A Syringe-Based Biosensor to Rapidly Detect Low Levels of Escherichia Coli (ECOR13) in Drinking Water Using Engineered Bacteriophages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials & Methods
2.1. Materials & Reagents
2.2. Bacterial Strains & Growth Conditions
2.3. DNA Isolation
2.4. Recombinant Phage Construction
2.5. Phage Characterization
2.6. Dose Response
2.7. Infection of Non-Viable Cells
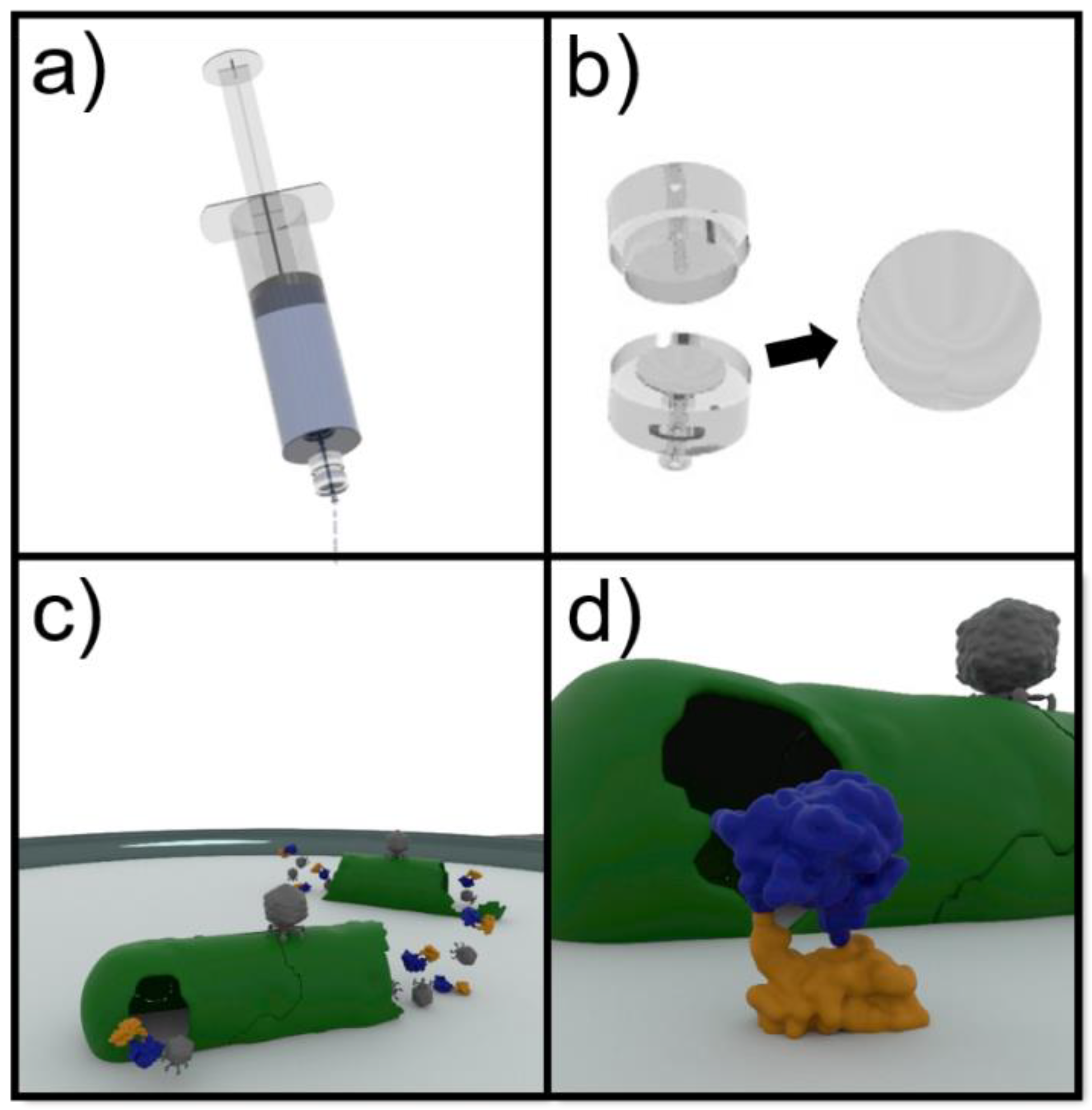
2.8. Phage-Based Syringe Filter Detection Assay
3. Results & Discussion
3.1. Recombinant Phage Construction
3.2. Phage Characterization
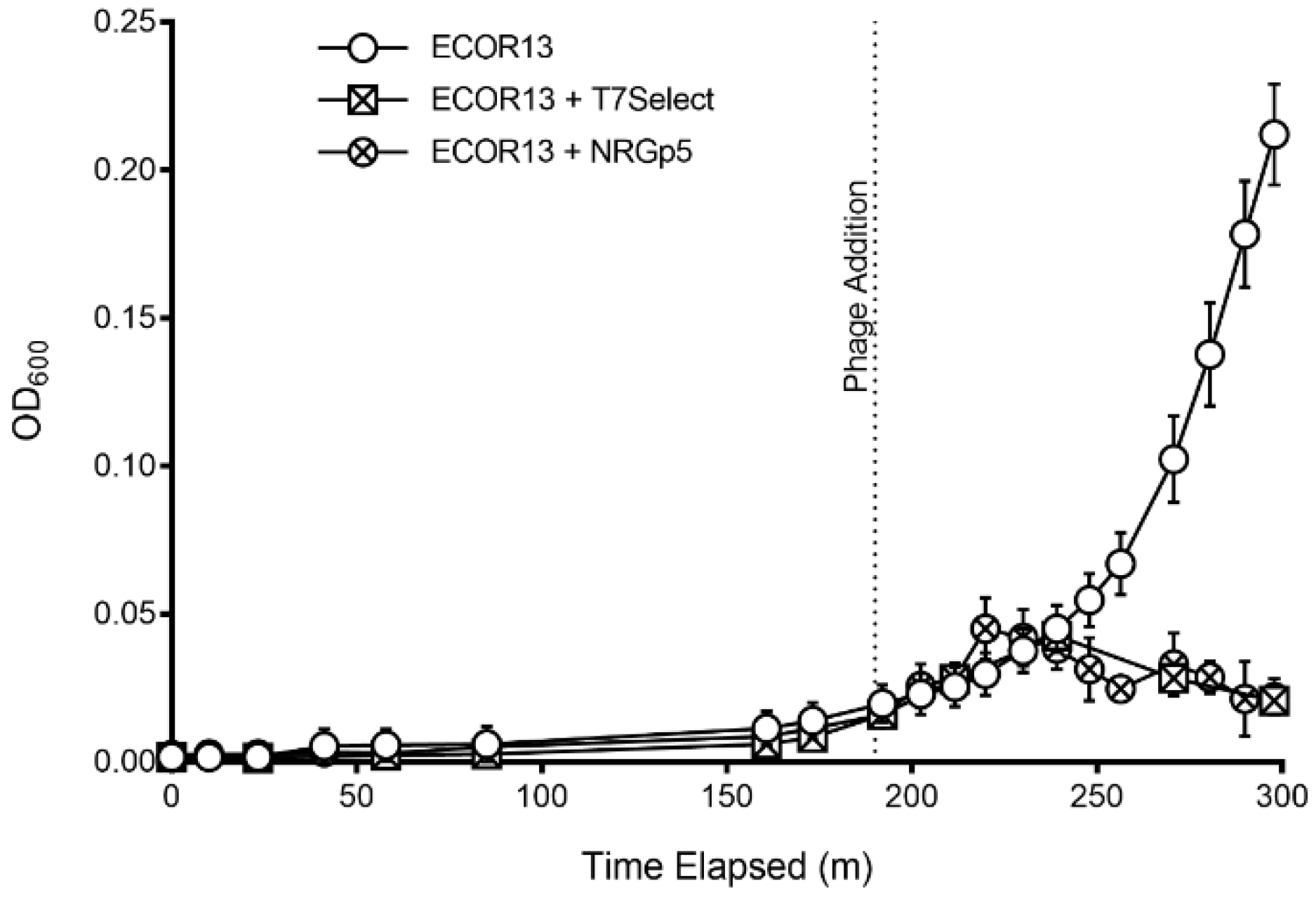
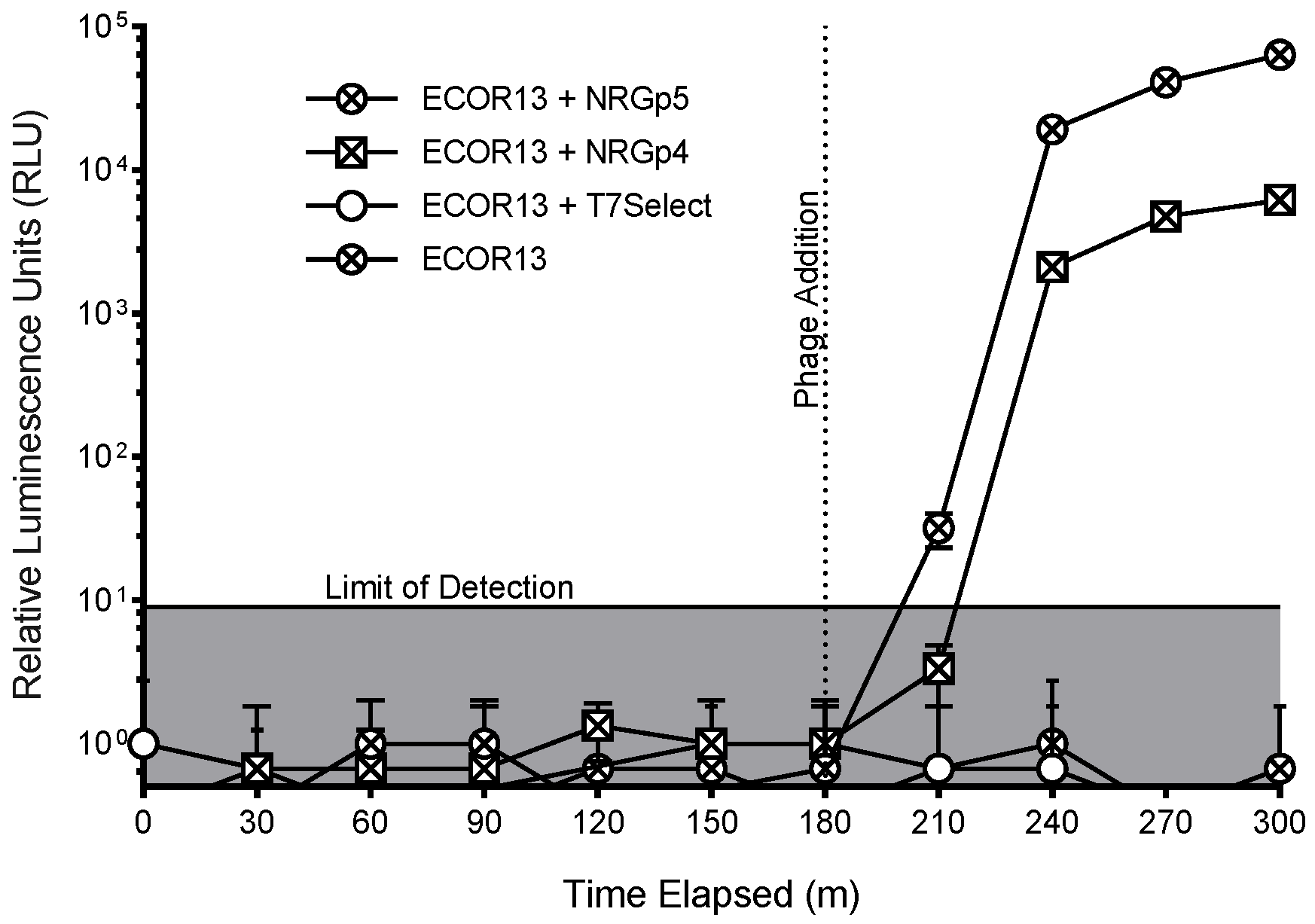
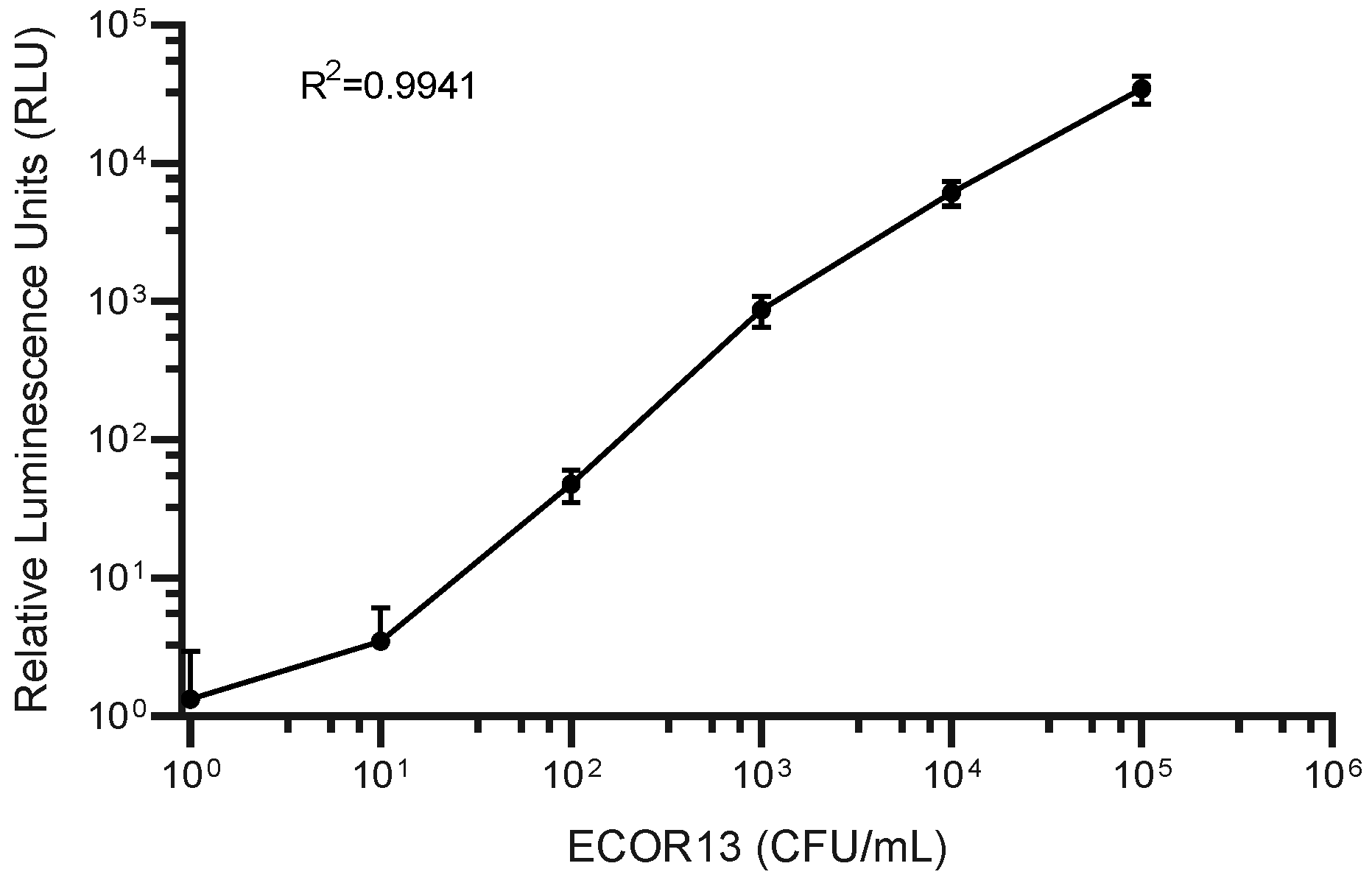
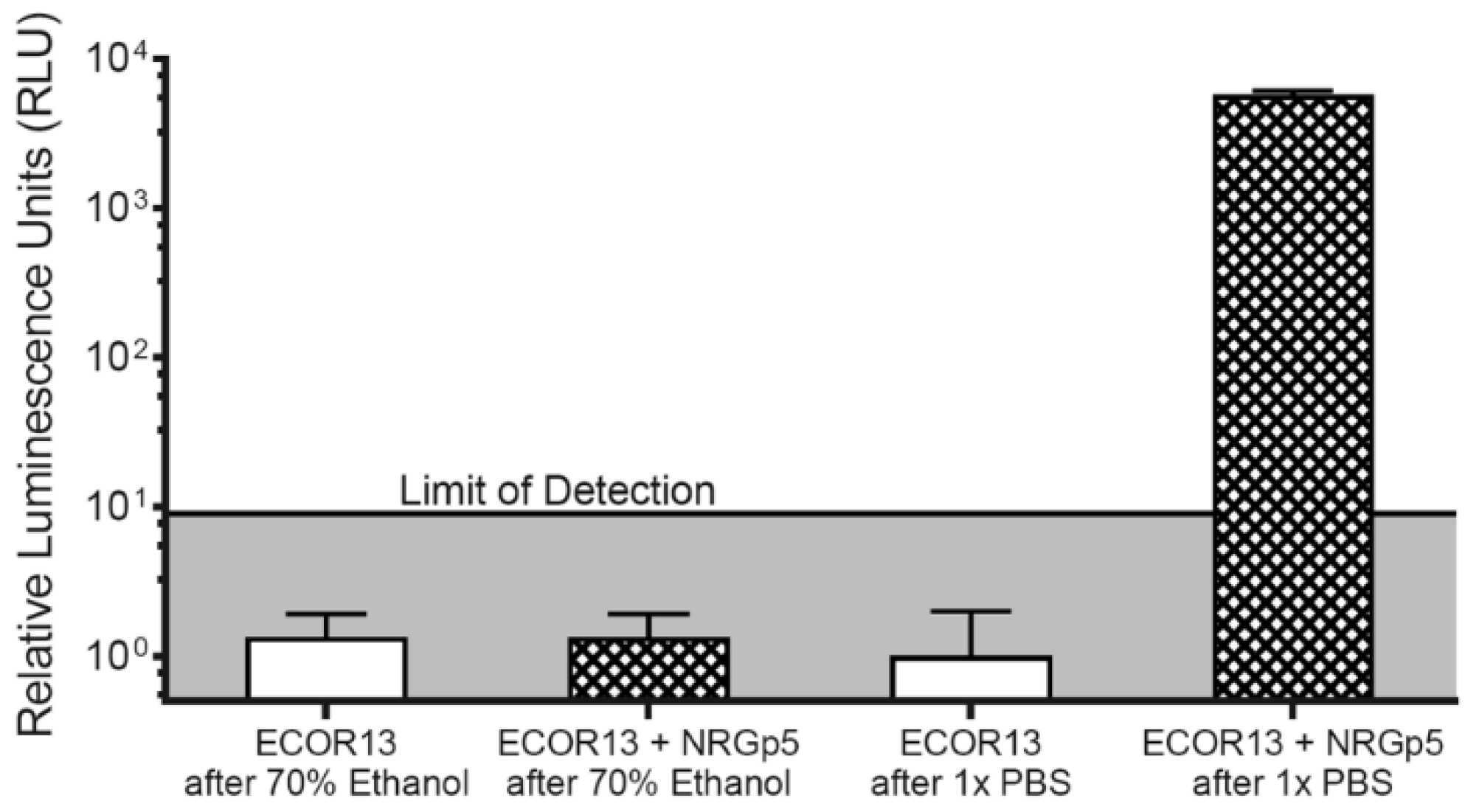
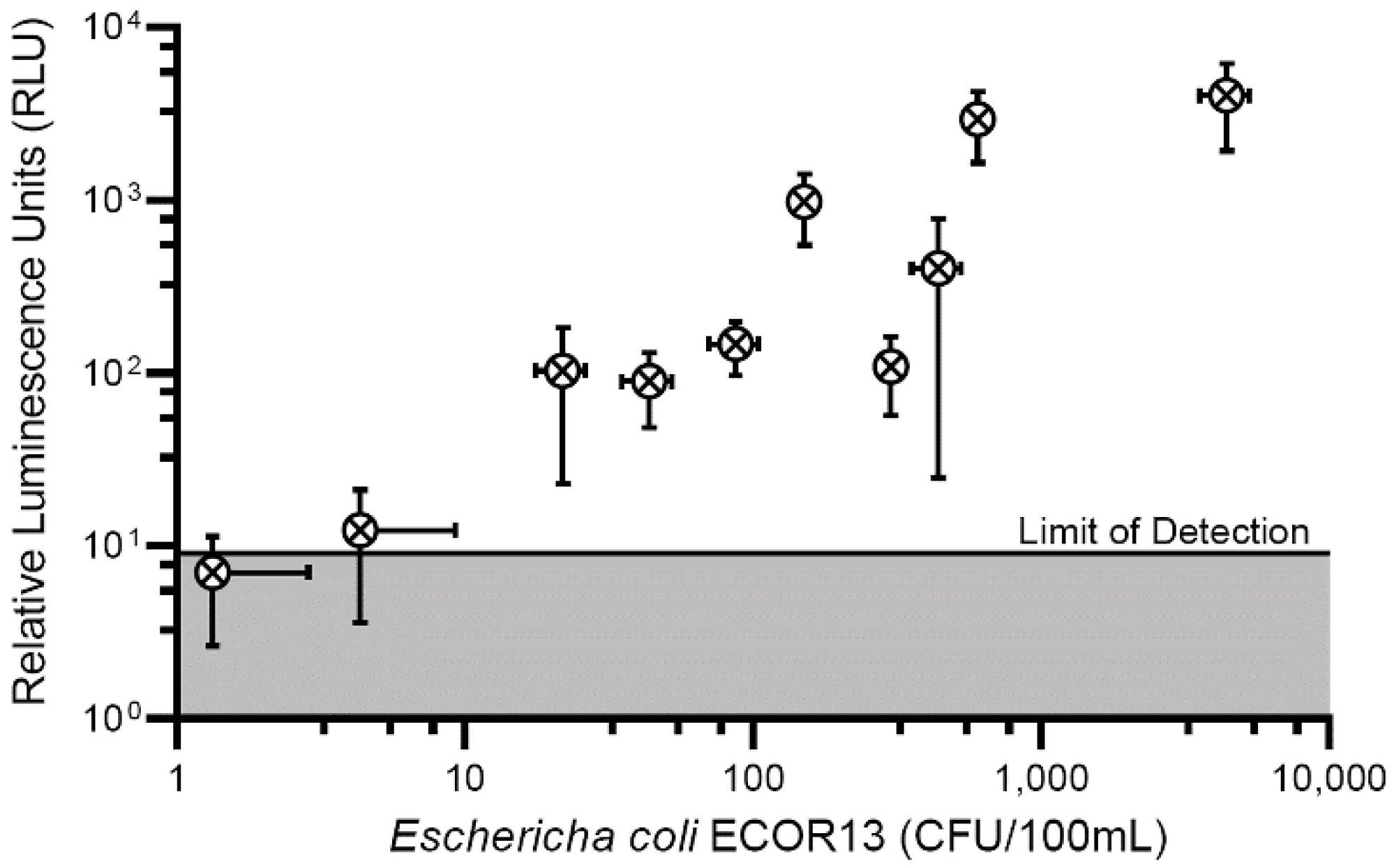
3.3. Phage-Based Syringe Filter Detection Assay
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Assembly, U.N.G. The human right to water and sanitation. UN Resolut. 2010, 64, 292. [Google Scholar]
- Bain, R.; Cronk, R.; Wright, J.; Yang, H.; Slaymaker, T.; Bartram, J. Fecal contamination of drinking-water in low-and middle-income countries: A systematic review and meta-analysis. PLoS Med. 2014, 11, e1001644. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.D.J.; Khush, R.; Peletz, R.; Kumpel, E. Efficacy of microbial sampling recommendations and practices in sub-Saharan Africa. Water Res. 2018, 134, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.; Fujioka, R.; Glymph, T.; McGee, C.; Schaub, S.; Sobsey, M.; Toranzos, G. Pathogen indicators, and indicators of fecal contamination. In Report of the Experts Scientific Workshop on Critical Research Needs for the Development of New or Revised Recreational Water Quality Criteria; EPA: Wasington, DC, USA, 2007; pp. 35–56. [Google Scholar]
- De Vrese, M.; Schrezenmeir, J. Probiotics, Prebiotics, and Synbiotics. In Food Biotechnology. Advances in Biochemical Engineering/Biotechnology; Stahl, U., Donalies, U.E., Nevoigt, E., Eds.; Springer: Berlin, Heidelberg, 2008; Volume 111. [Google Scholar]
