
The Forest Refugium of the Bükk Mountains, Hungary—Vegetation Change and Human Impact from the Late Pleistocene

Abstract
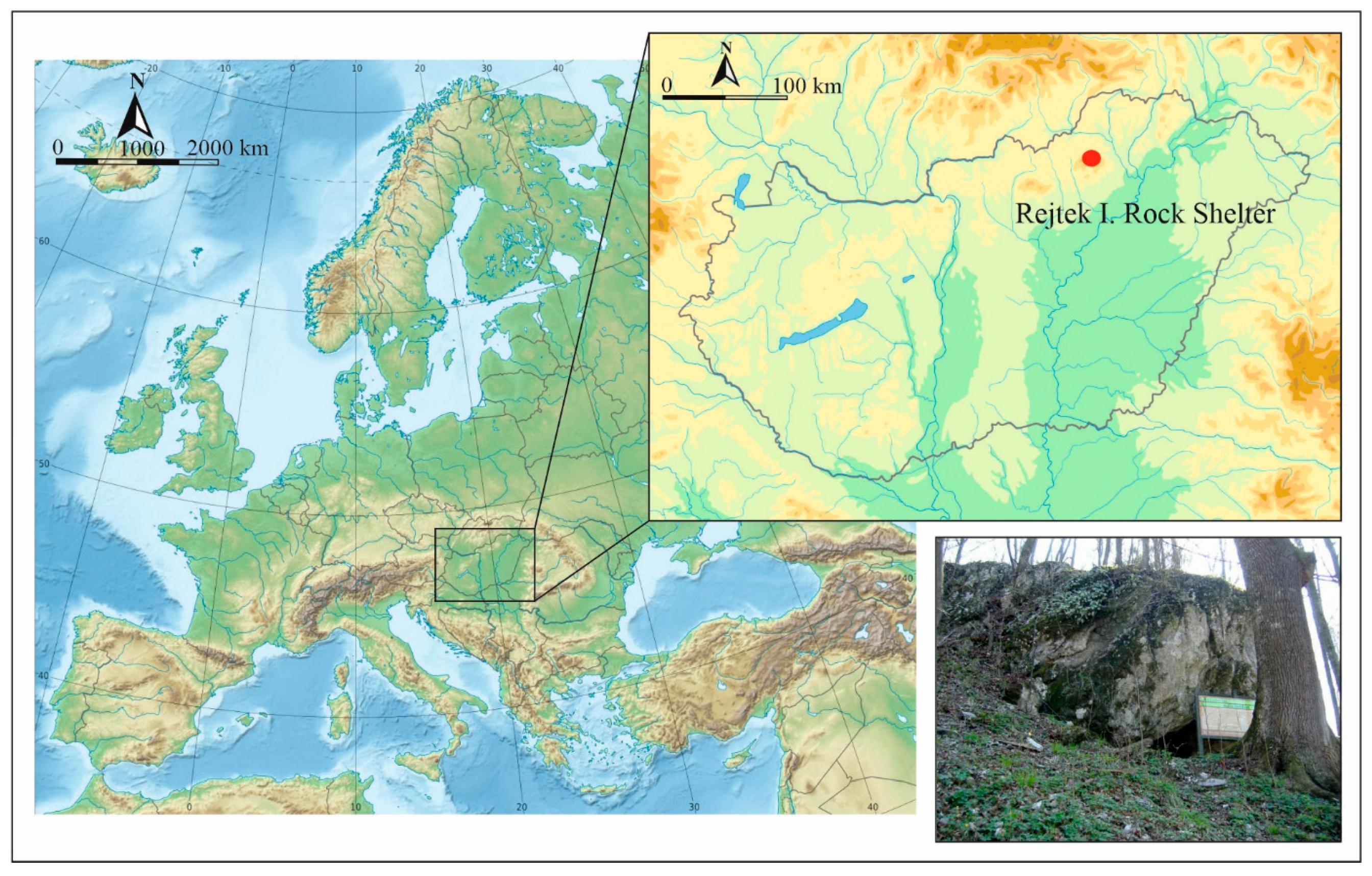
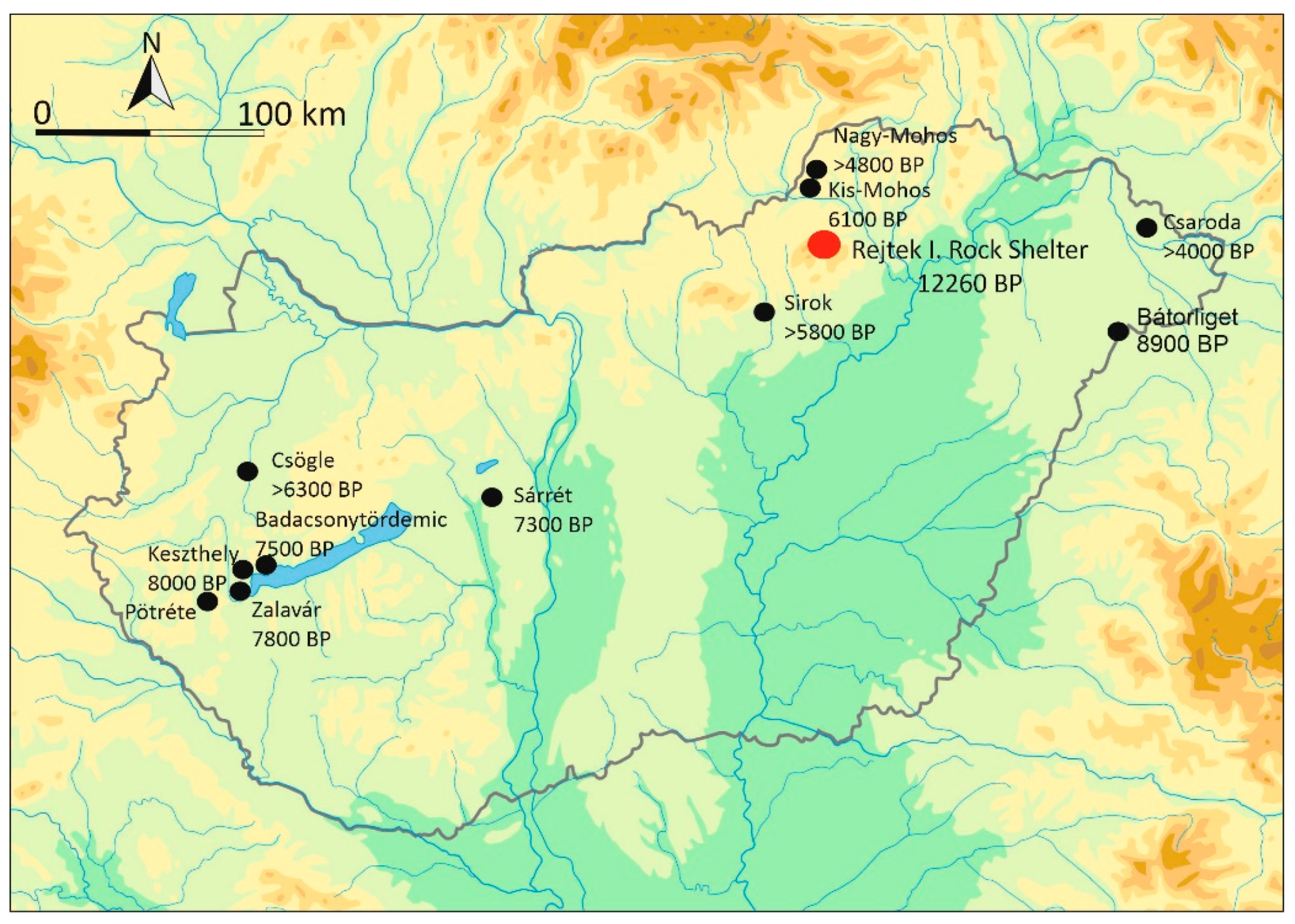
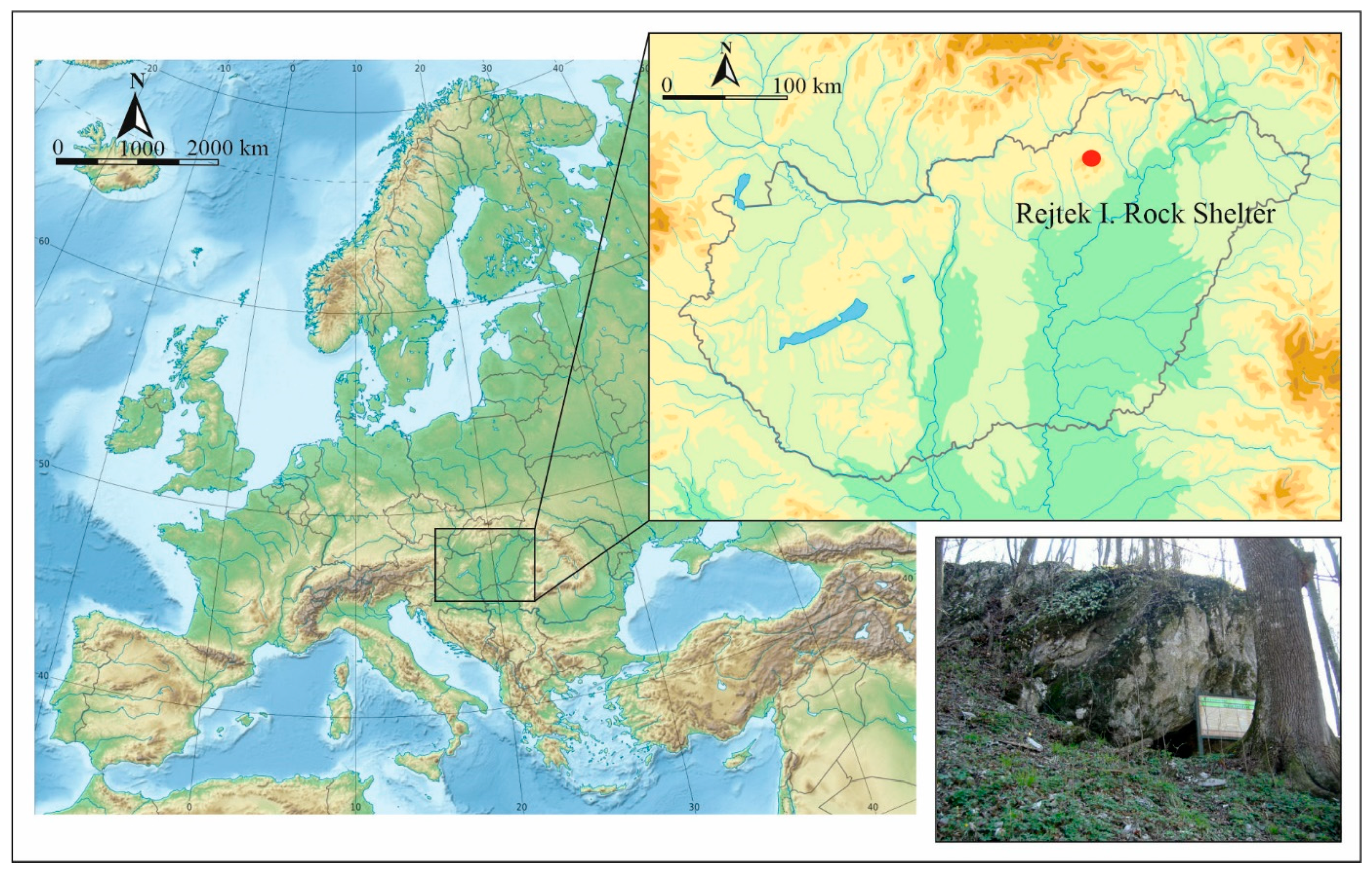
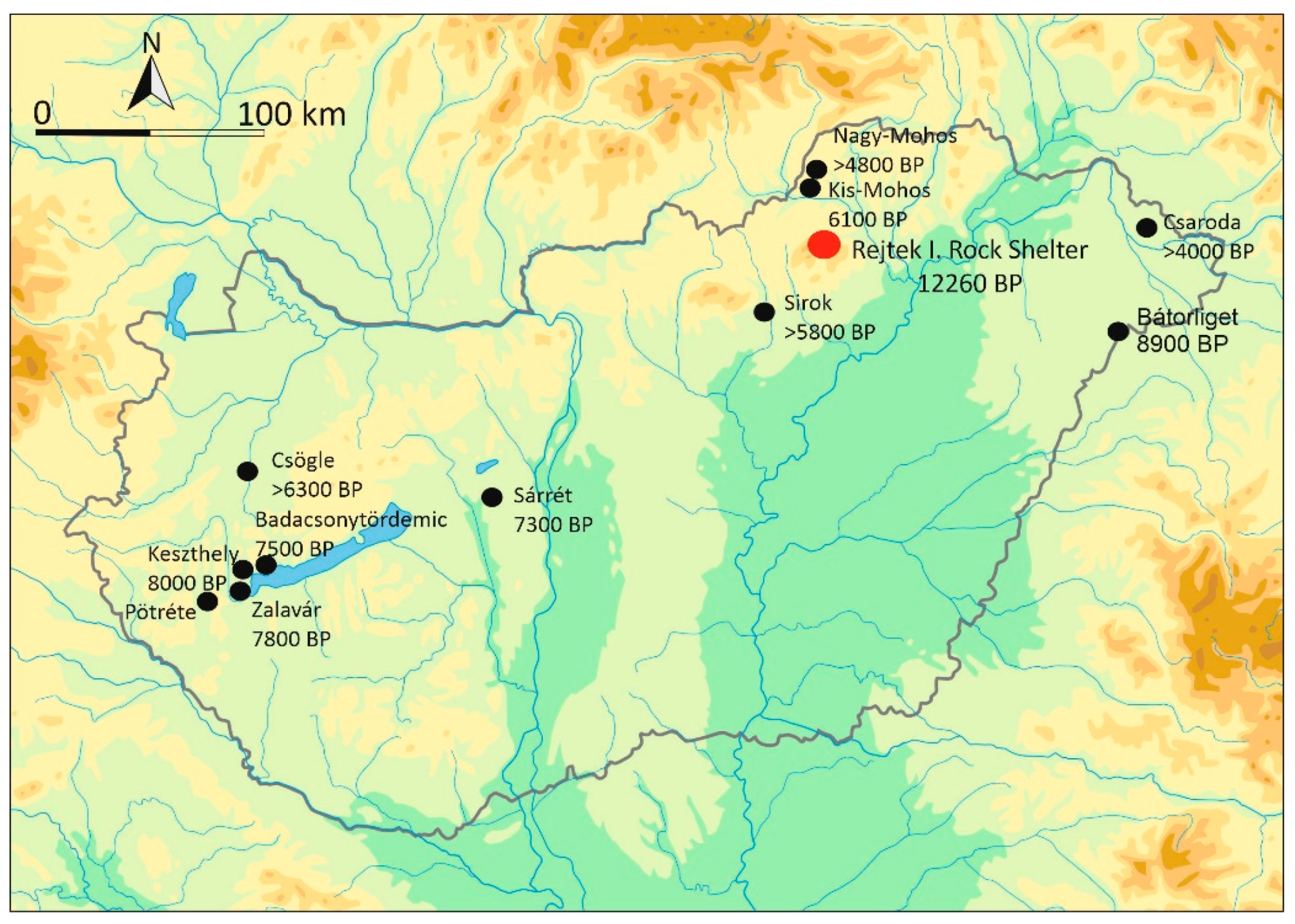
1. Introduction
2. Materials and Methods
3. Results
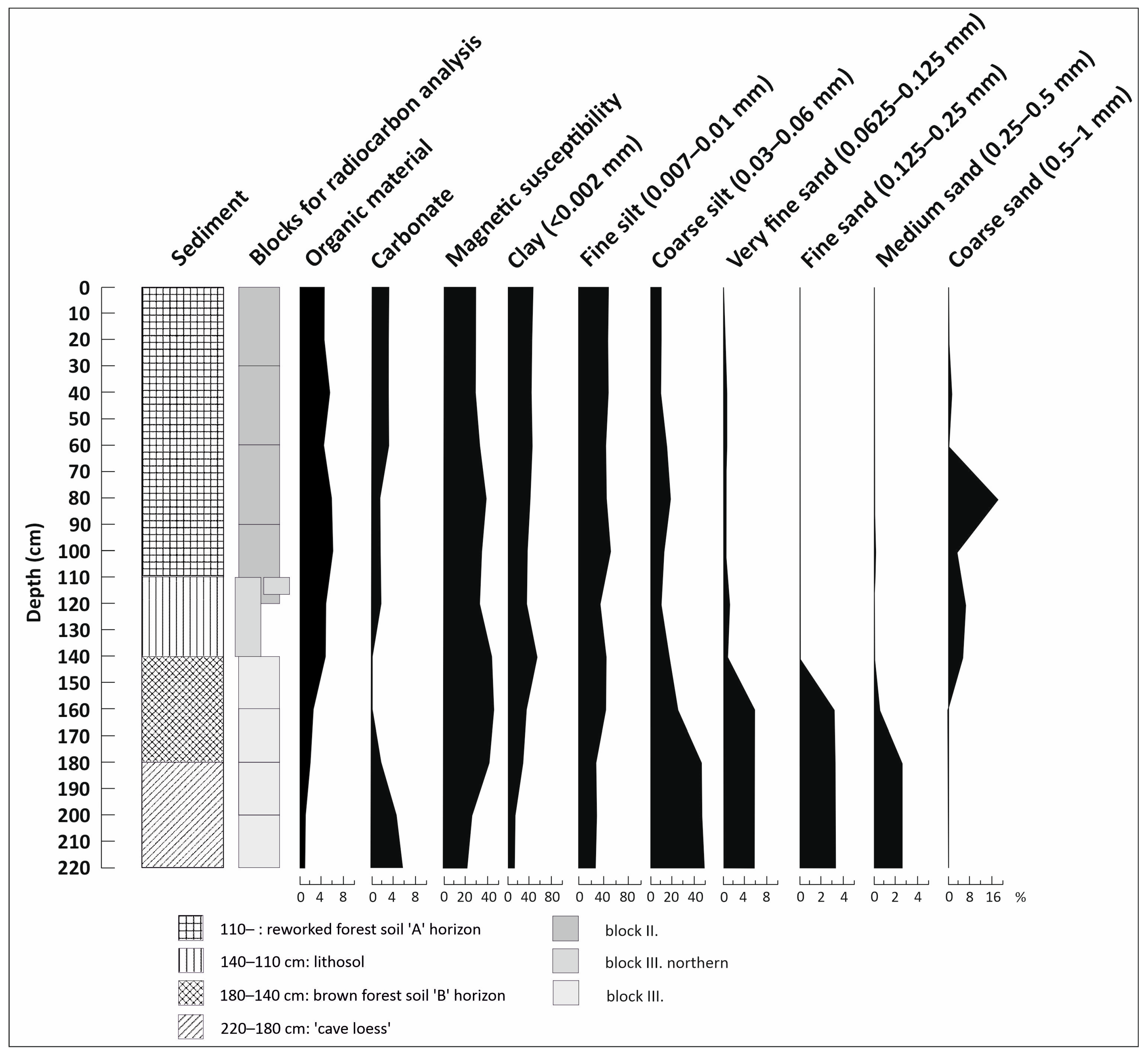
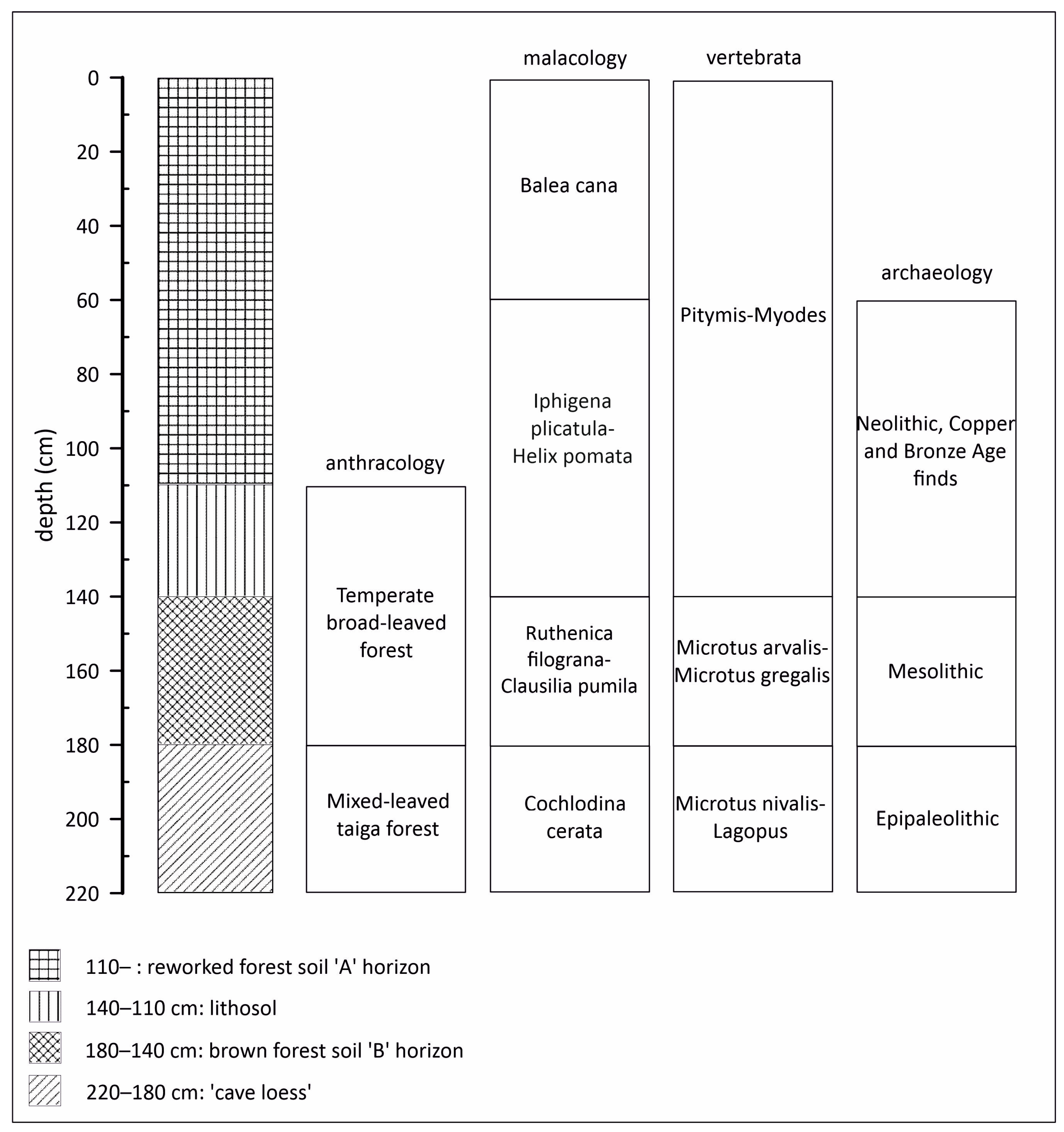
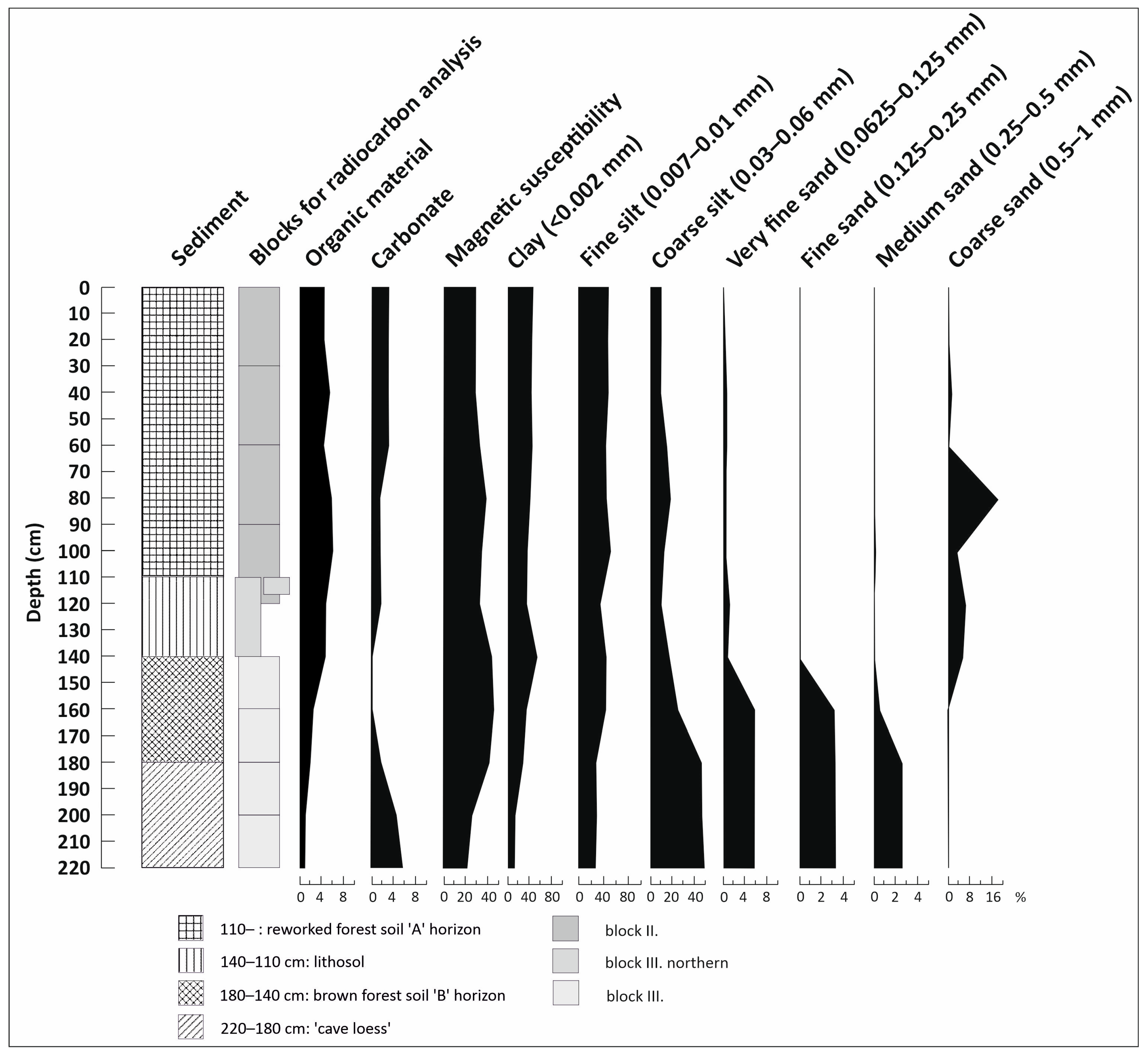
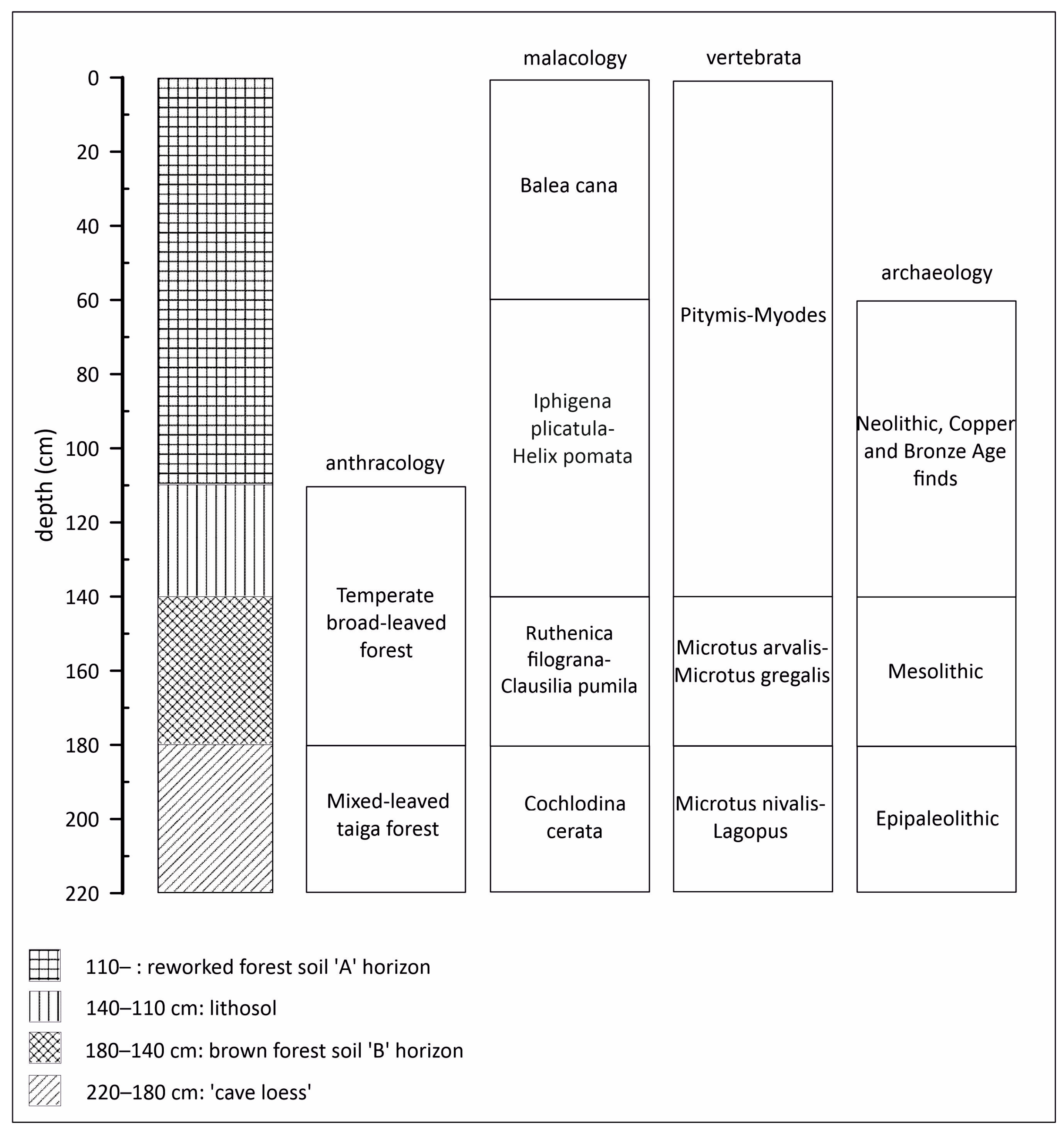
3.1. Lithological Observations and Analysis
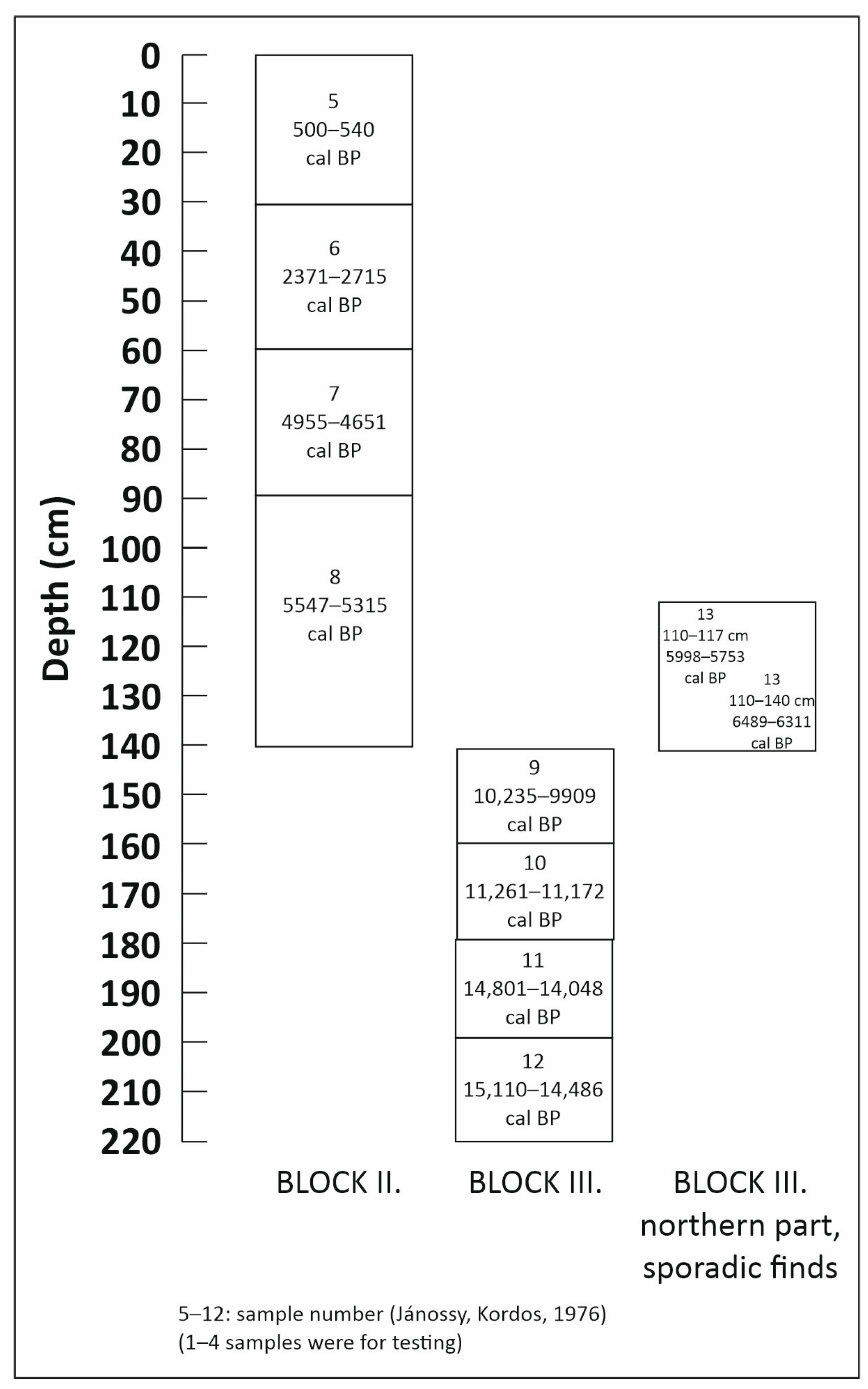
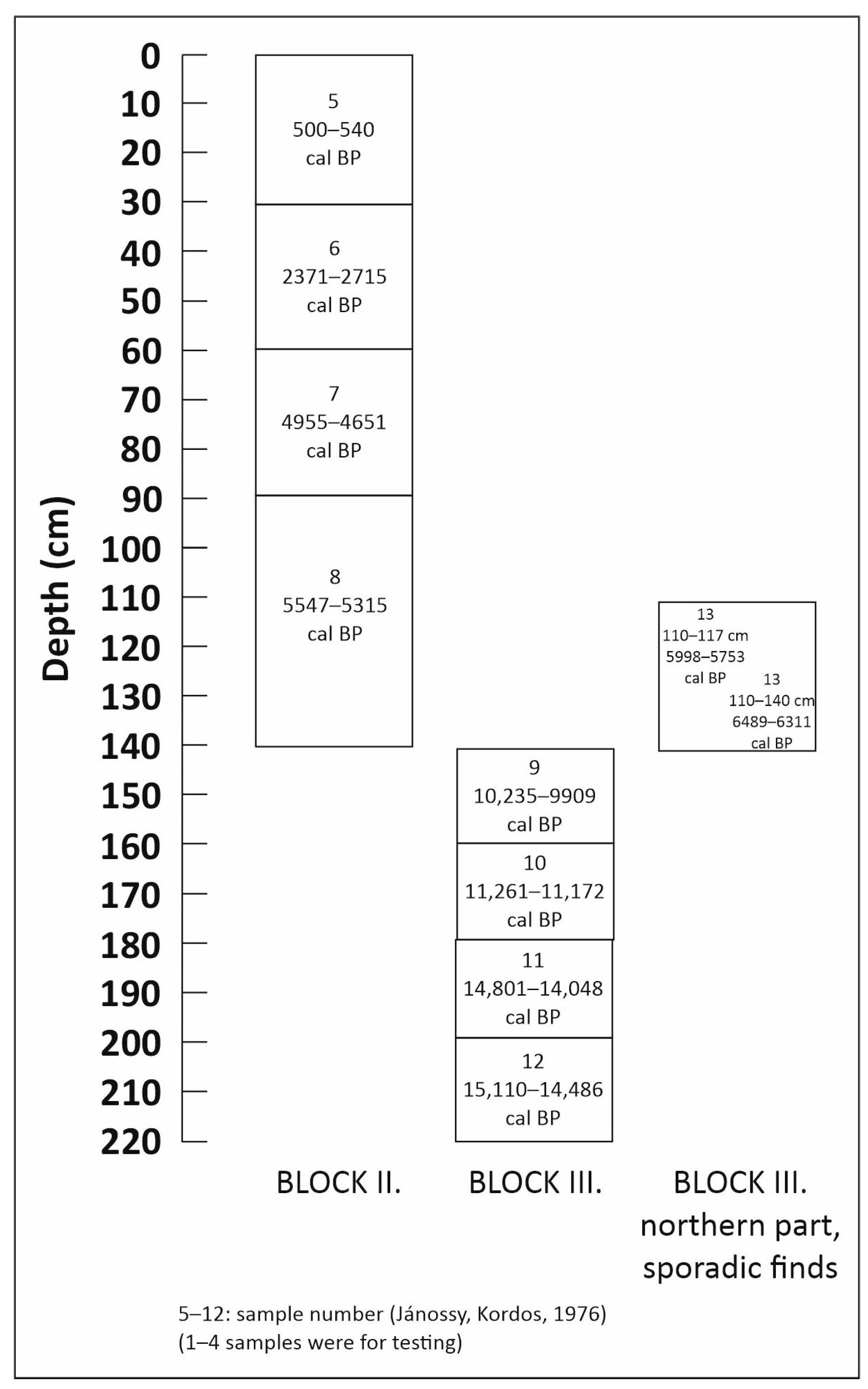
3.2. Radiocarbon Analysis and Archaeological Finds
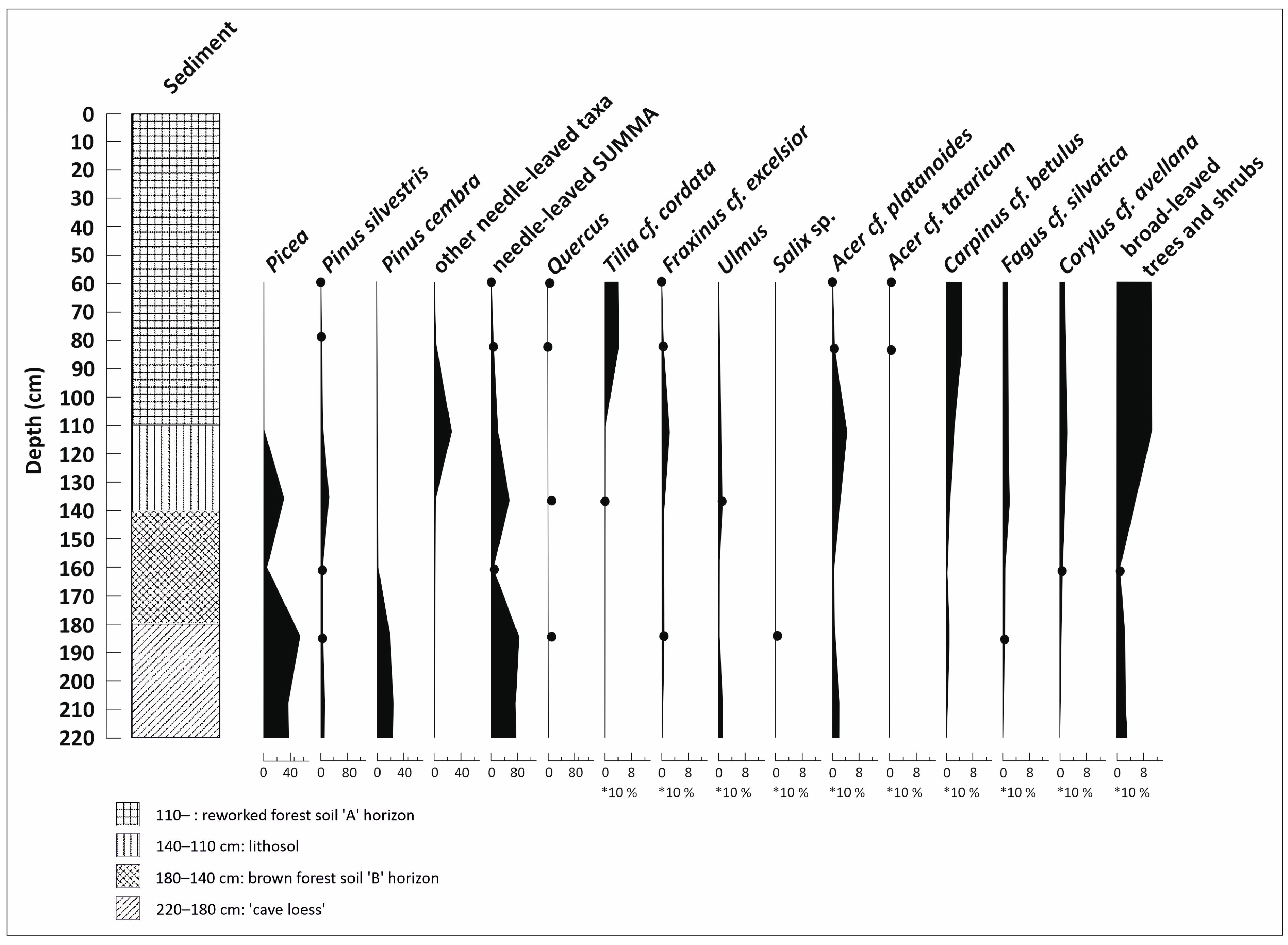
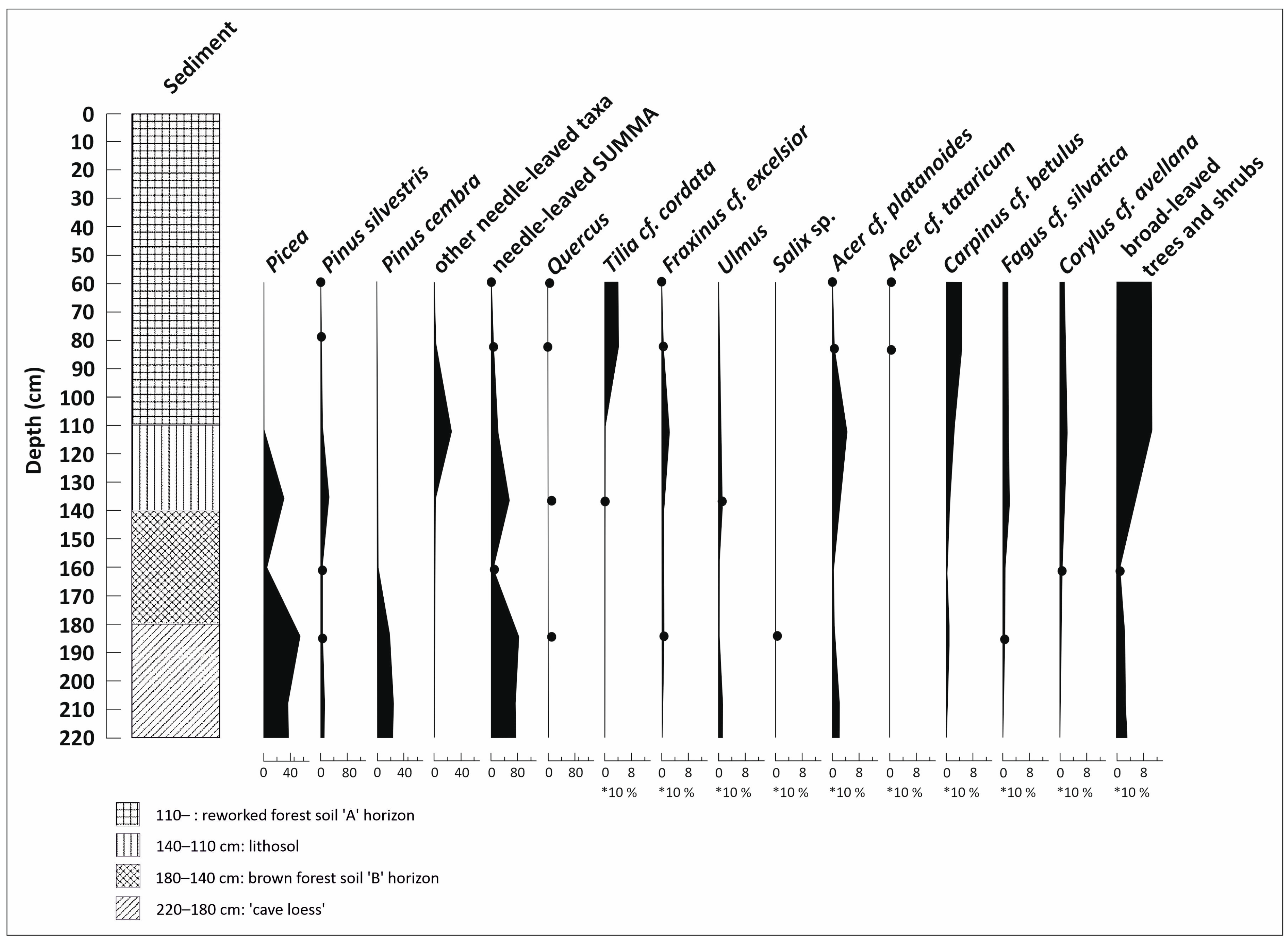
3.3. Paleobotanical Analysis
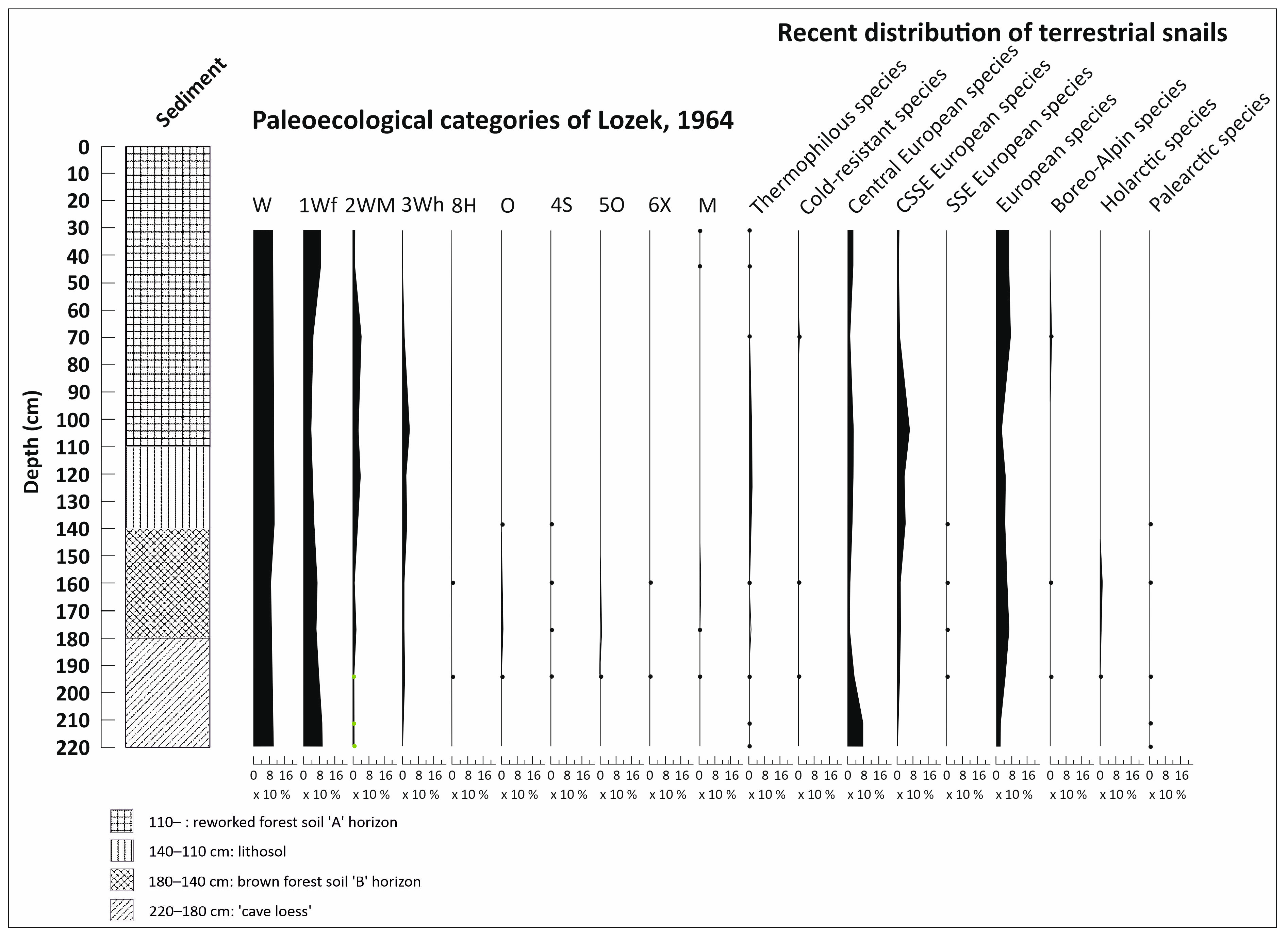
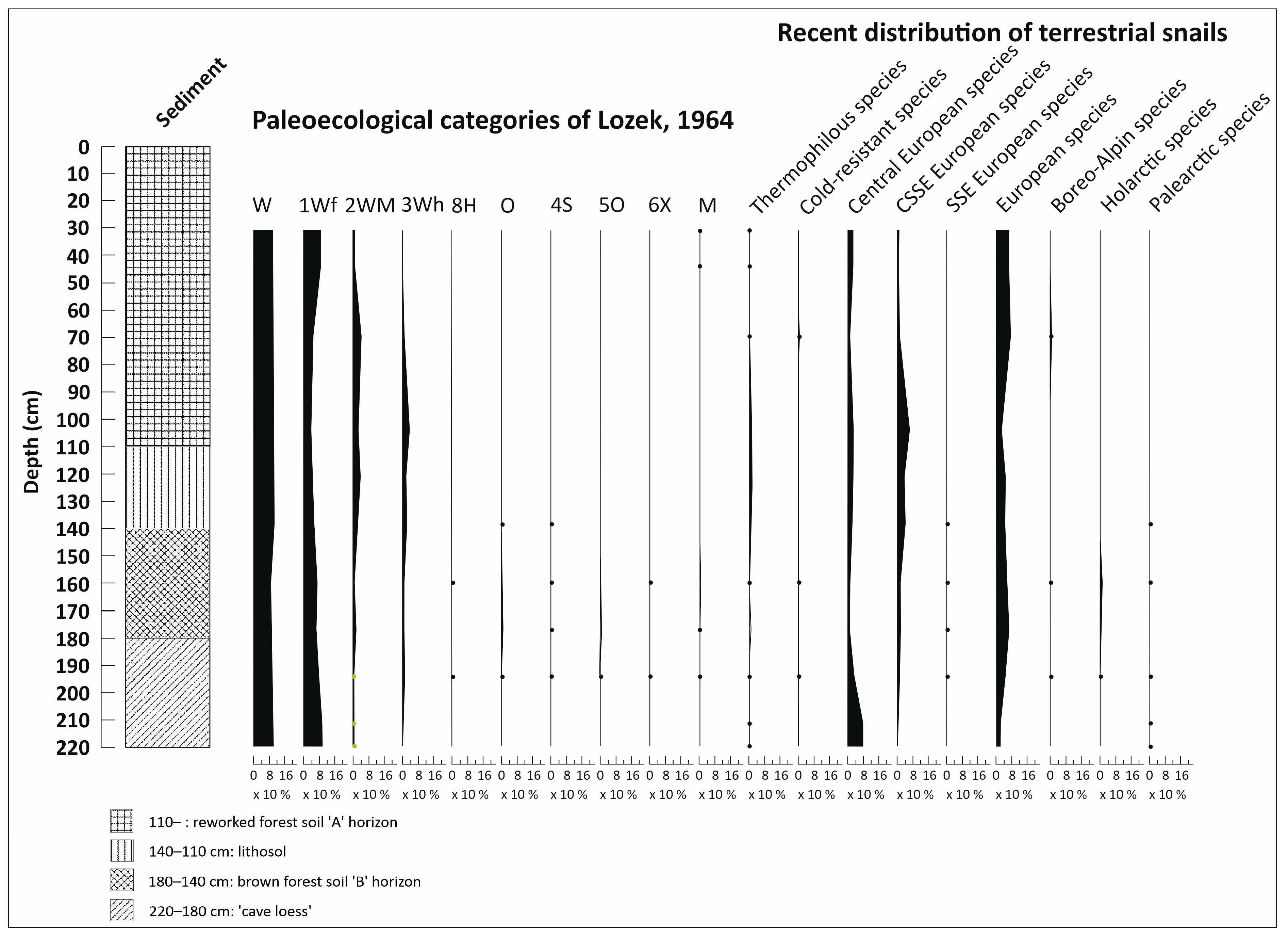
3.4. Malacological Analysis
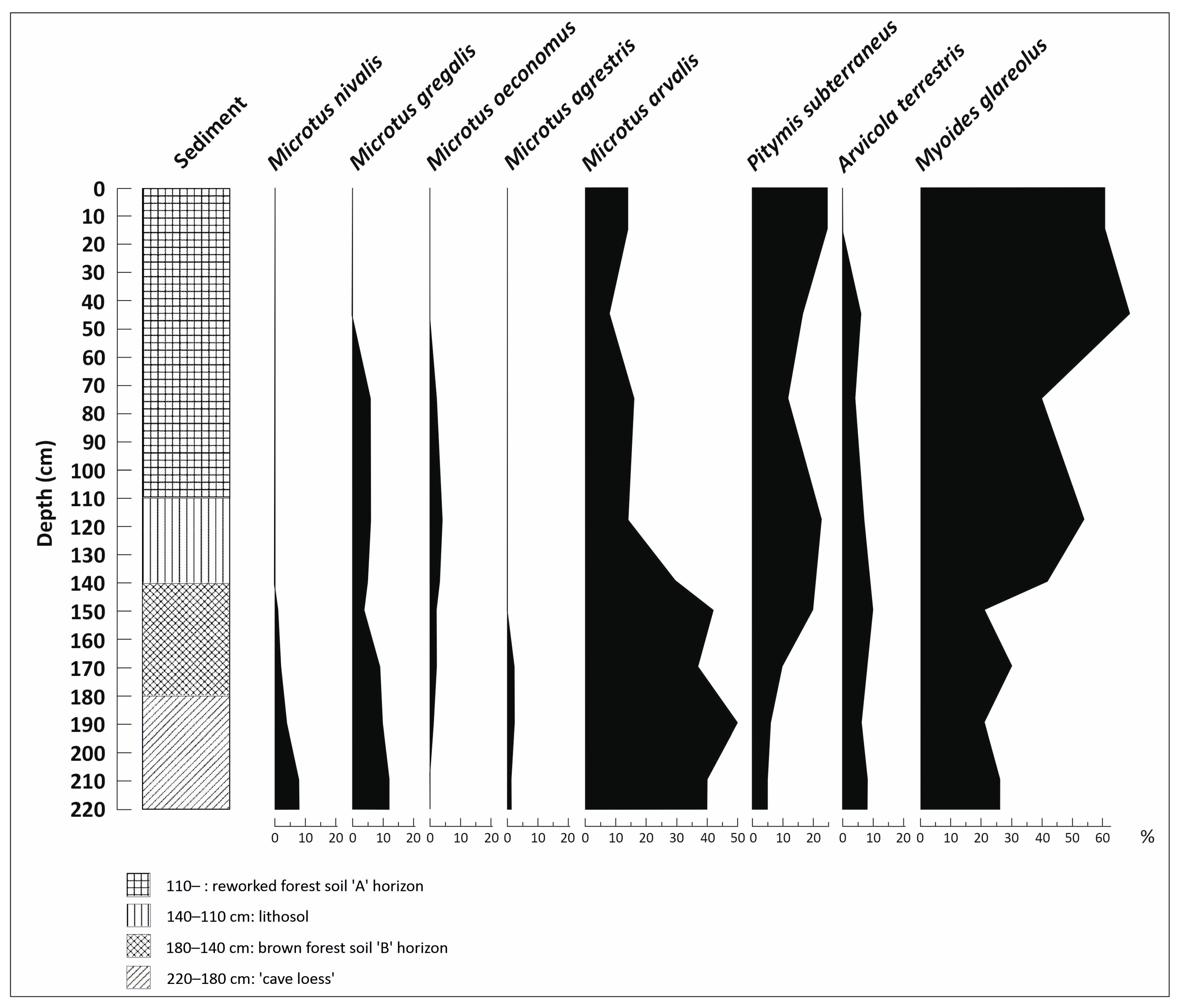
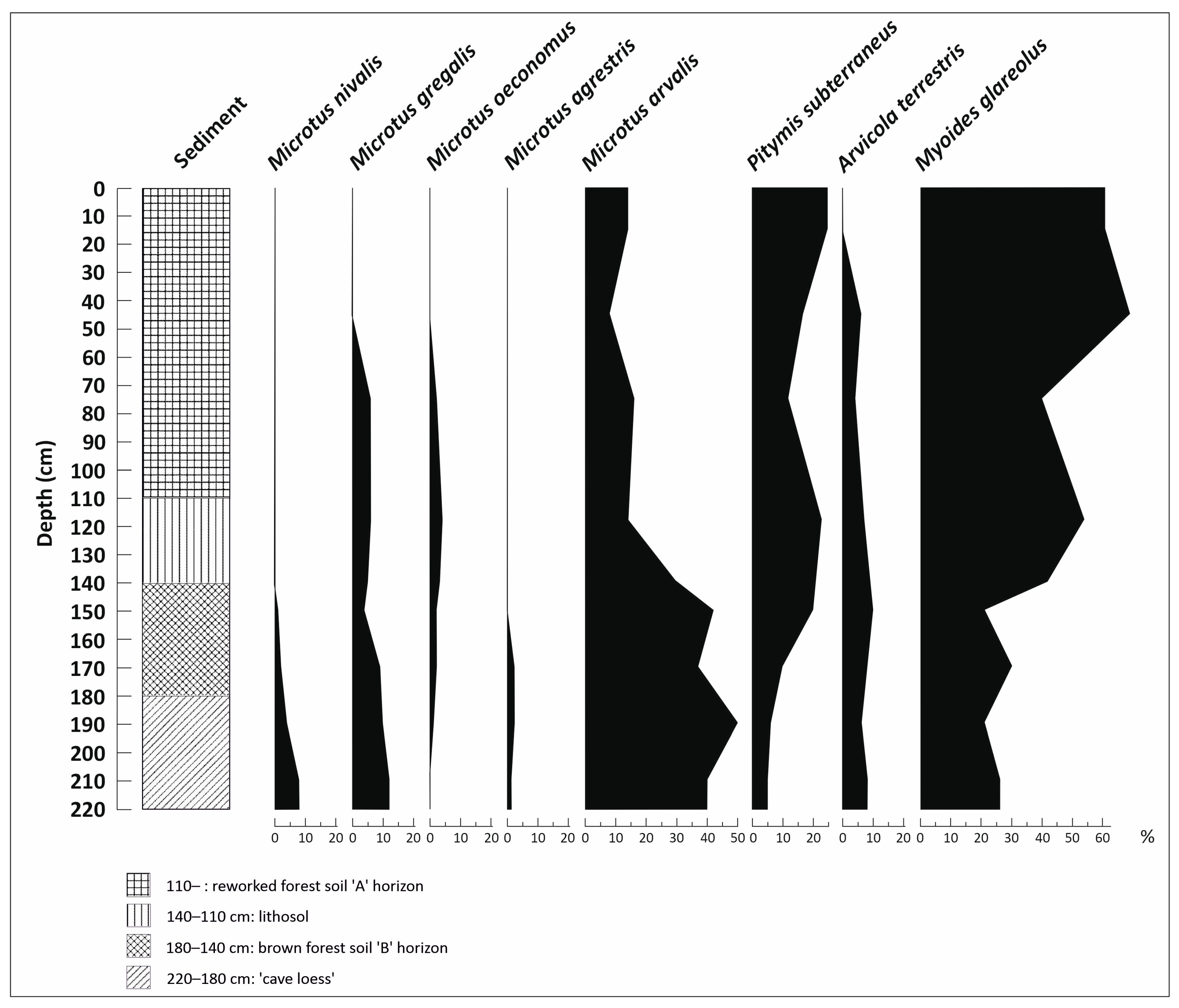
3.5. Vertebrate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- De Lattin, G Beiträge zur Zoogeographie des Mittelmeergebietes. Verhandlungen Der Dtsch. Zool. Ges. Kiel 1949, 13, 143–150.
- Stieber, J. Anthrakotonia, Kvarther Kronológia és a Hazai Pleisztocén Vegetáció. Ph.D. Thesis, Eötvös Loránd University, Budapest, Hungary, 1968. [Google Scholar]
- Jánossy, D. Vorlaufige Ergebnisse der Ausgrabungen in der Felsnische Rejtek 1. (Bükk- Gebirge, Gem. Répáshuta). Karszt És Barlangkutatás 1962, 3, 49–58. [Google Scholar]
- Stieber, J. A Hazai Felső-Pleisztocénből Származó Faszénmaradványok Antrakotómiai Vizsgálata. Candidate Thesis, Budapest, Hungary, 1957. (In Hungarian). [Google Scholar]
- Stieber, J. A magyarországi felső-pleisztocén vegetációtörténete az anthrakotómiai eredmények (1957-ig) tükrében. Földtani Közlöny 1967, 97, 308–316, (In Hungarian with German Summary). [Google Scholar]
- Stieber, J. A hazai későglaciális vegetációtörténet anthrakotómiai vizsgálatok alapján. Földtani Közlöny 1969, 99, 188–193, (In Hungarian with German summary). [Google Scholar]
- Willis, K.J.; Sümegi, P.; Braun, M.; Tóth, A. The Late Quaternary environmental history of Bátorliget, N.E. Hungary. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 118, 25–47. [Google Scholar] [CrossRef]
- Willis, K.J.; Braun, M.; Sümegi, P.; Tóth, A. Does soil change cause vegetation change or vice-versa? A temporal perpective from Hungary. Ecology 1997, 78, 740–750. [Google Scholar] [CrossRef]
- Willis, K.J.; Rudner, E.; Sümegi, P. The full-glacial forests of central and southeastern Europe: Evidence from Hungarian palaeoecological records. Quat. Res. 2000, 53, 203–213. [Google Scholar] [CrossRef]
