promises and challenges. Nat. Rev. Endocrinol. 2009, 5, 195–203. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, Y.; Xie, Y.; Zhang, L.; Tang, P. Cell Transplantation for Spinal Cord Injury: Tumorigenicity of Induced Pluripotent Stem Cell-Derived Neural Stem/Progenitor Cells. Stem Cells Int. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lund, R.J.; Närvä, E.; Lahesmaa, R. Genetic and epigenetic stability of human pluripotent stem cells. Nat. Rev. Genet. 2012, 13, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Guhr, A.; Kobold, S.; Seltmann, S.; Seiler Wulczyn, A.E.M.; Kurtz, A.; Löser, P. Recent Trends in Research with Human Pluripotent Stem Cells: Impact of Research and Use of Cell Lines in Experimental Research and Clinical Trials. Stem Cell Rep. 2018, 11, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Park, I.-H.; Zhao, R.; West, J.A.; Yabuuchi, A.; Huo, H.; Ince, T.A.; Lerou, P.H.; Lensch, M.W.; Daley, G.Q. Reprogramming of human somatic cells to pluripotency with defined factors. Nature 2008, 451, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Aasen, T.; Raya, A.; Barrero, M.J.; Garreta, E.; Consiglio, A.; Gonzalez, F.; Vassena, R.; Bilić, J.; Pekarik, V.; Tiscornia, G.; et al. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes. Nat. Biotechnol. 2008, 26, 1276–1284. [Google Scholar] [CrossRef]
- Aoi, T.; Yae, K.; Nakagawa, M.; Ichisaka, T.; Okita, K.; Takahashi, K.; Chiba, T.; Yamanaka, S. Generation of Pluripotent Stem Cells from Adult Mouse Liver and Stomach Cells. Science 2008, 321, 699–702. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Takahashi, K.; Ichisaka, T.; Aoi, T.; Okita, K.; Mochiduki, Y.; Takizawa, N.; Yamanaka, S. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 2008, 26, 101–106. [Google Scholar] [CrossRef]
- Okita, K.; Nakagawa, M.; Hyenjong, H.; Ichisaka, T.; Yamanaka, S. Generation of Mouse Induced Pluripotent Stem Cells Without Viral Vectors. Science 2008, 322, 949–953. [Google Scholar] [CrossRef]
- Stadtfeld, M.; Nagaya, M.; Utikal, J.; Weir, G.; Hochedlinger, K. Induced Pluripotent Stem Cells Generated Without Viral Integration. Science 2008, 322, 945–949. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell–Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Kavyasudha, C.; Macrin, D.; ArulJothi, K.N.; Joseph, J.P.; Harishankar, M.K.; Devi, A. Clinical Applications of Induced Pluripotent Stem Cells – Stato Attuale. In Cell Biology and Translational Medicine, Volume 1; Turksen, K., Ed.; Springer International Publishing: Cham, Switzerland, 2018; Volume 1079, pp. 127–149. [Google Scholar]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Seki, T.; Yuasa, S.; Fukuda, K. Generation of induced pluripotent stem cells from a small amount of human peripheral blood using a combination of activated T cells and Sendai virus. Nat. Protoc. 2012, 7, 718–728. [Google Scholar] [CrossRef]
- Soldner, F.; Jaenisch, R. iPSC Disease Modeling. Science 2012, 338, 1155–1156. [Google Scholar] [CrossRef]
- Ebert, A.D.; Yu, J.; Rose, F.F.; Mattis, V.B.; Lorson, C.L.; Thomson, J.A.; Svendsen, C.N. Induced pluripotent stem cells from a spinal muscular atrophy patient. Nature 2009, 457, 277–280. [Google Scholar] [CrossRef]
- Marchetto, M.C.N.; Carromeu, C.; Acab, A.; Yu, D.; Yeo, G.W.; Mu, Y.; Chen, G.; Gage, F.H.; Muotri, A.R. A Model for Neural Development and Treatment of Rett Syndrome Using Human Induced Pluripotent Stem Cells. Cell 2010, 143, 527–539. [Google Scholar] [CrossRef]
- Raya, Á.; Rodríguez-Pizà, I.; Guenechea, G.; Vassena, R.; Navarro, S.; Barrero, M.J.; Consiglio, A.; Castellà, M.; Río, P.; Sleep, E.; et al. Disease-corrected haematopoietic progenitors from Fanconi anaemia induced pluripotent stem cells. Nature 2009, 460, 53–59. [Google Scholar] [CrossRef]
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef]
- Song, J.; Olsen, R.H.J.; Sun, J.; Ming, G.; Song, H. Neuronal Circuitry Mechanisms Regulating Adult Mammalian Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018937. [Google Scholar] [CrossRef]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef]
