

Origin of Wild Polyploid Avena Species Inferred from Polymorphism of the ITS1 rDNA in Their Genomes
 ,
, 


Abstract
1. Introduction
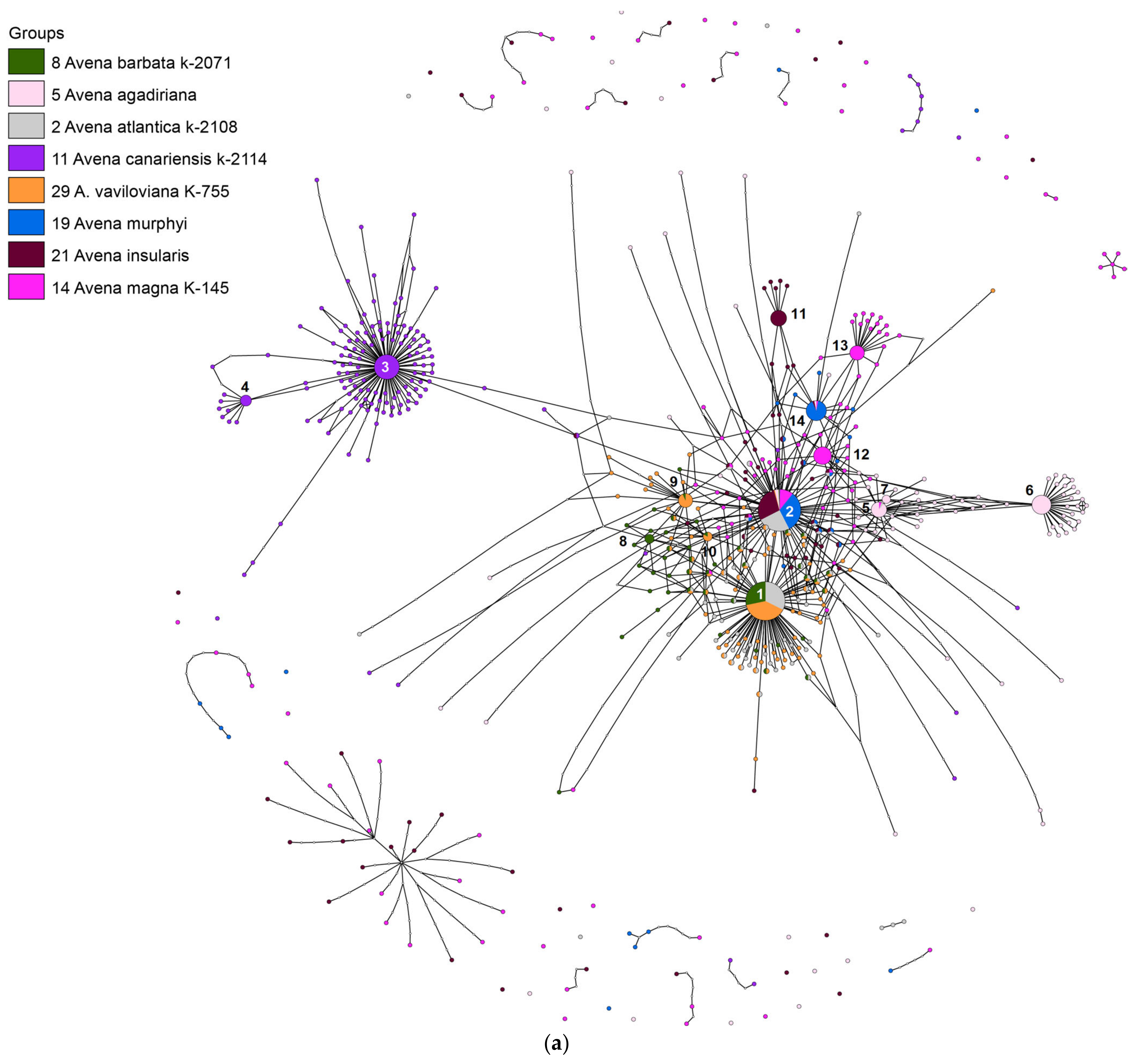
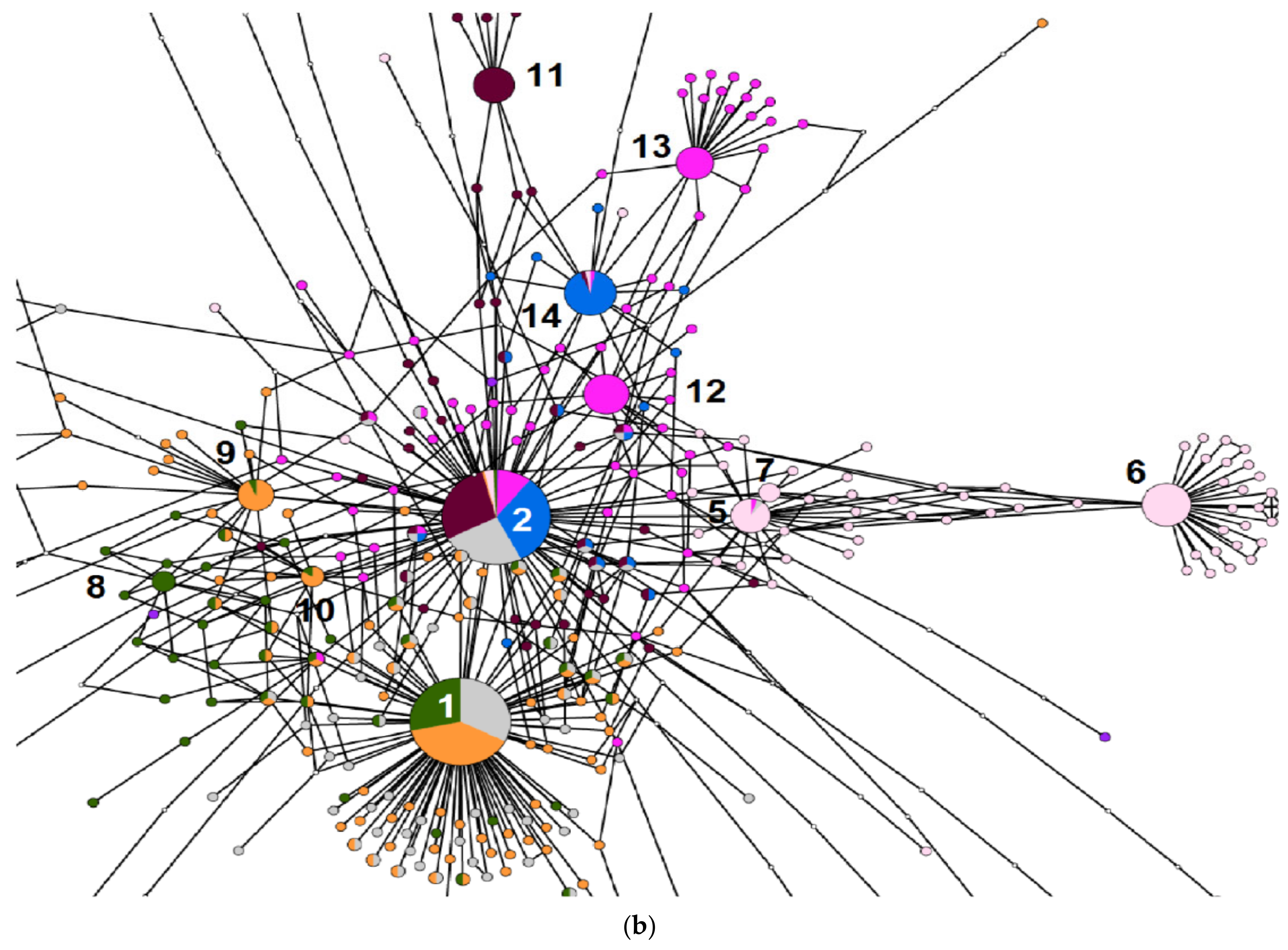
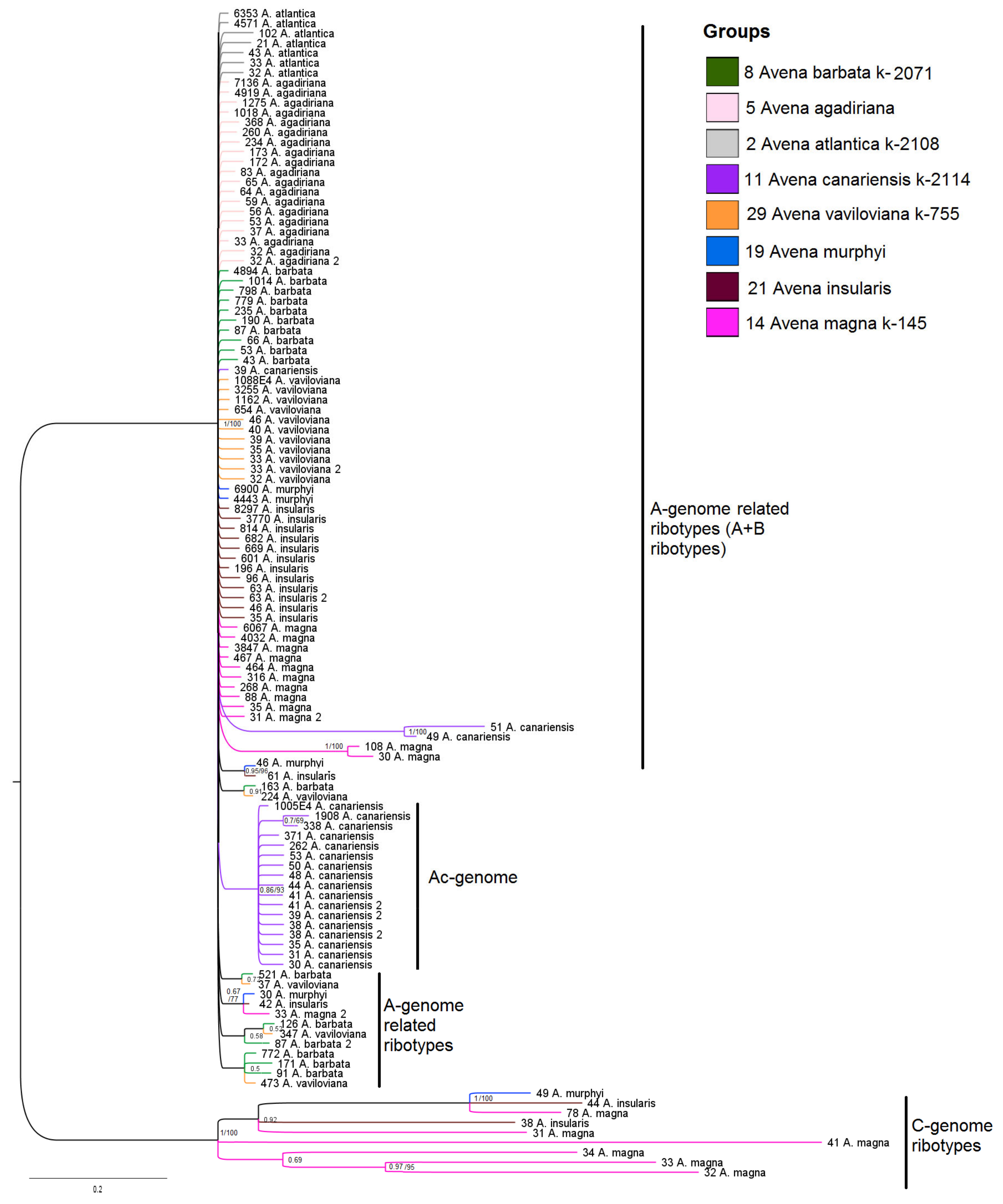
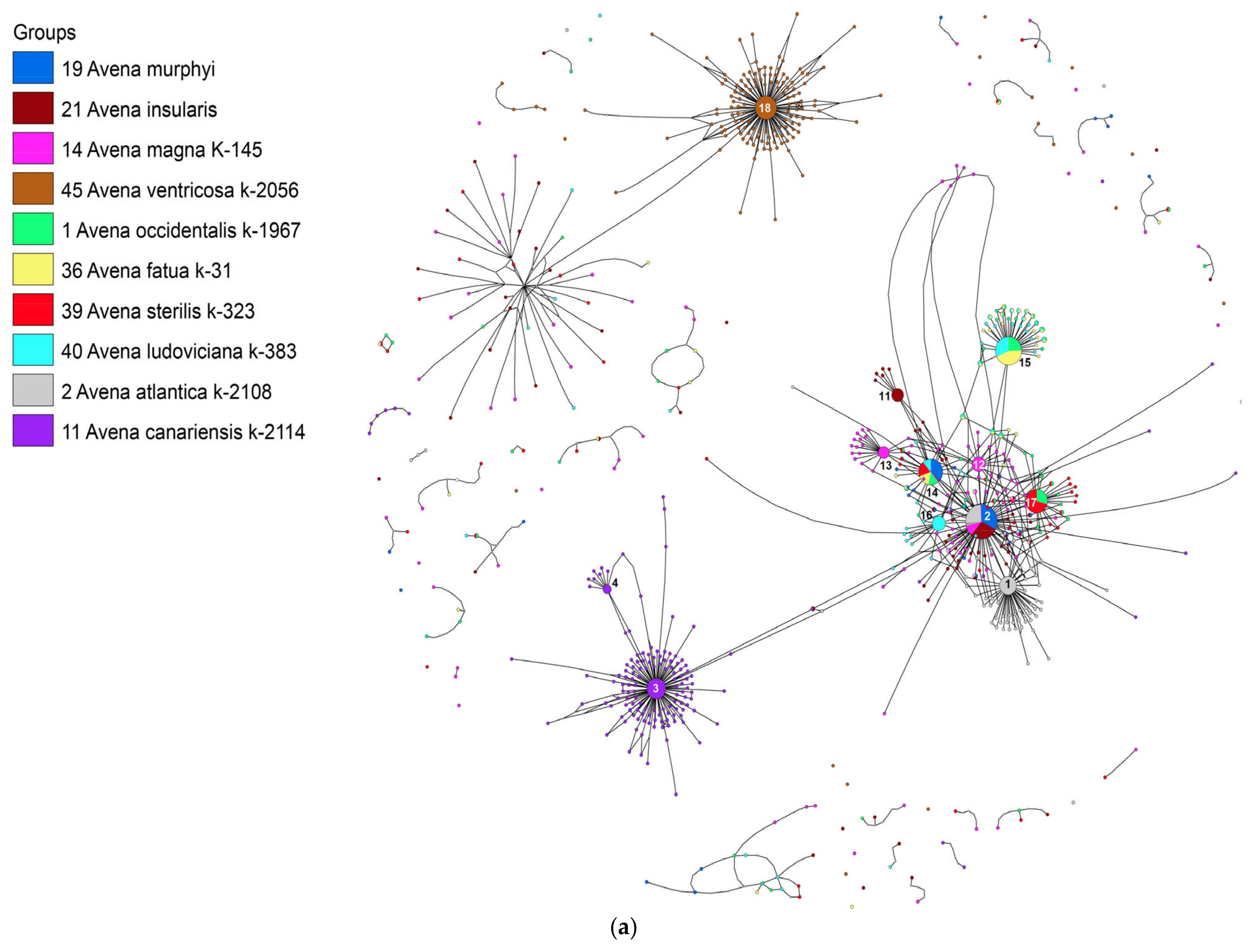
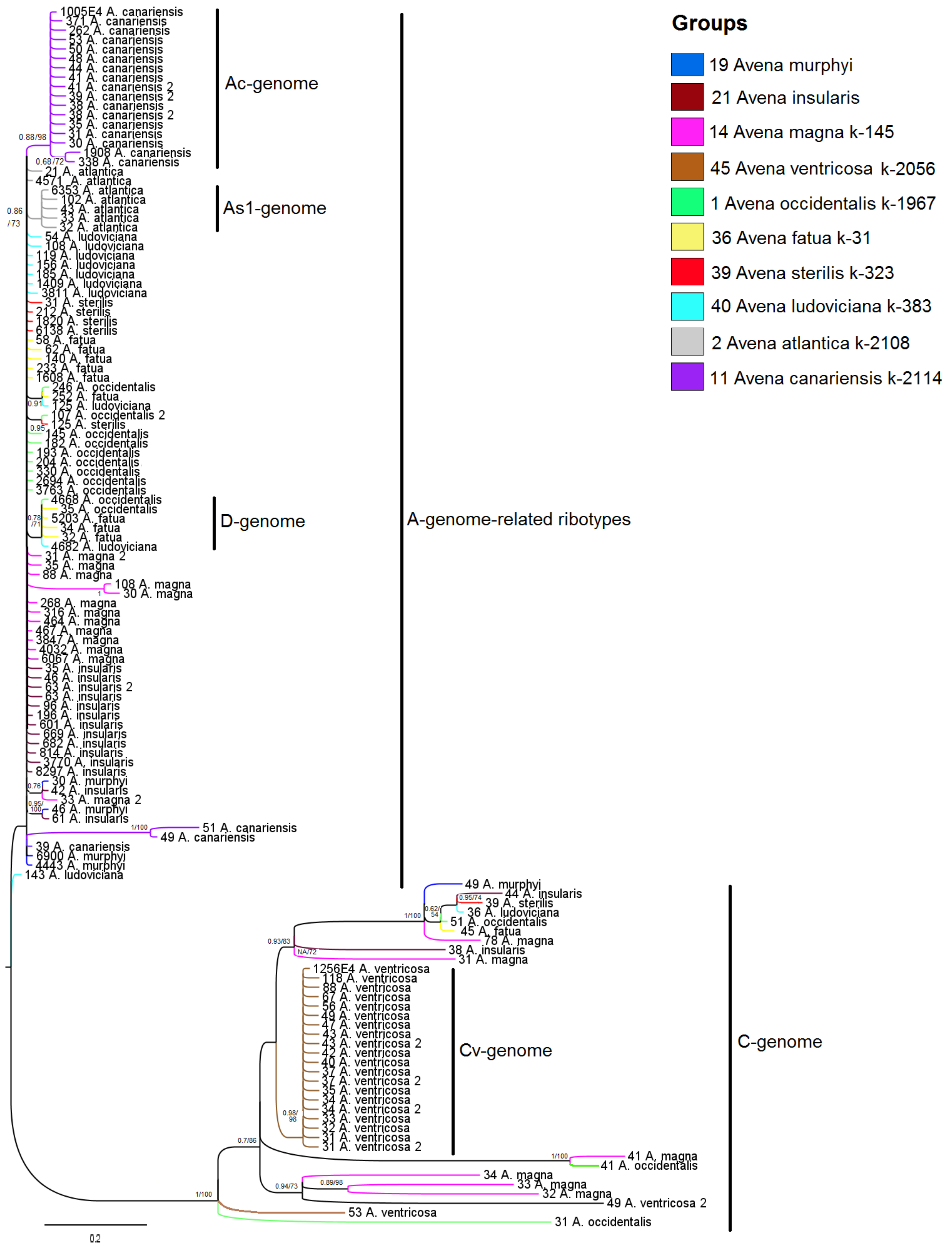
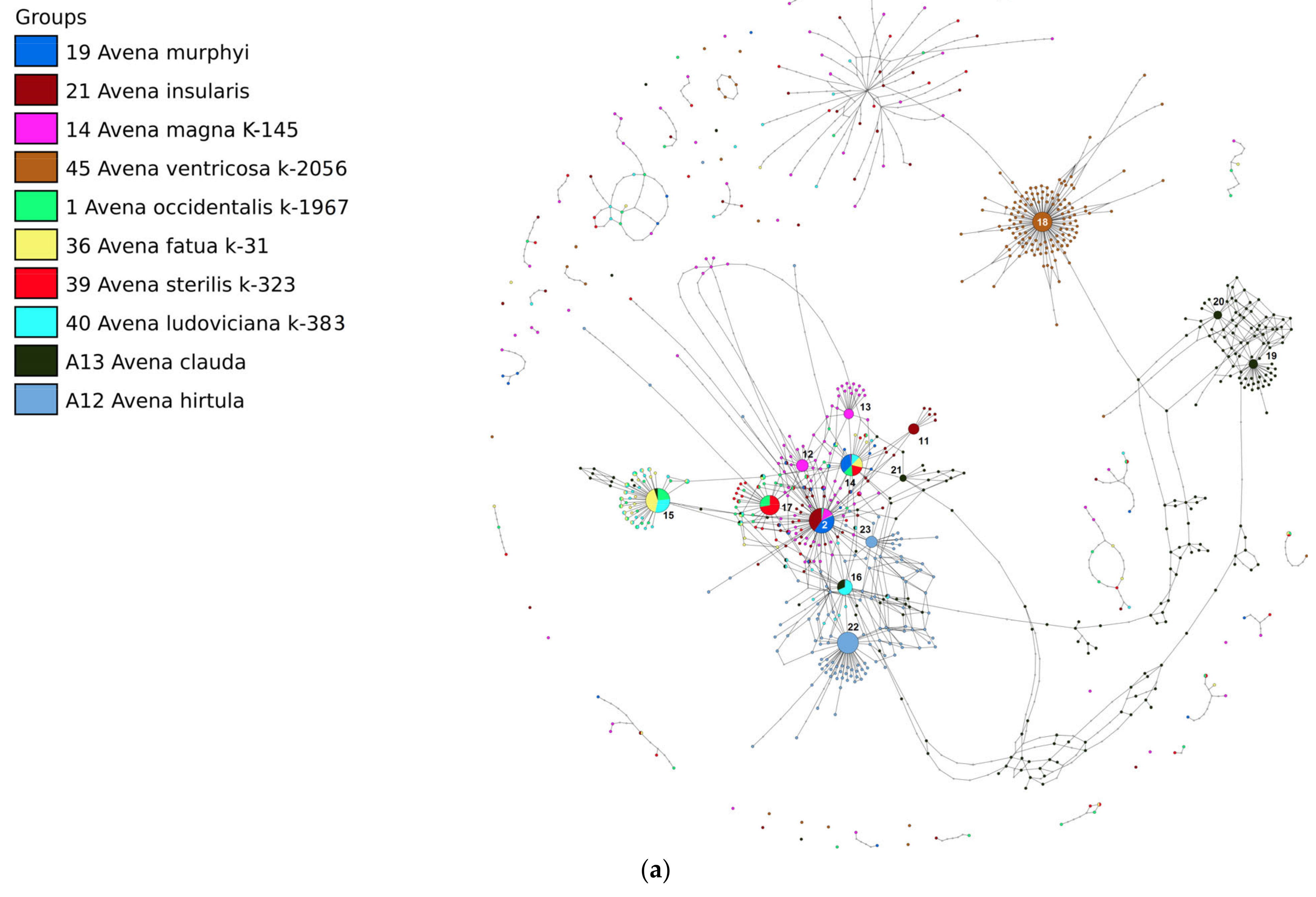
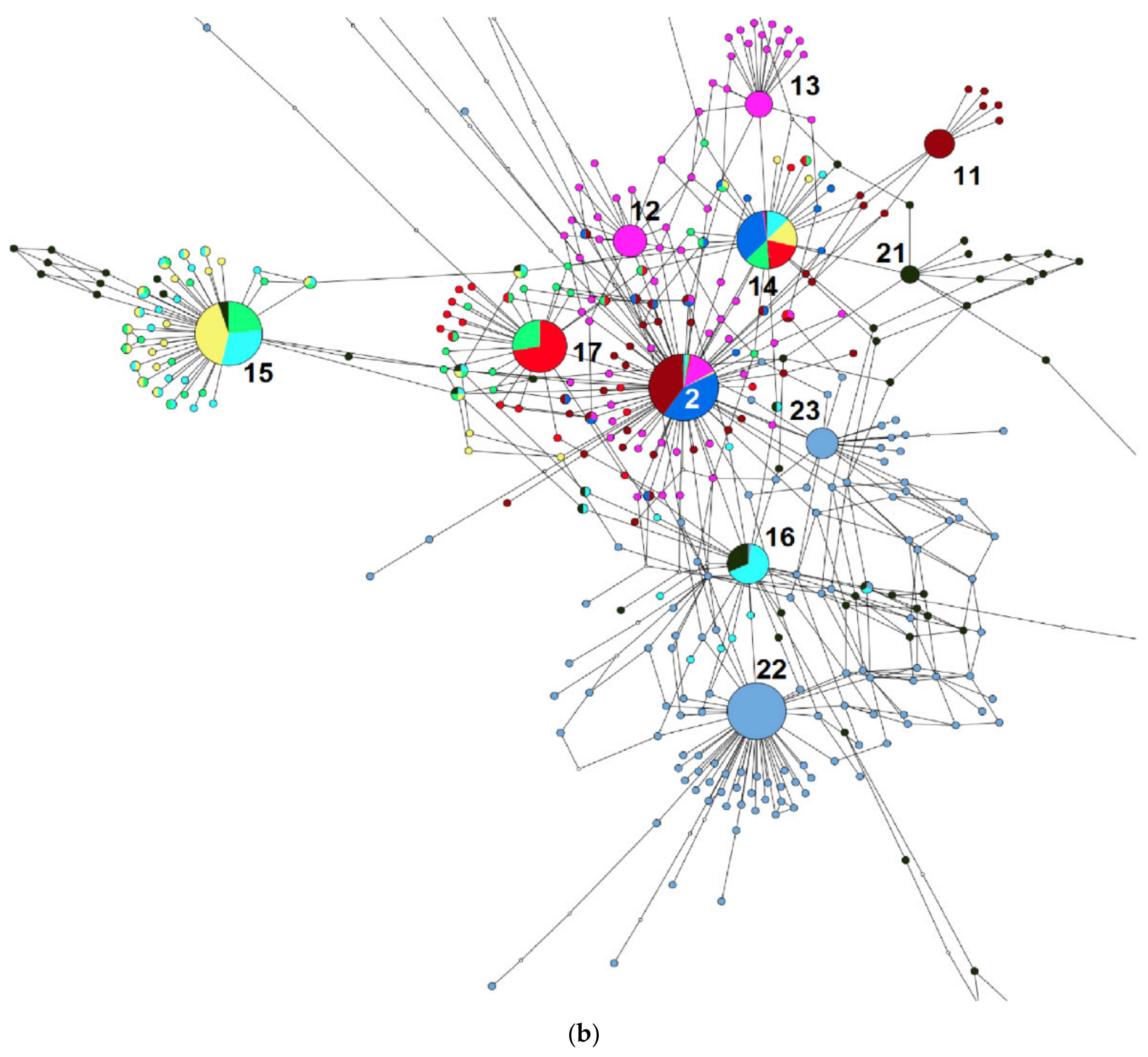
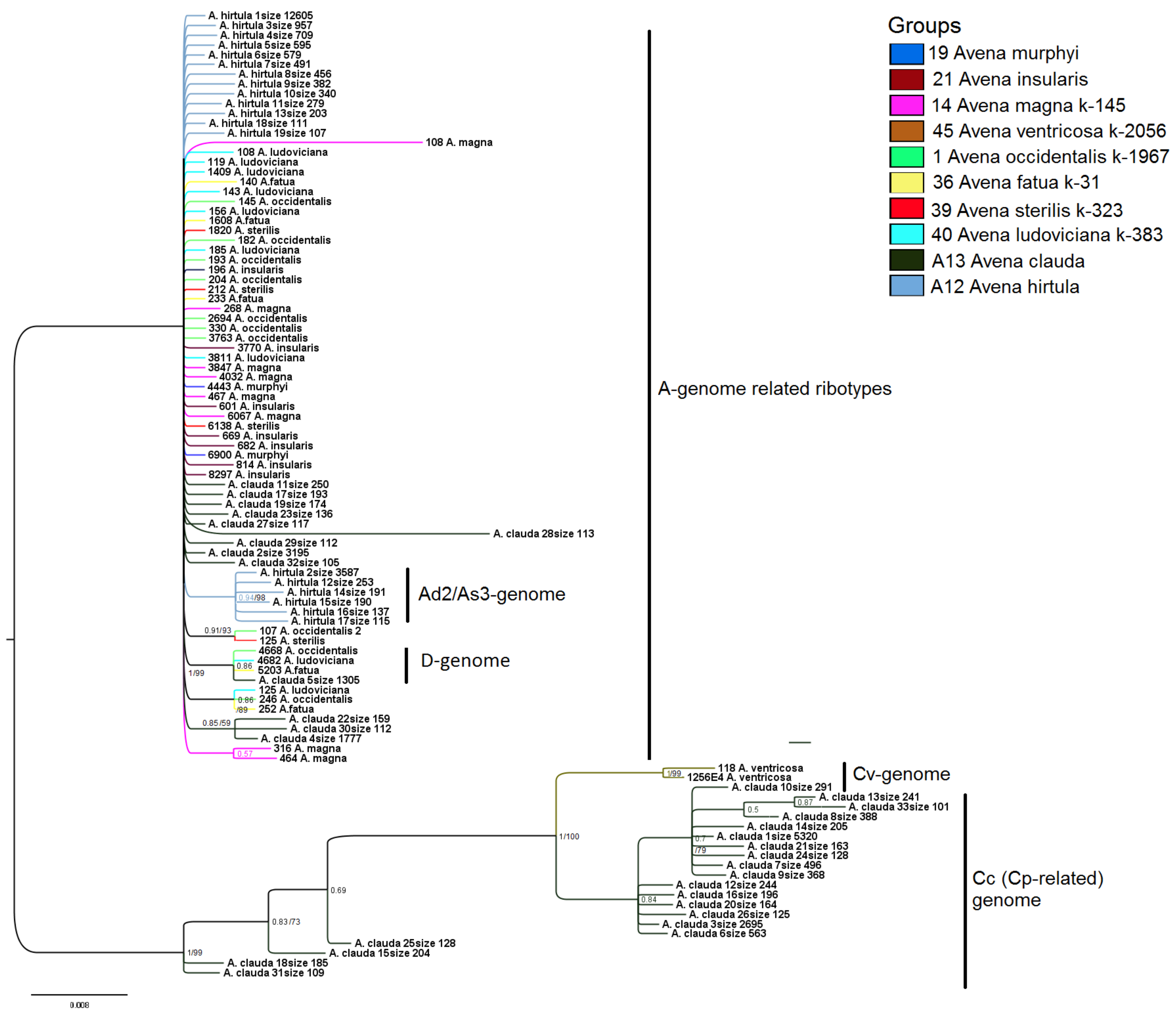
2. Results







3. Discussion
4. Conclusions
5. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitchell, C.C.; Parkinson, S.E.; Baker, T.J.; Jellen, E.N. C-banding and localization of 18S–5.8S S-26S rDNA in tall oatgrass species. Crop. Sci. 2003, 43, 32–36. [Google Scholar] [CrossRef]
- Leggett, J.M.; Markland, G.S. The genomic structure of Avena revealed by GISH. In Kew Chromosome Conference IV; Brandham, P.E., Bennett, M.D., Eds.; Royal Botanic Gardens: Kew, UK, 1995; pp. 133–139. [Google Scholar]
- Loskutov, I.G.; Rines, H.W. Avena. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Chapter 3; pp. 109–183. [Google Scholar]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold: London, UK, 1971; 216p. [Google Scholar]
- Fabijanski, S.; Fedak, G.; Armstrong, K.; Altosaar, I. A repeated sequence probe for the C genome in Avena (oats). Theor. Appl. Genet. 1990, 79, 1–7. [Google Scholar] [CrossRef]
- Katsiotis, A.; Hagidimitriou, M.; Heslop-Harrison, J.S. The close relationship between the A and B genomes in Avena L. (Poaceae) determined by molecular cytogenetic analysis of total genomic, tandemly and dispersed repetitive DNA sequences. Ann. Bot. 1997, 79, 103–109. [Google Scholar] [CrossRef]
- Peng, Y.; Zhou, P.; Zhao, J.; Li, J.; Lai, S.; Tinker, N.A.; Liao, S.; Yan, H. Phylogenetic relationships in the genus Avena based on the nuclear Pgk1 gene. PLoS ONE 2018, 13, e0200047. [Google Scholar] [CrossRef]
- Chew, P.; Meade, K.; Hayes, A.; Harjes, C.; Bao, Y.; Beattie, A.D.; Puddephat, I.; Gusmini, G.; Tanksley, S.D. A study on the genetic relationships of Avena taxa and the origins of hexaploid oat. Theor. Appl. Gen. 2016, 129, 1405–1415. [Google Scholar] [CrossRef]
- Chen, Q.; Armstrong, K. Genomic in situ hybridization in Avena sativa. Genome 1994, 37, 607–612. [Google Scholar] [CrossRef]
- Linares, C.; González, J.; Ferrer, E.; Fominaya, A. The use of double fluorescence in situ hybridization to physically map the positions of 5S rDNA genes in relation to the chromosomal location of 18S-5.8S-26S rDNA and a C genome specific DNA sequence in the genus Avena. Genome 1996, 39, 535–542. [Google Scholar] [CrossRef]
- Loskutov, I.G. Oat (Avena L.). Distribution, Taxonomy, Evolution and Breeding Value; VIR: St. Petersburg, Russia, 2007; 336p. [Google Scholar]
