Abstract
Epiphytic bryophytes are important components of forest ecosystems and play important roles in maintaining biodiversity and ecosystem function. However, the main factors driving epiphytic bryophyte diversity remain unclear. We collected the tree epiphytic bryophytes from a one-hectare plot within a temperate deciduous broadleaf forest (China). Canonical correspondence analyses and Mantel tests were used to establish linear regression models and thus dissect the effects of environmental variables (topography, light and bark physicochemical properties) on the species diversity, functional diversity, and phylogenetic diversity of epiphytic bryophytes. The relationship between environmental variables and epiphytic bryophyte diversity was analyzed using piecewise structural equation modeling. Results showed that the physicochemical properties of the bark directly influenced the species diversity and phylogenetic diversity of the epiphytic bryophytes. The physical and chemical properties of bark also indirectly affected the functional diversity of the epiphytic bryophytes. Elucidation of the factors driving epiphytic bryophyte diversity provides insights into their conservation.
1. Introduction
Bryophytes (mosses, liverworts, and hornworts) are extensively distributed throughout the world, except in deserts and arid areas. They are the earliest land plants, with 480 million years of evolution. Bryophyta (mosses) are the second most diverse phylum of land plants after angiosperms, comprising approximately 13,000 species [1]. Bryophytes play significant roles in ecosystems, such as contributing to the maintenance of forest biodiversity, water retention, soil improvement, and nutrient cycling [2], as well as being ecological indicators.
Forest biodiversity patterns and their formation are core components of community ecology research [3]. Understanding the formation, maintenance, and loss of biodiversity patterns can help in the sustainable development and conservation of biodiversity [4]. Tree epiphytic bryophytes are particularly sensitive to environmental changes in climate and air quality [5].
In a temperate deciduous broadleaf forest, environmental variables such as light, topography, and bark physicochemical properties (BPPs) are interrelated. Given these complex inter-relationships, an in-depth analysis of their combined effects is necessary for understanding and predicting the future impact of environmental changes. For example, BPPs reportedly influence the epiphytic specificity and distribution of epiphytic bryophytes [6,7,8]. Different woody plants have different BPPs, which in turn affect epiphyte colonization [9,10]. Light is also an important environmental factor in forest ecosystems: both directly via photosynthesis and indirectly by changing moisture and humidity. Topography such as elevation is a further determinant of plant communities [11].
Biodiversity studies include species diversity, phylogenetic diversity (PD), and functional diversity (i.e., functional richness (FRic)). Some community ecologists have sought to link woody plant biodiversity (species diversity, FRic, or PD) to abiotic environmental variables to infer the community–assembly mechanisms [12,13,14,15,16,17]. However, the particular effects of an individual environmental factor have been extensively studied, whereas few have considered their combined effects and interactions. Studying ecosystems goes beyond considering only species diversity, and should include phylogenetic, functional trait, habitat, and ecosystem diversity [18]. Accordingly, we investigated the relationship between diversity (species diversity, PD, and FRic) and environmental variables to understand the environmental factors driving the diversity in epiphytic bryophyte communities. We aimed to clarify the factors (light, topography, or BPPs) primarily driving the diversity. Our results may contribute to an understanding of the factors driving epiphytic bryophyte diversity and provide ideas for conserving epiphytic bryophytes.
2. Materials and Methods
2.1. Study Location and Sampling Design
This study was conducted in the Baiyunshan National Nature Reserve 10,000 m2 forest dynamic plot (111°480–112′160″ E, 33°330–33′560″ N), Songxian County, east China. The White River (a tributary of the Yangtze River), the Yi River (a tributary of the Yellow River), and the Ru River (a tributary of the Huai River) all originate at the Yuhuangding peak, which is 2216 m above sea level. Funiu Mountain in China is an important geographical boundary with a warm temperate zone and subtropical zone. It is also a complex ecosensitive area with a high percentage of forest cover (>90%). The reserve’s forest type and structure are intricate, varied, and rich, making it one of the relatively rare regions in central China with clearly defined vertical forest-distribution zones. The annual mean temperature in the region is 13.5 °C, the annual mean precipitation is 1200 mm, and the forest cover is 81.2% (Figure 1).
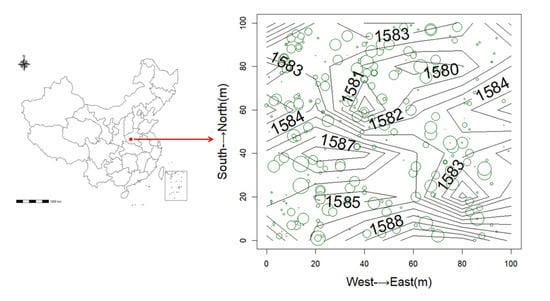
Figure 1.