- Weiss, S.; Dunne, C.; Hewson, J.; Wohl, C.; Wheatley, M.; Peterson, A.C.; Reynolds, B.A. Multipotent CNS Stem Cells Are Present in the Adult Mammalian Spinal Cord and Ventricular Neuroaxis. J. Neurosci. 1996, 16, 7599–7609. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.; Weiss, S. Generation of neurons and astrocytes from isolated cells of the adult mammalian central nervous system. Science 1992, 255, 1707–1710. [Google Scholar] [CrossRef]
- Morshead, C.M.; Reynolds, B.A.; Craig, C.G.; McBurney, M.W.; Staines, W.A.; Morassutti, D.; Weiss, S.; van der Kooy, D. Neural stem cells in the adult mammalian forebrain: A relatively quiescent subpopulation of subependymal cells. Neuron 1994, 13, 1071–1082. [Google Scholar] [CrossRef]
- Zhang, S.-C.; Wernig, M.; Duncan, I.D.; Brüstle, O.; Thomson, J.A. In vitro differentiation of transplantable neural precursors from human embryonic stem cells. Nat. Biotechnol. 2001, 19, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Kozhich, O.A.; Hamilton, R.S.; Mallon, B.S. Standardized Generation and Differentiation of Neural Precursor Cells from Human Pluripotent Stem Cells. Stem Cell Rev. Rep. 2013, 9, 531–536. [Google Scholar] [CrossRef]
- Meneghini, V.; Frati, G.; Sala, D.; De Cicco, S.; Luciani, M.; Cavazzin, C.; Paulis, M.; Mentzen, W.; Morena, F.; Giannelli, S.; et al. Generation of Human Induced Pluripotent Stem Cell-Derived Bona Fide Neural Stem Cells for Ex Vivo Gene Therapy of Metachromatic Leukodystrophy: iPSC-Derived NSCs for CNS Ex Vivo Gene Therapy. Stem Cells Transl. Med. 2017, 6, 352–368. [Google Scholar] [CrossRef]
- Banda, E.; Grabel, L. Directed Differentiation of Human Embryonic Stem Cells into Neural Progenitors. In Human Embryonic Stem Cell Protocols; Turksen, K., Ed.; Springer: New York, NY, USA, 2014; Volume 1307, pp. 289–298. [Google Scholar]
- Wen, Y.; Jin, S. Production of neural stem cells from human pluripotent stem cells. J. Biotechnol. 2014, 188, 122–129. [Google Scholar] [CrossRef]
- Bauwens, C.L.; Peerani, R.; Niebruegge, S.; Woodhouse, K.A.; Kumacheva, E.; Husain, M.; Zandstra, P.W. Control of Human Embryonic Stem Cell Colony and Aggregate Size Heterogeneity Influences Differentiation Trajectories. Stem Cells 2008, 26, 2300–2310. [Google Scholar] [CrossRef]
- Hong, S.-H.; Werbowetski-Ogilvie, T.; Ramos-Mejia, V.; Lee, J.B.; Bhatia, M. Multiparameter comparisons of embryoid body differentiation toward human stem cell applications. Stem Cell Res. 2010, 5, 120–130. [Google Scholar] [CrossRef]
- Sheridan, S.D.; Surampudi, V.; Rao, R.R. Analysis of Embryoid Bodies Derived from Human Induced Pluripotent Stem Cells as a Means to Assess Pluripotency. Stem Cells Int. 2012, 2012, 1–9. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of Stem Cell Fate by Physical Interactions with the Extracellular Matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef]
- Knight, E.; Przyborski, S. Advances in 3D cell culture technologies enabling tissue-like structures to be created in vitro. J. Anat. 2015, 227, 746–756. [Google Scholar] [CrossRef]
- Aghajanian, P.; Mohan, S. The art of building bone: Emerging role of chondrocyte-to-osteoblast transdifferentiation in endochondral ossification. Bone Res. 2018, 6, 19. [Google Scholar] [CrossRef]
- Selman, K.; Kafatos, F.C. Transdifferentiation in the labial gland of silk moths: Is DNA required for cellular metamorphosis? Cell Differ. 1974, 3, 81–94. [Google Scholar] [CrossRef]
- Xiao, D.; Liu, X.; Zhang, M.; Zou, M.; Deng, Q.; Sun, D.; Bian, X.; Cai, Y.; Guo, Y.; Liu, S.; et al. Direct reprogramming of fibroblasts into neural stem cells by single non-neural progenitor transcription factor Ptf1a. Nat. Commun. 2018, 9, 2865. [Google Scholar] [CrossRef]
- Christoforou, N.; Chakraborty, S.; Kirkton, R.D.; Adler, A.F.; Addis, R.C.; Leong, K.W. Core Transcription Factors, MicroRNAs and Small Molecules Drive Transdifferentiation of Human Fibroblasts towards the Cardiac Cell Lineage. Sci. Rep. 2017, 7, 40285. [Google Scholar] [CrossRef]
- Yao, H.; Gao, M.; Ma, J.; Zhang, M.; Li, S.; Wu, B.; Nie, X.; Jiao, J.; Zhao, H.; Wang, S.; et al. Transdifferentiation-Induced Neural Stem Cells Promote Recovery of Middle Cerebral Artery Stroke Rats. PLoS ONE 2015, 10, e0137211. [Google Scholar] [CrossRef]
- Cheng, L.; Hu, W.; Qiu, B.; Zhao, J.; Yu, Y.; Guan, W.; Wang, M.; Yang, W.; Pei, G. Generation of neural progenitor cells by chemical cocktails and hypoxia. Cell Res. 2014, 24, 665–679. [Google Scholar] [CrossRef]
