
InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data


Abstract
1. Introduction
2. Results
2.1. Identification of Kaluga Sturgeon (H. dauricus) and Hybrids
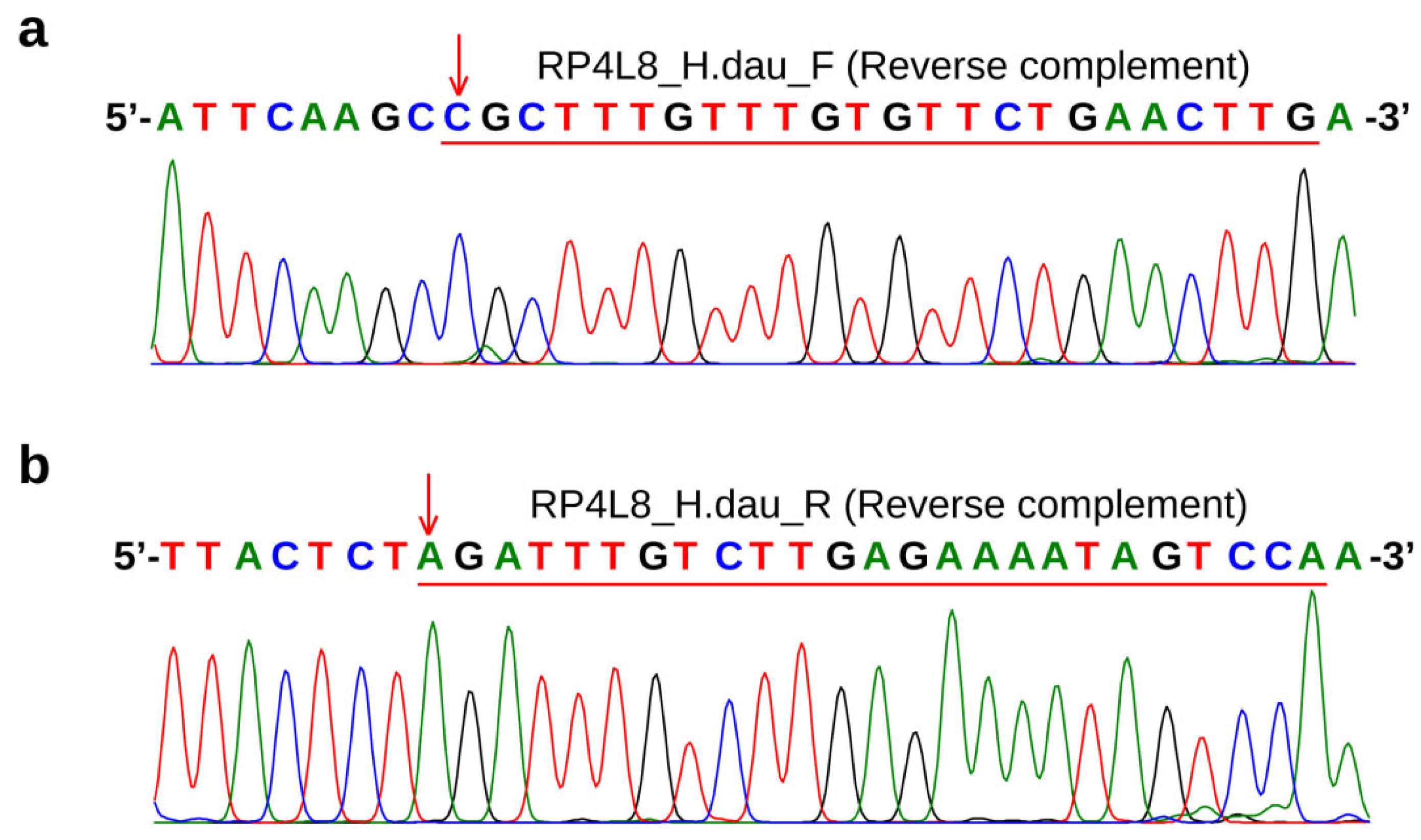
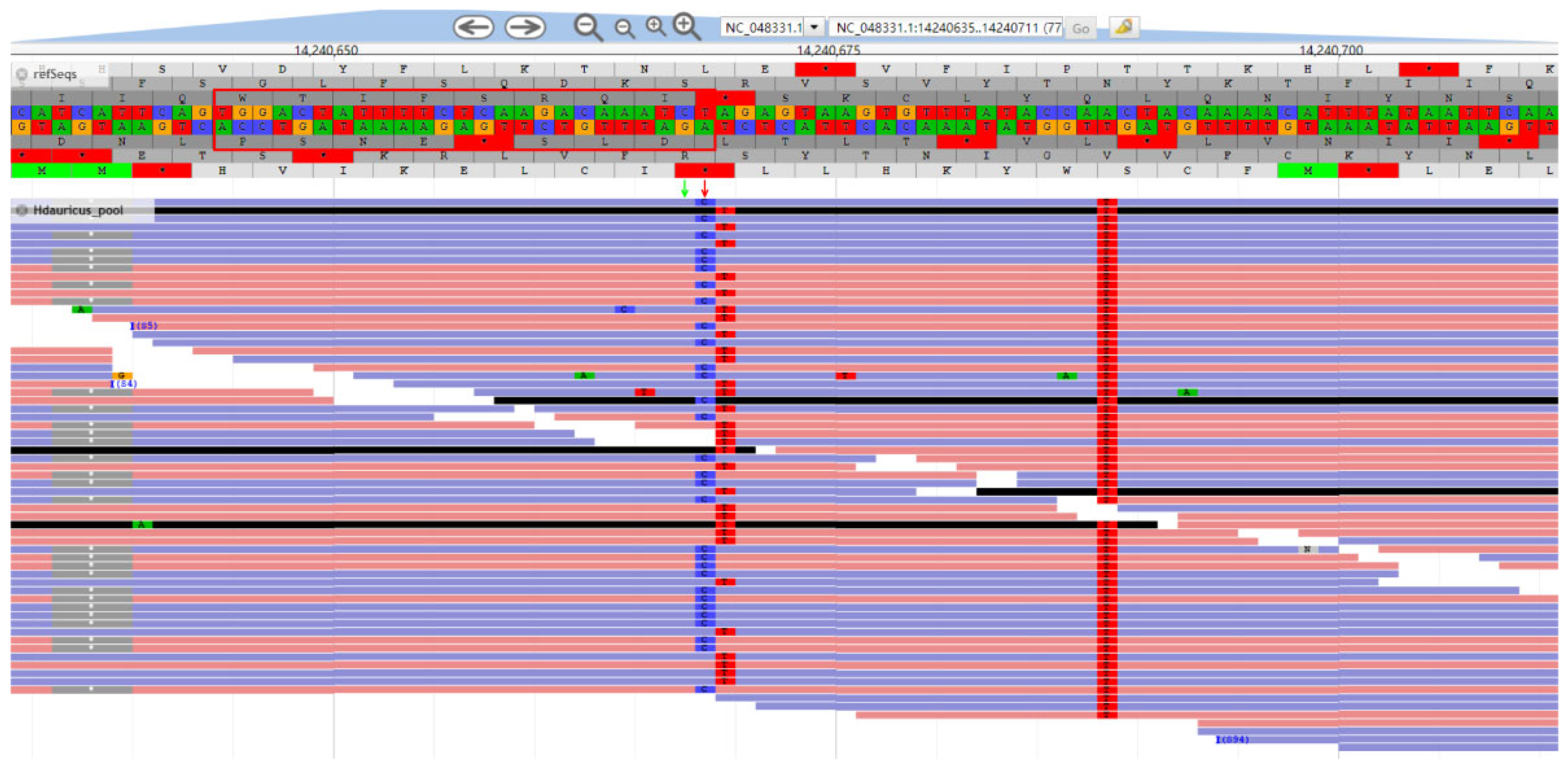