Location and topography of plots in the Baiyunshan National Reserve. The star indicates the location of the plot; the line indicates the elevation of the plot; the green circle indicates the woody plants in the plot; and the size of the circle indicates the diameter of woody plants at breast height.
All stems ≥ 1 cm diameter at breast height in the 10,000 m2 plot were tagged, mapped, and measured. The forest type of the plot is a deciduous broadleaf forest, and its dominant tree is Quercus aliena Bl. var. acuteserrata Maxim. ex Wenz. All epiphytic bryophytes in the plot were collected based on 10 m × 10 m subplots, with a total of 100 subplots. In August 2020, all epiphytic bryophytes growing on the same side of woody plants within the sample plots were collected. The collection range was from the roots of the woody plant at ground level to 1.5 m up the tree trunk. A piece of bark (5 cm × 5 cm) was collected at 1.3 m. The data for all epiphytic bryophytes and woody plant species are shown in Table 1.

Table 1.
Lists of the tree species and the epiphytic bryophytes.
2.2. Epiphytic Bryophyte DNA and Functional-Trait Data Collection
The epiphytic bryophytes were identified from macroscopic characteristics (including stem length, leaf length) and microscopic features (including cell width, cell shape).
The cetyltrimethylammonium bromide method was used to extract DNA from each bryophyte. Three common barcoding genes, namely, rbcL, rps4, and trnL-F, were amplified using the polymerase chain reaction. The amplified products were examined via gel electrophoresis and then sent to BGI for sequencing. The sequence results returned by BGI were retrieved in NCBI, and the bryophyte species were identified based on their macrocharacteristics [19,20]. The gene sequences were used for phylogenetic tree construction.
Eight functional traits for each species were measured: stem length, leaf area, leaf length-to-width ratio, middle cell length, middle cell width, capsule length, spore diameter, and spore thickness. The measurement protocols for these traits followed a previous work [21]. Each functional trait was measured three times. All functional data are shown in Table S7.
For stem lengths we measured heights t from the part of the plant in contact with the substrate to the top of the plant. Setal lengths were from the foot to the base of the capsule. Leaf area and leaf length-to-width ratios were calculated from fully hydrated leaves.
2.3. Environmental Data
Four topography variables were measured for each 10 m × 10 m subplot, with a total of 100 subplots. The elevation of the four corners of the subplot in the field was recorded. Based on the elevation data, we calculated the average elevation, slope, aspect, and convexity [15,22].
Six light variables were measured for each 10 m × 10 m subplot: subcanopy scattered radiation (SRUC), direct radiation under the canopy, transmittance of light (TT), canopy cover (CC), leaf area index (LAI), and average leaf inclination (ALA). For the collection method, an ultra-wide-angle fisheye lens was connected to a Canon 7D camera, which was then used to take hemispherical photos at the center of each 10 m × 10 m subplot. Light data were collected from 8 a.m. to 10 a.m. and from 4 p.m. to 6 p.m. on clear days without wind and fog, thus avoiding blurred images. A HemiView canopy light-analysis system was used to calculate the canopy light factor data of each 10 m × 10 m subplot [23].
A total of eight BPP variables (N, P, K, C, S, Si, pH, and bark cracking depth (CD)) were measured [24,25,26]. The total nitrogen (N) of barks was determined via the micro-Kjeldahl method of the means of micronutrients. The barks’ total phosphorus (P) was determined via molybdenum–antimony colorimetry with H2SO4–H2O2. Total potassium (K) was determined using the flame-photometry method (H2SO4–H2O2). Organic carbon (C) was determined using the K2Cr2O7-capacity method. Silicon (Si) content was determined using the colorimetric silicon–molybdenum blue method. Barks were boiled in concentrated HNO3 to determine the sulfur (S) content. Bark pH was determined by potentiometry, and crack depth was measured with a Vernier caliper.
2.4. Statistical Procedures
Epiphytic bryophyte diversity was calculated using three indices (species diversity, FRic, and PD) based on 10 m × 10 m subplots of epiphytic species samples. Species diversity was estimated using the Simpson index [27] in the R package “vegan.” Functional richness (FRic) was calculated with the R function “dbFD” [28]. Eight functional traits were used to calculate FRic because studies have demonstrated that considering only a single trait can lead to an oversimplification of results [29]. The species evolutionary differences in the community were described by PD [30], which was calculated using the phylogenetic tree of the studied epiphytic bryophyte community and the “picante” package in R.