- Feng, N.; Han, Q.; Li, J.; Wang, S.; Li, H.; Yao, X.; Zhao, R.C. Generation of Highly Purified Neural Stem Cells from Human Adipose-Derived Mesenchymal Stem Cells by Sox1 Activation. Stem Cells Dev. 2014, 23, 515–529. [Google Scholar] [CrossRef]
- Merrell, A.J.; Stanger, B.Z. Adult cell plasticity in vivo: De-differentiation and transdifferentiation are back in style. Nat. Rev. Mol. Cell Biol. 2016, 17, 413–425. [Google Scholar] [CrossRef]
- McDonald, J.W.; Liu, X.-Z.; Qu, Y.; Liu, S.; Mickey, S.K.; Turetsky, D.; Gottlieb, D.I.; Choi, D.W. Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nat. Med. 1999, 5, 1410–1412. [Google Scholar] [CrossRef] [PubMed]
- Kumamaru, H.; Saiwai, H.; Kubota, K.; Kobayakawa, K.; Yokota, K.; Ohkawa, Y.; Shiba, K.; Iwamoto, Y.; Okada, S. Therapeutic Activities of Engrafted Neural Stem/Precursor Cells Are Not Dormant in the Chronically Injured Spinal Cord: Cell Activity of NSPCs Transplanted in Chronic SCI. Stem Cells 2013, 31, 1535–1547. [Google Scholar] [CrossRef]
- Cheng, I.; Park, D.Y.; Mayle, R.E.; Githens, M.; Smith, R.L.; Park, H.Y.; Hu, S.S.; Alamin, T.F.; Wood, K.B.; Kharazi, A.I. Does timing of transplantation of neural stem cells following spinal cord injury affect outcomes in an animal model? J. Spine Surg. 2017, 3, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.X.; Hooshmand, M.J.; Saiwai, H.; Maddox, J.; Salehi, A.; Lakatos, A.; Nishi, R.A.; Salazar, D.; Uchida, N.; Anderson, A.J. Systemic Neutrophil Depletion Modulates the Migration and Fate of Transplanted Human Neural Stem Cells to Rescue Functional Repair. J. Neurosci. 2017, 37, 9269–9287. [Google Scholar] [CrossRef]
- Salazar, D.L.; Uchida, N.; Hamers, F.P.T.; Cummings, B.J.; Anderson, A.J. Human Neural Stem Cells Differentiate and Promote Locomotor Recovery in an Early Chronic Spinal coRd Injury NOD-scid Mouse Model. PLoS ONE 2010, 5, e12272. [Google Scholar] [CrossRef]
- Cheng, I.; Githens, M.; Smith, R.L.; Johnston, T.R.; Park, D.Y.; Stauff, M.P.; Salari, N.; Tileston, K.R.; Kharazi, A.I. Local versus distal transplantation of human neural stem cells following chronic spinal cord injury. Spine J. 2016, 16, 764–769. [Google Scholar] [CrossRef]
- Tashiro, S.; Nishimura, S.; Iwai, H.; Sugai, K.; Zhang, L.; Shinozaki, M.; Iwanami, A.; Toyama, Y.; Liu, M.; Okano, H.; et al. Functional Recovery from Neural Stem/Progenitor Cell Transplantation Combined with Treadmill Training in Mice with Chronic Spinal Cord Injury. Sci. Rep. 2016, 6, 30898. [Google Scholar] [CrossRef]
- Xiong, L.; Zou, Y.; Shi, Y.; Zhang, P.; Zhang, R.; Dai, X.; Liu, B.; Wang, T. Tree shrew neural stem cell transplantation promotes functional recovery of tree shrews with a hemi‑sectioned spinal cord injury by upregulating nerve growth factor expression. Int. J. Mol. Med. 2018, 41, 3267–3277. [Google Scholar] [CrossRef]
- Riemann, L.; Younsi, A.; Scherer, M.; Zheng, G.; Skutella, T.; Unterberg, A.W.; Zweckberger, K. Transplantation of Neural Precursor Cells Attenuates Chronic Immune Environment in Cervical Spinal Cord Injury. Front. Neurol. 2018, 9, 428. [Google Scholar] [CrossRef]
- Kadoya, K.; Lu, P.; Nguyen, K.; Lee-Kubli, C.; Kumamaru, H.; Yao, L.; Knackert, J.; Poplawski, G.; Dulin, J.N.; Strobl, H.; et al. Spinal cord reconstitution with homologous neural grafts enables robust corticospinal regeneration. Nat. Med. 2016, 22, 479–487. [Google Scholar] [CrossRef]
- Robinson, J.; Lu, P. Optimization of trophic support for neural stem cell grafts in sites of spinal cord injury. Exp. Neurol. 2017, 291, 87–97. [Google Scholar] [CrossRef]
- Hwang, I.; Hahm, S.-C.; Choi, K.-A.; Park, S.-H.; Jeong, H.; Yea, J.-H.; Kim, J.; Hong, S. Intrathecal Transplantation of Embryonic Stem Cell-Derived Spinal GABAergic Neural Precursor Cells Attenuates Neuropathic Pain in a Spinal Cord Injury Rat Model. Cell Transplant. 2016, 25, 593–607. [Google Scholar] [CrossRef]