2.2. Specific Variant Identification of Kaluga Sturgeon
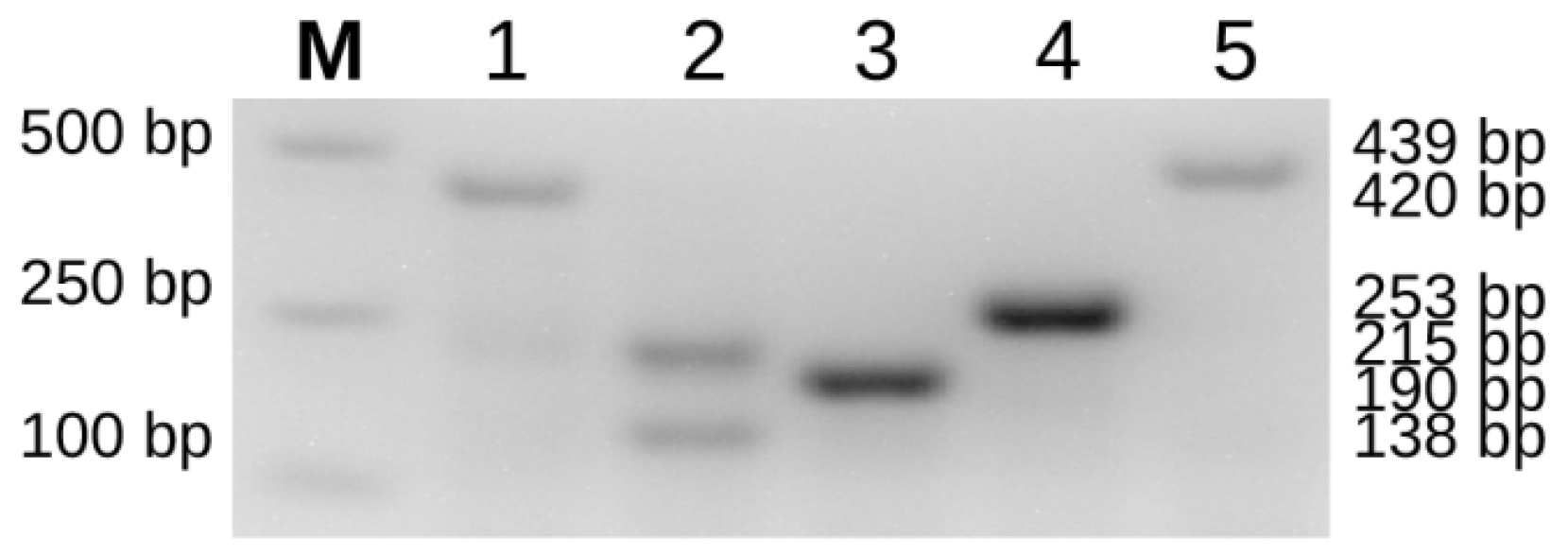
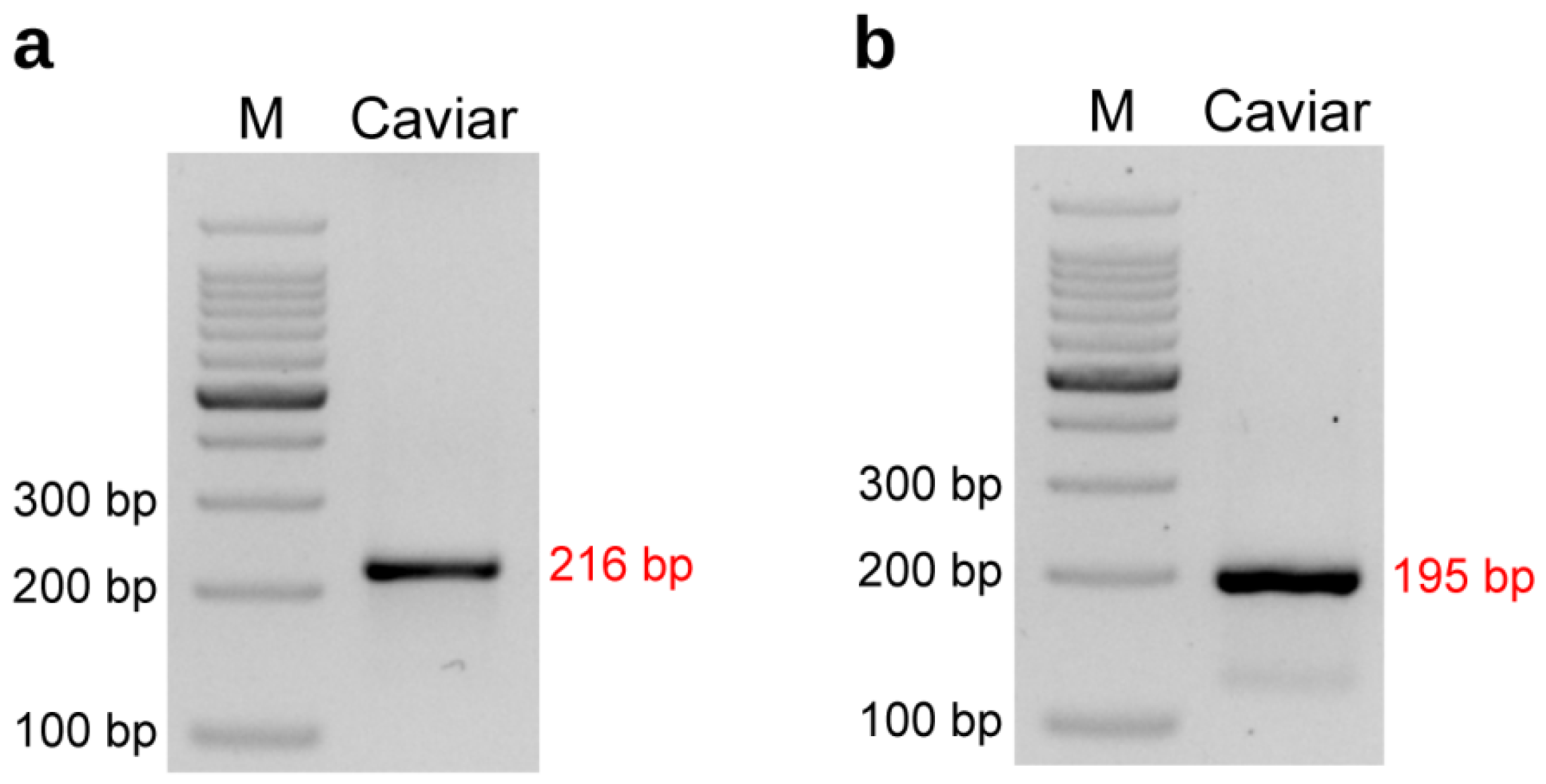
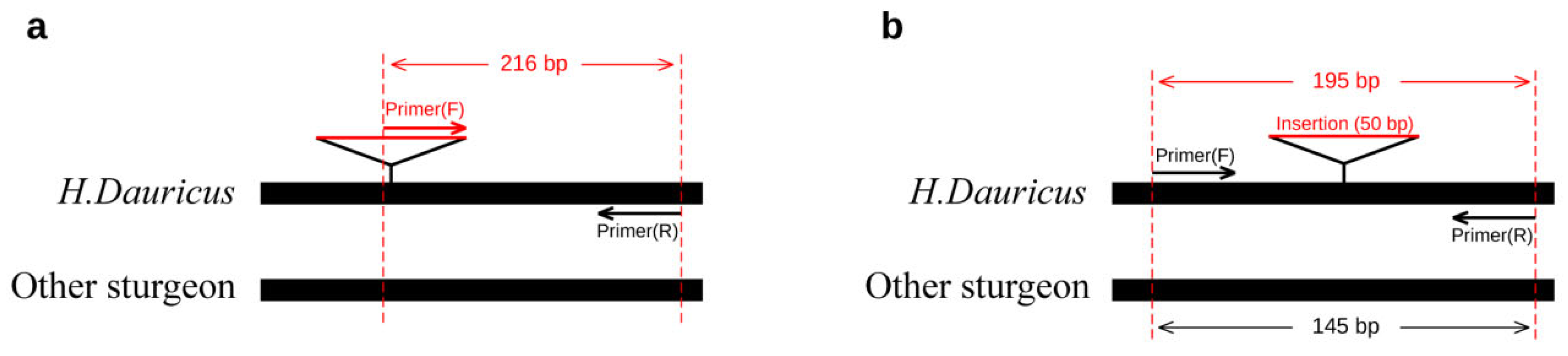
2.3. Dominant/Co-Dominant Primers Design for Kaluga Sturgeon and Validation