Given the large number and complex data types of light variables and bark physical and chemical properties variables, principal component analysis (PCA) was applied to light variables and BPP variables and thus reduced secondary variables [31]. In the PCA of the light factor, the first four main components (PC1, PC2, PC3, and PC4) captured 99.7% of the total variations in these variables. The first four PCs (largely reflecting variables in parentheses) were as follows: PC1 (CC, LAI, and ALA), 66.7%; PC2 (CC), 30.3%; PC3 (TT, SRUC, and ALA), 2.2%. In the PCA of BPP factors, the first four main components (PC1, PC2, PC3, and PC4) captured 82.6% of the total variations in these variables. The first four PCs (largely reflecting variables in parentheses) were as follows: PC1 (N, P, S, and Si), 36.7%; PC2 (K, S, pH, and CD), 20.0%; PC3 (N, C, and Si), 13.7%; and PC4 (P, S, and pH), 12.3% [31].
Canonical correspondence analysis (CCA) was used to elucidate relationships between environmental factors and epiphytic bryophyte communities. We also used forward selection to reduce the number of explanatory variables in search of a parsimonious model. The significance of each environmental variable was tested in R using the envfit function. The CCA results were tested using Monte Carlo permutation to determine whether the model reached a significant level (p < 0.05). A CCA analysis and significance test were implemented using the “vegan” package in R [31].
To determine the correlation between environmental factors on the studied epiphytic bryophyte community, the Mantel test was used [32]. The Mantel test assessed the correlation of environmental factors (Topography, LightPC1–PC4, and BPPPC1–PC4) with the diversity of the epiphytic bryophyte community (species diversity, PD, and FRic), respectively. They were represented by the package “ggcor” in R.
To further explore the effects of environmental factors on the diversity of epiphytic bryophyte, linear regression analyses were conducted to test the correlations of different environmental factors with the epiphytic bryophyte species diversity index (Simpson), PD index, and FRic. Variance partitioning [33] based on the “packfor” and “vegan” software packages were used to assess the relative importance of three sets of environmental variables on epiphytic bryophyte species diversity index (Simpson), PD, and FRic, respectively.
Piecewise structural equation models (“piecewiseSEM” package) were used to assess the direct and indirect links between environmental factors and the studied epiphytic bryophyte community. SEM is extensively used in multivariable-related studies due to its ability to partition the causal influences among multiple variables and separate the direct and indirect effects of model predictors [34]. These analyses were conducted using “piecewiseSEM” [35]. The models were modified stepwise according to the pathway significance (p < 0.05) and the goodness of the model (0 ≤ Fisher’s C/df ≤ 2 and 0.05 < p ≤ 1.00) [36].
3. Results
3.1. Composition of the Epiphytic Bryophyte Community
The total number of epiphytic bryophytes records, from the 1310 trees in the plot, was 1650. A total of 25 species were recorded. The species abundance per host tree ranged from 2 to 697. The abundance of epiphytic bryophytes ranged from 5 to 353. The values for the subgroups are given in Table S2 (Additional file). In the studied epiphytic bryophyte community, the dominant species was Pylaisiadelpha yokohamae (Broth.) Buck. In the host tree community, the dominant species was Quercus aliena Bl. var. Acuteserrata Maxim. ex Wenz. The network of interactions between host trees and epiphytic bryophytes was largely symmetric in species richness (Figure 2). The ratio of host tree species-to-epiphytic bryophyte species was 25:24, with a mean of 1.04 bryophyte species per tree species.
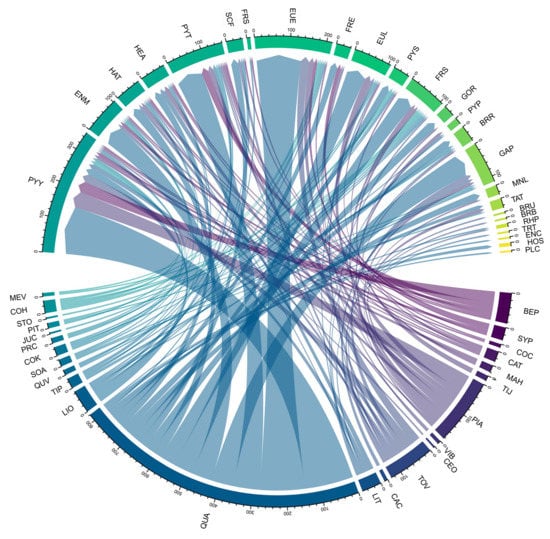
Figure 2.