- Karova, K.; Wainwright, J.V.; Machova-Urdzikova, L.; Pisal, R.V.; Schmidt, M.; Jendelova, P.; Jhanwar-Uniyal, M. Transplantation of neural precursors generated from spinal progenitor cells reduces inflammation in spinal cord injury via NF-κB pathway inhibition. J. Neuroinflammation 2019, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.-J.; Oh, S.-H.; Lee, N.; Choi, C.; Jeon, I.; Kim, H.S.; Shin, D.A.; Lee, S.E.; Kim, D.; Song, J. Contralaterally transplanted human embryonic stem cell-derived neural precursor cells (ENStem-A) migrate and improve brain functions in stroke-damaged rats. Exp. Mol. Med. 2013, 45, 53. [Google Scholar] [CrossRef]
- Lien, B.V.; Tuszynski, M.H.; Lu, P. Astrocytes migrate from human neural stem cell grafts and functionally integrate into the injured rat spinal cord. Exp. Neurol. 2019, 314, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Ceto, S.; Wang, Y.; Graham, L.; Wu, D.; Kumamaru, H.; Staufenberg, E.; Tuszynski, M.H. Prolonged human neural stem cell maturation supports recovery in injured rodent CNS. J. Clin. Investig. 2017, 127, 3287–3299. [Google Scholar] [CrossRef]
- Kumamaru, H.; Kadoya, K.; Adler, A.F.; Takashima, Y.; Graham, L.; Coppola, G.; Tuszynski, M.H. Generation and post-injury integration of human spinal cord neural stem cells. Nat. Methods 2018, 15, 723–731. [Google Scholar] [CrossRef]
- Amemori, T.; Ruzicka, J.; Romanyuk, N.; Jhanwar-Uniyal, M.; Sykova, E.; Jendelova, P. Comparison of intraspinal and intrathecal implantation of induced pluripotent stem cell-derived neural precursors for the treatment of spinal cord injury in rats. Stem Cell Res. Ther. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed]
- Romanyuk, N.; Amemori, T.; Turnovcova, K.; Prochazka, P.; Onteniente, B.; Sykova, E.; Jendelova, P. Beneficial Effect of Human Induced Pluripotent Stem Cell-Derived Neural Precursors in Spinal Cord Injury Repair. Cell Transplant. 2015, 24, 1781–1797. [Google Scholar] [CrossRef]
- Hosseini, M.; Yousefifard, M.; Aziznejad, H.; Nasirinezhad, F. The Effect of Bone Marrow–Derived Mesenchymal Stem Cell Transplantation on Allodynia and Hyperalgesia in Neuropathic Animals: A Systematic Review with Meta-Analysis. Biol. Blood Marrow Transplant. 2015, 21, 1537–1544. [Google Scholar] [CrossRef]
- Yousefifard, M.; Rahimi-Movaghar, V.; Nasirinezhad, F.; Baikpour, M.; Safari, S.; Saadat, S.; Moghadas Jafari, A.; Asady, H.; Razavi Tousi, S.M.T.; Hosseini, M. Neural stem/progenitor cell transplantation for spinal cord injury treatment; A systematic review and meta-analysis. Neuroscience 2016, 322, 377–397. [Google Scholar] [CrossRef]
- Kojima, K.; Miyoshi, H.; Nagoshi, N.; Kohyama, J.; Itakura, G.; Kawabata, S.; Ozaki, M.; Iida, T.; Sugai, K.; Ito, S.; et al. Selective Ablation of Tumorigenic Cells Following Human Induced Pluripotent Stem Cell-Derived Neural Stem/Progenitor Cell Transplantation in Spinal Cord Injury: Suicide System in hiPSC-NS/PC Therapy. Stem Cells Transl. Med. 2019, 8, 260–270. [Google Scholar] [CrossRef]
- Miura, K.; Okada, Y.; Aoi, T.; Okada, A.; Takahashi, K.; Okita, K.; Nakagawa, M.; Koyanagi, M.; Tanabe, K.; Ohnuki, M.; et al. Variation in the safety of induced pluripotent stem cell lines. Nat. Biotechnol. 2009, 27, 743–745. [Google Scholar] [CrossRef]
- Roy, N.S.; Cleren, C.; Singh, S.K.; Yang, L.; Beal, M.F.; Goldman, S.A. Functional engraftment of human ES cell–derived dopaminergic neurons enriched by coculture with telomerase-immortalized midbrain astrocytes. Nat. Med. 2006, 12, 1259–1268. [Google Scholar] [CrossRef]
- Itakura, G.; Kawabata, S.; Ando, M.; Nishiyama, Y.; Sugai, K.; Ozaki, M.; Iida, T.; Ookubo, T.; Kojima, K.; Kashiwagi, R.; et al. Fail-Safe System against Potential Tumorigenicity after Transplantation of iPSC Derivatives. Stem Cell Rep. 2017, 8, 673–684. [Google Scholar] [CrossRef]
- Kim, J.B.; Zaehres, H.; Wu, G.; Gentile, L.; Ko, K.; Sebastiano, V.; Araúzo-Bravo, M.J.; Ruau, D.; Han, D.W.; Zenke, M.; et al. Pluripotent stem cells induced from adult neural stem cells by reprogramming with two factors. Nature 2008, 454, 646–650. [Google Scholar] [CrossRef]
- Cheng, I.; Mayle, R.E.; Cox, C.A.; Park, D.Y.; Smith, R.L.; Corcoran-Schwartz, I.; Ponnusamy, K.E.; Oshtory, R.; Smuck, M.W.; Mitra, R.; et al. Functional assessment of the acute local and distal transplantation of human neural stem cells after spinal cord injury. Spine J. 2012, 12, 1040–1044. [Google Scholar] [CrossRef]
- Bottai, D.; Madaschi, L.; Giulio, A.M.D.; Gorio, A. Viability-Dependent Promoting Action of Adult Neural Precursors in Spinal Cord Injury. Mol. Med. 2008, 14, 634–644. [Google Scholar] [CrossRef]