- Leggett, J.M.; Thomas, H. Oat evolution and cytogenetics. In The Oat Crop Production and Utilization; Welch, R.W., Ed.; Chapman & Hall: London, UK, 1995; pp. 120–149. [Google Scholar]
- Ladizinsky, G. A new species of Avena from Sicily, possible the tetraploid progenitor of hexaploid oats. Genet. Resour. Crop Evol. 1998, 45, 263–269. [Google Scholar] [CrossRef]
- Li, C.D.; Rossnagel, B.G.; Scoles, G.J. The development of oar microsatellite markers and their use in identifying relationships among Avena species and oat cultivars. Theor. Appl. Genet. 2000, 101, 1259–1268. [Google Scholar] [CrossRef]
- Tomaszewska, P.; Schwarzacher, T.; Heslop-Harrison, J.S. Oat chromosome and genome evolution defined by widespread terminal intergenomic translocations in polyploids. Front. Plant Sci. 2022, 13, 1026364. [Google Scholar] [CrossRef]
- Yan, H.; Ren, Z.; Deng, D.; Yang, K. New evidence confirming the CD genomic constitutions of the tetraploid Avena species in the section Pachycarpa Baum. PLoS ONE 2021, 16, e0240703. [Google Scholar] [CrossRef]
- Morikawa, T. Isozyme and chromosome polymorphisms of the genus Avena and its geographic distribution in Morocco. Wheat Inform. Serv. 1991, 72, 104–105. [Google Scholar]
- Yan, H.-H.; Baum, B.R.; Zhou, P.-P.; Zhao, J.; Wei, Y.-M.; Ren, C.-Z.; Xiong, F.-Q.; Liu, G.; Zhong, L.; Zhao, G.; et al. Phylogenetic analysis of the genus Avena based on chloroplast intergenic spacer psbA-trnH and single-copy nuclear gene Acc1. Genome 2014, 57, 267–277. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, H.; Guo, L.; Deng, C.; Wang, C.; Wang, Y.; Kang, L.; Zhou, P.; Yu, K.; Dong, X.; et al. Reference genome assemblies reveal the origin and evolution of allohexaploid oat. Nat. Genet. 2022, 54, 1248–1258. [Google Scholar] [CrossRef]
- Jellen, E.N.; Phillips, R.L.; Rines, H.W. Chromosomal localization and polymorphisms of ribosomal DNA in oat (Avena spp.). Genome 1994, 37, 23–32. [Google Scholar] [CrossRef]
- Linares, C.; Ferrer, E.; Fominaya, A. Discrimination of the closely related A and D genomes of the hexaploid oat Avena sativa L. PNAS 1998, 95, 12450–12455. [Google Scholar] [CrossRef] [PubMed]
- Ladizinsky, G. New evidence on the origin of hexaploidy oats. Evolution 1969, 23, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.C.; Sadanaga, K.; Zilinsky, F.J.; Terrell, E.; Smith, R.T. Avena magna: An important new tetraploid species in oats. Science 1968, 159, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jellen, E.N.; Murphy, J.P. Progenitor germplasm of domesticated hexaploid oat. Crop. Sci. 1999, 39, 1208–1214. [Google Scholar] [CrossRef]
- Baum, B.R. Avena occidentalis, a hitherto overlooked species of oats. Can. J. Bot. 1971, 49, 1055–1057. [Google Scholar] [CrossRef]