- Birks, H.H. The Late-Quaternary history of arctic and alpine plants. Plant Ecol. Divers. 2008, 1, 135–146. [Google Scholar] [CrossRef]
- Vértes, L. Az Őskőkor és az Átmeneti Kőkor Emlékei Magyarországon; Akadémiai Kiadó: Budapest, Hungary, 1965. [Google Scholar]
- Jánossy, D. Pleistocene Vertebrate Faunas of Hungary; Akadémiai Kiadó: Budapest, Hungary, 1979. (In Hungarian) [Google Scholar]
- Jánossy, D.; Kordos, L. Pleistocene-Holocene Mollusc and Vertebrate Fauna of Two Caves in Hungary. Ann. Musei Hist. Nat. Hung. 1976, 68, 5–29. [Google Scholar]
- Pazonyi, P. A Kárpát-Medence Kvarter Emlősfauna Közösségeinek Paleoökológiai és Rétegtani Vizsgálata. Ph.D. Thesis, ELTE Őslénytani Tanszék, Budapest, Hungary, 2006. [Google Scholar]
- Jánossy, D. Pleistocene Vertebrate Faunas of Hungary; Elsevier: Amsterdam, The Netherlands, 2011; 205p. [Google Scholar]
- Troels-Smith, J. Karakterisering af lose jordater. Dan. Geol. Unders. 1955, 4, 10. [Google Scholar]
- Munsell Color Company. Munsell Soil Color Charts; Macbeth, Division of Kollmorgen Corporation, Munsell Color: New York, NY, USA, 1992. [Google Scholar]
- Njalsson, T.; Novosselov, I. Design and optimization of a compact low-cost optical particle sizer. J. Aerosol Sci. 2018, 119, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.W.; Qiang, X.K.; Fu, C.F.; Zhao, H.; Chen, T.; Sun, Y.F. Characteristics of frequency-dependent magnetic susceptibility in Bartington MS2 and Kappabridge MFK1-FA, and its application in loess-paleosol, red clay and lacustrine sediments. Chin. J. Geophys. 2012, 55, 197–206. [Google Scholar]
- Dean, W.E., Jr. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: Comparison with other methods. J. Sediment. Res. 1974, 44, 242–248. [Google Scholar]
- Magyari, E.K.; Gasparik, M.; Major, I.; Lengyel, G.; Pál, I.; Virág, A.; Korponai, J.; Haliuc, A.; Szabó, Z.; Pazonyi, P. Mammal extinction facilitated biome shift and human population change during the last glacial termination in East-Central Europe. Sci. Rep. 2022, 12, 6796. [Google Scholar] [CrossRef]
- Sümegi, P.; Persaits, G.; Gulyás, S. Woodland-Grassland Ecotonal Shifts in Environmental Mosaics: Lessons Learnt from the Environmental History of the Carpathian Basin (Central Europe) during the Holocene and the Last Ice Age Based on Investigation of Paleobotanical and Mollusk Remains. In Ecotones Between Forest and Grassland; Myster, R., Ed.; Springer: New York, NY, USA, 2012; pp. 15–57. [Google Scholar]
- Sümegi, P.; Náfrádi, K. A radiocarbon, dated cave sequence and the Pleistocene/Holocene transition in Hungary. Open Geosci. 2015, 7, 783–798. [Google Scholar] [CrossRef]
- Gaál, I. A Gerecse-hegység egyik legérdekesebb barlangcsoportja. Földgömb 1934, 5, 321–330. [Google Scholar]