Correspondence between epiphytic bryophytes and host tree species. The upper part of the figure shows epiphytic bryophyte species, and the lower part of the figure shows host tree species. Lines of the same color connect the epiphytic bryophytes with the host tree species.
3.2. Effect of Environmental Variables on the Epiphytic Bryophyte Community
CCA results showed that the studied epiphytic bryophyte community was primarily affected by Light-PC4 and BPP-PC2 (Figure 3A). The results of the Mantel test showed that the species diversity of epiphytic bryophytes (Simpson) correlated significantly with Light-PC2, and the PD correlated significantly with aspect. Among the environmental factors, the light variables correlated with the topography and BPP variables (Figure 3B).
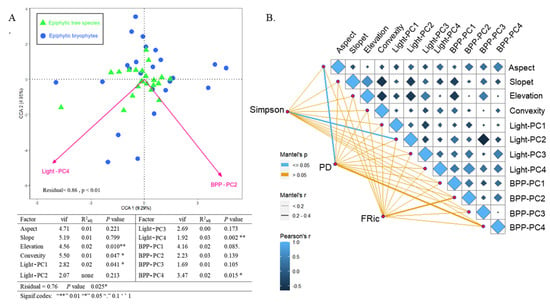
Figure 3.
CCA ordination diagram of the epiphytic bryophytic community with environmental variables. The ranking results were forward selected; the direction of the arrow indicates the value of the environmental parameter, and its length indicates the degree of correlation with the representative axis. The table shows the variance inflation factors (VIFs), the degree of contribution of all environmental factors, and the significance of environmental variables (A). Environmental factors and epiphytic bryophyte diversity are also shown. Pairwise comparisons of environmental factors are shown at the upper-right, with a color gradient representing Pearson’s correlation coefficient. The diversity of epiphytic bryophyte communities was correlated with environmental variables as shown by partial Mantel tests. The line width indicates the partial Mantel r statistic of the corresponding correlation, and the color of the line indicates significance (B).
Across all observations, the species diversity of tree epiphytic bryophytes (Simpson) correlated positively with elevation, while Simpson correlated positively with altitude and negatively with light-PC1 and BPP-PC3. The PD of the tree epiphytic bryophytes correlated negatively with light-PC1 and BPP-PC2. The FRic of the epiphytic bryophytes correlated positively only with light-PC3 (Figure 4).
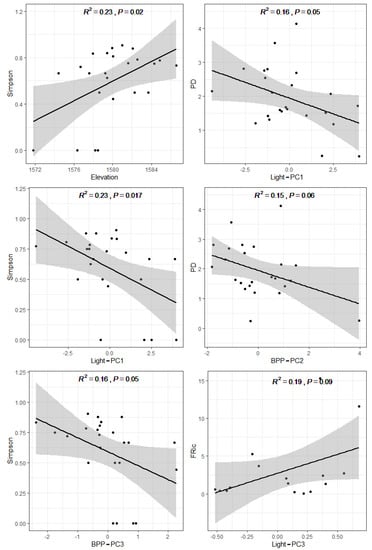
Figure 4.
Linear relationship between diversity indices of epiphytic bryophytes community and environmental factors. The solid line represents the linear regression model with a 95% confidence interval (shaded area).
3.3. The Main Environmental Processes Determining the Coexistence of Bryophyte Communities
In the studied epiphytic bryophyte community, the variations in species diversity (Simpson) and PD were primarily explained by BPP variables (10%) and light variables (16%). Conversely, the variation in FRic was primarily explained by topography (32%) and light (44%) variables (Figure 5).
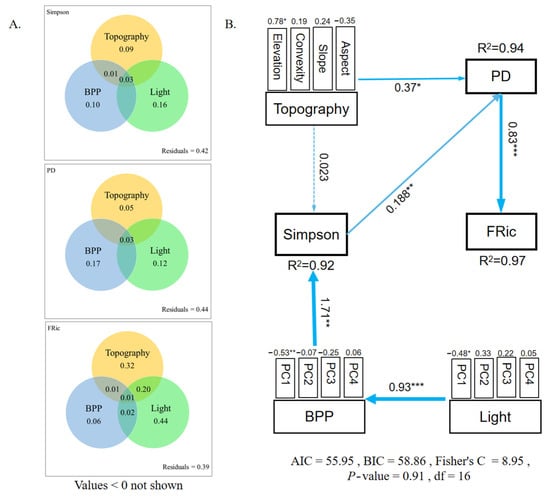
Figure 5.
