

From Source to Tap: Tracking Microbial Diversity in a Riverbank Filtration-Based Drinking Water Supply System under Changing Hydrological Regimes

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
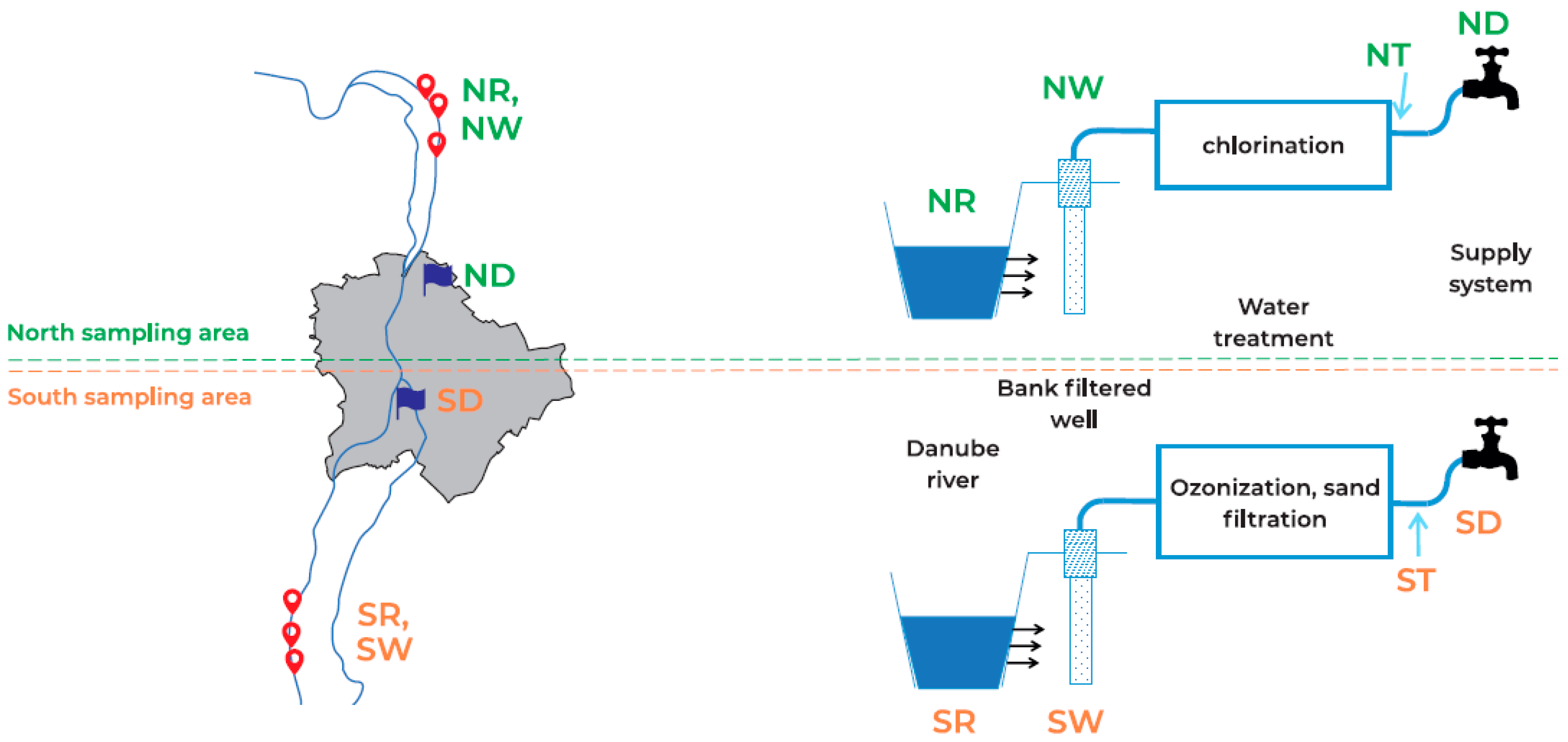
2.1. Study Site and Sampling
2.2. Chemical and Classic Microbiological Analysis
2.3. Direct Cell Count Detection
2.4. Concentration, DNA Extraction, and Illumina Sequencing
2.5. Statistical Analyses
3. Results
3.1. Physicochemical Characteristics and Microbiological Indicators
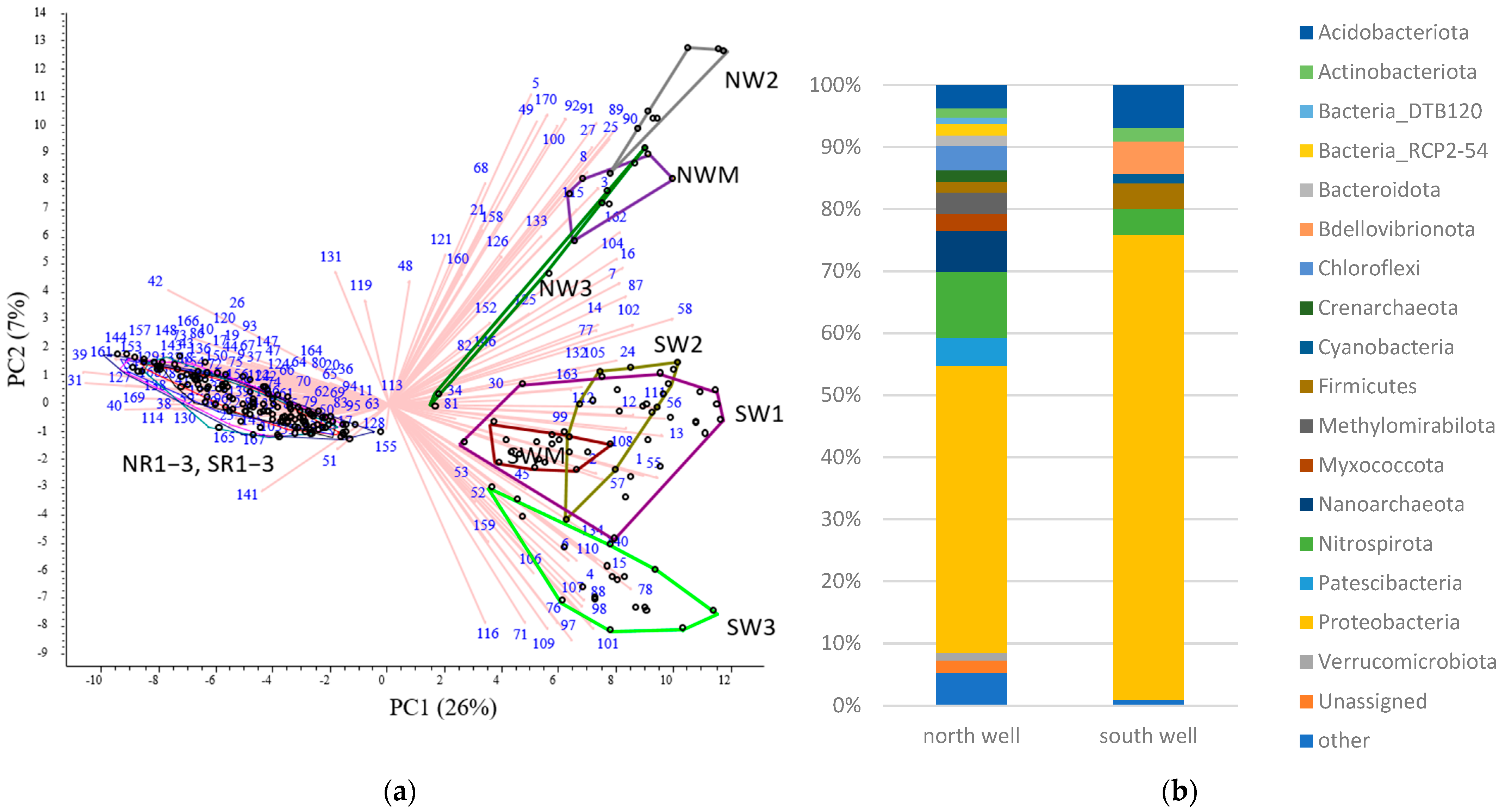
3.2. Microbial Community Diversity
3.3. Impact of Riverbank Filtration on Microbial Communities in the Upstream and Downstream Sites
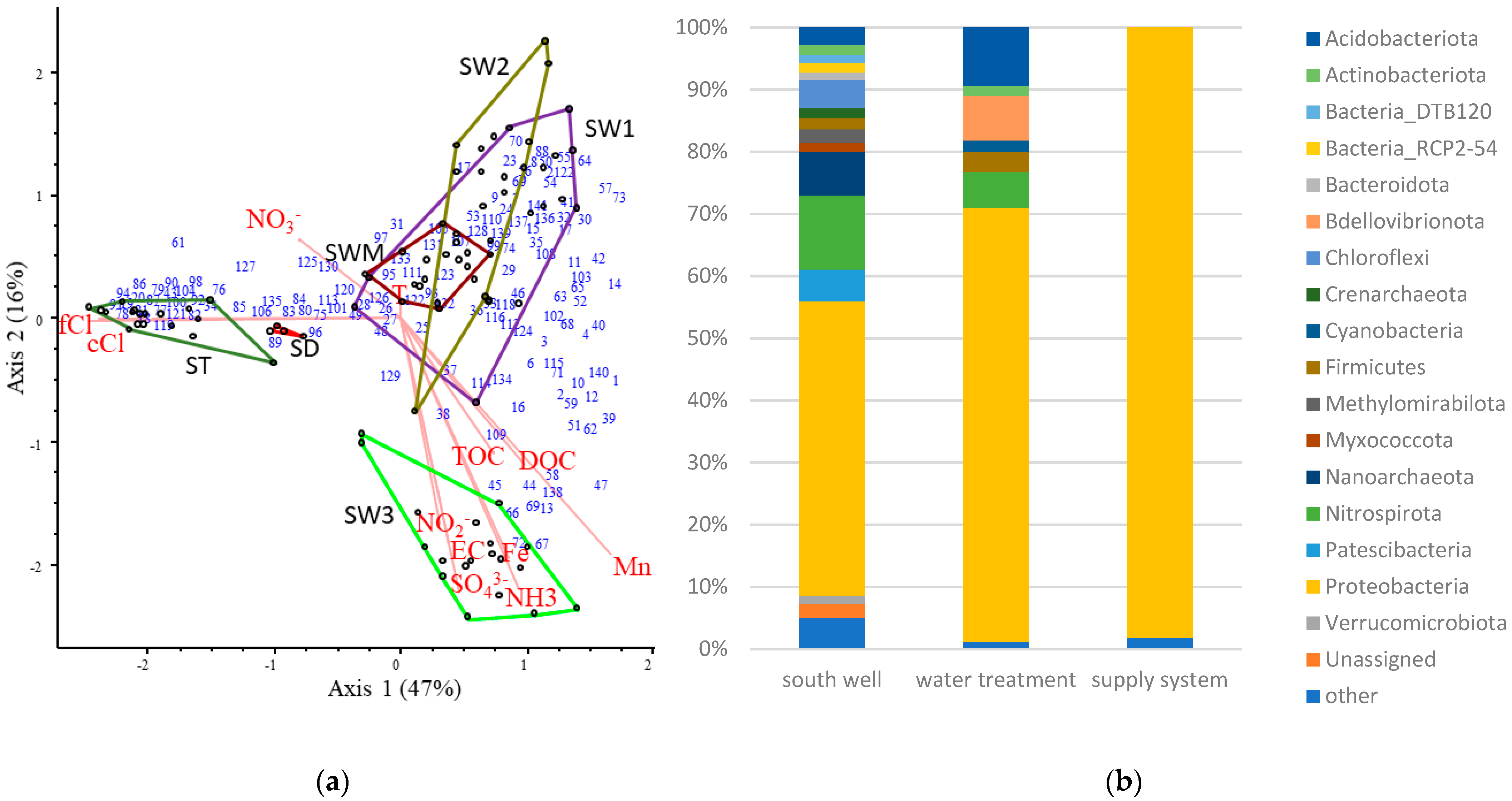
3.4. Impact of Water Treatment and Distribution
3.5. Hydrological Conditions and Seasonality
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoppe-Jones, C.; Oldham, G.; Drewes, J.E. Attenuation of Total Organic Carbon and Unregulated Trace Organic Chemicals in U.S. Riverbank Filtration Systems. Water Res. 2010, 44, 4643–4659. [Google Scholar] [CrossRef]
- Stegen, J.C.; Fredrickson, J.K.; Wilkins, M.J.; Konopka, A.E.; Nelson, W.C.; Arntzen, E.V.; Chrisler, W.B.; Chu, R.K.; Danczak, R.E.; Fansler, S.J.; et al. Groundwater–Surface Water Mixing Shifts Ecological Assembly Processes and Stimulates Organic Carbon Turnover. Nat. Commun. 2016, 7, 11237. [Google Scholar] [CrossRef]
- Wang, H.; Knabe, D.; Engelhardt, I.; Droste, B.; Rohns, H.-P.; Stumpp, C.; Ho, J.; Griebler, C. Dynamics of Pathogens and Fecal Indicators during Riverbank Filtration in Times of High and Low River Levels. Water Res. 2022, 209, 117961. [Google Scholar] [CrossRef]