- Nishimura, S.; Yasuda, A.; Iwai, H.; Takano, M.; Kobayashi, Y.; Nori, S.; Tsuji, O.; Fujiyoshi, K.; Ebise, H.; Toyama, Y.; et al. Time-dependent changes in the microenvironment of injured spinal cord affects the therapeutic potential of neural stem cell transplantation for spinal cord injury. Mol. Brain 2013, 6, 3. [Google Scholar] [CrossRef]
- Osaka, M.; Honmou, O.; Murakami, T.; Nonaka, T.; Houkin, K.; Hamada, H.; Kocsis, J.D. Intravenous administration of mesenchymal stem cells derived from bone marrow after contusive spinal cord injury improves functional outcome. Brain Res. 2010, 1343, 226–235. [Google Scholar] [CrossRef]
- Okubo, T.; Nagoshi, N.; Kohyama, J.; Tsuji, O.; Shinozaki, M.; Shibata, S.; Kase, Y.; Matsumoto, M.; Nakamura, M.; Okano, H. Treatment with a Gamma-Secretase Inhibitor Promotes Functional Recovery in Human iPSC- Derived Transplants for Chronic Spinal Cord Injury. Stem Cell Rep. 2018, 11, 1416–1432. [Google Scholar] [CrossRef]
- Ruzicka, J.; Romanyuk, N.; Jirakova, K.; Hejcl, A.; Janouskova, O.; Machova, L.U.; Bochin, M.; Pradny, M.; Vargova, L.; Jendelova, P. The Effect of iPS-Derived Neural Progenitors Seeded on Laminin-Coated pHEMA-MOETACl Hydrogel with Dual Porosity in a Rat Model of Chronic Spinal Cord Injury. Cell Transplant. 2019. [Google Scholar] [CrossRef]
- Pomeshchik, Y.; Puttonen, K.A.; Kidin, I.; Ruponen, M.; Lehtonen, S.; Malm, T.; Åkesson, E.; Hovatta, O.; Koistinaho, J. Transplanted Human Induced Pluripotent Stem Cell-Derived Neural Progenitor Cells Do Not Promote Functional Recovery of Pharmacologically Immunosuppressed Mice with Contusion Spinal Cord Injury. Cell Transplant. 2015, 24, 1799–1812. [Google Scholar] [CrossRef]
- Ghobrial, G.M.; Anderson, K.D.; Dididze, M.; Martinez-Barrizonte, J.; Sunn, G.H.; Gant, K.L.; Levi, A.D. Human Neural Stem Cell Transplantation in Chronic Cervical Spinal Cord Injury: Functional Outcomes at 12 Months in a Phase II Clinical Trial. Neurosurgery 2017, 64, 87–91. [Google Scholar] [CrossRef]
- Okubo, T.; Iwanami, A.; Kohyama, J.; Itakura, G.; Kawabata, S.; Nishiyama, Y.; Sugai, K.; Ozaki, M.; Iida, T.; Matsubayashi, K.; et al. Pretreatment with a γ-Secretase Inhibitor Prevents Tumor-like Overgrowth in Human iPSC-Derived Transplants for Spinal Cord Injury. Stem Cell Rep. 2016, 7, 649–663. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhu, W.; Cao, K.; Wu, F.; Li, J.; Wang, G.; Li, H.; Lu, M.; Ren, Y.; He, X. Anti-Inflammatory Mechanism of Neural Stem Cell Transplantation in Spinal Cord Injury. Int. J. Mol. Sci. 2016, 17, 1380. [Google Scholar] [CrossRef]
- Salewski, R.P.; Mitchell, R.A.; Shen, C.; Fehlings, M.G. Transplantation of Neural Stem Cells Clonally Derived from Embryonic Stem Cells Promotes Recovery After Murine Spinal Cord Injury. Stem Cells Dev. 2015, 24, 36–50. [Google Scholar] [CrossRef]
- Jin, Y.; Bouyer, J.; Shumsky, J.S.; Haas, C.; Fischer, I. Transplantation of neural progenitor cells in chronic spinal cord injury. Neuroscience 2016, 320, 69–82. [Google Scholar] [CrossRef]
- Cohrs, R.J.; Martin, T.; Ghahramani, P.; Bidaut, L.; Higgins, P.J.; Shahzad, A. Translational Medicine definition by the European Society for Translational Medicine. Eur. J. Mol. Clin. Med. 2014, 2, 86. [Google Scholar] [CrossRef]
- Barreau, K.; Lépinoux-Chambaud, C. Review of Clinical Trials Using Neural Stem Cells. JSM Biotechnol. Bioeng. 2016, 3, 1057. [Google Scholar]
- Sharif-Alhoseini, M.; Khormali, M.; Rezaei, M.; Safdarian, M.; Hajighadery, A.; Khalatbari, M.M.; Safdarian, M.; Meknatkhah, S.; Rezvan, M.; Chalangari, M.; et al. Animal models of spinal cord injury: A systematic review. Spinal Cord 2017, 55, 714–721. [Google Scholar] [CrossRef]
- Shin, J.C.; Kim, K.N.; Yoo, J.; Kim, I.-S.; Yun, S.; Lee, H.; Jung, K.; Hwang, K.; Kim, M.; Lee, I.-S.; et al. Clinical Trial of Human Fetal Brain-Derived Neural Stem/Progenitor Cell Transplantation in Patients with Traumatic Cervical Spinal Cord Injury. Neural Plast. 2015, 2015, 1–22. [Google Scholar] [CrossRef]
- Long-Term Follow-Up of Transplanted Human Central Nervous System Stem Cells (HuCNS-SC) in Spinal Cord Trauma Subjects. Available online: https://clinicaltrials.gov/ct2/show/NCT01725880 (accessed on 12 March 2019).
