

Diverse Communities of Endemic and Cosmopolitan Lineages at Local Sites in the Lecanora polytropa Aggregate (Ascomycota)

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction, Sequencing and Alignment
2.3. Candidate Species Delimitation Using the Standard Fungal DNA Barcode
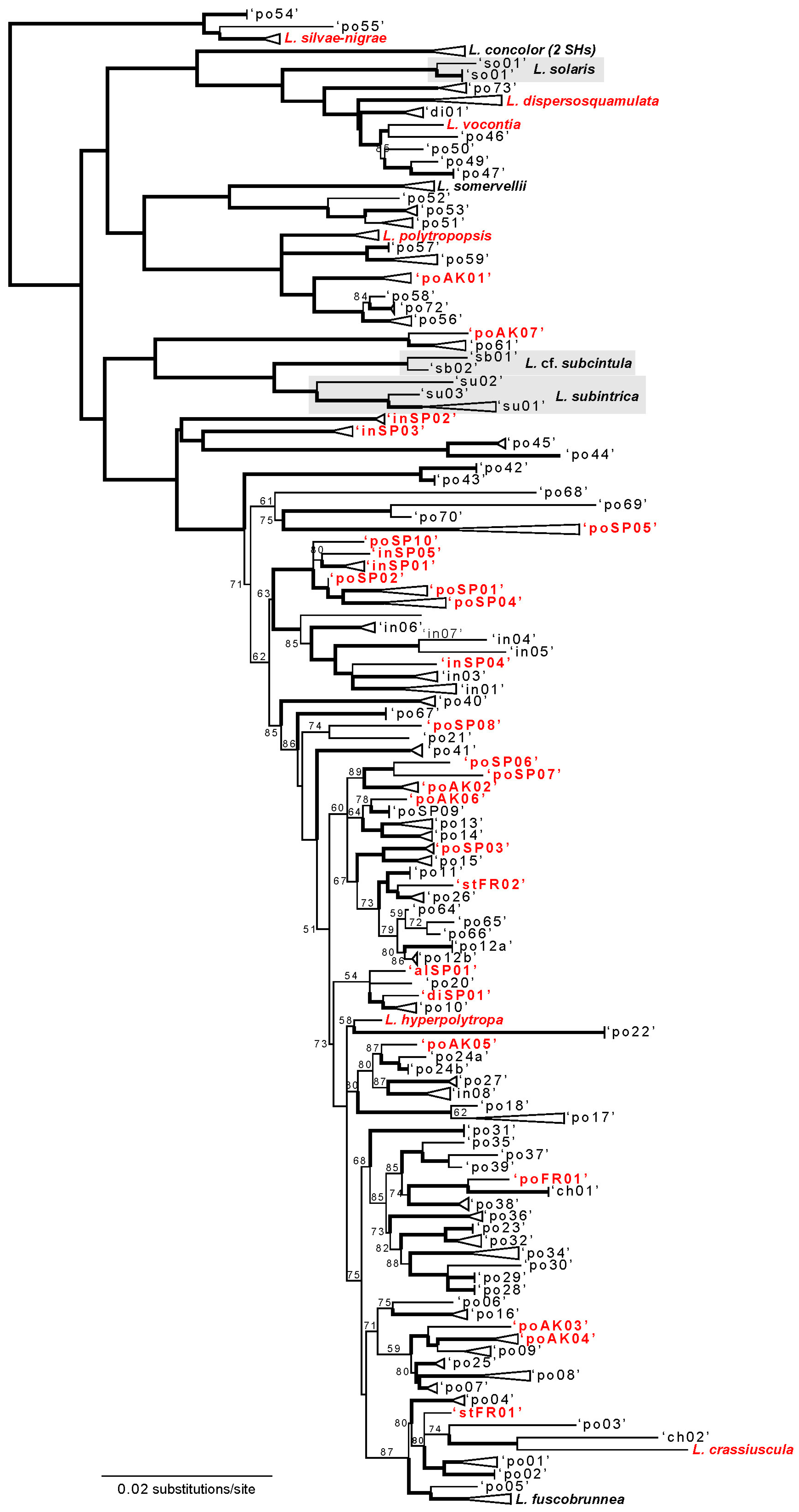
3. Results
3.1. ASAP Candidate Species Delimitation
3.1.1. ASAP Candidate Species at Two Sites in the Pacific Coast Ranges
3.1.2. ASAP Candidate Species at Three Sites in Montane Habitats in Spain
3.1.3. ASAP Candidate Species in Species of Interest from France
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, J.L.; Lendemer, J.C. A call to reconceptualize lichen symbioses. Trends Ecol. Evol. 2022, 37, 582–589. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Pérez-Ortega, S. Past, present, and future research in bipolar lichen-forming fungi and their photobionts. Am. J. Bot. 2017, 104, 1660–1674. [Google Scholar] [CrossRef]
- Werth, S. Biogeography and phylogeography of lichen fungi and their photobionts. In Biogeography of Microscopic Organisms: Is Everything Small Everywhere? Fontaneto, D., Ed.; Cambridge University Press: Cambridge, UK, 2011; pp. 191–208. [Google Scholar]
- Galloway, D.J. Lichen Biogeography. In Lichen Biology, 2nd ed.; Nash, T.H.I., Ed.; Cambridge University Press: New York, NY, USA, 2008; pp. 315–335. [Google Scholar]
- Geiser, L.H.; Jovan, S.E.; Glavich, D.A.; Porter, M.K. Lichen-based critical loads for atmospheric nitrogen deposition in Western Oregon and Washington Forests, USA. Environ. Pollut. 2010, 158, 2412–2421. [Google Scholar] [CrossRef]
- Geiser, L.H.; Neitlich, P.N. Air pollution and climate gradients in western Oregon and Washington indicated by epiphytic macrolichens. Environ. Pollut. 2007, 145, 203–218. [Google Scholar] [CrossRef]
- Branquinho, C.; Catarino, F.; Brown, D.H.; Pereira, M.J.; Soares, A. Improving the use of lichens as biomonitors of atmospheric pollution. Sci. Total Environ. 1999, 232, 67–77. [Google Scholar] [CrossRef]
- Printzen, C. Lichen Systematics: The Role of Morphological and Molecular Data to Reconstruct Phylogenetic Relationships. In Progress in Botany 71; Springer: Berlin/Heidelberg, Germany, 2009; Volume 71, pp. 233–275. [Google Scholar]
- Lumbsch, H.T.; Leavitt, S.D. Goodbye morphology? A paradigm shift in the delimitation of species in lichenized fungi. Fungal Divers. 2011, 50, 59–72. [Google Scholar] [CrossRef]
- Muggia, L.; Nelsen, M.P.; Kirika, P.M.; Barreno, E.; Beck, A.; Lindgren, H.; Lumbsch, H.T.; Leavitt, S.D. Formally described species woefully underrepresent phylogenetic diversity in the common lichen photobiont genus Trebouxia (Trebouxiophyceae, Chlorophyta): An impetus for developing an integrated taxonomy. Mol. Phylogenetics Evol. 2020, 149, 106821. [Google Scholar] [CrossRef]
