
Population Structure of Aetobatus narinari (Myliobatiformes) Caught by the Artisanal Fishery in Northeast Brazil

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
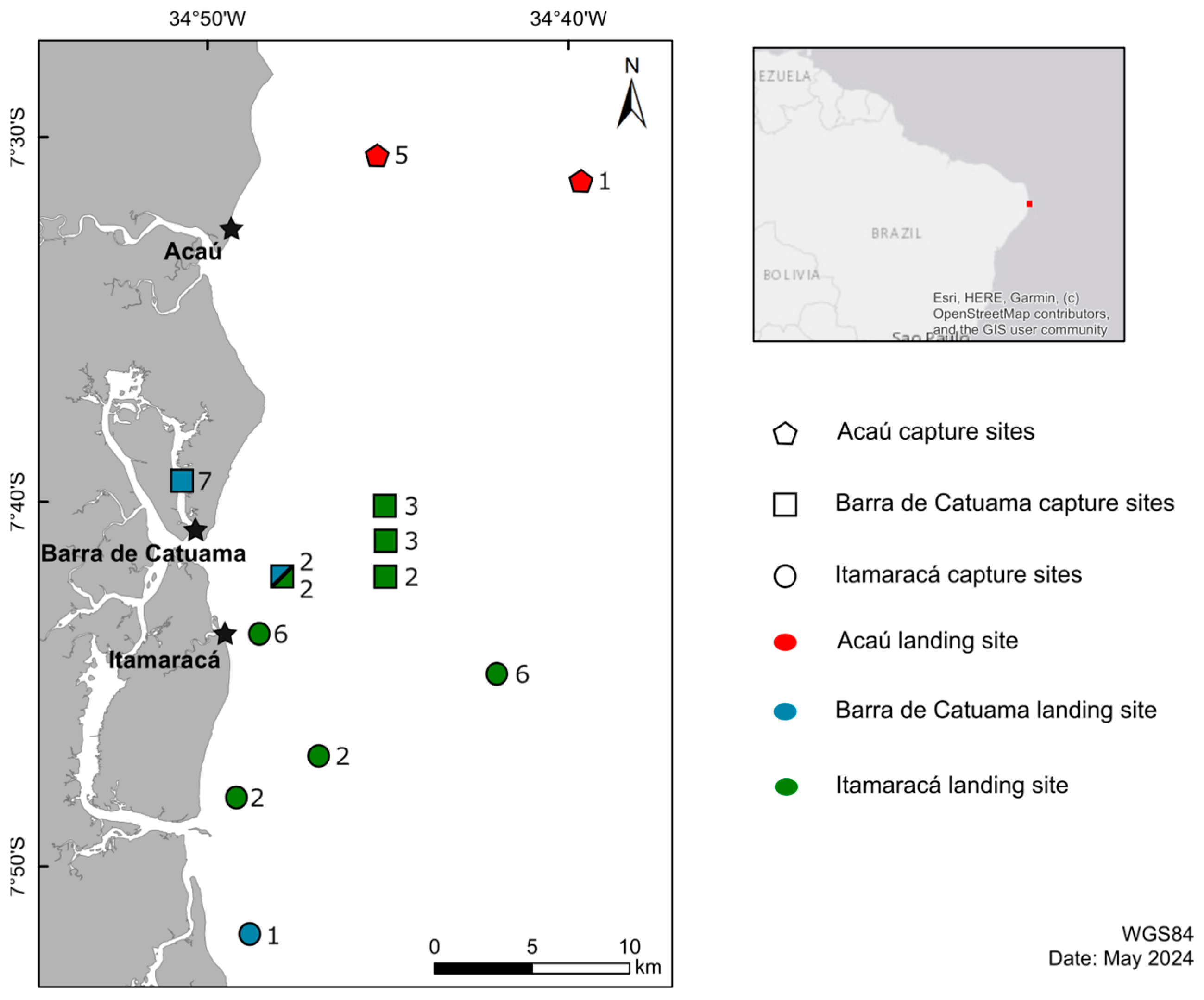
2.1. Study Area
2.2. Sampling
2.3. DNA Extraction, PCR, and Sequencing
2.4. Analysis of Genetic Diversity and Structure
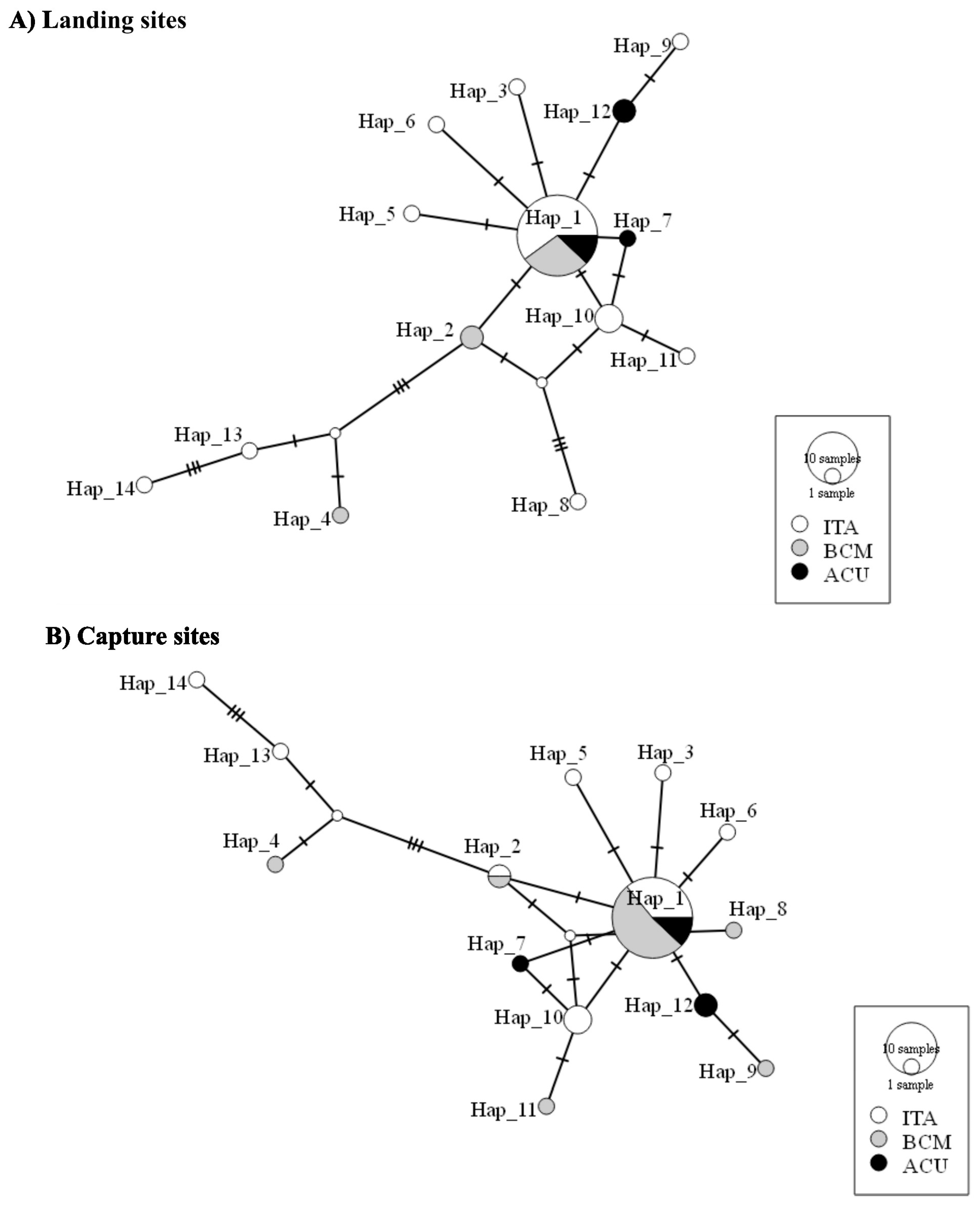
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Euphrasen, B.A. Raja (Narinari). Kongliga Vetensk. Akad. Nya Handl. 1790, 11, 217–219. [Google Scholar]
- Bigelow, H.B.; Schroeder, W.C. Sawfishes, Guitarfishes, Skates and Rays. In Fishes of the Western North Atlantic; Memoir Sears Foundation for Marine Research: New Haven, CT, USA, 1953; pp. 1–514. [Google Scholar]
- White, W.; Last, P.; Naylor, G.; Jensen, K.; Caira, J.N. Clarification of Aetobatus ocellatus (Kuhl, 1823) as a Valid Species, and a Comparison with Aetobatus narinari (Euphrasen, 1790) (Rajiformes: Myliobatidae). Descr. New Sharks Rays Borneo 2010, 32, 141–164. [Google Scholar]
- Sales, J.B.L.; De Oliveira, C.N.; Dos Santos, W.C.R.; Rotundo, M.M.; Ferreira, Y.; Ready, J.; Sampaio, I.; Oliveira, C.; Cruz, V.P.; Lara-Mendoza, R.E.; et al. Phylogeography of Eagle Rays of the Genus Aetobatus: Aetobatus narinari Is Restricted to the Continental Western Atlantic Ocean. Hydrobiologia 2019, 836, 169–183. [Google Scholar] [CrossRef]
- Dulvy, N.; Carlson, J.; Charvet, P.; Bassos-Hull, K.; Blanco-Parra, M.-D.-P.; Chartrain, E.; Derrick, D.; Dia, M.; Diop, M.; Doherty, P.; et al. Aetobatus narinari-Whitespotted Eagle Ray. The IUCN Red List of Threatened Species 2021. 2021. Available online: https://www.iucnredlist.org/species/42564343/2924463 (accessed on 12 December 2023).
- ICMBIO. Avaliação Do Risco de Extinção Dos Elasmobrânquios e Quimeras No Brasil: 2010–2012; ICMBIO/MMA/CEPSUL: Itajaí, Brazil, 2016. [Google Scholar]
- Tagliafico, A.; Rago, N.; Rangel, M.; Mendoza, J. Exploitation and Reproduction of the Spotted Eagle Ray (Aetobatus narinari) in the Los Frailes Archipelago, Venezuela. Fish. Bull. 2012, 110, 307–316. [Google Scholar]