- Mozsolics, A.A. Goldfunde des Depotfundhorizontes von Hajdúsámson. In Bericht der Römisch–Germanischen Kommission; J. Baer & Company: Berlin, Germany, 1968; Volume 46–47, pp. 2–76. [Google Scholar]
- Rudner, E.; Sümegi, P. Recurring taiga forest steppe habitats in the Carpathian Basin in the Upper Weichselian. Quat. Int. 2001, 76/77, 177–189. [Google Scholar] [CrossRef]
- Rudner, E.; Sümegi, P. Charcoal as a remain of natural and human-set fires of Palaeolithic Times—Case study from Hungary. Br. Archaeol. Rep. 2002, 1089, 11–18. [Google Scholar]
- Welter-Schultes, F. European Non-Marine Mollusc, a Guide for Species Identification; Planet Poster Editions: Göttingen, Germany, 2012; 680p. [Google Scholar]
- Lozek, V. Quartärmollusken der Tschechoslowakei—Rozpravy Ustredniho Ústavu Geologického; Ústřední Ústav Geologický: Prague, Czech Republic, 1964; p. 31. [Google Scholar]
- Vörös, I. Large mammalian faunal changes during the late Upper Pleistocene and early Holocene times in the Carpathian Basin. In Pleistocene Environment in Hungary; Pécsi, M., Ed.; Geographical Research Institute of HAS: Budapest, Hungary, 1987; pp. 81–101. [Google Scholar]
- Wilson, K.R. The last glaciation in the western Mourne Mountains, Northern Ireland. Scott. Geogr. J. 2004, 120, 199–210. [Google Scholar] [CrossRef]
- Sissons, J.B. The Loch Lomond Stadial in the British Isles. Nature 1979, 280, 199–203. [Google Scholar] [CrossRef]
- Sümegi, P. The results of paleoenvironmental reconstruction and comparative geoarcheological analysis for the examined area. In The Geohistory of Bátorliget Marshland; Sümegi, P., Gulyás, S., Eds.; Archaeolingua Press: Budapest, Hungary, 2004; pp. 301–348. [Google Scholar]
- Sümegi, P. Reconstruction of flora, soil and landscape evolution, and human impact on the Bereg Plain from late-glacial up to the present, based on palaeoecological analysis. In The Upper Tisa Valley; Hamar, J., Sárkány Kiss, A., Eds.; Tiscia Monograph Series; Tisza Klub & Liga Pro Europa: Szeged, Hungary, 1999; pp. 173–204. [Google Scholar]
- Szabó, J. Csuszamlásos Folyamatok Szerepe a Magyarországi Tájak Geomorfológiai Fejlődésében. Habilitation Thesis, Kossuth Egyetemi Kiadó, Debrecen, Hungary, 1996. (In Hungarian). [Google Scholar]
- Stieber, J. Anthrakotomische Untersuchung. Folia Archaeol. 1956, 8, 13–14, (In Hungarian with German and French Summaries). [Google Scholar]
- Hollendonner, F. A magyarországi praehistorikus fák és faszenek mikroszkópikus vizsgálata. Math. És Természettudományi Értesítő 1926, 42, 204–207. [Google Scholar]
- Hollendonner, F. A fosszilis faszenek vizsgálata. In A cserépfalui Mussolini barlang (Subalyuk). Geol. Hung. Ser. Palaeonthologica 1938, 14, 311–315. (In Hungarian) [Google Scholar]