- Gnutikov, A.A.; Nosov, N.N.; Loskutov, I.G.; Machs, E.M.; Blinova, E.V.; Probatova, N.S.; Langdon, T.; Rodionov, A.V. New insights into the genomic structure of the oats (Avena L., Poaceae): Intragenomic polymorphism of ITS1 sequences of rare endemic species Avena bruhnsiana Gruner and its relationship to other species with C-genomes. Euphytica 2022, 218, 3. [Google Scholar] [CrossRef]
- Gnutikov, A.A.; Nosov, N.N.; Loskutov, I.G.; Blinova, E.V.; Shneyer, V.S.; Probatova, N.S.; Rodionov, A.V. New Insights into the Genomic Structure of Avena L.: Comparison of the Divergence of A-Genome and One C-Genome Oat Species. Plants 2022, 11, 1103. [Google Scholar] [CrossRef]
- Brassac, J.; Blattner, F.R. Species-Level Phylogeny and Polyploid Relationships in Hordeum (Poaceae) Inferred by Next-Generation Sequencing and In Silico Cloning of Multiple Nuclear Loci. Syst. Biol. 2015, 64, 792–808. [Google Scholar] [CrossRef]
- Schultz, J.; Maisel, S.; Gerlach, D.; Müller, T.; Wolf, M. A common core of secondary structure of 33 the internal transcribed spacer 2 (ITS2) throughout the Eukaryota. RNA 2005, 11, 361–364. [Google Scholar] [CrossRef]
- Coleman, A.W. Nuclear rRNA transcript processing versus internal transcribed spacer 29 secondary structure. Trends Genet. 2015, 31, 157–163. [Google Scholar] [CrossRef]
- Zhang, X.; Cao, Y.; Zhang, W.; Simmons, M.P. Adenine· cytosine substitutions are an alternative 13 pathway of compensatory mutation in angiosperm ITS2. RNA 2020, 26, 209–217. [Google Scholar] [CrossRef]
- Peng, Y.Y.; Wei, Y.M.; Baum, B.R.; Zheng, Y.L. Molecular diversity of the 5S rRNA gene and genomic relationships in the genus Avena (Poaceae: Aveneae). Genome 2008, 51, 137–154. [Google Scholar] [CrossRef]
- Maughan, P.J.; Lee, R.; Walstead, R.; Vickerstaff, R.J.; Fogarty, M.C.; Brouwer, C.R.; Reid, R.R.; Jay, J.J.; Bekele, W.A.; Jackson, E.W.; et al. Genomic insights from the first chromosome-scale assemblies of oat (Avena spp.) diploid species. BMC Biol. 2019, 17, 92. [Google Scholar] [CrossRef]
- Peng, Y.-Y.; Baum, B.R.; Re, C.-Z.; Jiang, Q.-T.; Chen, G.-Y.; Zheng, Y.-L.; Wei, Y.-M. The evolution pattern of rDNA ITS in Avena and phylogenetic relationship of the Avena species (Poaceae: Aveneae). Hereditas 2010, 147, 183–204. [Google Scholar] [CrossRef]
- Rodionov, A.V.; Tiupa, N.B.; Kim, E.S.; Machs, E.M.; Loskutov, I.G. Genomic structure of the autotetraploid oat species Avena macrostachya inferred from comparative analysis of the ITS1 and ITS2 sequences: On the oat karyotype evolution during the early stages of the Avena species divergence. Genetika 2005, 41, 646–656. (In Russian) [Google Scholar]