- Janse, M.; Kappe, A.; van Kuijk, B.L.M. Paternity Testing Using the Poisonous Sting in Captive White-Spotted Eagle Rays Aetobatus narinari: A Non-Invasive Tool for Captive Sustainability Programmes. J. Fish Biol. 2013, 82, 1082–1085. [Google Scholar] [CrossRef]
- Bassos-Hull, K.; Wilkinson, K.; Hull, P.; Dougherty, D.; Omori, K.; Ailloud, L.; Morris, J.; Hueter, R. Life History and Seasonal Occurrence of the Spotted Eagle Ray, Aetobatus narinari, in the Eastern Gulf of Mexico. Environ. Biol. Fishes 2014, 97, 1039–1056. [Google Scholar] [CrossRef]
- Harmon, T.S.; Kamerman, T.Y.; Corwin, A.L.; Sellas, A.B. Consecutive Parthenogenetic Births in a Spotted Eagle Ray Aetobatus narinari. J. Fish Biol. 2016, 88, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Araújo, P.; Oddone, M.C.; Evêncio-Neto, J.; LESSA, R. Reproductive Biology of the Whitespotted Eagle Ray Aetobatus narinari (Myliobatiformes) Captured in the Coast of Paraíba and Pernambuco, Brazil. J. Fish Biol. 2022, 100, 944–957. [Google Scholar] [CrossRef]
- Sellas, A.; Bassos-Hull, K.; Pérez-Jiménez, J.; Valdes, J.; Bernal, M.; Hueter, R. Population Structure and Seasonal Migration of the Spotted Eagle Ray, Aetobatus narinari. J. Hered. 2015, 106, 266–275. [Google Scholar] [CrossRef]
- Sellas, A.; Bassos-Hull, K.; Hueter, R.; Feldheim, K. Isolation and Characterization of Polymorphic Microsatellite Markers from the Spotted Eagle Ray (Aetobatus narinari). Conserv. Genet. Resour. 2011, 3, 609–611. [Google Scholar] [CrossRef]
- Newby, J.; Darden, T.; Shedlock, A. Population Genetic Structure of Spotted Eagle Rays, Aetobatus narinari, off Sarasota, Florida and the Southeastern United States. Copeia 2014, 2014, 503–512. [Google Scholar] [CrossRef]
- Ajemian, M.J.; Powers, S.P.; Murdoch, T.J.T. Estimating the Potential Impacts of Large Mesopredators on Benthic Resources: Integrative Assessment of Spotted Eagle Ray Foraging Ecology in Bermuda. PLoS ONE 2012, 7, e40227. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Flores, F.; Pérez-Jiménez, J.C.; Méndez-Loeza, I.; Bassos-Hull, K.; Ajemian, M.J. Comparison between the Feeding Habits of Spotted Eagle Ray (Aetobatus narinari) and Their Potential Prey in the Southern Gulf of Mexico. J. Mar. Biol. Assoc. United Kingd. 2019, 99, 661–672. [Google Scholar] [CrossRef]