- Greguss, P. Az első alföldi paleolit-lelet faszén maradványai. Dolg. A Magy. Királyi Ferenc József Tudegy. Archeol. Intézetéből 1936, 12, 14–19. [Google Scholar]
- Sárkány, S.; Stieber, J. A Szelim-barlang újabb faszénmaradványainak anthrakotómiai vizsgálata. In Budapesti Tudományegyetem Biológiai Intézetének Évkönyve; Budapesti Tudományegyetem Biológiai Intézetének: Budapest, Hungary, 1950; pp. 32–37. [Google Scholar]
- Miháltz, I. A Duna-Tisza köze déli részének földtani felvétele. In A Magyar Állami Földtani Intézet Évi jelentése 1950-ről; Müszaki Könyvkiadó: Budapest, Hungary, 1953; pp. 113–143. Available online: https://epa.oszk.hu/02900/02934/00118/pdf/EPA02934_mafi_evi_jel_1950_113-143.pdf. (accessed on 30 September 2023).
- Csinády, G. A bátorligeti láp története a pollenanalízis tükrében. (The evolutionary history of Bátorliget marshland from a pollen analytical point of view). Földrajzi Értesítő 1953, 3, 684–694. (In Hungarian) [Google Scholar]
- O’Connor, T.P.; Evans, J.G. Environmental Archaeology: Principles and Methods; Sutton Publishing: London, UK, 1999; 242p. [Google Scholar]
- Gardner, A.R. Biotic response to Early Holocene human activity: Results from palaeoenvironmental analyses of sediments from Podpesko Jezero. Dok. Prehist. 1998, 24, 63–77. [Google Scholar]
- Gardner, A.R. The ecology of Neolithic environmental impacts–re-evaluation of existing theory using case studies from Hungary. Dok. Prehist. 1999, 26, 163–183. [Google Scholar]
- Gardner, A.R.; Willis, K.J. Prehistoric farming and the postglacial expansion of beech and hornbeam: A comment on Küster. Holocene 1999, 9, 119–122. [Google Scholar] [CrossRef]
- Magyari, E. A bükk (Fagus silvatica) és a gyertyán (Carpinus betulus) terjedésének problematikája a Kárpát-medence holocén vegetációfejlődésében. A 4. Magyar Őslénytani Vándorgyűlés előadásainak kivonatai. Pécsvárad. 2001. Available online: https://foldtan.hu/sites/default/files/3071150_2001_pecsvarad_fuzet.pdf. (accessed on 30 September 2023).
- Juhász, I.E. A Délnyugat Dunántúl Negyedkori Vegetációtörténetének Palinológiai Rekonstrukciója. (Reconstitution Palynologique de la Végétation Depuis le Tardiglaciaire Dans la Région de Zala, Sud-Ouest de la Hongrie). Ph.D. Thesis, University Aix Marseille III., Marseille, France, 2002. [Google Scholar]
- Magyari, E. Climatic Versus Human Modification of the Late Quaternary Vegetation in Eastern Hungary. Ph.D. Thesis, Department of Mineralogy and Geology, University of Debrecen, Debrecen, Hungary, 2002. [Google Scholar]