The contribution of environmental factors to the diversity of epiphytic bryophyte communities. Topography: elevation, aspect, slope, and convexity. Light: Light-PC1-PC4. BPP: BPPPC1-PC4. Among them, yellow = topography; blue = light; green = bark physicochemical properties; yellow + blue = topography+light; yellow + green = topography + BPP; blue + green = light + BPP; yellow + blue + green = topography + light + BPP (A). PiecewiseSEM accounts for the direct and indirect effects of topography variables, light variables, and bark physicochemical property variables on the variation of the diversity of epiphytic bryophyte. The topography, light, and BPP variables were divided into composite variables. Numbers adjacent to measured variables are their coefficients with composite variables. Numbers adjacent to arrows are path coefficients and are the directly standardized effect size of the relationship. Significance levels of each predictor are * p < 0.05, ** p < 0.01, *** p < 0.001. (B).
“PiecewiseSEM” was used to assess the direct and indirect links between environmental factors and the studied epiphytic bryophyte community as well as the relationship among species diversity, PD, and FRic in epiphytic bryophyte communities. In such communities, apart from the direct effects of BPPs on species diversity and topography on PD, the indirect effects of light on species diversity, PD, and FRic were also highly significant. Species diversity was further closely correlated with PD, which was closely correlated with FRic (Figure 5).
4. Discussion
The results of our correlation analyses between the studied epiphytic bryophyte community and environmental factors were consistent with previous studies [6,7,8]. Epiphytic bryophytes species did not show strong positive relationships with light compared with other bryophytes [37]. Bark pH is one of the ecological factors affecting the occurrence of bryophytes [38]. Our results showed a correlation between bark pH [38] and epiphytic bryophyte communities and BPPs (N, P, K, C, S, Si, and bark CD) (Figure 3A). The environmental variables affected the species diversity, PD, and FRic of the epiphyte community.
Previous research has shown that light influences the PD and FRic of vascular plant communities [39]. In the present study, variance-partitioning results showed that light was the main factor driving the diversity variation in epiphytic bryophyte communities. Light drove the variation in species diversity, PD, and FRic (Figure 4) and affected the humidity and temperature in the microenvironment.
Quantification of the effect of BPPs on the Simpson, FRic, and PD (Figure 5A) showed that the BPPs significantly affected the species diversity and PD of epiphytic bryophytes. Studies on vascular plants have also confirmed that physicochemical properties can directly affect the functional traits of plants [40,41]. This finding was similar to a previous one from a study on four tree species, namely, Notholithocarpus densiflorus (Hook. and Arn.) Manos, Cannon and S.H.Oh; Quercus chrysolepis Liebm.; Arbutus menziesii Pursh; and Umbellularia californica (Hook. and Arn.). Their bark characteristics were found to affect epiphytic bryophyte cover across tree species [42]. In temperate forests, the diversity of epiphytic bryophytes is influenced first and foremost by large-scale environmental factors, such as geography and climate [43]. Small-scale environmental factors such as light, temperature, and humidity in the forest also greatly affect bryophyte diversity [42,43,44,45], leading to changes in the distribution pattern of bryophytes in the forest [6,7,8,46,47,48]. BPPs such as bark water content, drying rate, and pH, as well as some chemical properties, influence the epiphytic preference of bryophytes [7]. The results of the present study revealed that the species diversity and PD of epiphytic bryophytes were influenced by bark pH and fissure depth and by some chemical properties of the bark, such as K, P, S, and Si. In a study on epiphytic bryophytes on a broadleaf tree species in the Heishiding Nature Reserve [49], the epiphytic bryophyte bias was most likely correlated with specific bark properties [44]. Bryophytes are very sensitive to the physicochemical properties of epiphytic bark [50,51]. Low bark pH is not conducive to bryophyte growth, and bark CD affects bryophyte attachment [6,8]. Therefore, the BPPs of different tree species have different effects on epiphytic bryophytes and thus on bryophyte diversity. Thus, increasing the diversity of forest tree species and maintaining a certain proportion of specific tree species in the stand are conducive to maintaining the highepiphyte diversity of epiphytic bryophytes [52,53].
“PiecewiseSEM” results showed that FRic, species diversity, and PD depended on and influenced one another in the studied epiphytic bryophyte community. Related studies on microorganism analyses provided evidence that Shannon diversity was positively and strongly correlated with biodiversity components, such as PD and species richness [54].