- World Health Organization. WHO estimates of the global burden of foodborne diseases: Foodborne disease burden epidemiology reference group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Hodge, J.; Chang, H.H.; Boisson, S.; Collin, S.M.; Peletz, R.; Clasen, T. Assessing the association between thermotolerant coliforms in drinking water and diarrhea: An analysis of individual-level data from multiple studies. Env. Health Perspect. 2016, 124, 1560–1567. [Google Scholar] [CrossRef] [PubMed]
- Stoll, B.J.; Hansen, N.I.; Sanchez, P.J.; Faix, R.G.; Poindexter, B.B.; van Meurs, K.P.; Bizzarro, M.J.; Goldberg, R.N.; Frantz, I.D.; Hale, E.C.; et al. Early onset neonatal sepsis: The burden of group B streptococcal and E. coli disease continus. Pediatrics 2011, 127, 817–826. [Google Scholar] [CrossRef]
- Caugant, D.; Levin, B.; Lidin-Janson, G.; Whittam, T.; Eden, C.S.; Selander, R. Host Parasite Relationships in Gram-Negative Infections; Karger Publishers: Bazel, Switzerland, 1983; Volume 33, pp. 203–227. [Google Scholar]
- Figueras, M.J.; Borrego, J.J. New perspectives in monitoring drinking water microbial quality. Int. J. Environ. Res. Public Health 2010, 7, 4179–4202. [Google Scholar] [CrossRef]
- Edberg, S.C.; Rice, E.W.; Karlin, R.J.; Allen, M.J. Escherichia coli: The best biological drinking water indicator for public health protection. J. Appl. Microbiol. 2000, 88, 106S–116S. [Google Scholar] [CrossRef]
- Costan-Longares, A.; Montemayor, M.; Payan, A.; Mendez, J.; Jofre, J.; Mujeriego, R.; Lucena, F. Water reclamation technologies for safe mangeed aquifer rechangrge. Water Res. 2008, 42, 4439–4448. [Google Scholar]
- Bain, R.; Bartram, J.; Elliott, M.; Matthews, R.; McMahan, L.; Tung, R.; Chuang, P.; Gundry, S. A summary catalogue of microbial drinking water tests for low and medium resource settings. Int. J. Environ. Res. Public Health 2012, 9, 1609–1625. [Google Scholar] [CrossRef]