- Baldwin, B.G.; Sanderson, M.J.; Porter, J.M.; Wojciechowski, M.F.; Campbell, C.S.; Donoghue, M.J. The ITS Region of Nuclear Ribosomal DNA: A Valuable Source of Evidence on Angiosperm Phylogeny. Ann. Mis. Bot. Gard. 1995, 82, 247–277. [Google Scholar] [CrossRef]
- Mahelka, V.; Krak, K.; Kopecký, D.; Fehrer, J.; Šafář, J.; Bartoš, J.; Hobza, R.; Blavet, N.; Blattner, F.R. Multiple horizontal transfers of nuclear ribosomal genes between phylogenetically distinct grass lineages. Biol. Sci. 2017, 114, 1726–1731. [Google Scholar] [CrossRef] [PubMed]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.D.P.N.; Riina, R.; Valduga, E.; Caruzo, M.B.R. A new species of Croton (Euphorbiaceae) endemic to the Brazilian Pampa and its phylogenetic affinities. Pl. Syst. Evol. 2022, 308, 14. [Google Scholar] [CrossRef]
- Suyama, Y.; Hirota, S.K.; Matsuo, A.; Tsunamoto, Y.; Mitsuyuki, C.; Shimura, A.; Okano, K. Complementary combination of multiplex high-throughput DNA sequencing for molecular phylogeny. Ecol. Res. 2022, 37, 171–181. [Google Scholar] [CrossRef]
- Saarela, J.M.; Bull, R.D.; Paradis, M.J.; Ebata, S.N.; Peterson, P.M.; Soreng, R.J.; Paszko, B. Molecular phylogenetics of cool-season grasses in the subtribes Agrostidinae, Anthoxanthinae, Aveninae, Brizinae, Calothecinae, Koeleriinae and Phalaridinae (Poaceae, Pooideae, Poeae, Poeae chloroplast group 1). PhytoKeys 2017, 87, 1–139. [Google Scholar] [CrossRef]
- Tkach, N.; Schneider, J.; Döring, E.; Wölk, A.; Hochbach, A.; Nissen, J.; Winterfeld, G.; Meyer, S.; Gabriel, J.; Hoffmann, M.H.; et al. Phylogenetic lineages and the role of hybridization as driving force of evolution in grass supertribe Poodae. Taxon 2020, 69, 234–277. [Google Scholar] [CrossRef]
- Fu, Y.B. Oat evolution revealed in the maternal lineages of 25 Avena species. Sci. Rep. 2018, 8, 4252. [Google Scholar] [CrossRef]
- Ladizinsky, G. The cytogenetic position of Avena prostrata among the diploid oats. Can. J. Genet. Cytol. 1973, 15, 443–450. [Google Scholar] [CrossRef]
- Malzew, A.I. Wild and Cultivated Oat. Sectio Euavena Griseb; Publishers of the All-Union Institute of Applied Botany and New Cultures under the Council of People’s Commissars of the USSR: Leningrad, Russia, 1930; 522p. [Google Scholar]
- Emme, E.K. Karyosystematic research of oat of section Euavena Griseb. Works Appl. Bot. Plant Breed Seria II 1932, 1, 147–168. [Google Scholar]
- Baum, B.R.; Fedak, G. A new tetraploid species of Avena discovered in Morocco. Can. J. Bot. 1985, 63, 1379–1385. [Google Scholar] [CrossRef]
- Morikawa, T. Isozyme and chromosome variations of the Avena species in the Canary Islands and Morocco. In Proceedings of the 4th International Oat Conference, Adelaide, SA, Australia, 19–23 October 1992; Volume 3, pp. 138–140. [Google Scholar]