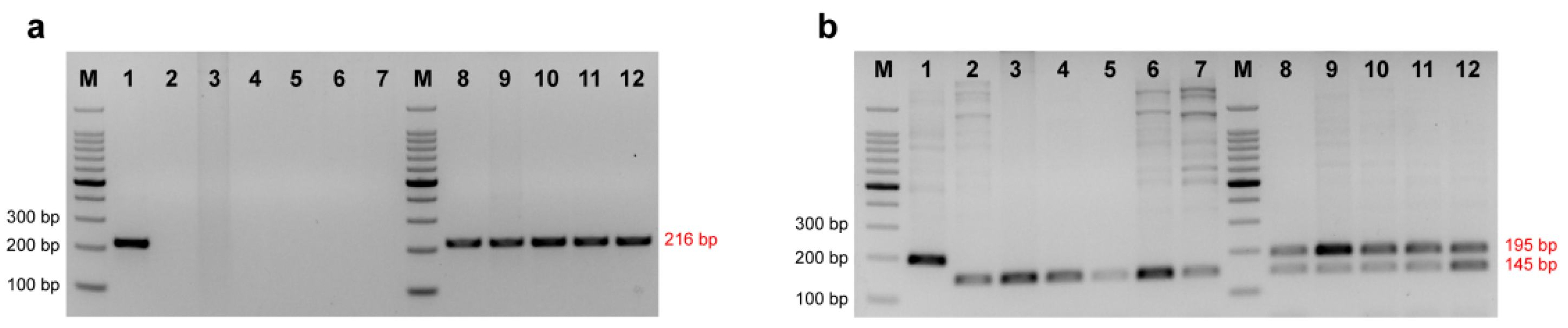
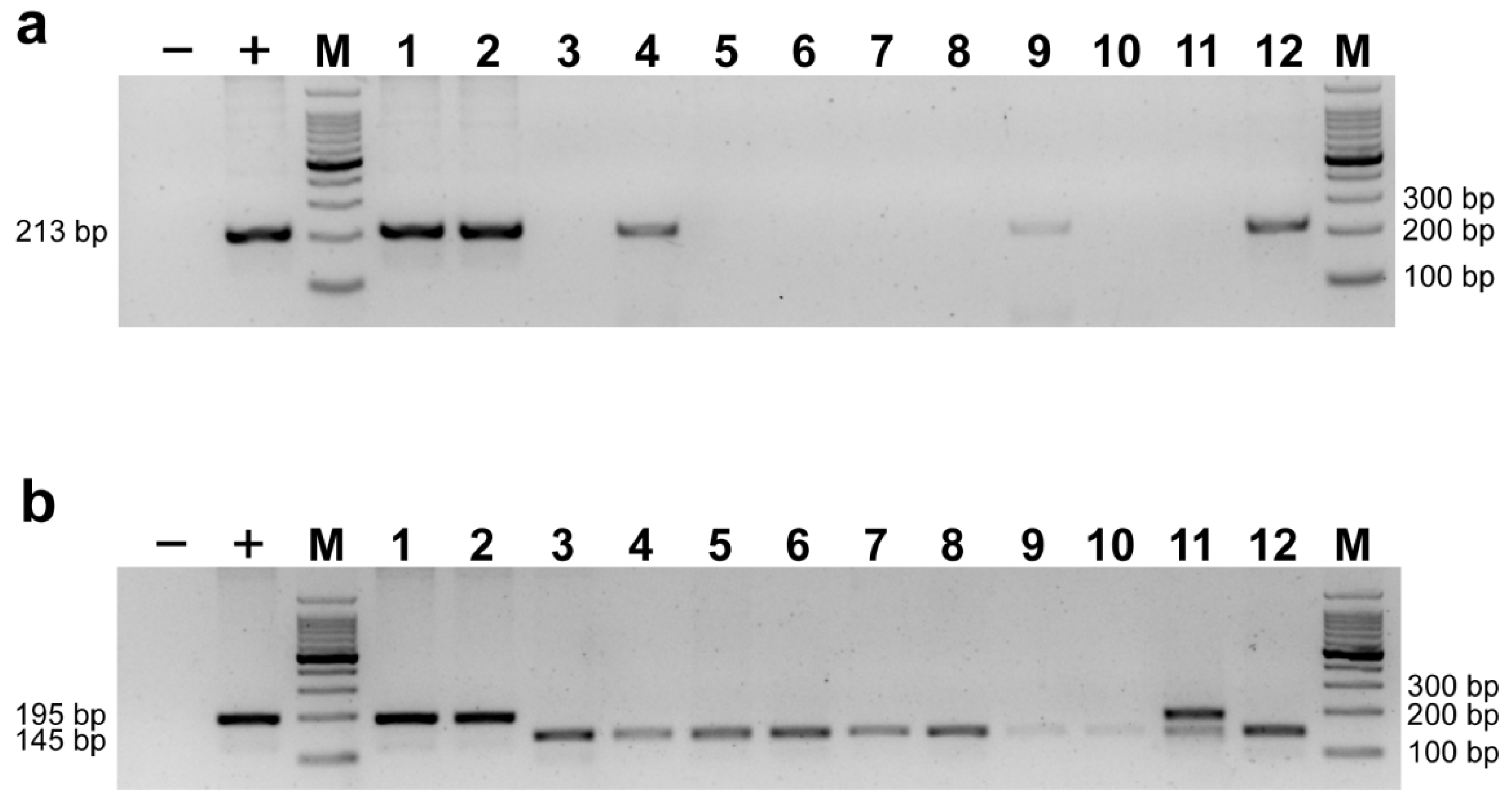
2.4. Validation and Comparison with Other Markers
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling and DNA Extraction
5.2. Whole-Genome Resequencing and Variant Identification
5.3. Primer Design and Validation
5.4. Species-Specific Marker RP4L8 PCR Validation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bemis, W.E.; Findeis, E.K.; Grande, L. An overview of Acipenseriformes. Environ. Biol. Fishes 1997, 48, 25–71. [Google Scholar] [CrossRef]
- International Union for Conservation of Nature. The IUCN Red List of Threatened Species, Version 2022-1. 2022. Available online: www.iucnredlist.org (accessed on 21 July 2022).
- Convention on International Trade in Endangered Species of Wild Fauna and Flora. The tenth meeting of the Conference of the Parties to CITES (COP10), Harare, Zimbabwe. 9-20 June 1997. Available online: https://www.govinfo.gov/content/pkg/FR-1997-08-22/html/97-22402.htm (accessed on 21 July 2022).