- Magri, D.; Vendramin, G.G.; Comps, B.; Dupanloup, I.; Geburek, T.; Gömöry, D.; Latałowa, M.; Litt, T.; Paule, L.; Roure, J.M.; et al. A new scenario for the Quaternary history of European beech populations: Palaeobotanical evidence and genetic consequences. New Phytol. 2006, 171, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Fűköh, L. A magyarországi középhegységi holocén Mollusca-fauna zoogeográfiai vizsgálata. Malacol. Newsl. 1994, 13, 9–43. [Google Scholar]
- Fűköh, L. Biostratigraphical and geomorphological investigation in areas of Hungarian subsided during the Holocene. Z. Geomorphol. Suppl. 1997, 110, 45–56. [Google Scholar]
- Medzihradszky, Z.; Gasparik, M.; Fűköh, L. A Vallonia costata biozóna (Fűköh, L. 1990) sztratotípus lelőhely újravizsgálata. Folia Hist. Nat. Musei Matra. 2002, 26, 37–46. [Google Scholar]
- Alexandrowicz, W.P. Molluscan assemblages of Late Glacial and Holocene calcareous tufas in Southern Poland. Folia Quat. 2004, 75, 309. [Google Scholar]
- Alexandrowicz, W.P. Malacological evidence of the natural and anthropogenic changes of the environment in the eastern part of the Carpathian Foreland: The studies in the Glinne stream valley near Rzeszów (southern Poland). Carpathian J. Earth Environ. Sci. 2019, 14, 367–384. [Google Scholar] [CrossRef]
- Alexandrowicz, W.P.; Szymanek, M.; Rybska, E. Molluscan assemblages from Holocene calcareous tufa and their significance for palaeoenvironmental reconstructions. A study in the Pieniny Mountains (Carpathians, southern Poland). Carpathian J. Earth Environ. Sci. 2016, 11, 37–54. [Google Scholar]
- Sümegi, P.; Deli, T. Results of the quartermalacological analysis of the profiles from the central and marginal areas of Bátorliget marshland. In The Geohistory of Bátorliget Marshland; Sümegi, P., Gulyás, S., Eds.; Archaeolingua Press: Budapest, Hungary, 2004; pp. 183–207. [Google Scholar]
- Preece, R.C. Mapping snails in time: The prospect of elucidating the historical biogeography of the European malacofauna. In Proceedings of the Tenth International Malacological Congress (Tübingen 1989); Meier Brook, C., Ed.; Unitas Malacologica: Sidney, Australia, 1991; pp. 477–479. [Google Scholar]
- Preece, R.C.; Day, S.P. Comparison of the molluscan and vegetational successions from a radiocarbon-dated tufa in Oxfordshire. J. Biogeogr. 1994, 21, 463–478. [Google Scholar] [CrossRef]
- Pigati, J.S.; Quade, J.; Shahanan, T.M.; Haynes, C.V., Jr. Radiocarbon dating of minute gastropods and new constraints on the timing of late Quaternary spring-discharge deposits in southern Arizona, USA. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 204, 33–45. [Google Scholar] [CrossRef]
- Pigati, J.S.; Rech, J.A.; Nekola, J.C. Radiocarbon dating of small terrestrial gastropod shells in North America. Quat. Geochronol. 2010, 5, 519–532. [Google Scholar] [CrossRef]