- Bartak, R.; Page, D.; Sandhu, C.; Grischek, T.; Saini, B.; Mehrotra, I.; Jain, C.K.; Ghosh, N.C. Application of Risk-Based Assessment and Management to Riverbank Filtration Sites in India. J. Water Health 2015, 13, 174–189. [Google Scholar] [CrossRef] [PubMed]
- Nagy-Kovács, Z.; Davidesz, J.; Czihat-Mártonné, K.; Till, G.; Fleit, E.; Grischek, T. Water Quality Changes during Riverbank Filtration in Budapest, Hungary. Water 2019, 11, 302. [Google Scholar] [CrossRef]
- Sprenger, C.; Hartog, N.; Hernández, M.; Vilanova, E.; Grützmacher, G.; Scheibler, F.; Hannappel, S. Inventory of Managed Aquifer Recharge Sites in Europe: Historical Development, Current Situation and Perspectives. Hydrogeol. J. 2017, 25, 1909–1922. [Google Scholar] [CrossRef]
- Sprenger, C.; Lorenzen, G.; Hülshoff, I.; Grützmacher, G.; Ronghang, M.; Pekdeger, A. Vulnerability of Bank Filtration Systems to Climate Change. Sci. Total Environ. 2011, 409, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, L.; Hug, K.; Griebler, C. Aquifer Recharge Viewed through the Lens of Microbial Community Ecology: Initial Disturbance Response, and Impacts of Species Sorting versus Mass Effects on Microbial Community Assembly in Groundwater during Riverbank Filtration. Water Res. 2021, 189, 116631. [Google Scholar] [CrossRef]
- Meng, L.; Zuo, R.; Wang, J.; Yang, J.; Li, Q.; Chen, M. The Spatial Variations of Correlation between Microbial Diversity and Groundwater Quality Derived from a Riverbank Filtration Site, Northeast China. Sci. Total Environ. 2020, 706, 135855. [Google Scholar] [CrossRef]
- Zuo, R.; Xue, Z.; Wang, J.; Meng, L.; Zhao, X.; Pan, M.; Cai, W. Spatiotemporal Variations of Redox Conditions and Microbial Responses in a Typical River Bank Filtration System with High Fe2+ and Mn2+ Contents. J. Hydrol. 2022, 609, 127777. [Google Scholar] [CrossRef]