5. Conclusions
The environmental variables of light, topography, BPPs, all affect the composition of epiphytic bryophyte communities. A range of host trees is essential for maintaining forest epiphyte diversity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15050688/s1: Table 1: Lists of the tree species and the epiphytic bryophytes; Table S1: PCA first four axes of BPP variables; Table S2: List of epiphytic bryophytes and woody plants species in the plot; Table S3: Overview of topographic factor forest canopy light factors in the plot; Table S4: PCA first four axes of light variables; Table S5: Light variables; Table S6: BPP variables; Table S7: List of functional data; Figure S1: Tree epiphytic bryophyte phylogenetic tree.
Author Contributions
Conceptualization, Y.S., S.W. and Y.L.; methodology, Y.S. and Y.L.; software, Y.S. and S.W.; validation, Y.S., Y.C. and Y.L.; formal analysis, Y.S. and S.W.; investigation, Y.S. and Y.C.; resources, Y.S. and Y.L.; data curation, Y.S., Y.C. and H.Z.; writing—original draft preparation, Y.S., S.W. and Y.L.; writing—review and editing, S.W., Y.C. and J.W.; visualization, Y.L., and F.L.; supervision, Z.Y.; project administration, J.W., F.L. and Z.Y.; funding acquisition, H.Z., F.L. and Z.Y. All authors have read and agreed to the published version of the manuscript.
Funding
Project funded by China Postdoctoral Science Foundation (2021M693400); Supported by Natural Science Foundation of Henan (No. 212300410166); Construction of biodiversity survey and monitoring network in the ecologically sensitive area of the Yellow River in Henan Province (HNGD2023018).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article/Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goffnet, B.; Buck, W.R.; Shaw, A.J. Morphology, anatomy, and classification of the Bryophyta. Bryophyt. Biol. 2009, 2, 55–138. [Google Scholar]
- Gao, B.; Li, X.; Zhang, D.; Liang, Y.; Yang, H.; Chen, M.; Zhang, Y.; Zhang, J.; Wood, A.J. Desiccation tolerance in bryophytes: The dehydration and rehydration transcriptomes in the desiccation-tolerant bryophyte Bryum argenteum. Sci. Rep. 2017, 7, 7571. [Google Scholar] [CrossRef] [PubMed]
- Ehbrecht, M.; Seidel, D.; Annighöfer, P.; Kreft, H.; Köhler, M.; Zemp, D.C.; Puettmann, K.; Nilus, R.; Babweteera, F.; Willim, K.; et al. Global patterns and climatic controls of forest structural complexity. Nat. Commun. 2021, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Bowler, D.E.; Nilsen, E.B.; Bischof, R.; O’Hara, R.B.; Yu, T.T.; Oo, T.; Aung, M.; Linnell, J.D. Integrating data from different survey types for population monitoring of an endangered species: The case of the Eld’s deer. Sci. Rep. 2019, 9, 7766. [Google Scholar] [CrossRef]
- Slack, N.G. The Ecological Value of Bryophytes as Indicators of Climate Change. In Bryophyte Ecology and Climate Change; Cambridge University Press: Cambridge, UK, 2011; pp. 3–12. [Google Scholar]
- Kuusinen, M. Epiphyte Flora and Diversity on Basal Trunks of Six Old-Growth Forest Tree Species in Southern and Middle Boreal Finland. Lichenologist 1996, 28, 443–463. [Google Scholar] [CrossRef]
- Slack, N.G. Host specificity of bryophytic epiphytes in Eastern North America. J. Hattori Bot. Lab. 1976, 41, 107–132. [Google Scholar]
- Studlar, S.M. Host Specificity of Epiphytic Bryophytes near Mountain Lake, Virginia. Bryologist 1982, 85, 37–50. [Google Scholar] [CrossRef]
- Yu, S.; Zinati, G.M. Physical and chemical changes in container media in response to bark substitution for peat. Compost. Sci. Util. 2006, 14, 222–230. [Google Scholar] [CrossRef]
- Chemetova, C.; Quilhó, T.; Braga, S.; Fabio, A.M.; Ribeiro, D. Aged Acacia melanoxylon bark as an organic peat replacement in container media. J. Clean. Prod. 2019, 232, 1103–1111. [Google Scholar] [CrossRef]