- Pigati, J.S.; McGeehin, J.P.; Muhs, D.R.; Bettis III, E.A. Radiocarbon dating late Quaternary loess deposits using small terrestrial gastropod shells. Quat. Sci. Rev. 2013, 76, 114–128. [Google Scholar] [CrossRef]
- Újvári, G.; Molnár, M.; Novothny, Á.; Páll-Gergely, B.; Kovács, J.; Várhegyi, A. AMS 14C and OSL/IRSL dating of the Dunaszekcső loess sequence (Hungary): Chronology for 20 to 150 ka and implications for establishing reliable age–depth models for the last 40 ka. Quat. Sci. Rev. 2014, 106, 140–154. [Google Scholar] [CrossRef]
- Roberts, N. The Holocene. An Environmental History; Blackwell Press: Oxford, UK, 1998; 384p. [Google Scholar]
- Nyilas, I.; Sümegi, P. Pointing out Cochlicopa nitens (Gallenstein, 1848) a new species for the Pleistocene in Hungary, in the territory of the Hortobágy National Park. Soosiana 1989, 17, 113–115. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BLOCK II. | BLOCK III. | BLOCK III. Northern Part, Sporadic Finds | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Depth (cm) | Original Sample Number 1957 | Renumbered 1962 | Renumbered 1976 | Original Sample Number 1957 | Renumbered 1962 | Renumbered 1976 | Original Sample Number 1957 | Renumbered 1962 | Renumbered 1976 |
| 0–30 | II/1 | 1 | 5 | ||||||
| 30–60 | II/2 | 6 | |||||||
| 60–90 | II/3 | 7 | |||||||
| 90–120 (140) | II/4 “Neolithic” | 2 | 8 | block III. northern part at 1.10 m/110–140 cm | 3 (110–140 cm) | 13 (at 110 cm) | |||
| direct continuity of layers between Blocks II. and III. is lacking [13] | |||||||||
| 140–160 | BLOCK III. 140–160 | 4 | 9 | ||||||
| 160–180 | BLOCK III. 160–180 | 5 | 10 | ||||||
| 180–200 | BLOCK III. 180–200 | 6 | 11 | ||||||
| 200–220 | BLOCK III. 200–220 | 7 | 12 | ||||||
| Depth (cm) | Uncal BP | +/− | Cal BP | +/− | Range Cal BP | Cal BC/AD | +/− | Range Cal BC | Lab Code |
|---|---|---|---|---|---|---|---|---|---|
| 0–30 | 480 | 25 | 520 | 20 | 500–540 | 1431 AD | 20 | 1411–1451 AD | Poz-7881 |
| 30–60 | 2470 | 30 | 2887 | 172 | 3059–2715 | 593 BC | 172 | 765–421 BC | Poz-7880 |
| 60–90 | 4265 | 35 | 4803 | 152 | 4955–4651 | 2804 BC | 152 | 2956–2652 BC | Poz-7887 |
| 90–120 | 4675 | 35 | 5431 | 116 | 5547–5315 | 3714 BC | 116 | 3830–3598 BC | Poz-7877 |
| 110–117 | 5160 | 40 | 5876 | 123 | 5999–5753 | 3927 BC | 123 | 4050–3804 BC | Poz-7879 |
| 110–140 | 5630 | 40 | 6400 | 89 | 6489–6311 | 4451 BC | 89 | 4540–4362 BC | Poz-7872 |
| 140–160 | 8970 | 50 | 10,072 | 163 | 10,235–9909 | 8019 BC | 159 | 8169–7869 BC | Poz-7876 |
| 160–180 | 9790 | 40 | 11,217 | 45 | 11,262–11,172 | 9268 BC | 45 | 9313–9223 BC | Poz-7878 |
| 180–200 | 12,260 | 50 | 14,425 | 377 | 14,802–14,048 | 12,476 BC | 377 | 12,853–12,099 BC | Poz-7963 |