- Leggett, J.M. Interspecific diploid hybrids in Avena. Genome 1989, 32, 346–348. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Shelukhina, O.; Diederichsen, A.; Loskutov, I.G.; Pukhalskiy, V.A. Comparative cytogenetic analysis of Avena macrostachya and diploid C-genome Avena species. Genome 2010, 53, 125–137. [Google Scholar] [CrossRef]
- Murray, B.E.; Craig, J.L.; Rajhathy. T. A protein electrophoretic study of three amphiploids and eight species in Avena. Can. J. Genet. Cytol. 1970, 12, 651–655. [Google Scholar] [CrossRef]
- Yan, H.; Bekele, W.A.; Wight, C.P.; Peng, Y.; Langdon, T.; Latta, R.G.; Fu, Y.-B.; Diederichsen, A.; Howarth, C.J.; Jellen, E.N.; et al. High-density marker profiling confirms ancestral genomes of Avena species and identifies D-genome chromosomes of hexaploid oat. Theor. Appl. Genet. 2016, 129, 2133–2149. [Google Scholar] [CrossRef]
- Sadasivaiah, R.S.; Rajhathy, T. Genome relationships in tetraploid Avena. Can. J. Genet. Cytol. 1968, 10, 655–669. [Google Scholar] [CrossRef]
- Rajhathy, T. Chromosome polymorphism in Avena ventricosa. Chromosoma 1971, 35, 206–216. [Google Scholar] [CrossRef]
- Rodionov, A.V.; Shneyer, V.S.; Gnutikov, A.A.; Nosov, N.N.; Punina, E.O.; Zhurbenko, P.M.; Loskutov, I.G.; Muravenko, O.V. Species dialectics: From initial uniformity, through the greatest possible diversity to ultimate uniformity. Bot. Zhurn. 2020, 105, 835–853. (In Russian) [Google Scholar] [CrossRef]
- Rajhathy, T.; Thomas, H. Cytogenetics of Oats (Avena L.) Miscellaneous Publications of the Genetics Society of Canada 2; Ontario (Canada) Genetics Society of Canada: Ottawa, ON, Canada, 1974; 90p. [Google Scholar]
- Rajhathy, T. A standard kariotype for Avena sativa. Can. J. Genet. Cytol. 1963, 5, 127–132. [Google Scholar] [CrossRef]
- Rajhathy, T.; Morrison, J.W. Chromosome morphology in the genus Avena. Can. J. Bot. 1959, 37, 331–337. [Google Scholar] [CrossRef]
- Nishiyama, I. The genetic and cytology of certain cereals. I. Morphological and cytological studies in triploid, pentaploid and hexaploid Avena hybrids. Jpn. J. Genet. 1929, 5, 1–48. [Google Scholar] [CrossRef]
- Fominaya, A.; Loarce, Y.; González, J.M.; Ferrer, E. Cytogenetic evidence supports Avena insularis being closely related to hexaploid oats. PLoS ONE 2021, 16, e0257100. [Google Scholar] [CrossRef] [PubMed]
- Ladizinsky, G.; Zohary, D. Notes on species delimination species relationships and poliploidy in Avena L. Euphytica 1971, 20, 380–395. [Google Scholar] [CrossRef]
- Sadanaga, K. Chromosome Homology of Avena magna Murphy and Terrell and the Hexaploid Species A. sativa L. and A. byzantina C. Koch. Crop Sci. 1976, 16, 425–428. [Google Scholar] [CrossRef]