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef]
- Shivaramu, S.; Vuong, D.T.; Havelka, M.; Šachlová, H.; Lebeda, I.; Kašpar, V.; Flajšhans, M. Influence of interspecific hybridization on fitness-related traits in Siberian sturgeon and Russian sturgeon. Czech J. Anim. Sci. 2019, 64, 78–88. [Google Scholar] [CrossRef]
- Havelka, M.; Arai, K. Hybridization and polyploidization in sturgeon. Sex Control. Aquac. 2018, Chapter 34, 669–687. [Google Scholar]
- Ludwig, A.; May, B.; Debus, L.; Jenneckens, I. Heteroplasmy in the mtDNA control region of sturgeon (Acipenser, Huso and Scaphirhynchus). Genetics 2000, 156, 1933–1947. [Google Scholar] [CrossRef]
- Ludwig, A.; Debus, L.; Jenneckens, I. A molecular approach to control the international trade in black caviar. Int. Rev. Hydrobiol. A J. Cover. All Asp. Limnol. Mar. Biol. 2002, 87, 661–674. [Google Scholar]
- Mugue, N.S.; Barmintseva, A.E.; Rastorguev, S.M.; Mugue, V.N.; Barmintsev, V.A. Polymorphism of the mitochondrial DNA control region in eight sturgeon species and development of a system for DNA-based species identification. Russ. J. Genet. 2008, 44, 793–798. [Google Scholar] [CrossRef]
- Havelka, M.; Arai, K.; Boscari, E.; Congiu, L.; Sergeev, A.; Mugue, N. A new marker, isolated by ddRAD sequencing, detects Siberian and Russian sturgeon in hybrids. Anim. Genet. 2019, 50, 115–116. [Google Scholar] [CrossRef] [PubMed]
- Havelka, M.; Fujimoto, T.; Hagihara, S.; Adachi, S.; Arai, K. Nuclear DNA markers for identification of Beluga and Sterlet sturgeons and their interspecific Bester hybrid. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Boscari, E.; Barmintseva, A.; Zhang, S.; Yue, H.; Li, C.; Shedko, S.V.; Lieckfeldt, D.; Ludwig, A.; Wei, Q.W.; Mugue, N.S.; et al. Genetic identification of the caviar-producing Amur and Kaluga sturgeons revealed a high level of concealed hybridization. Food Control. 2017, 82, 243–250. [Google Scholar] [CrossRef]
- Yuan, H.; Yang, W.; Zou, J.; Cheng, M.; Fan, F.; Liang, T.; Yu, Y.; Qiu, R.; Li, S.; Hu, J. InDel Markers Based on 3K Whole-Genome Re-Sequencing Data Characterise the Subspecies of Rice (Oryza sativa L.). Agriculture 2021, 11, 655. [Google Scholar] [CrossRef]
- Omoto, N.; Maebayashi, M.; Hara, A.; Adachi, S.; Yamauchi, K. Gonadal maturity in wild sturgeons, Huso dauricus, Acipenser mikadoi and A. schrenckii caught near Hokkaido, Japan. Environ. Biol. Fishes 2004, 70, 381–391. [Google Scholar] [CrossRef]
- Shedko, S.V.; Miroshnichenko, I.L.; Nemkova, G.A. Asymmetric Hybridization of Kaluga Acipenser dauricus Georgi, 1775 and Amur Sturgeon A. schrenckii Brandt, 1869 (Acipenseridae) in Nature as Follows from Analysis of Mitochondrial and Nuclear DNA Markers. Russ. J. Genet. 2020, 56, 718–724. [Google Scholar] [CrossRef]
- Du, K.; Stöck, M.; Kneitz, S.; Klopp, C.; Woltering, J.M.; Adolfi, M.C.; Feron, R.; Prokopov, D.; Makunin, A.; Kichigin, I.; et al. The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization. Nat. Ecol. Evol. 2020, 4, 841–852. [Google Scholar] [CrossRef]