- Cahill, B.V.; Eckert, R.J.; Bassos-Hull, K.; Ostendorf, T.J.; Voss, J.D.; DeGroot, B.C.; Ajemian, M.J. Diet and Feeding Ecology of the Whitespotted Eagle Ray (Aetobatus narinari) from Florida Coastal Waters Revealed via DNA Barcoding. Fishes 2023, 8, 388. [Google Scholar] [CrossRef]
- Boggio-Pasqua, A.; Bassos-Hull, K.; Aeberhard, W.H.; Hoopes, L.A.; Swider, D.A.; Wilkinson, K.A.; Dureuil, M. Whitespotted Eagle Ray (Aetobatus narinari) Age and Growth in Wild (in Situ) versus Aquarium-Housed (Ex Situ) Individuals: Implications for Conservation and Management. Front. Mar. Sci. 2022, 9, 960822. [Google Scholar] [CrossRef]
- Cuevas-Zimbrón, E.; Pérez-Jiménez, J.C.; Méndez-Loeza, I. Spatial and Seasonal Variation in a Target Fishery for Spotted Eagle Ray Aetobatus narinari in the Southern Gulf of Mexico. Fish. Sci. 2011, 77, 723–730. [Google Scholar] [CrossRef]
- Lessa, R.P.; Santana, F.M.; Rincón, G.; Gadig, O.B.F.; El-Deir, A.C.A. Biodiversidade de Elasmobrânquios Do Brasil; Ministério do Meio Ambiente (MMA), Projeto de Conservação e Utilização Sustentável da Diversidade Biológica Brasileira (PROBIO): Recife, Brazil, 1999. [Google Scholar]
- Dubick, J.D. Age and Growth of the Spotted Eagle Ray, Aetobatus narinari (Euphrasen, 1790), from Southwest Puerto Rico with Notes on Its Biology and Life History. Master’s Thesis, Universidade de Porto Rico, Mayaguez, Puerto Rico, 2000. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A.; McInness, K.H. A Primer of Conservation Genetics; Reprinted; Cambridge University Press: Cambridge, UK, 2005; ISBN 978-0-521-83110-9. [Google Scholar]
- Heithaus, M.R.; Frid, A.; Wirsing, A.J.; Worm, B. Predicting Ecological Consequences of Marine Top Predator Declines. Trends Ecol. Evol. 2008, 23, 202–210. [Google Scholar] [CrossRef]
- Clarke, C.R.; Karl, S.A.; Horn, R.L.; Bernard, A.M.; Lea, J.S.; Hazin, F.H.; Prodöhl, P.A.; Shivji, M.S. Global Mitochondrial DNA Phylogeography and Population Structure of the Silky Shark, Carcharhinus falciformis. Mar. Biol. 2015, 162, 945–955. [Google Scholar] [CrossRef]