| 200–220 | 12,530 | 50 | 14,812 | 326 | 15,138–14,486 | 12,835 BC | 326 | 13,161–12,509 BC | Poz-7975 |
| Depth (cm) | Cal BP (Range) | Cal BC/AD (Range) | Archaeological Finds [11,12,15] |
|---|---|---|---|
| 0–30 | 500–540 | 1411–1451 AD | coin from 18th century |
| hiatus | |||
| 30–60 | 3059–2715 | 765–421 BC | - |
| hiatus | |||
| 60–90 | 4955–4651 | 2956–2652 BC | Prehistoric (Neolithic-Bronze Age) ceramics pieces |
| 90–120 | 5547–5315 | 3830–3598 BC | Prehistoric (Neolithic-Bronze Age) ceramics pieces |
| 110–117 (sample 13) | 5999–5753 | 4050–3804 BC | Prehistoric (Neolithic-Bronze Age) ceramics pieces |
| 110–140 (sample 13) | 6489–6311 | 4540–4362 BC | Prehistoric (Neolithic-Bronze Age) ceramics pieces |
| hiatus | |||
| 140–160 | 10,235–9909 | 8169–7869 BC | Mezolithic (Tardonasien) trapezoid blade |
| 160–180 | 11,262–11,172 | 9313–9223 BC | |
| hiatus | |||
| 180–200 | 14,802–14,048 | 12,853–12,099 BC | Epipaleolit (?) finds |
| 220–200 | 15,138–14,486 | 13,161–12,509 BC | Epipaleolit (?) finds |
| Sample Number [13] | 12 | 11 | 10 | 9 | 13 | 8 |
|---|---|---|---|---|---|---|
| cal BP years | 15,138–14,486 | 14,802–14,048 | 11,262–11,172 | 10,235–9909 | 6489–6311 | 5547–5315 |
| Depth (cm) | 220–200 cm | 200–180 cm | 180–160 cm | 160–140 cm | 140–110 cm | 90–120 cm |
| Charcoal taxa | number of pieces | number of pieces | number of pieces | number of pieces | number of pieces | number of pieces |
| Picea (original Picea-Larix) | 5 | 26 | 1 | 30 | ||
| Pinus sylvestris | 1 | 1 | 1 | 16 | 1 | |
| Pinus cembra | 2 | 5 | ||||
| Needle-leaf tree | 6 | 32 | 2 | 46 | 1 | |
| Quercus sp. | 1 | 4 | 2 | |||
| Tilia cf. cordata | 2 | 25 | ||||
| Fraxinus cf. excelsior | 1 | 2 | 4 | |||
| Ulmus cf. campestre | 1 | 4 | ||||
| Salix sp. | 1 | |||||
| Acer cf. platanoides | 2 | 2 | 15 | 5 | 4 | |
| Acer cf. tataricum | 2 | |||||
| Carpinus cf. betulus | 2 | 4 | 2 | 31 | ||
| Fagus cf. sylvatica | 1 | 11 | 1 | 8 | ||
| Corylus cf. avellana | 1 | 8 | 2 | 10 | ||
| Broad leaf trees and shrub | 3 | 8 | 1 | 48 | 14 | 84 |
| SUMMA | 11 | 40 | 3 | 94 | 14 | 85 |
| Sample Number [13] | 12 | 11 | 10 | 9 | 13 | 8 | 7 | 6 | 5 |
|---|---|---|---|---|---|---|---|---|---|
| cal BP years (range) | 15,138–14,486 | 14,802–14,048 | 11,262–11,172 | 10,235–9909 | 6489–6311 | 5547–5313 | 4955–4651 | 3059–2715 | 540–500 |
| Depth (cm) | 220–200 | 200–180 | 180–160 | 160–140 | 140–110 | 120–90 | 90–60 | 60–30 | 30–0 |
| Piece (i = individual) | i | i | i | i | i | i | i | i | i |
| Acicula polita | 1 | ||||||||
| Carychium tridentatum | 1 | 7 | |||||||
| Cochlicopa lubricella | 1 | ||||||||
| Chondrina clienta | 1 | 1 | |||||||
| Granaria frumentum | 1 | 2 | 1 | ||||||