- Barmintseva, A.E.; Mugue, N.S. The use of microsatellite loci for identification of sturgeon species (Acipenseridae) and hybrid forms. Russ. J. Genet. 2013, 49, 950–961. [Google Scholar] [CrossRef]
- Taboada, X.; Robledo, D.; Bouza, C.; Piferrer, F.; Viñas, A.M.; Martínez, P. Reproduction and sex control in turbot. Sex Control Aquac. 2018, Chapter 28, 565–582. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Buels, R.; Yao, E.; Diesh, C.M.; Hayes, R.D.; Munoz-Torres, M.; Helt, G.; Goodstein, D.M.; Elsik, C.G.; Lewis, S.E.; Stein, L.; et al. JBrowse: A dynamic web platform for genome visualization and analysis. Genome Biol. 2016, 17, 66. [Google Scholar] [CrossRef] [PubMed]
- Lalitha, S. Primer premier 5. Biotech Softw. Internet Rep. Comput. Softw. J. Sci. 2000, 1, 270–272. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Pair | Sequence (5′-3′) | Target Band (bp) | |
|---|---|---|---|
| Hdau216p | Forward | CCCGGGGCACAAACATATC | 216 (Kaluga-Specific) |
| Reverse | CAAACAACAAGGAACATTGATACTG | ||
| Hdau195l | Forward | AAACATTTGTCACTTAGGGTTGCCA | 195 (Kaluga-Specific) |
| Reverse | TAACTCGTGCTCATTGTACRGGAA | 145 (Other sturgeon) | |
| Primer Pair | Target Band (bp) | Purebred Specimens | ||||||
|---|---|---|---|---|---|---|---|---|
| H. dauricus | A. schrenckii | A. ruthenus | A. baerii | A. gueldenstaedtii | A. stellatus | H. huso | ||
| Hdau216p | 216 (Kaluga-Specific) | 78/78 | 0/53 | 0/32 | 0/41 | 0/37 | 0/5 | 0/5 |
| Hdau195l | 195 (Kaluga-Specific) | 78/78 | 0/53 | 0/32 | 0/41 | 0/37 | 0/5 | 0/5 |
| 145 (Other sturgeon) | 0/78 | 53/53 | 32/32 | 41/41 | 37/37 | 5/5 | 5/5 | |
| Primer Pair | Target Band (bp) | Hybrid Specimens (♀ × ♂) | ||||
|---|---|---|---|---|---|---|
| A. gueldenstaedtii × H. dauricus | A. baerii × H. dauricus | A. ruthenus × H. dauricus | A. schrenckii × H. dauricus | H. dauricus × A. schrenckii | ||
| Hdau216p | 216 (Kaluga-Specific) | 12/12 | 7/7 | 5/5 | 12/12 | 22/22 |
| Hdau195l | 195 (Kaluga-Specific) | 12/12 | 7/7 | 5/5 | 12/12 | 22/22 |
| 145 (Other sturgeon) | 12/12 | 7/7 | 5/5 | 12/12 | 22/22 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, X.; Dong, Y.; Dong, T.; Song, H.; Wang, W.; Hu, H. InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data. Diversity 2023, 15, 689. https://doi.org/10.3390/d15050689
Yan X, Dong Y, Dong T, Song H, Wang W, Hu H. InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data. Diversity. 2023; 15(5):689. https://doi.org/10.3390/d15050689
Chicago/Turabian StyleYan, Xiaoyu, Ying Dong, Tian Dong, Hailiang Song, Wei Wang, and Hongxia Hu. 2023. "InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data" Diversity 15, no. 5: 689. https://doi.org/10.3390/d15050689
APA StyleYan, X., Dong, Y., Dong, T., Song, H., Wang, W., & Hu, H. (2023). InDel DNA Markers Potentially Unique to Kaluga Sturgeon Huso dauricus Based on Whole-Genome Resequencing Data. Diversity, 15(5), 689. https://doi.org/10.3390/d15050689