- Schluessel, V.; Bennett, M.B.; Collin, S.P. Diet and Reproduction in the White-Spotted Eagle Ray Aetobatus narinari from Queensland, Australia and the Penghu Islands, Taiwan. Mar. Freshw. Res. 2010, 61, 1278–1289. [Google Scholar] [CrossRef]
- Yokota, L.; Lessa, R.P. A Nursery Area for Sharks and Rays in Northeastern Brazil. Environ. Biol. Fishes 2006, 75, 349–360. [Google Scholar] [CrossRef]
- Meyer, C.P. Molecular Systematics of Cowries (Gastropoda: Cypraeidae) and Diversification Patterns in the Tropics. Biol. J. Linn. Soc. 2003, 79, 401–459. [Google Scholar] [CrossRef]
- Richards, V.P.; Henning, M.; Witzell, W.; Shivji, M.S. Species Delineation and Evolutionary History of the Globally Distributed Spotted Eagle Ray (Aetobatus narinari). J. Hered. 2009, 100, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Arèvalo, E.; Davis, S.K.; Sites, J.W., Jr. Mitochondrial DNA Sequence Divergence and Phylogenetic Relationships among Eight Chromosome Races of the Sceloporus Grammicus Complex (Phrynosomatidae) in Central Mexico. Syst. Biol. 1994, 43, 387–418. [Google Scholar] [CrossRef]
- Inoue, J.G.; Miya, M.; Tsukamoto, K.; Nishida, M. A Mitogenomic Perspective on the Basal Teleostean Phylogeny: Resolving Higher-Level Relationships with Longer DNA Sequences. Mol. Phylogenet. Evol. 2001, 20, 275–285. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walke, P.; Posada, D.; Crandall, K. TCS: Estimating Gene Genealogies. In Proceedings of the 16th International Parallel and Distributed Processing Symposium, Fort Lauderdale, FL, USA, 15–19 April 2002; p. 7. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Reiss, H.; Hoarau, G.; Dickey-Collas, M.; Wolff, W.J. Genetic Population Structure of Marine Fish: Mismatch between Biological and Fisheries Management Units. Fish Fish. 2009, 10, 361–395. [Google Scholar] [CrossRef]
- DeGroot, B.C.; Bassos-Hull, K.; Wilkinson, K.A.; Lowerre-Barbieri, S.; Poulakis, G.R.; Ajemian, M.J. Variable Migration Patterns of Whitespotted Eagle Rays Aetobatus narinari along Florida’s Coastlines. Mar. Biol. 2021, 168, 18. [Google Scholar] [CrossRef]