- Pescador, D.S.; de Bello, F.; Valladares, F.; Escudero, A.; Weigelt, A. Plant Trait Variation along an Altitudinal Gradient in Mediterranean High Mountain Grasslands: Controlling the Species Turnover Effect. PLoS ONE 2015, 10, e0118876. [Google Scholar] [CrossRef]
- William, K.; Cornwell, D.W.; Schwilk, D.D.; Ackerly, A. Trait-Based Test for Habitat Filtering: Convex Hull Volume. Ecology 2006, 87, 1465–1471. [Google Scholar]
- Swenson, N.G.; Enquist, B.J.; Thompson, J.; Zimmerman, J.K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology 2007, 88, 1770–1780. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Renato, V.D.D. Functional traits and niche-based tree community assembly in an Amazonian forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef]
- Valencia, R.; Foster, R.B.; Villa, G.; Condit, R.; Svenning, J.C.; Hernández, C.; Romoleroux, K.; Losos, E.; Magård, E.; Balslev, H. Tree Species Distributions and Local Habitat Variation in the Amazon: Large Forest Plot in Eastern Ecuador. J. Ecol. 2004, 92, 214–229. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Opposing assembly mechanisms in a Neotropical dry forest: Implications for phylogenetic and functional community ecology. Ecology 2009, 90, 2161–2170. [Google Scholar] [CrossRef]
- Uriarte, M.; Swenson, N.G.; Chazdon, R.L.; Comita, L.S.; John Kress, W.; Erickson, D.; Forero-Montaña, J.; Zimmerman, J.K.; Thompson, J. Trait similarity, shared ancestry and the structure of neighbourhood interactions in a subtropical wet forest: Implications for community assembly. Ecol. Lett. 2010, 13, 1503–1514. [Google Scholar] [CrossRef]
- Hoffmann, S.; Beierkuhnlein, C.; Field, R.; Provenzale, A.; Chiarucci, A. Uniqueness of Protected Areas for Conservation Strategies in the European Union. Sci. Rep. 2018, 8, 6445. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L.; Swenson, N.G.; Thompson, J.; Uriarte, M.; Zimmerman, J.K. Advances in the Use of DNA Barcodes to Build a Community Phylogeny for Tropical Trees in a Puerto Rican Forest Dynamics Plot. PLoS ONE 2010, 5, e15409. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Ter Steege, H.; Morgan, H.D.; Van Der Heijden, M.G.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Harms, K.; Richard, E.; Stephen, C.; Hubbell, P.; Robin, B. Foster Habitat Associations of Trees and Shrubs in a 50-Ha Neotropical Forest Plot. J. Ecol. 2001, 89, 947–959. [Google Scholar] [CrossRef]
- Han, B.; Umaña, M.N.; Mi, X.; Liu, X.; Chen, L.; Wang, Y.; Liang, Y.; Wei, W.; Ma, K. The role of transcriptomes linked with responses to light environment on seedling mortality in a subtropical forest, China. J. Ecol. 2017, 105, 592–601. [Google Scholar] [CrossRef]
- Zheng, Y.; Hou, L.; Newell, S.; Liu, M.; Zhou, J.; Zhao, H.; Cheng, X. Community Dynamics and Activity of Ammonia-Oxidizing Prokaryotes in Intertidal Sediments of the Yangtze Estuary. Appl. Environ. Microb. 2014, 80, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Conant, R.T.; Zhou, G.; Wang, Y.; Wang, N.; Li, Y.; Zhang, K. Effects of moso bamboo encroachment into native, broad-leaved forests on soil carbon and nitrogen pools. Sci. Rep. 2016, 6, 31480. [Google Scholar] [CrossRef]
- Lin, Y.; Sun, Z.; Li, Z.; Xue, R.; Cui, W.; Sun, S.; Liu, T.; Zeng, R.; Song, Y. Deficiency in Silicon Transporter Lsi1 Compromises Inducibility of Anti-herbivore Defense in Rice Plants. Front. Plant. Sci. 2019, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. Package ‘FD’. 2014: 1-0. Available online: https://www.imsbio.co.jp/RGM/R_rdfile?f=FD/man/FD-package.Rd&d=R_CC (accessed on 3 April 2022).
- Legras, G.; Loiseau, N.; Gaertner, J.C. Functional richness: Overview of indices and underlying concepts. ACTA Oecol. 2018, 87, 34–44. [Google Scholar] [CrossRef]
- Daniel, P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 10. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0. 2012. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1654-1103.2003.tb02228.x (accessed on 19 April 2022).