- Truchado, P.; Hernandez, N.; Gil, M.I.; Ivanek, R.; Allende, A. Microbial water quality: Monitoring and modeling. Water Res. 2018, 128, 226–233. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency (USEPA). Method 1604: Total Coliforms and Escherichia coli in Water by Membrane Filtration Using a Simultaneous Detection Technique (MI Medium). 2002. Available online: https://nepis.epa.gov/Exe/ZyNET.exe/P1002D57.TXT?ZyActionD=ZyDocument&Client=EPA&Index=2000+Thru+2005&Docs=&Query=&Time=&EndTime=&SearchMethod=1&TocRestrict=n&Toc=&TocEntry=&QField=&QFieldYear=&QFieldMonth=&QFieldDay=&IntQFieldOp=0&ExtQFieldOp=0&XmlQuery=&File=D%3A%5Czyfiles%5CIndex%20Data%5C00thru05%5CTxt%5C00000017%5CP1002D57.txt&User=ANONYMOUS&Password=anonymous&SortMethod=h%7C-&MaximumDocuments=1&FuzzyDegree=0&ImageQuality=r75g8/r75g8/x150y150g16/i425&Display=hpfr&DefSeekPage=x&SearchBack=ZyActionL&Back=ZyActionS&BackDesc=Results%20page&MaximumPages=1&ZyEntry=1&SeekPage=x&ZyPURL (accessed on 10 February 2020).
- US Environmental Protection Agency. Method 1603: Escherichia coli (E. coli) in Water by Membrane Filtration Using Modified Membrane-Thermotolerant Escherichia coli Agar (Modified mTEC). Available online: https://www.epa.gov/sites/production/files/2015-08/documents/method_1603_2009.pdf (accessed on 10 February 2020).
- Association, A.W.W. Water Chlorination/Chloramination Practices and Principles; American Water Works Association: Denver, CO, USA, 2006. [Google Scholar]
- Cho, I.H.; Ku, S. Current technical approaches for the early detection of foodbore pathogens: Challenges and opportunitieds. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Richter, L.; Janczuk-Richter, M.; Niedziolka-Jonsson, J.; Paczesny, J.; Holyst, R. Recent advances in bacteriophage-based methods for bacteria detection. Drug Discov. Today 2018, 23, 448–455. [Google Scholar] [CrossRef] [PubMed]