- Argüello, A.; Del Prado, R.; Cubas, P.; Crespo, A. Parmelina quercina (Parmeliaceae, Lecanorales) includes four phylogenetically supported morphospecies. Biol. J. Linn. Soc. 2007, 91, 455–467. [Google Scholar] [CrossRef]
- Spribille, T. Relative symbiont input and the lichen symbiotic outcome. Curr. Opin. Plant Biol. 2018, 44, 57–63. [Google Scholar] [CrossRef]
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.-H.; Liow, L.H.; Nowak, M.D.; et al. Finding Evolutionary Processes Hidden in Cryptic Species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [CrossRef]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Clancy, J.; Jensen, J.; McMullin, R.T.; Wang, L.; Leavitt, S.D. Providing scale to a known taxonomic unknown—At least a 70-fold increase in species diversity in a cosmopolitan nominal taxon of lichen-forming fungi. J. Fungi 2022, 8, 490. [Google Scholar] [CrossRef]
- Roux, C.; Bertrand, M.; Poumarat, S.; Uriac, P. Quelques espèces nouvelles saxicoles–calcifuges de Lecanora du groupe polytropa (Ascomycota, Lecanoraceae) découvertes en France. Bull. Soc. Linn. Provence 2022, 73, 79–120. [Google Scholar]
- Huneck, S. Mitteilung über Flechteninhaltsstoffe. Die Inhaltsstoffe von Lecanora viridula (Flk.) Hillm., Lecanora polytropa (Ehrh.) Rabenh., Lecanora badia (Hoffm.) Ach. v. milvina Rabenh. und Lecanora sulphurea (Hoffm.) Ach. Z. Für Naturforschung 1966, 21B, 888–890. [Google Scholar] [CrossRef]
- Pérez-Ortega, S.; Spribille, T.; Palice, Z.; Elix, J.; Printzen, C. A molecular phylogeny of the Lecanora varia group, including a new species from western North America. Mycol. Prog. 2010, 9, 523–535. [Google Scholar] [CrossRef]
- Śliwa, L.; Flakus, A. Lecanora microloba, a new saxicolous species from Poland. Lichenol. 2011, 43, 1–6. [Google Scholar] [CrossRef]
- Yakovchenko, L.S.; Davydov, E.A.; Ohmura, Y.; Printzen, C. The phylogenetic position of species of Lecanora s. l. containing calycin and usnic acid, with the description of Lecanora solaris Yakovchenko & Davydov sp. nov. Lichenol. 2019, 51, 147–156. [Google Scholar] [CrossRef]
- Medeiros, I.D.; Mazur, E.; Miadlikowska, J.; Flakus, A.; Rodriguez-Flakus, P.; Pardo-De la Hoz, C.J.; Cieślak, E.; Śliwa, L.; Lutzoni, F. Turnover of lecanoroid mycobionts and their Trebouxia photobionts along an elevation gradient in Bolivia highlights the role of environment in structuring the lichen symbiosis. Front. Microbiol. 2021, 12, 774839. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. Notes 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Zhao, X.; Leavitt, S.D.; Zhao, Z.T.; Zhang, L.L.; Arup, U.; Grube, M.; Pérez-Ortega, S.; Printzen, C.; Śliwa, L.; Kraichak, E.; et al. Towards a revised generic classification of lecanoroid lichens (Lecanoraceae, Ascomycota) based on molecular, morphological and chemical evidence. Fungal Divers. 2016, 78, 293–304. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Rozewicki, J.; Yamada, K.D.; Katoh, K. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. 2019, 20, 1160–1166. [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Chethana, K.W.T.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Sites, J.W.; Marshall, J.C. Operational criteria for delimiting species. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 199–227. [Google Scholar] [CrossRef]
- Bradshaw, M.J.; Carey, J.; Liu, M.; Bartholomew, H.P.; Jurick II, W.M.; Hambleton, S.; Hendricks, D.; Schnittler, M.; Scholler, M. Genetic time traveling: Sequencing old herbarium specimens, including the oldest herbarium specimen sequenced from kingdom Fungi, reveals the population structure of an agriculturally significant rust. New Phytol. 2023, 237, 1463–1473. [Google Scholar] [CrossRef]
- Barclay, D.J.; Wiles, G.C.; Calkin, P.E. Holocene glacier fluctuations in Alaska. Quat. Sci. Rev. 2009, 28, 2034–2048. [Google Scholar] [CrossRef]
- Shafer, A.B.A.; Cullingham, C.I.; Côté, S.D.; Coltman, D.W. Of glaciers and refugia: A decade of study sheds new light on the phylogeography of northwestern North America. Mol. Ecol. 2010, 19, 4589–4621. [Google Scholar] [CrossRef]
- Gómez, A.; Lunt, D.H. Refugia within Refugia: Patterns of Phylogeographic Concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 155–188. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Vargas, P. Congruence between distribution modelling and phylogeographical analyses reveals Quaternary survival of a toadflax species (Linaria elegans) in oceanic climate areas of a mountain ring range. New Phytol. 2013, 198, 1274–1289. [Google Scholar] [CrossRef]
- Olalde, M.; Herrán, A.; Espinel, S.; Goicoechea, P.G. White oaks phylogeography in the Iberian Peninsula. For. Ecol. Manag. 2002, 156, 89–102. [Google Scholar] [CrossRef]
- Abellán, P.; Svenning, J.-C. Refugia within refugia—Patterns in endemism and genetic divergence are linked to Late Quaternary climate stability in the Iberian Peninsula. Biol. J. Linn. Soc. 2014, 113, 13–28. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Penas, Á.; Díaz González, T.E.; Cantó, P.; del Río, S.; Costa, J.C.; Herrero, L.; Molero, J. Biogeographic Units of the Iberian Peninsula and Baelaric Islands to District Level. A Concise Synopsis. In The Vegetation of the Iberian Peninsula: Volume 1; Loidi, J., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 131–188. [Google Scholar] [CrossRef]