- Lee, J.-H.; Lee, B.-J.; Unno, T. Bacterial Communities in Ground-and Surface Water Mixing Zone Induced by Seasonal Heavy Extraction of Groundwater. Geomicrobiol. J. 2018, 35, 768–774. [Google Scholar] [CrossRef]
- Yang, F.; Yue, S.; Wu, X.; Zhang, C.; Li, D.; Zhu, R. Effects of Flood Inundation on Biogeochemical Processes in Groundwater during Riverbank Filtration. J. Hydrol. 2023, 617, 129101. [Google Scholar] [CrossRef]
- Pinar-Méndez, A.; Wangensteen, O.S.; Præbel, K.; Galofré, B.; Méndez, J.; Blanch, A.R.; García-Aljaro, C. Monitoring Bacterial Community Dynamics in a Drinking Water Treatment Plant: An Integrative Approach Using Metabarcoding and Microbial Indicators in Large Water Volumes. Water 2022, 14, 1435. [Google Scholar] [CrossRef]
- Waak, M.B.; Hozalski, R.M.; Hallé, C.; LaPara, T.M. Comparison of the Microbiomes of Two Drinking Water Distribution Systems—With and without Residual Chloramine Disinfection. Microbiome 2019, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Verberk, J.Q.J.C.; Van Dijk, J.C. Bacteriology of Drinking Water Distribution Systems: An Integral and Multidimensional Review. Appl. Microbiol. Biotechnol. 2013, 97, 9265–9276. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, C.J.; Schönher, C.; Proksch, P.; Kerschbaumer, D.J.; Mayr, E.; Zunabovic-Pichler, M.; Domig, K.J.; Perfler, R. Assessment of Microbial Community Dynamics in River Bank Filtrate Using High-Throughput Sequencing and Flow Cytometry. Front. Microbiol. 2018, 9, 2887. [Google Scholar] [CrossRef]
- Gomez-Alvarez, V.; Siponen, S.; Kauppinen, A.; Hokajärvi, A.-M.; Tiwari, A.; Sarekoski, A.; Miettinen, I.T.; Torvinen, E.; Pitkänen, T. A Comparative Analysis Employing a Gene- and Genome-Centric Metagenomic Approach Reveals Changes in Composition, Function, and Activity in Waterworks with Different Treatment Processes and Source Water in Finland. Water Res. 2023, 229, 119495. [Google Scholar] [CrossRef]
