Abstract
The diversity of parasite communities is mainly driven by evolutionary history, as well as the ecology of the host species. To test whether the diversity of the parasite community of four related Stromateidae (Pisces: Scombriformes) is related to evolutionary history (the host phylogeny) or the host’s geographical distribution, we analyzed the metazoan parasite fauna of four species of fishes of this family, from the Pacific and Atlantic coasts of South America. Studied species were Peprilus snyderi (samples from Callao, Perú, and Antofagasta, Chile), Peprilus medius (Chorrillos, Perú), Peprilus paru (Rio de Janeiro, Brazil) and Stromateus stellatus (Talcahuano, Chile). Our multivariate analysis strongly suggests that the diversity of the parasite fauna of the studied fishes is driven mainly by the host’s geographical distribution and not the host phylogeny.
1. Introduction
The composition of the metazoan parasite communities of a given host species is strongly influenced by the evolutionary history, as well as the ecology of their host species, (i.e., diet and geographical distribution of the host species) [1,2,3]. Through their evolutionary history, hosts species can lose or acquire parasites due the evolution of native parasites or gain new parasite species from other host species due to sympatric distribution and/or phylogenetic affinities [4,5]. Although many studies suggest the importance of host phylogeny as a force that drives the parasite’s community structure, few have integrated this information into diversity studies and parasite community analyses [1,6,7]. The importance of ecological and phylogenetic factors, when analyzed simultaneously, have been explored [2,3,8,9,10], among a handful of others, mainly due that is not easy to found close related host species (i.e., same genus or family) living in sympatry. Specifically, similarity in parasite fauna tends to decrease exponentially, but not strongly, with increasing phylogenetic divergence, as well as geographical distance between host species [8,11]. However, a clear pattern between fish phylogeny and some biological traits of the host (diet, body size) with the characteristics of the endohelminth fauna of five species of Cheilininae (Labridae) from Lizard Island, Austalia, was not found [9], except for the two most closely phylogenetically related species, which showed the most similar diets and body size, as well as the most similar parasite communities. Fish ecology rather than host phylogeny was the most important factor driving similarity in the parasite composition of five species of Gadiformes belonging to three families from Southern Chile; similarity in parasite communities was explained by the similar diet of the hosts and the predominance of generalist larval species [2]. A similar conclusion was reached [3] based on the parasitological analysis of five deep-sea teleosts (>1000 m deep), from Northern Chile, but phylogenetic relationships cannot be discarded, as indicated by the presence of highly specific digeneans such as Gibsonia hastata Gaevskaya and Rodyuk, 1988, Profundivermis intercalarius Bray & Gibson, 1991 and Glomericirrus macrouri Gaevskaja, 1979; therefore, host phylogeny was a critical issue in determining endoparasite richness. Another approach, but scarcely explored, is the analysis of the parasite fauna of closely phylogenetically related host species from contrasting environments that in turn will explain an ecological relationship, i.e., members of the same genus but from different biogeographic regions or oceans. The infracommunity analysis of two species of Menticirrhus (Sciaenidae), M. ophicephalus (Yenyns, 1840) from the Pacific (12°30′ S) and M. americanus (Linnaeus, 1758) in the Atlantic (21–23° S) showed a differential dominance of ectoparasites (higher in the Pacific) and endoparasites (higher in the Atlantic). The lower richness of the endoparasite fauna in the Pacific was explained because of an unstable environment due to an upwelling system, periodically affected by the El Niño Southern oscillation (ENSO), characteristic of the Humboldt Current System [12].
The effect of host phylogeny, when searching for forces behind the structure of parasite communities, is a well-known confounding factor. Closely related host species are expected to harbor similar numbers of parasite species and possibly taxonomically related parasite species, if these were inherited from a recent common ancestor [13]. This is especially true when the studied dataset includes host species that are not closely related. Consequently, the study of parasite communities of close related species, ideally members of the same genus, will avoid the impact of host phylogeny. Few studies have been developed under this approach in the marine realm [9,10,14]. The Pacific Stromateidae studied herein shows a wide geographical range, with Peprilus medius (Peters, 1879) and Peprilus snyderi Gilberts and Starks, 1904 overlapping along their geographical range, from Baja California Sur (25° N) to Chorrillos, Central Perú (12°10′ S), but P. snyderi extend as far as Antofagasta (23°30′ S). Stromateus stellatus Cuvier, 1829 is distributed from Southern Chile (47° S) to Central Perú (12°), overlapping with P. snyderi between 23°30′ S and 12°10′ S, whereas Peprilus paru (Linnaeus, 1758) is found from the Elizabeth Islands (41°30′ N) to Argentina (35°40′ S). (Figure 1). Here, we evaluated the metazoan parasite fauna of two closely related genera of Stromateidae, Stromateus Linnaeus, 1758 and Peprilus Cuvier, 1829, from the Atlantic and Pacific coasts of South America, in order to test if the phylogeny of the host or their geographical distribution explain similarities in their parasite diversity.