- DeLong, E.F.; Pace, N.R. Environmental diversity of bacteria and archaea. Syst. Biol. 2001, 50, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Kim, Y.T.; Ryu, S.; Lee, J.H. Biocontral and detection of food-borne pathogens using bacteriophages and endolysins. Front. Microbiol. 2016, 7, 474. [Google Scholar] [CrossRef] [PubMed]
- McElroy, W.D. The Energy Source for Bioluminescence in an Isolated System. Proc. Natl. Acad. Sci. USA 1947, 33, 342. [Google Scholar] [CrossRef]
- Ding, Y.; Hua, X.; Chen, H.; Liu, F.; Gonzalez-Sapien, G.; Wang, M. Rapid antibody-based technologies in food analysis. Anal. Chem. 2018, 90, 2230–2237. [Google Scholar] [CrossRef]
- Zhang, D.; Coronel-Aguilera, C.P.; Romero, P.L.; Perry, L.; Minocha, U.; Rosenfield, C.; Gehring, A.G.; Paoli, G.C.; Bhunia, A.K.; Applegate, B. Methods for detection of viable foodborne pathogens: Current state-of-art and future prospects. Sci. Rep. 2016, 6, 33235. [Google Scholar] [CrossRef]
- Masser, A.E.; Kandasamy, G.; Kaimal, J.M.; Andreasson, C. Luciferase nanoluc as a reporter for gene expression and protein in saccharomyces cerevisiae. Yeast 2016, 33, 191–200. [Google Scholar] [CrossRef]
- Cevenini, L.; Calabretta, M.M.; Lopreside, A.; Tarantino, G.; Tassoni, A.; Ferri, M.; Roda, A.; Michelini, E. Exploiting nanoluc luciferase for smartphone-based bioluminescence cell biosensors for anti-inflammatory activity and toxicity. Anal. Bioanal. Chem. 2016, 408, 8859–8868. [Google Scholar] [CrossRef]
- Oh-Hashi, K.; Hirata, Y.; Kiuchi, K. SOD1 dimerization monitoring using a novel split nanoluc, nanobit. Cell Biochem. Funct. 2016, 34, 497–504. [Google Scholar] [CrossRef]
- England, C.G.; Ehlerding, E.B.; Cai, W. Nanoluc: A small luciferase is brightening up the field of bioluminescence. Bioconjug Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. Nanoluc complementation reporter optimized for accurate measurement of protein interactions in cells. ACS Chem. Biol. 2016, 11, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.-J.; Song, G.; Zhang, Z.; Guo, Z.-Y. Efficient overexpression of human interleukin-6 in Escherichia coli using nanoluciferase as a fusion partner. Process Biochem. 2015, 50, 1618–1622. [Google Scholar] [CrossRef]