- Han, Z.; An, W.; Yang, M.; Zhang, Y. Assessing the Impact of Source Water on Tap Water Bacterial Communities in 46 Drinking Water Supply Systems in China. Water Res. 2020, 172, 115469. [Google Scholar] [CrossRef]
- Rudolf von Rohr, M.; Hering, J.G.; Kohler, H.-P.E.; von Gunten, U. Column Studies to Assess the Effects of Climate Variables on Redox Processes during Riverbank Filtration. Water Res. 2014, 61, 263–275. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Podani, J. SYN-TAX Computer Programs for Data Analysis in Ecology and Systematics. User’s Manual; Scientia: Budapest, Hungary, 2001. [Google Scholar]
- European Council Directive 98/83/EC of 3 November 1998 on the Quality of Water Intended for Human Consumption. Off. J. Eur. Union 1998, L330, 32–54. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:31998L0083 (accessed on 20 March 2023).
- Zhang, Y.; Oh, S.; Liu, W. Impact of Drinking Water Treatment and Distribution on the Microbiome Continuum: An Ecological Disturbance’s Perspective. Environ. Microbiol. 2017, 19, 3163–3174. [Google Scholar] [CrossRef]
- Bruno, A.; Agostinetto, G.; Fumagalli, S.; Ghisleni, G.; Sandionigi, A. It’s a Long Way to the Tap: Microbiome and DNA-Based Omics at the Core of Drinking Water Quality. Int. J. Environ. Res. Public Health 2022, 19, 7940. [Google Scholar] [CrossRef]
- van Driezum, I.H.; Chik, A.H.S.; Jakwerth, S.; Lindner, G.; Farnleitner, A.H.; Sommer, R.; Blaschke, A.P.; Kirschner, A.K.T. Spatiotemporal Analysis of Bacterial Biomass and Activity to Understand Surface and Groundwater Interactions in a Highly Dynamic Riverbank Filtration System. Sci. Total Environ. 