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning Beta Diversity in a Subtropical Broad-Leaved Forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef]
- Grace, J.B. Structural Equation Modeling Natural Systems; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Lefcheck, J.S. PiecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Tian, P.; Liu, S.; Zhao, X.; Sun, Z.; Yao, X.; Niu, S.; Crowther, T.W.; Wang, Q. Past climate conditions predict the influence of nitrogen enrichment on the temperature sensitivity of soil respiration. Commun. Earth Env. Environ. 2021, 2, 251. [Google Scholar] [CrossRef]
- Tinya, F.; Márialigeti, S.; Király, I.; Németh, B.; Odor, P. The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant. Ecol. 2009, 204, 69–81. [Google Scholar] [CrossRef]
- Kovářová, M.; Pyszko, P.; Plášek, V. How Does the pH of Tree Bark Change with the Presence of the Epiphytic Bryophytes from the Family Orthotrichaceae in the Interaction with Trunk Inclination? Plants 2022, 11, 63. [Google Scholar] [CrossRef]
- Liu, M.; Che, Y.; Jiao, J.; Li, L.; Jiang, X. Exploring the community phylogenetic structure along the slope aspect of subalpine meadows in the eastern Qinghai–Tibetan Plateau, China. Ecol. Evol. 2019, 9, 5270–5280. [Google Scholar] [CrossRef]
- Win, K.T.; Okazaki, K.; Ookawa, T.; Yokoyama, T.; Ohwaki, Y. Influence of rice-husk biochar and Bacillus pumilus strain TUAT-1 on yield, biomass production, and nutrient uptake in two forage rice genotypes. PLoS ONE 2019, 14, e0220236. [Google Scholar]
- Gil-Martínez, M.; López-García, Á.; Domínguez, M.T.; Navarro-Fernández, C.M.; Kjøller, R.; Tibbett, M.; Marañón, T. Ectomycorrhizal Fungal Communities and Their Functional Traits Mediate Plant–Soil Interactions in Trace Element Contaminated Soils. Front. Plant Sci. 2018, 9, 1682. [Google Scholar] [CrossRef]
- Becker, D.; Dobson, A.; Klitgaard, K. Bark characteristics affect epiphytic bryophyte cover across tree species. In California Ecology and Conservation Research; University of California: Riverside, CA, USA, 2019; pp. 1–8. [Google Scholar]
- Schmitt, C.K.; Slack, N.G. Host specificity of epiphytic lichens and bryophyte: A comparison of the Adirondack Mountains (New York) and the Southern Blue Ridge Mountains (North Carolina). Bryologist 1990, 93, 661–665. [Google Scholar] [CrossRef]
- Trynoski, S.E.; Glime, J.M. Direction and height of bryophytes on four species of Northern Trees. Bryologist 1982, 85, 281–300. [Google Scholar] [CrossRef]
- González-Mancebo, J.M.; Romaguera, F.; Losada-Lima, A.; Suárez, A. Epiphytic bryophytes growing on Laurus azorica (Seub.) Franco in three laurel forest area in Tenerife (CanaryIs lands). ACTA Oecol. 2004, 25, 159–167. [Google Scholar] [CrossRef]
- González-Mancebo, J.M.; Losada-Lima, A.; McAlister, S. Host specificity of epiphytic bryophyte communities of a Laurel forest on Tenerife (Canary Islands, Spain). Bryologist 2003, 106, 383–394. [Google Scholar] [CrossRef]
- Király, I.; Nascimbene, J.; Tinya, F.; Ódor, P. Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers. Conserv. 2013, 22, 209–223. [Google Scholar] [CrossRef]
- Palmer, M.W. Pattern in Corticolous Bryophyte Communities of the North Carolina Piedmont: Do Mosses See the Forest or the Trees? Bryologist 1986, 89, 59–65. [Google Scholar] [CrossRef]
- Weiqiu, L.; Dai, X.; Wang, Y.; Lei, C. Analysis of environmental factors affecting thedistribution of epiphytic bryophyte at Heishiding Nature Reserve, GuangdongProvince. ShengTaiXueBao 2008, 28, 1080–1088. [Google Scholar]
- Turetsky, M.R. The Role of Bryophytes in Carbon and Nitrogen Cycling. Bryologist 2003, 106, 395–409. [Google Scholar] [CrossRef]
- Glime, J.M. Physiological Ecology. Vol.1. In Bryophyte Ecology; Michigan Technological University and the International Association of Bryologists: Houghton, MI, USA, 2007; Available online: http://www.bryoecol.mtu.edu (accessed on 10 April 2022).
- Steele, A. Bryophyte communities of central Idaho forests. Northwest Sci. 1978, 52, 310–322. [Google Scholar]
- Kenkel, N.C.; Bradfield, G.E. Ordination of Epiphytic Bryophyte Communities in a Wet-Temperate Coniferous Forest, South-Coastal British Columbia. Vegetatio 1981, 45, 147–154. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).