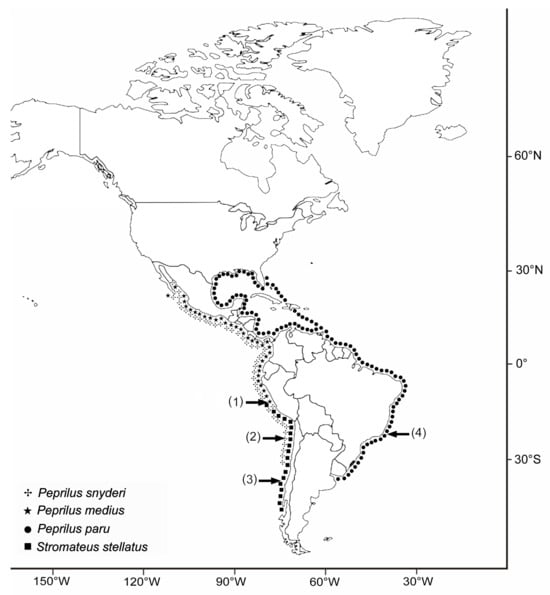
Figure 1.
Known distribution of the host species. Approximate position of localities where samples were obtained. 1: Chorrillos and Callao (Perú), 2: Antofagasta (Chile), 3: Talcahuano (Chile), 4: Rio de Janeiro (Brazil).
2. Materials and Methods
We used published data of parasites of P. snyderi (n = 214) from Central Perú [15], P. paru (n = 81) from Brazil [16], P. medius (n = 115) from Perú [17] and our unpublished data for S. stellatus (n = 25) from Central Chile (Talcahuano) and a small sample of P. snyderi (n = 9) from Northern Chile (Antofagasta). All fishes were obtained from local fishermen. Specimens of S. stellatus and P. snyderi from Chile (Talcahuno, 36°40′ S and Antofagasta, 23°30′ S, respectively) were examined for parasites. Fishes were caught by local fishermen, transported to the laboratory and were deep frozen (−18 °C) until examination. After thawing, fishes were measured for total length (cm) and sexed. All specimens were examined first for ectoparasites, including skin, gills and mouth cavity, and then for endoparasites. All the viscera, including heart and blood vessels (arterial cone and branchial artery) were examined. To count endoparasites, each visceral organ was dissected separately and washed in running water and all the material retained on a 0.3 mm mesh was examined under a LEICA M 125 stereoscope, with an incorporated LEICA MC120 HD camera (Heerbrugg, Switzerland). Parasites were fixed, preserved and stained according to standard techniques in parasitology [18].
The prevalence and mean intensity of infection for each parasite species were calculated [19]. Cluster analyses (based on Euclidean distance and complete linkage algorithm) and the SIMPROF test (999 permutations) as implemented in PRIMER 6.0 [20] were used to determine similarity in the parasite composition (prevalence) among host species. Correspondence analyses allowed us to evaluate host–parasite associations. All multivariate analyses were performed with Statistica 6.0 software (StatSoft Inc., Tulsa, OK, USA).
3. Results
Twenty-five parasite species were found, of which six were ectoparasites; there were eight endoparasite species at larval stages, and the remaining eleven species were adults. (Table 1). Trematoda (Digenea and Aspidogastrea) represented 48% of the recorded species. The number of parasite species ranged from seven (S. stellatus) to ten species (P. snyderi Peruvian sample). The richness was not correlated with sample size, previous log transformation of data (r = 0.686, gl = 3, p = 0.201).

Table 1.
Metazoan parasite species found in four species of Stromateid from South America. P = prevalence of infection, MI = mean intensity of infection.
Two species, the digenean Lecithocladium cristatum (Rudolphi, 1819) and the larval nematode Hysterothylacium sp., were common to all samples. The monogenean Microcotyle sp. Was found only in P. parus from Brazil, whereas Oaxacotyle oaxacensis was found only in Peprilus spp. from the Pacific coast of America. The copepod Caligus sp. was found in Peprilus spp. from the Atlantic and Pacific Ocean, but not in S. stellatus.