2018, 627, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Zhai, Y.; Teng, Y. Microbial Response to Biogeochemical Profile in a Perpendicular Riverbank Filtration Site. Ecotoxicol. Environ. Saf. 2022, 244, 114070. [Google Scholar] [CrossRef]
- Kojima, H.; Shinohara, A.; Fukui, M. Sulfurifustis Variabilis Gen. Nov., Sp. Nov., a Sulfur Oxidizer Isolated from a Lake, and Proposal of Acidiferrobacteraceae Fam. Nov. and Acidiferrobacterales Ord. Nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 3709–3713. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Herrmann, M.; Kampe, B.; Lehmann, R.; Totsche, K.U.; Küsel, K. Environmental Selection Shapes the Formation of Near-Surface Groundwater Microbiomes. Water Res. 2020, 170, 115341. [Google Scholar] [CrossRef]
- Liu, X.; Li, M.; Castelle, C.J.; Probst, A.J.; Zhou, Z.; Pan, J.; Liu, Y.; Banfield, J.F.; Gu, J.-D. Insights into the Ecology, Evolution, and Metabolism of the Widespread Woesearchaeotal Lineages. Microbiome 2018, 6, 102. [Google Scholar] [CrossRef]
- Oh, S.; Hammes, F.; Liu, W.-T. Metagenomic Characterization of Biofilter Microbial Communities in a Full-Scale Drinking Water Treatment Plant. Water Res. 2018, 128, 278–285. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Lovejoy, C.; Vincent, W.F. Genomic Evidence of Functional Diversity in DPANN Archaea, from Oxic Species to Anoxic Vampiristic Consortia. ISME Commun. 2022, 2, 4. [Google Scholar] [CrossRef]
- Begmatov, S.; Beletsky, A.V.; Dedysh, S.N.; Mardanov, A.V.; Ravin, N.V. Genome Analysis of the Candidate Phylum MBNT15 Bacterium from a Boreal Peatland Predicted Its Respiratory Versatility and Dissimilatory Iron Metabolism. Front. Microbiol. 2022, 13, 951761. [Google Scholar] [CrossRef]