- Sun, C.; Gardner, C.L.; Watson, A.M.; Ryman, K.D.; Klimstra, W.B. Screening for novel small-molecule inhibitors targeting the assembly of influenza virus polymerase complex by a bimolecular luminescence complementation-based reporter system. J. Virol. 2014, 88, 2035–2046. [Google Scholar] [CrossRef]
- Tran, V.; Moser, L.A.; Poole, D.S.; Mehle, A. Highly sensitive real-time in vivo of an influence reporter virus reveals dynamics of replication and spread. J. Virol. 2013, 87, 13321–13329. [Google Scholar] [CrossRef]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilization a novel imidazopyrazinonc substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.; Kim, S.; Ryu, S. Sensitive detection of viable Escherichia coli O157:H7 from foods using a luciferase-reporter phage phiV10lux. Int. J. Food Microbiol. 2017, 254, 11–17. [Google Scholar] [CrossRef]
- Oosterik, L.H.; Tuntufye, H.N.; Tsonos, J.; Luyten, T.; Noppen, S.; Liekens, S.; Lavigne, R.; Butaye, P.; Goddeeris, B.M. Bioluminescent avian pathogenic Escherichia coil for monitoring colibacillosis in experimentally infected chickens. Vet. J. 2016, 216, 87–92. [Google Scholar] [CrossRef]
- Loessner, M.J.; Rees, C.E.; Stewart, G.S.; Scherer, S. The terminally redundant, nonpermuted genome of listeria bacteriophage A511: A model for the SPO1-like myoviruses of gram-positive bacteria. Appl. Environ. Microbiol. 1996, 62, 1133–1140. [Google Scholar] [CrossRef]
- Kim, S.; Kim, M.; Ryu, S. Noncanonical DNA-binding mode of repressor and its disassembly by antirepressor. Anal. Chem. 2014, 86, 5858–5864. [Google Scholar] [CrossRef]