The cluster analysis based on parasite prevalence previous arcsine transformation, showed that the shorter Euclidean distance (60.18) was between P. medius and P. snyderi (Chilean sample), whereas the distance between this clade and P. snyderi (Peruvian sample) was 69.06. The distance between S. stellatus and Pacific samples of Peprilus was 89.43 (π = 2.7, p = 0.731), and finally the distance between P. paru from Brazil and the Pacific samples was 153.66 (π = 2.7, p = 0.50). (Figure 2).
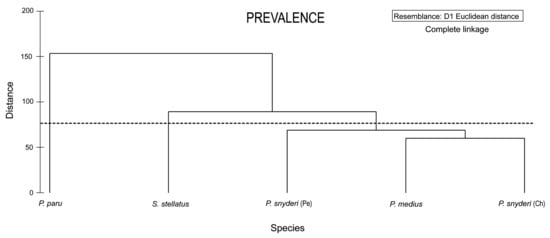
Figure 2.
Similarity dendrogram of parasite communities of P. medius, P. snyderi, P. paru and S. stellatus based on prevalence data. Horizontal line indicates significant groups.
The correspondence analysis from the prevalence data (Figure 3) showed significant differences in parasite composition among the host species (χ2 = 408, df = 96, p < 0.001); 77.3% of the variation was explained by the first two dimensions (43.2% and 34.1% for the first and second dimensions, respectively). High values for inertia corresponded to the monogenean Microcotyle sp., the digenean Opechona sp., the nematode Rhapidascaris sp. and the isopod C. gaudichaudii. P. paru was well separated in the first dimension.
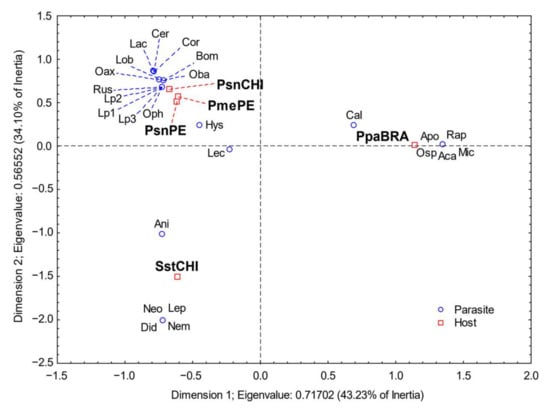
Figure 3.
Results of the correspondence analysis based on the prevalence of infection. PsnCHI= P. snyderi sample from Chile, Psn(PE) = P. snyderi sample from Perú, Pme = P. medius, Sst = S. stellatus. Codes for parasites are as in Table 1.
The correspondence analysis based on the mean intensity of infection (Figure 4) showed significant differences among host species (χ2 = 1846, df = 96, p < 0.001); 76.8% of the variation was explained by the first two dimensions (46.1% and 30.7% for the first and second dimensions, respectively). Stromateus stellatus was well separated in the first dimension, and high values for inertia corresponded to the digenean Lepocreadiidae gen. sp., L. cristatum and Dydimozoidea gen. sp., and the monogenean Neogrubea seriolellae Dillon and Hargis, 1968.
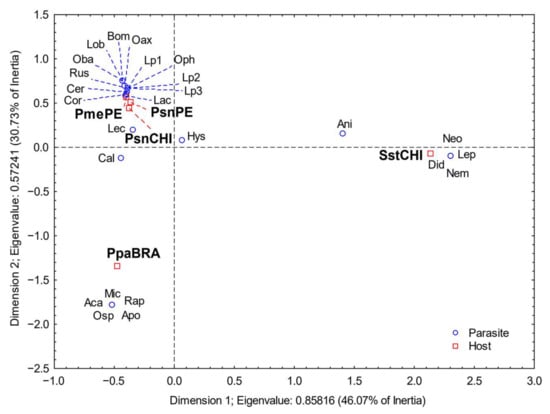
Figure 4.
Results of the correspondence analysis based on mean intensity of infection. PsnCHI= P. snyderi sample from Chile, Psn(PE) = P. snyderi sample from Perú, Pme = P. medius, Sst = S. stellatus. Code for parasites as in Table 1.
4. Discussion
The resemblance among fish parasite communities could be explained by the critical role of host phylogeny and/or ecological convergence, specifically in terms of habitat use or trophic levels [21]. The study of three closely related and sympatric flounders from Argentina suggests that similar ecological filters produce analogous infections across host species driving towards homogeneous parasite communities [10]; that is, ecology will be the force behind the structure of parasite communities, rather than host phylogeny.