- Flowers, K.I.; Henderson, A.C.; Lupton, J.L.; Chapman, D.D. Site Affinity of Whitespotted Eagle Rays Aetobatus narinari Assessed Using Photographic Identification. J. Fish Biol. 2017, 91, 1337–1349. [Google Scholar] [CrossRef]
- Ensing, D.; Crozier, W.W.; Boylan, P.; O’Maoiléidigh, N.; McGinnity, P. An Analysis of Genetic Stock Identification on a Small Geographical Scale Using Microsatellite Markers, and Its Application in the Management of a Mixed-Stock Fishery for Atlantic Salmon Salmo salar in Ireland. J. Fish Biol. 2013, 82, 2080–2094. [Google Scholar] [CrossRef] [PubMed]
- Postaire, B.D.; Devloo-Delva, F.; Brunnschweiler, J.M.; Charvet, P.; Chen, X.; Cliff, G.; Daly, R.; Drymon, J.M.; Espinoza, M.; Fernando, D.; et al. Global Genetic Diversity and Historical Demography of the Bull Shark. J. Biogeogr. 2024, 51, 632–648. [Google Scholar] [CrossRef]
- Schluessel, V.; Broderick, D.; Collin, S.P.; Ovenden, J.R. Evidence for Extensive Population Structure in the White-Spotted Eagle Ray within the Indo-Pacific Inferred from Mitochondrial Gene Sequences. J. Zool. 2010, 281, 46–55. [Google Scholar] [CrossRef]
- Cruz, V.P.; Adachi, A.M.C.L.; Oliveira, P.H.; Ribeiro, G.S.; Paim, F.G.; Souza, B.C.; Rodrigues, A.S.F.; Vianna, M.; Delpiani, S.M.; Díaz de Astarloa, J.M.; et al. Genetic Diversity in Two Threatened Species of Guitarfish (Elasmobranchii: Rhinobatidae) from the Brazilian and Argentinian Coasts: An Alert for Conservation. Neotropical Ichthyol. 2021, 19, e210012. [Google Scholar] [CrossRef]
- Mendonça, F.F.; Oliveira, C.; Gadig, O.B.F.; Foresti, F. Phylogeography and Genetic Population Structure of Caribbean Sharpnose Shark Rhizoprionodon porosus. Rev. Fish Biol. Fish. 2011, 21, 799–814. [Google Scholar] [CrossRef]
- Sandoval-Castillo, J.; Rocha-Olivares, A. Deep Mitochondrial Divergence in Baja California Populations of an Aquilopelagic Elasmobranch: The Golden Cownose Ray. J. Hered. 2011, 102, 269–274. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Landing Sites | |||
| Itamaracá | Barra de Catuama | Acaú | |
| Itamaracá | 0.00000 | ||