- Nikoloudakis, N.; Skaracis, G.A.; Katsiotis, A. Evolutionary insights inferred by molecular analysis of the ITS1-5.8S-ITS2 and IGS Avena sp. sequences. Mol. Phyl. Evol. 2008, 46, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Viegas, W.; Silva, M. 45S rDNA external transcribed spacer organization reveals new phylogenetic relationships in Avena genus. PLoS ONE 2017, 12, e0176170. [Google Scholar] [CrossRef]
- Clarkson, P.M.; Hoffman, E.P.; Zambraski, E.; Gordish-Dressman, H.; Kearns, A.; Hubal, M.; Harmon, B.; Devan, J.M. ACTN3 and MLCK genotype associations with exertional muscle damage. J. Appl. Physiol. 2005, 99, 564–569. [Google Scholar] [CrossRef]
- Mandáková, T.; Pouch, M.; Harmanová, K.; Zhan, S.H.; Mayrose, I.; Lysak, M.A. Multispeed genome diploidization and diversification after an ancient allopolyploidization. Mol. Ecol. 2017, 26, 6445–6462. [Google Scholar] [CrossRef]
- Nikoloudakis, N.; Katsiotis, A. The origin of the C-genome and cytoplasm of Avena polyploids. Theor. Appl. Genet. 2008, 117, 273–281. [Google Scholar] [CrossRef]
- Ridgway, K.P.; Duck, J.M.; Young, J.P.W. Identification of roots from grass swards using PCR-RFLP and FFLP of the plastid trnL (UAA) intron. BMC Ecol. 2003, 3, 8. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef]
- Múrias dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2016, 32, 627–628. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Sample ID | Country of Origin | Accession Number | Collected by | Genome | Total Number of Reads | Ribotype Number in Figure 2 and Figure 4 | Ribotype Symbol (of the Major Ribotypes) | Number of Reads | % from the Total Number of the Reads |
|---|---|---|---|---|---|---|---|---|---|---|
| Avena atlantica | K-2108 | Morocco | from OM004717 to OM004731; from OQ338511 to OQ338567 | A. Diederichsen | As | 16,241 | 1 | As1 | 6353 | 39 |
| 2 | As2 | 4571 | 28 | |||||||
| Avena canariensis | K-2114 | Spain | from OM004567 to OM004604; from OQ338401 to OQ338510 | A. Diederichsen | Ac | 20,606 | 3 | Ac1 | 10,050 | 49 |
| 4 | Ac2 | 1908 | 9 | |||||||
| Avena agadiriana | K-2123 | Morocco | from OQ135344 to OQ135431 | AB? | 25,194 | 5 | B1 | 7136 | 28 | |
| 6 | B2 | 4919 | 41 | |||||||
| 7 | B3 | 1275 | 5 | |||||||
| 2 | As2 | 1018 | 4 | |||||||
| Avena barbata | K-2071 | Lebanon | from OQ135432 to OQ135477 | AB | 14,896 | 1 | As1 | 4894 | 33 | |
| 8 | B4 | 1014 | 6 | |||||||
| Avena vaviloviana | K-755 | Ethiopia | from OQ135478 to OQ135562 | AB | 23,292 | 1 | As1 | 10,884 | 47 | |
| 9 | B5 | 3255 | 14 | |||||||
| 10 | B6 | 1162 | 5 | |||||||
| Avena insularis | K-2067 | Italy | from OQ135592 to OQ135663 | AC or CD | 21,046 | 2 | As2 | 8297 | 39 | |
| 11 | Ai | 3770 | 18 | |||||||
| Avena magna | K-145 | Morocco | from OQ135664 to OQ135782 | E. V. Mazhorov | AC or CD | 25,167 | 12 | Am1 | 6067 | 24 |