With regard to host phylogeny, closely related host species are expected to harbor taxonomically related parasite species because these were inherited from a recent common ancestor [13]; consequently, high host specificity is expected. In a similar way, the similarity in the composition of host parasite fauna and communities will decrease as geographical distance increases, and also phylogenetic distance will increase with increased geographical distance [8,11]. Then, it is crucial to take phylogenetic relationships into account when trying to determine which host features are associated with diverse parasite fauna [21,22], specifically if biological traits of many unrelated host species (i.e., different families, orders or higher taxonomic categories) are analyzed in the search for patterns in parasite communities. But if closely related host species are analyzed, congeneric species (or members of closely related genera) will be the best biological model in order to avoid the expected effect of host phylogeny.
Stromateidae include three genera: Pampus Bonaparte, 1834, Peprilus and Stromateus [23]; Pampus is widely distributed along the coasts of the Northwest Pacific and north Indian Oceans [24], whereas Peprilus is found in both Pacific and Atlantic coasts of America [25]. Stromateus is widely distributed along the coasts of the Mediterranean, the West African coast to Cape Town and the southern Atlantic and Pacific coasts of South America. It is the only genus of the family with a transoceanic distribution along South America, with the Atlantic species S. paru and S. brasiliensis Fowler, 1906 and the Pacific S. stellatus. Among the Stromateidae, Peprilus and Stromateus have been considered as the closest related genera in the family [26]. Due to the close phylogenetic relationship between both genera, the search for a phylogenetic signal is not required.
The lower Euclidean distance between members of Peprilus from the Pacific coast of South America suggests a high similitude in their parasite fauna in accordance with the expected relationship between host phylogeny and parasite communities, but the higher distance of P. paru from the Atlantic coast of South America could reject the hypothesis of host phylogeny, and even more so when P. paru is the most distant species and S. stellatus is more related to Peprilus from the Pacific Ocean, suggesting that geographical origin is a better explanation for the specific characteristics of the observed parasite communities. As stated earlier [27], parasitological relationships between widely distributed species (of similar families) are influenced by the overlapping distribution of the host species, which in turn can be associated with biogeographic regions. Along the Pacific coast of South America, the distribution of Peprilus spp. overlaps along the Peruvian Province [28] but does not reach the transitional zone, whereas S. stellatus is found from the Magellanic province to the Peruvian province. P. snyderi and S. stellatus overlap between 12° S (ca. Callao Perú) and 23°30′ S (ca. Antofagasta, Chile). That is, host species from the same biogeographic province show higher similarity in their parasite fauna evaluated at the community level. A similar picture is evident from the results of the correspondence analysis. The biogeographical patterns of prey distribution are key determinants of the endoparasite community of the red rockfish Sebastes capensis, along their whole distribution [29]. Contrarily, the ectoparasite community of the same host species does not follow the biogeographical pattern from free-living organisms along the whole distribution of the host, but allows the transitional zone of the south-eastern Pacific to be distinguished, and on a broad geographical scale. Therefore, it is possible to distinguish the ectoparasite communities of S. capensis in the south-eastern Pacific (as a whole) from those of the Atlantic coast of South America and South Africa [30]. In a similar way, the assemblage of larval parasites of Percophis brasiliensis Quoy and Gaimard, 1825 (Percophidae: Perciformes) reflect the biogeography of the Southwest Atlantic [31]. Our results show that the structure of the whole metazoan community of parasites of the four species of Stromateidae are a good indicator of the biogeographic provinces [28] in the Southeastern region. Data from P. paru from Brazil suggest that similarity in parasite communities in closely related host species (Genus Peprilus) that inhabit different biogeographical provinces is not driven by host phylogeny. For instance, the monogenean O. oaxacensis was found only in Peprilus spp. from the Pacific coast of North and South America [15,17,32], and not from P. parus from Brazil, whereas a non-related monogenea of the genus Microcotyle was found (see Table 1). In addition, the monogenea N. seriolellae from S. stellatus from central Chile has been found in the pelagic Seriolella porosa Guichenot 1848 (Centrolophidae) from the Patagonian coast of Argentina, and also in S. brasiliensis Fowler, 1906 in the Southwestern Atlantic, but not in P. paru [33]. Our results strongly suggest that biogeography is the main force behind the diversity as well as the structure of metazoan parasites of four species of Stromateidae from South America.