- Kumar, V.; Loganathan, P.; Sivaramakrishnan, G.; Kriakov, J.; Dusthakeer, A.; Subramanyam, B.; Chan, J.; Jacobs, W.R., Jr.; Rama, N.P. Characterization of temperate phage che12 and construction of a new tool for diagnosis of tuberculosis. Tuberculosis 2008, 88, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Jin, Y.; Lin, H.; Wang, J.; Khan, M.N. Application of the VPp1 bacteriophage combined with a coupled enzyme system in the rapid detection of vibro parahaemolyticus. J. Microbiol. Methods 2014, 98, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Sharp, N.J.; Vandamm, J.P.; Molineux, I.J.; Schofield, D.A. Rapid detection of Bacillus anthracis in complex food matrices using phage-mediated bioluminescence. J. Food Prot. 2015, 78, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Vandamm, J.; Rajanna, C.; Sharp, N.; Molineux, I.; Schofield, D. Rapid detection and simulation antibiotic susceptibility analysis of yersinsia pestis directly from clinical specimens using reporter pahge. J. Clin. Microbiol. 2014, 314–316. [Google Scholar]
- Schofield, D.; Wray, D.; Molineux, I. The Europen journal of clinical microbiology and infectious diseases: Quality and quantity in 2013. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 395–403. [Google Scholar] [CrossRef]
- Schofield, D.A.; Bull, C.T.; Rubio, I.; Wechter, W.P.; Westwater, C.; Molineux, I.J. Development of an engineed bioluminescent reporter phage for detection of bacterial blight of crucifers. Appl. Environ. Microbiol. 2012, 78, 3592–3598. [Google Scholar] [CrossRef]
- Nguyen, C.; Makkar, R.; Sharp, N.J.; Page, M.A.; Molineux, I.J.; Schofield, D.A. Detection of Bacillus anthracis spores from environmental water using bioluminescent reporter phage. J. Appl. Microbiol. 2017, 123, 1184–1193. [Google Scholar] [CrossRef]
- Schofield, D.; Bull, C.T.; Rubio, I.; Wechter, W.P.; Westwater, C.; Molineux, I.J. “Light-tagged” bacteriophage as a diagnostic tool for the detection of phytopathogents. Bioengineered 2013, 4, 50–54. [Google Scholar] [CrossRef]
- Kutter, E. Phage production and maintenance of stocks, including expected stock lifetimes. Methods Mol.Biol. 2009, 501, 141–149. [Google Scholar]