| Orcula dolium | 1 | 1 | 1 | 1 | 1 | ||||
| Sphyradium doliolum | 1 | 4 | 1 | ||||||
| Truncatellina cylindrica | 1 | ||||||||
| Vallonia pulchella | 1 | ||||||||
| Vallonia costata | 1 | 2 | 17 | ||||||
| Chondrula tridens | 2 | 1 | |||||||
| Cochlodina laminata | 4 | 3 | 7 | 1 | 1 | ||||
| Cochlodina cerata | 35 | 52 | 0 | 2 | 6 | 1 | 1 | ||
| Clausilia dubia | 2 | 2 | 1 | 2 | 12 | ||||
| Clausilia pumila | 17 | 1 | 16 | 10 | 1 | 2 | 1 | ||
| Iphigena ventricosa | 2 | ||||||||
| Iphigena plicatula | 1 | 1 | |||||||
| Laciniaria plicata | 1 | 3 | 4 | 18 | 1 | 2 | 1 | ||
| Laciniaria biplicata | 5 | 1 | |||||||
| Balea cana | 4 | ||||||||
| Ruthenica filograna | 3 | 19 | 6 | 16 | |||||
| Clausilia sp. | 10 | 75 | 27 | 150 | 37 | 2 | 1 | 9 | 56 |
| Vitrea crystallina | 1 | 6 | |||||||
| Vitrea contracta | 1 | ||||||||
| Vitrea diaphana | 1 | ||||||||
| Aegopinella minor | 1 | 5 | 5 | ||||||
| Aegopinella pura | 4 | ||||||||
| Oxychilus glaber | 8 | ||||||||
| Oxychilus orientalis | 1 | 4 | 1 | 1 | |||||
| Oxychilus depressus | 1 | ||||||||
| Nesovitrea hammonis | 3 | 1 | |||||||
| Zonitidae | 4 | 2 | 21 | ||||||
| Euconulus fulvus | 1 | ||||||||
| Daudebardia rufa | 4 | ||||||||
| Limax maximus | 11 | 2 | 1 | 8 | |||||
| Limacidae | 4 | ||||||||
| Discus ruderatus | 1 | 7 | 1 | ||||||
| Discus rotundatus | 2 | 1 | 2 | ||||||
| Discus perspectivus | 1 | ||||||||
| Bradybaena fruticum | 1 | 2 | |||||||
| Euomphalia strigella | 1 | 3 | 1 | 1 | |||||
| Trichia unidentata | 1 | ||||||||
| Perforatella incarnata | 2 | 1 | |||||||
| Isognomostoma isognomostoma | 1 | ||||||||
| Helicigona faustina | 3 | 1 | |||||||
| Helicodonta obvoluta | 1 | 2 | 1 | ||||||
| Helix pomatia | 1 | 3 | 2 | 1 | 1 | 4 | |||
| Helicidae | 2 | 4 | 5 | 1 | 4 | ||||
| SUMMA | 49 | 168 | 57 | 307 | 128 | 11 | 11 | 23 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Náfrádi, K.; Sümegi, P. The Forest Refugium of the Bükk Mountains, Hungary—Vegetation Change and Human Impact from the Late Pleistocene. Diversity 2024, 16, 109. https://doi.org/10.3390/d16020109
Náfrádi K, Sümegi P. The Forest Refugium of the Bükk Mountains, Hungary—Vegetation Change and Human Impact from the Late Pleistocene. Diversity. 2024; 16(2):109. https://doi.org/10.3390/d16020109
Chicago/Turabian StyleNáfrádi, Katalin, and Pál Sümegi. 2024. "The Forest Refugium of the Bükk Mountains, Hungary—Vegetation Change and Human Impact from the Late Pleistocene" Diversity 16, no. 2: 109. https://doi.org/10.3390/d16020109
APA StyleNáfrádi, K., & Sümegi, P. (2024). The Forest Refugium of the Bükk Mountains, Hungary—Vegetation Change and Human Impact from the Late Pleistocene. Diversity, 16(2), 109. https://doi.org/10.3390/d16020109