Author Contributions
Conceptualization, M.E.O. and L.A.Ñ.; methodology, M.E.O., L.A.Ñ. and J.L.L.; software, L.A.Ñ.; validation, M.E.O.; L.A.Ñ., R.E. and J.L.L.; formal analysis, M.E.O. and L.A.Ñ.; investigation, M.E.O., L.A.Ñ., R.E. and J.L.L.; resources, M.E.O. and R.E.; data curation, L.A.Ñ.; writing—original draft preparation, M.E.O. and L.A.Ñ.; review and editing, M.E.O., L.A.Ñ., R.E. and J.L.L.; funding acquisition, M.E.O. and R.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by MINEDUC-UA project, ANT 1855, Plan de Fortalecimiento Universidades Estatales—Chile RES21992 and Millennium Institute of Oceanography (IMO), IC120019.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available from the corresponding authors on request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Poulin, R. Phylogeny, ecology, and the richness of parasite communities in vertebrates. Ecol. Monogr. 1995, 65, 283–302. [Google Scholar] [CrossRef]
- Chávez, R.A.; Gonzalez, M.T.; Oliva, M.E.; Valdivia, I.M. Endoparasite fauna of five Gadiformes fish species from the coast of Chile: Host ecology versus phylogeny. J. Helminthol. 2012, 86, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Ñacari, L.A.; Oliva, M.E. Metazoan parasites of deep-sea fishes from the Southeastern Pacific: Exploring the role of ecology and host phylogeny. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2016, 115, 123–130. [Google Scholar] [CrossRef]
- MacLeod, C.J.; Paterson, A.M.; Tompkins, D.M.; Duncan, R.P. Parasites lost—Do invaders miss the boat or drown on arrival? Ecol. Lett. 2010, 13, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R.; Rohde, K. Comparing the richness of metazoan ectoparasite communities of marine fishes: Controlling for host phylogeny. Oecologia 1997, 110, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Morand, S.; Cribb, T.H.; Kulbicki, M.; Rigby, M.C.; Chauvet, C.; Dufour, V.; Faliex, E.; Galzin, R.; Lo, C.M.; Lo-Yat, A.; et al. Endoparasite species richness of New Caledonian butterfly fishes: Host density and diet matter. Parasitology 2000, 121, 65–73. [Google Scholar] [CrossRef]
- Bush, A.O.; Aho, J.M.; Kennedy, C.R. Ecological versus phylogenetic determinants of helminth parasite community richness. Evol. Ecol 1990, 4, 1–20. [Google Scholar] [CrossRef]
- Poulin, R. Decay of similarity with host phylogenetic distance in parasite faunas. Parasitology 2010, 137, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, G.; Grutter, A.S.; Cribb, T.H. Endoparasite communities of five fish species (Labridae: Cheilininae) from Lizard Island: How important is the ecology and phylogeny of the hosts? Parasitology 2006, 132, 363–374. [Google Scholar] [CrossRef]
- Alarcos, A.J.; Timi, J.T. Parasite communities in three sympatric flounder species (Pleuronectiformes: Paralichthyidae) Similar ecological filters driving toward repeatable assemblages. Parasitol. Res. 2012, 110, 2155–2166. [Google Scholar] [CrossRef]
- Oliva, M.E.; González, M.T. The decay of similarity over geographic distance in parasite communities of marine fishes. J. Biogeogr. 2005, 32, 1327–1332. [Google Scholar] [CrossRef]
- Luque, J.L.; Oliva, M.E. Metazoan Parasite Infracommunities of Menticirrhus (Teleostei: Sciaenidae): An Amphi-Oceanic Approximation. J. Parasitol. 1999, 85, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Luque, J.L.; Mouillot, D.; Poulin, R. Parasite biodiversity and its determinants in coastal marine teleost fishes of Brazil. Parasitology 2004, 128, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, G.; Cortés, Y. Parasite communities of a fish assemblage from the intertidal rocky zone of central Chile: Similarity and host specificity between temporal and resident fish. Parasitology 2009, 136, 1291–1303. [Google Scholar] [CrossRef] [PubMed]
- Ñacari, L.A.; Sanchez, L. Helminth fauna of Peprilus snyderi Gilbert & Starks, 1904 (Stromateidae) of Chorrillos fishmarket, Lima, Perú. Neotrop. Helminthol. 2014, 8, 1–17. [Google Scholar]