| 13 | Am2 | 4032 | 16 | |||||||
| 2 | As2 | 3847 | 15 | |||||||
| Avena murphyi | K-2088 | Spain | from OQ135563 to OQ135591 | AC or CD | 15,596 | 2 | As2 | 6900 | 44 | |
| 14 | Amp | 4443 | 28 | |||||||
| Avena fatua | K-31 | Kazakhstan | from OQ144753 to OQ144793 | ACD | 12,280 | 15 | D | 5203 | 42 | |
| 14 | Amp | 1608 | 13 | |||||||
| Avena ludoviciana | K-383 | Russian Federation | from OQ144842 to OQ144891 | ACD | 15,740 | 15 | D | 4682 | 30 | |
| 16 | Ald/Cc2A | 3811 | 24 | |||||||
| 14 | Amp | 1409 | 9 | |||||||
| Avena occidentalis | K-1967 | Spain, Canary is. | from OQ144691 to OQ144752 | ACD | 21,870 | 15 | D/Cc2C | 4668 | 21 | |
| 17 | Ast (D′?) | 3763 | 17 | |||||||
| 14 | Amp | 2694 | 12 | |||||||
| Avena sterilis | K-323 | Russian Federation | from OQ144794 to OQ144841 | ACD | 13,147 | 17 | Ast (D′?) | 6138 | 47 | |
| 14 | Amp | 1820 | 14 | |||||||
| Avena ventricosa | K-2056 | Algeria | from OK301935 to OK302014; from OQ520145 to OQ520214 | Cv | 15,506 | 18 | Cv | 12,560 | 81 | |
| Avena clauda | K-269 | Azerbaijan | from OK273905 to OK274031; from OQ834465 to OQ834560 | V. N. Soldatov | Cp | 40,168 | 19 | Cc1A | 5320 | 13 |
| 20 | Cc1B | 2695 | 7 | |||||||
| 16 | Ald/Cc2A | 3195 | 8 | |||||||
| 21 | Cc2B | 1777 | 4 | |||||||
| 15 | D/Cc2C | 1305 | 3 | |||||||
| Avena hirtula | k-1878 | Spain | from OM004668 toOM004716; from OQ834671 to OQ834744 | M. Leggett | As | 31,625 | 22 | As5 | 12,605 | 40 |
| 23 | Ad2/As3 | 3587 | 11 |
| 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | |||
| 8 | 9 | 0 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 3 | 3 | 3 | 4 | 4 | 4 | 7 | 7 | 7 | 7 | 8 | 8 | 8 | 9 | 0 | 0 | 3 | 3 | 4 | 4 | 4 | 4 | 4 | 5 | 5 | 6 | 6 | 6 | 7 | 8 | 1 | |
| 7 | 6 | 8 | 2 | 3 | 5 | 6 | 8 | 2 | 3 | 0 | 4 | 8 | 3 | 5 | 9 | 1 | 4 | 5 | 7 | 2 | 4 | 9 | 0 | 3 | 8 | 0 | 7 | 0 | 1 | 3 | 4 | 7 | 0 | 7 | 0 | 5 | 9 | 4 | 1 | 6 | |
| As1 | A | A | C | T | C | T | G | T | T | G | A | D | C | A | C | A | T | A | G | G | C | T | T | A | G | G | C | T | G | G | C | A | A | G | T | C | C | T | G | T | C |
| As2 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| B1 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | A | . | . | . | . | A | . | . |
| B2 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| B3 | . | . | T | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| B4 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | T |
| Ac1 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | - | . | . | . | . | . | T | . | . | . | . | . |
| Ac2 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | T | . | . | - | . | . | . | . | . | T | . | . | . | G | . |