- Pardo-De la Hoz, C.J.; Magain, N.; Lutzoni, F.; Goward, T.; Restrepo, S.; Miadlikowska, J. Contrasting Symbiotic Patterns in Two Closely Related Lineages of Trimembered Lichens of the Genus Peltigera. Front. Microbiol. 2018, 9, 2770. [Google Scholar] [CrossRef]
- Fernández-Mendoza, F.; Domaschke, S.; García, M.A.; Jordan, P.; Martin, M.P.; Printzen, C. Population structure of mycobionts and photobionts of the widespread lichen Cetraria aculeata. Mol. Ecol. 2011, 20, 1208–1232. [Google Scholar] [CrossRef]
- Pino-Bodas, R.; Stenroos, S. Global biodiversity patterns of the photobionts associated with the genus Cladonia (Lecanorales, Ascomycota). Microb. Ecol. 2021, 82, 173–187. [Google Scholar] [CrossRef]
- Altermann, S.; Goward, T.; Leavitt, S.D. Tidying up the genus Letharia: Introducing L. lupina sp. nov. and a new circumscription for L. columbiana. Lichenol. 2016, 48, 423–439. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Antarctica | 2 (n = 4) |
| Asia | 21 (n = 91) |
| Europe | 49 (n = 97) |
| North America | 26 (n = 84) |
| South America | 2 (n = 4) |
| Intercontinental | 21 (228) |
| unknown | 2 (n = 2) |
| Current worldwide sampling | 123 (n = 510) |
| Distribution of SHs | Chugach Mountains | Kenai Mountains |
|---|---|---|
| Intercontinental | 8 1 | 6 1 |
| Unique to site | 3 | 3 |
| Total SHs | 11 | 9 |
| Distribution of SHs | Sierra de Guadarrama | Laguna Negra y Circos Glaciares de Urbion | Ponga Biosphere Reserve |
|---|---|---|---|
| Intercontinental SHs | 6 | 6 | 2 |
| SHs unique to site | 10 | 6 | 4 |
| Regional SHs | 2 | 2 | 4 |
| Total SHs | 18 | 14 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woodhouse, J.; Pérez-Ortega, S.; Roux, C.; Bertrand, M.; Leavitt, S.D. Diverse Communities of Endemic and Cosmopolitan Lineages at Local Sites in the Lecanora polytropa Aggregate (Ascomycota). Diversity 2024, 16, 88. https://doi.org/10.3390/d16020088
Woodhouse J, Pérez-Ortega S, Roux C, Bertrand M, Leavitt SD. Diverse Communities of Endemic and Cosmopolitan Lineages at Local Sites in the Lecanora polytropa Aggregate (Ascomycota). Diversity. 2024; 16(2):88. https://doi.org/10.3390/d16020088
Chicago/Turabian StyleWoodhouse, John, Sergio Pérez-Ortega, Claude Roux, Michel Bertrand, and Steven D. Leavitt. 2024. "Diverse Communities of Endemic and Cosmopolitan Lineages at Local Sites in the Lecanora polytropa Aggregate (Ascomycota)" Diversity 16, no. 2: 88. https://doi.org/10.3390/d16020088
APA StyleWoodhouse, J., Pérez-Ortega, S., Roux, C., Bertrand, M., & Leavitt, S. D. (2024). Diverse Communities of Endemic and Cosmopolitan Lineages at Local Sites in the Lecanora polytropa Aggregate (Ascomycota). Diversity, 16(2), 88. https://doi.org/10.3390/d16020088