| Barra de Catuama | 0.01049 | 0.00000 | |
| Acaú | −0.00713 | 0.09579 | 0.00000 |
| Capture Sites | |||
| Itamaracá | Barra de Catuama | Acaú | |
| Itamaracá | 0.00000 | ||
| Barra de Catuama | −0.01053 | 0.00000 | |
| Acaú | 0.01698 | −0.00429 | 0.00000 |
| Landing Sites | ||||
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation |
| Among groups | 1 | 0.885 | −0.00802 Va | −0.93 |
| Among populations within groups | 1 | 1.054 | 0.01396 Vb | 1.63 |
| Within populations | 39 | 33.251 | 0.85260 Vc | 99.31 |
| Total | 41 | 35.190 | 0.85854 | |
| Fixation Index | ΦST: 0.00692 | |||
| Capture Sites | ||||
| Source of Variation | d.f. | Sum of squares | Variance components | Percentage of variation |
| Among groups | 1 | 0.885 | 0.00698 Va | 0.81 |
| Among populations within groups | 1 | 0.750 | −0.00613 Vb | −0.71 |
| Within populations | 39 | 33.556 | 0.86040 Vc | 99.90 |
| Total | 41 | 35.190 | 0.85854 | |
| Fixation Index | ΦST: 0.00099 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo, P.R.V.; Postaire, B.D.; Feldheim, K.A.; Bassos-Hull, K.; Lessa, R.; Ajemian, M.J. Population Structure of Aetobatus narinari (Myliobatiformes) Caught by the Artisanal Fishery in Northeast Brazil. Diversity 2024, 16, 377. https://doi.org/10.3390/d16070377
Araújo PRV, Postaire BD, Feldheim KA, Bassos-Hull K, Lessa R, Ajemian MJ. Population Structure of Aetobatus narinari (Myliobatiformes) Caught by the Artisanal Fishery in Northeast Brazil. Diversity. 2024; 16(7):377. https://doi.org/10.3390/d16070377
Chicago/Turabian StyleAraújo, Priscila R. V., Bautisse D. Postaire, Kevin A. Feldheim, Kim Bassos-Hull, Rosangela Lessa, and Matthew J. Ajemian. 2024. "Population Structure of Aetobatus narinari (Myliobatiformes) Caught by the Artisanal Fishery in Northeast Brazil" Diversity 16, no. 7: 377. https://doi.org/10.3390/d16070377
APA StyleAraújo, P. R. V., Postaire, B. D., Feldheim, K. A., Bassos-Hull, K., Lessa, R., & Ajemian, M. J. (2024). Population Structure of Aetobatus narinari (Myliobatiformes) Caught by the Artisanal Fishery in Northeast Brazil. Diversity, 16(7), 377. https://doi.org/10.3390/d16070377