| B5 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| B6 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ai | G | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . |
| Am1 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | A | . | . | . | . | . | . | . | . | . | . | . |
| Am2 | . | . | . | . | . | . | . | . | . | T | C | D | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Amp | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| D/Cc2C | . | . | . | . | . | . | A | . | . | . | C | D | . | . | . | . | C | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ald/Cc2A | . | . | . | . | . | . | . | . | . | . | C | D | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | |
| Ast (D′?) | . | . | . | . | . | . | . | . | . | . | C | D | . | . | A | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Cv | . | C | . | D | D | C | . | C | C | . | C | C | T | G | . | G | C | G | A | . | A | C | A | T | A | A | . | C | T | . | T | G | G | . | . | . | T | G | . | . | . |
| Cc1A | . | C | . | D | D | C | . | . | C | . | C | C | T | G | . | G | C | G | A | . | . | C | . | . | A | A | . | C | T | . | T | G | G | . | C | T | T | G | . | . | . |
| Cc1B | . | C | . | D | D | C | . | C | C | . | C | C | T | G | . | G | C | G | A | . | . | C | . | . | A | A | . | C | T | . | T | G | G | . | C | T | T | . | . | . | . |
| Cc2B | . | . | G | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| As5 | . | . | . | . | . | . | . | . | . | . | . | D | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
| Ad2/As3 | . | . | . | . | . | . | . | . | . | . | C | D | . | . | . | . | . | . | T | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . | . |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnutikov, A.A.; Nosov, N.N.; Loskutov, I.G.; Blinova, E.V.; Shneyer, V.S.; Rodionov, A.V. Origin of Wild Polyploid Avena Species Inferred from Polymorphism of the ITS1 rDNA in Their Genomes. Diversity 2023, 15, 717. https://doi.org/10.3390/d15060717
Gnutikov AA, Nosov NN, Loskutov IG, Blinova EV, Shneyer VS, Rodionov AV. Origin of Wild Polyploid Avena Species Inferred from Polymorphism of the ITS1 rDNA in Their Genomes. Diversity. 2023; 15(6):717. https://doi.org/10.3390/d15060717
Chicago/Turabian StyleGnutikov, Alexander A., Nikolai N. Nosov, Igor G. Loskutov, Elena V. Blinova, Victoria S. Shneyer, and Alexander V. Rodionov. 2023. "Origin of Wild Polyploid Avena Species Inferred from Polymorphism of the ITS1 rDNA in Their Genomes" Diversity 15, no. 6: 717. https://doi.org/10.3390/d15060717
APA StyleGnutikov, A. A., Nosov, N. N., Loskutov, I. G., Blinova, E. V., Shneyer, V. S., & Rodionov, A. V. (2023). Origin of Wild Polyploid Avena Species Inferred from Polymorphism of the ITS1 rDNA in Their Genomes. Diversity, 15(6), 717. https://doi.org/10.3390/d15060717