- Curtis, E.; Martin, J.R.; Gabel, B.; Sidhu, N.; Rzesiewicz, T.K.; Mandeville, R.; Van Gorp, S.; Leerink, M.; Tadokoro, T.; Marsala, S.; et al. A First-in-Human, Phase I Study of Neural Stem Cell Transplantation for Chronic Spinal Cord Injury. Cell Stem Cell 2018, 22, 941–950. [Google Scholar] [CrossRef]
- Study of Human Central Nervous System (CNS) Stem Cell Transplantation in Cervical Spinal Cord Injury. Available online: https://clinicaltrials.gov/ct2/show/NCT02163876 (accessed on 12 March 2019).
- INSTrUCT-SCI: INdependent Observational STUdy of Cell Transplantation in SCI. Available online: https://clinicaltrials.gov/ct2/show/NCT03069404 (accessed on 6 April 2019).
- Neural Stem Cell Transplantation in Traumatic Spinal Cord Injury. Available online: https://clinicaltrials.gov/ct2/show/NCT02326662 (accessed on 12 March 2019).
- NeuroRegen ScaffoldTM Combined With Stem Cells for Chronic Spinal Cord Injury Repair. Available online: https://clinicaltrials.gov/ct2/show/NCT02688049 (accessed on 12 March 2019).
- Safety Study of GRNOPC1 in Spinal Cord Injury. Available online: https://clinicaltrials.gov/ct2/show/NCT01217008 (accessed on 12 March 2019).
- Dose Escalation Study of AST-OPC1 in Spinal Cord Injury. Available online: https://clinicaltrials.gov/ct2/show/NCT02302157 (accessed on 12 March 2019).
- Piltti, K.M.; Avakian, S.N.; Funes, G.M.; Hu, A.; Uchida, N.; Anderson, A.J.; Cummings, B.J. Transplantation dose alters the dynamics of human neural stem cell engraftment, proliferation and migration after spinal cord injury. Stem Cell Res. 2015, 15, 341–353. [Google Scholar] [CrossRef]
- Sontag, C.J.; Nguyen, H.X.; Kamei, N.; Uchida, N.; Anderson, A.J.; Cummings, B.J. Immunosuppressants Affect Human Neural Stem Cells In Vitro but Not in an In Vivo Model of Spinal Cord Injury. Stem Cells Transl. Med. 2013, 2, 731–744. [Google Scholar] [CrossRef]
- Sontag, C.J.; Uchida, N.; Cummings, B.J.; Anderson, A.J. Injury to the Spinal Cord Niche Alters the Engraftment Dynamics of Human Neural Stem Cells. Stem Cell Rep. 2014, 2, 620–632. [Google Scholar] [CrossRef]
- Levi, A.D.; Okonkwo, D.O.; Park, P.; Jenkins, A.L.; Kurpad, S.N.; Parr, A.M.; Ganju, A.; Aarabi, B.; Kim, D.; Casha, S.; et al. Emerging Safety of Intramedullary Transplantation of Human Neural Stem Cells in Chronic Cervical and Thoracic Spinal Cord Injury. Neurosurgery 2018, 82, 562–575. [Google Scholar] [CrossRef]
- Levi, A.D.; Anderson, K.D.; Okonkwo, D.O.; Park, P.; Bryce, T.N.; Kurpad, S.N.; Aarabi, B.; Hsieh, J.; Gant, K. Clinical Outcomes from a Multi-Center Study of Human Neural Stem Cell Transplantation in Chronic Cervical Spinal Cord Injury. J. Neurotrauma 2019, 36, 891–902. [Google Scholar] [CrossRef]
- Tsuji, O.; Sugai, K.; Yamaguchi, R.; Tashiro, S.; Nagoshi, N.; Kohyama, J.; Iida, T.; Ohkubo, T.; Itakura, G.; Isoda, M.; et al. Concise Review: Laying the Groundwork for a First-In-Human Study of an Induced Pluripotent Stem Cell-Based Intervention for Spinal Cord Injury: iPSC-Based Intervention for Spinal Cord Injury. Stem Cells 2019, 37, 6–13. [Google Scholar] [CrossRef]
- Nagoshi, N.; Okano, H. iPSC-derived neural precursor cells: Potential for cell transplantation therapy in spinal cord injury. Cell. Mol. Life Sci. 2018, 75, 989–1000. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Ha, K.-Y.; Kim, S.-I. Spinal Cord Injury and Related Clinical Trials. Clin. Orthop. Surg. 2017, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Dalamagkas, K.; Tsintou, M.; Seifalian, A.; Seifalian, A. Translational Regenerative Therapies for Chronic Spinal Cord Injury. Int. J. Mol. Sci. 2018, 19, 1776. [Google Scholar] [CrossRef] [PubMed]
- Badhiwala, J.H.; Ahuja, C.S.; Fehlings, M.G. Time is spine: A review of translational advances in spinal cord injury. J. Neurosurg. Spine 2019, 30, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Wang, J.; Lin, X.; Zhang, L. Stem cell transplantation for spinal cord injury: A meta-analysis of treatment effectiveness and safety. Neural Regen. Res. 2017, 12, 815. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Advantages | Limitations |