- Baker, B.J.; Lazar, C.S.; Teske, A.P.; Dick, G.J. Genomic Resolution of Linkages in Carbon, Nitrogen, and Sulfur Cycling among Widespread Estuary Sediment Bacteria. Microbiome 2015, 3, 14. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Ning, D.; He, Z.; Zhang, P.; Spencer, S.J.; Gao, S.; Shi, W.; Wu, L.; Zhang, Y.; Yang, Y.; et al. Small and Mighty: Adaptation of Superphylum Patescibacteria to Groundwater Environment Drives Their Genome Simplicity. Microbiome 2020, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Nagy-Kovács, Z.; László, B.; Simon, E.; Fleit, E. Operational Strategies and Adaptation of RBF Well Construction to Cope with Climate Change Effects at Budapest, Hungary. Water 2018, 10, 1751. [Google Scholar] [CrossRef]
- Tiwari, A.; Hokajärvi, A.-M.; Domingo, J.S.; Elk, M.; Jayaprakash, B.; Ryu, H.; Siponen, S.; Vepsäläinen, A.; Kauppinen, A.; Puurunen, O.; et al. Bacterial Diversity and Predicted Enzymatic Function in a Multipurpose Surface Water System—From Wastewater Effluent Discharges to Drinking Water Production. Environ. Microbiome 2021, 16, 11. [Google Scholar] [CrossRef]
- Inkinen, J.; Siponen, S.; Jayaprakash, B.; Tiwari, A.; Hokajärvi, A.-M.; Pursiainen, A.; Ikonen, J.; Kauppinen, A.; Miettinen, I.T.; Paananen, J.; et al. Diverse and Active Archaea Communities Occur in Non-Disinfected Drinking Water Systems–Less Activity Revealed in Disinfected and Hot Water Systems. Water Res. X 2021, 12, 100101. [Google Scholar] [CrossRef]
- Zhou, W.; Li, W.; Chen, J.; Zhou, Y.; Wei, Z.; Gong, L. Microbial Diversity in Full-Scale Water Supply Systems through Sequencing Technology: A Review. RSC Adv. 2021, 11, 25484–25496. [Google Scholar] [CrossRef]