- Wu, J.; Long, S.C.; Das, D.; Dorner, S.M. Are microbial indicators and pathogens correlated? A statistical analysis of 40 yers of research. J. Water Health 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Hinkley, T.C.; Singh, S.; Garing, S.; le Ny, A.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. A phage-based assay for the rapid, quantitative, and single CFU visualization of E.coli in drinking water. Sci. Rep. 2018, 8, 14630. [Google Scholar] [CrossRef] [PubMed]
- Hinkley, T.C.; Garing, S.; Singh, S.; le Ny, A.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. Reporter bacteriophage T7 NLC utilizes a novel NanLuc:CBM fusion for the ultrasensitive detection of Escherichia cooli in water. Analyst 2018, 143, 4074–4082. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Hinkley, T.; Nugen, S.R.; Talbert, J.N. Fusion of carbohyfrate binding module to mutant alkaline phosphatase for immobilization on cellulose. Biocatal. Agric. Biotechnol. 2018, 13, 265–271. [Google Scholar] [CrossRef]
- E. I. L. L. C. Office of The Federal Register. In Title 40 Protection of Environment Parts 136 to 149 (Revised as of July 1, 2013): 40-CFR-Vol-24; U.S. Government Printing Office: Wasington, DC, USA, 2014. [Google Scholar]
- Clokie, M.R.J.; Kropinski, A.M. Bacteriophages: Methods and Protocols, Isolation, Characterization, and Interactions; Humana Press: Totowa, NJ, USA, 2009; Volume 1. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 1989. [Google Scholar]
- Dunn, J.J.; Studier, F.W. Complete nucleotide sequence of bacteriophage T7 DNA and the location of T7 genetic elements. J. Mol. Biol. 1983, 166, 477–535. [Google Scholar] [CrossRef]
- Jackson, A.A.; Hinkley, T.C.; Talbert, J.N.; Nugen, S.R.; Sela, D.A. Genetic optimization of a bacteriophage-delivered alkaline phosphatase reporter to detect Escherichia coli. Analyst 2016. [Google Scholar] [CrossRef]