|---|---|
| No ethical concerns | Risk of tumor formation |
| Evidence on a pluripotent profile (ES-like cells) | Epigenetic and genetic alterations |
| Highly flexible technique | Oncogenes reactivation |
| Derivation from any somatic cell | Expensive and time-consuming |
| Patient-specific source (no immune rejection) |
| SCI | Animal Model | Injury | Transplanted Cells | Time | Additional Treatments | Outcomes | REF |
|---|---|---|---|---|---|---|---|
| Transection | Fischer 344 Rat C57BL/6 Mice | T3 C4 | Rat E14 SC-derived NPCs Mouse E12 SC-derived cells | 2 w | Cell grafts survival Full-fill of the cavitation site Axonal CST regeneration and functional synaptic formation Improved forelimb function | [144] | |
| Contusion | C57BL/6 Mice | T9/10 | Mouse Fetal Brain NSCs | 1 w | Migration from the injection site toward the injury Locomotor improvement Reduction in neutrophils and iNOS+/Mac-2+ cells Downregulation of TNF-α, IL-1β, IL-6 and IL-12 | [170] | |
| Hemisection | Fischer 344 Rat | C5 | Rat E14 SC-derived NPCs | 2 w | 4-factor cocktail | Consistent graft survival Neuronal differentiation Reduction of the lesion site | [145] |
| Compression | Wistar Rat | T10 | Human Fetal Spinal Cord SPC-01 cell line | 1 w | Downregulation of TNF-α Inhibition of p65 NF-κB Reduction of glial scar and cavity size Gray matter preservation | [147] | |
| Compression | C57BL/6 Mice | T6 | ES-dNSC | 1 w | Enhancement of spared neural tissue Differentiation into oligodendrocytes Motor improvement | [171] | |
| Hemisection | Nude Rat | C5 | H9 ESC-derived NSCs | 2 w | Graft size stable over time Differentiation into mature neurons and glia Long axonal regrowth Glial migration to host white matter | [150] | |
| Contusion | NOD-scid Mice | T9 | hCNS-derived NSCs | 0 | Astroglial differentiation of donor cells in the lesion site No locomotor recovery | [138] | |
| Transection | Nude Rat | C4 | hPSC-derived Spinal Cord NSCs | 2 w | NSCs committed to a spinal cord phenotype Differentiation into excitatory neurons Regeneration of the CST Host-to-graft synaptic connectivity | [151] | |
| Compression | WT Mouse C3Fe.SWV-Mbpshi/J Mice | T6 | iPS-derived NSCs | 1 w | Integration within the lesion site Differentiation to oligodendrocytes wt-iPS-dNSCs promote remyelination and axonal function Motor Improvements | [53] | |
| Compression | Wistar Rat | T8 | iPS-derived NPs | 1 w | Intraspinal implantation promote: > gray and white matter sparing > axonal sprouting > astrogliosis reduction Moderate functional recovery | [152] | |
| Compression | Wistar Rat | T8/T9 | hiPSC-derived NPs | 1 w | Cell survival and tissue preservation Differentiation into the three germ layers Motor improvement Increased expression of NFs Neuronal regeneration | [153] | |
| Contusion | C57BL/6 Mice | T10 | iPSC-derived NPCs | 1 w | Neuronal lineage differentiation No tumor formation No locomotor recovery | [167] | |
| Contusion | Long-Evans hooded Rat | T10 | Human Fetal Brain NSCs | 4 w | Trophic effect in the CSF Motor improvement | [140] | |
| Contusion | C57BL/6 Mice | T9 | Mouse Striatal NS/PCs | 7–10 d | Treadmill Training | Differentiation into neurons, oligodendrocytes, and astrocytes Electrophysiologic recovery Locomotor improvements | [141] |
| Contusion | Rat | T10 | Rat Spinal Cord NSCs | 13 w | Ch combined with NFs | 60% of survival < 40% of the lesion site covered Improvement in bladder function | [172] |
| Hemisection | Tree Shrew | T10 | Shrew Fetal NSCs | 9 d | Self-renewal potential Differentiation into neurons and astrocytes Production of NFs (CNTF, TGF-β1, GDNF, NGF, BDNF and IGF) | [142] | |
| Contusion | Wistar Rat | C6/C7 | Rat Fetal Brain NSCs | 10 d | Long-term survival Differentiation along the oligodendroglial lineage Reduction in M1 macrophages Lower density of iNOS Functional recovery Reduction in apoptosis | [143] | |