- Azevedo, R.K.; Abdallah, V.D.; Luque, J.L. Aspectos quantitativos da comunidade de metazoários parasitos do gordinho Peprilus paru (Linnaeus, 1758) (Perciformes: Stromateidae), do litoral do estado do Rio de Janeiro, Brasil. Rev. Bras. Parasitol. Vet. 2007, 16, 10–14. [Google Scholar] [PubMed]
- Iannacone, J.; Alvarino, L. Influencia del tamaño y sexo de Peprilus medius (Peters) (Stromateidae: Perciformes) capturados en Chorrillos, Lima, Perú, sobre su comunidad parasitaria. Neotrop. Helminthol. 2008, 2, 62–70. [Google Scholar] [CrossRef]
- Pritchard, M.H.; Kruse, G.O. The Collection and Preservation of Animal Parasites; Technical Bulletin 1; The Harold W. Manter Laboratory, University of Nebraska Press: Lincoln, NE, USA, 1982; p. 141. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial. Primer-E; Plymouth Marine Laboratory: Plymouth, UK, 2006; p. 193. [Google Scholar]
- Carrassón, M.; Dallarés, S.; Cartes, J.E.; Constenla, M.; Pérez-del-Olmo, A.; Zucca, L.; Kostadinova, A. Drivers of parasite community structure in fishes of the continental shelf of the Western Mediterranean: The importance of host phylogeny and autecological traits. Int. J. Parasitol. 2019, 49, 668–669. [Google Scholar] [CrossRef]
- Poulin, R. Parasite species richness in New Zealand fishes: A grossly underestimated component of biodiversity? Diversity Distrib. 2004, 10, 31–37. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; Wiley: Hoboken, NJ, USA, 2006; p. 752. [Google Scholar]
- Yin, G.; Pan, Y.; Sarker, A.; Baki, M.A.; Kim, J.-K.; Wu, H.; Li, C. Molecular systematics of Pampus (Perciformes: Stromateidae) based on thousands of nuclear loci using target-gene enrichment. Mol. Phyl. Evol. 2019, 140, 106595. [Google Scholar] [CrossRef]
- Marceniuk, A.P.; Caires, R.; Siccha-Ramirez, R.; Oliveira, C. Review of the harvestfishes, genus Peprilus (Perciformes: Stromateidae), of the Atlantic coast of South America. Zootaxa 2016, 4098, 311–332. [Google Scholar] [CrossRef]
- Doiuchi, R.; Sato, T.; Nakabo, T. Phylogenetic relationships of the stromateoid fishes (Perciformes). Ichthyol. Res. 2004, 51, 202–212. [Google Scholar] [CrossRef]
- Marques, J.F.; Santos, M.J.; Cabral, H.N. Zoogeographical patterns of flatfish (Pleuronectiformes) parasites in the Northeast Atlantic and the importance of the Portuguese coast as a transitional area. Sci. Mar. 2009, 73, 461–471. [Google Scholar] [CrossRef][Green Version]
- Camus, P.A. Biogeografía marina de Chile continental. Rev. Chil. Hist Nat. 2001, 74, 587–617. [Google Scholar] [CrossRef]
- González, M.T.; Barrientos, C.; Moreno, C.A. Biogeographical patterns in endoparasite communities of a marine fish (Sebastes capensis Gmelin) with extended range in the Southern Hemisphere. J. Biogeogr. 2006, 33, 1086–1095. [Google Scholar] [CrossRef]
- González, M.T.; Moreno, C.A. The distribution of the ectoparasite fauna of Sebastes capensis from the southern hemisphere does not correspond with zoogeographical provinces of free-living marine animals. J. Biogeogr. 2005, 32, 1539–1547. [Google Scholar] [CrossRef]
- Braicovich, P.E.; Pantoja, C.; Pereira, A.N.; Luque, J.L.; Timi, J.T. Parasites of the Brazilian flathead Percophis brasiliensis reflect West Atlantic biogeographic regions. Parasitology 2017, 144, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Garfias, B.; Garcia-Prieto, L.; Perez-Ponce de Leon, G. Checklist of the monogenea (Platyhelminthes) parasitic in mexican aquatic vertebrates. Zoosystema 2017, 39, 501–598. [Google Scholar] [CrossRef]
- Hernández-Orts, J.S.; Alama-Bermejo, G.; Crespo, E.A.; Raga, J.A.; Montero, F.E. A revision of the genus Neogrubea Dillon & Hargis, 1968 (Monogenea: Mazocraeidae): New morphological and molecular data from off the Patagonian coast of Argentina. Syst. Parasitol. 2014, 89, 59–72. [Google Scholar] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).