- Alcaine, S.D.; Tilton, L.; Serrano, M.A.; Wang, M.; Vachet, R.W.; Nugen, S.R. Phage-protease-peptide: A novel trifecta enabling multiplex detection of viable bacterial pathogens. Appl. Microbiol. Biotechnol. 2015, 99, 8177–8185. [Google Scholar] [CrossRef]
- Food and Drug Administration. Final Report and Executive Summaries from the AOAC International Presidential Task Force on Best Practices in Microbiological Methodology; AOAC International: Rockville, MD, USA, 2011. [Google Scholar]
- Food and Drug Administration. Guidelines for the Validation of Analytical Methods for the Detection of Microbial Pathogens in Foods and Feeds; US Food and Drug Administration: Washington, DC, USA, 2015.






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinkley, T.C.; Garing, S.; Jain, P.; Williford, J.; Le Ny, A.-L.M.; Nichols, K.P.; Peters, J.E.; Talbert, J.N.; Nugen, S.R. A Syringe-Based Biosensor to Rapidly Detect Low Levels of Escherichia Coli (ECOR13) in Drinking Water Using Engineered Bacteriophages. Sensors 2020, 20, 1953. https://doi.org/10.3390/s20071953
Hinkley TC, Garing S, Jain P, Williford J, Le Ny A-LM, Nichols KP, Peters JE, Talbert JN, Nugen SR. A Syringe-Based Biosensor to Rapidly Detect Low Levels of Escherichia Coli (ECOR13) in Drinking Water Using Engineered Bacteriophages. Sensors. 2020; 20(7):1953. https://doi.org/10.3390/s20071953
Chicago/Turabian StyleHinkley, Troy C., Spencer Garing, Paras Jain, John Williford, Anne-Laure M. Le Ny, Kevin P. Nichols, Joseph E. Peters, Joey N. Talbert, and Sam R. Nugen. 2020. "A Syringe-Based Biosensor to Rapidly Detect Low Levels of Escherichia Coli (ECOR13) in Drinking Water Using Engineered Bacteriophages" Sensors 20, no. 7: 1953. https://doi.org/10.3390/s20071953
APA StyleHinkley, T. C., Garing, S., Jain, P., Williford, J., Le Ny, A.-L. M., Nichols, K. P., Peters, J. E., Talbert, J. N., & Nugen, S. R. (2020). A Syringe-Based Biosensor to Rapidly Detect Low Levels of Escherichia Coli (ECOR13) in Drinking Water Using Engineered Bacteriophages. Sensors, 20(7), 1953. https://doi.org/10.3390/s20071953