| Contusion | Sprague–Dawley Rat | T12 | mESC-derived NPCs | 3 w | In vitro differentiation into a spinal GABAergic phenotype Attenuation of chronic neuropathic pain | [146] | |
| Hemisection | Nude Rat | C5 | Human H9 ESC-derived NSCs | 2 w | No cellular migration Improvement in skilled forelimb motor function | [149] | |
| Contusion | C57BL/6 Mice | C6/C7 | iPS-derived NSCs | 8 w | Intrathecal ChABC | Cell survival Remyelination and synaptic formation Behavioral recovery of the forelimb grip strength and locomotion | [27] |
| Contusion | NOD-SCID Mice | T10 | hiPSC-derived NS/PCs | 6 w | GSI | Axonal regrowth and remyelination Reticulo-spinal tract fiber formation Motor functional recovery | [165] |
| Compression | Wistar Rat | T8/T8 | hiPSC-derived NS/PCs | 5 w | Laminin-coated pHEMA-MOETACl hydrogel | Survival and integration within the lesion spinal cord Reduction in cavity depth and axonal growth Increased number of astrocytes, blood vessels, and TH+ fibers No locomotor recovery | [166] |
| Start Year | Sponsor | Country | NTC/I.D. | Clinical Phase | SCI Cohort | Cell-Type | Cell Source | Safety | Improvements | Others | REF |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2005 | Yonsei University Health System, Severance Hospital | KR | KCT0000879 | Phase I/II | Cervical | hNSPCs | Fetal brain | Safe and well-tolerable | Partial sensorimotor function | No cord damage, syrinx or tumor formation No neurological deterioration, and exacerbating neuropathic pain or spasticity Incomplete sensory recovery | [176] |
| 2011 | StemCells, Inc. | CAN CH | NCT01321333 | Phase I/II | T2-T11 | HuCNS-SCs | Fetal brain | Safe and well-tolerable | Segmental sensory | Decline in sensory gains lost after withdrawal of the immunosuppressive | [168] |
| 2012 | StemCells, Inc. | CH | NCT01725880 | Phase I/II | T2-T11 | HuCNS-SCs | Fetal brain | Study terminated based on a business decision | [177] | ||
| 2013 | Neuralstem Inc. | US | NCT01772810 | Phase I | T2-T12 | NSI-566 cell line | Fetal spinal cord (cervical and upper thoracic regions) | Safe and no side effects 18–27 months after cell delivery | Low sample size (n = 4) Still Recruiting | [178] | |
| 2014 | StemCells, Inc. | US CAN | NCT02163876 | Phase I/II | C5-C7 | HuCNS-SCs | Fetal brain | Slight motor strength but the study was terminated based on a business decision | [179] | ||
| 2017 | University of Zurich | CH | NCT03069404 | Phase I/II | T2-T11 | HuCNS-SCs | Fetal brain | No data | [180] | ||
| 2014 | Federal Research Clinical Center of Federal Medical & Biological Agency | RU | NCT02326662 | Phase I/II | Neck, thoracic or lumbar | drNSCs | BMCs | Safe with any complications | Neurologic state | [181] | |
| 2016 | Chinese Academy of Sciences | CN | NCT02688049 | Phase I/II | C5-T12 | NSCs | No data Still recruiting | [182] | |||
| 2010 | Asterias Biotherapeutics | US | NCT01217008 | Phase I | Neurologically Complete, Subacute | GRNOPC1 | hESCs | The study was terminated based on financial issues | [183] | ||
| 2015 | Asterias Biotherapeutics | US | NCT02302157 | Phase I/II | C4-C7 | AST-OPC1 | hESCs | Favorable safety profile | Some hand functions | [184] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, I.M.; Marote, A.; Salgado, A.J.; Silva, N.A. Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury. Pharmaceuticals 2019, 12, 65. https://doi.org/10.3390/ph12020065
Pereira IM, Marote A, Salgado AJ, Silva NA. Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury. Pharmaceuticals. 2019; 12(2):65. https://doi.org/10.3390/ph12020065
Chicago/Turabian StylePereira, Inês M., Ana Marote, António J. Salgado, and Nuno A. Silva. 2019. "Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury" Pharmaceuticals 12, no. 2: 65. https://doi.org/10.3390/ph12020065
APA StylePereira, I. M., Marote, A., Salgado, A. J., & Silva, N. A. (2019). Filling the Gap: Neural Stem Cells as A Promising Therapy for Spinal Cord Injury. Pharmaceuticals, 12(2), 65. https://doi.org/10.3390/ph12020065