- Choi, Y.; Cha, Y.; Kim, B. Characteristics of Bacterial Communities in Biological Filters of Full-Scale Drinking Water Treatment Plants. J. Microbiol. Biotechnol. 2019, 29, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Fish, K.E.; Boxall, J.B. Biofilm Microbiome (Re)Growth Dynamics in Drinking Water Distribution Systems Are Impacted by Chlorine Concentration. Front. Microbiol. 2018, 9, 2519. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Han, X.; Liu, C.; Bai, X. Intrinsic Chlorine Resistance of Bacteria Modulated by Glutaminyl-TRNA Biosynthesis in Drinking Water Supply Systems. Chemosphere 2022, 308, 136322. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, W.; Zhang, J.; Qi, W.; Li, Y.; Chen, S.; Zhou, W. Prevalence of Antibiotic Resistance Genes in Drinking Water and Biofilms: The Correlation with the Microbial Community and Opportunistic Pathogens. Chemosphere 2020, 259, 127483. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, I.F.; Tunsjø, H.S.; Andreassen, M.; Nielsen, K.M.; Lund, V.; Charnock, C. Detection of Aminoglycoside Resistant Bacteria in Sludge Samples From Norwegian Drinking Water Treatment Plants. Front. Microbiol. 2019, 10, 487. [Google Scholar] [CrossRef]
- Issotta, F.; Moya-Beltrán, A.; Mena, C.; Covarrubias, P.C.; Thyssen, C.; Bellenberg, S.; Sand, W.; Quatrini, R.; Vera, M. Insights into the Biology of Acidophilic Members of the Acidiferrobacteraceae Family Derived from Comparative Genomic Analyses. Res. Microbiol. 2018, 169, 608–617. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
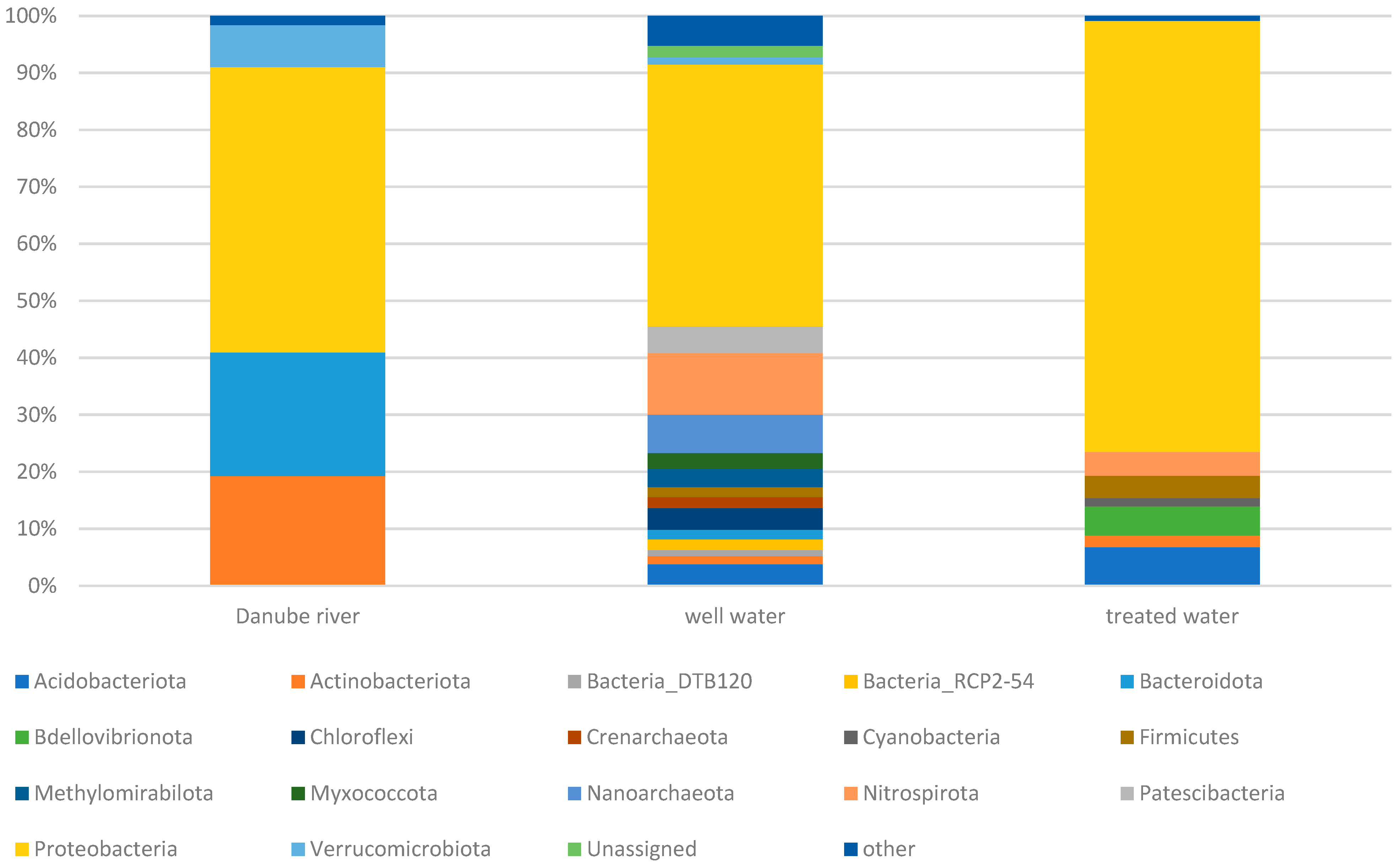{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NR | NW | NT | ND | SR | SW | ST | SD | |
|---|---|---|---|---|---|---|---|---|
| Parameter | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) |
| T (°C) | 11.5 (1.6–23.8) | 11.7 (7.2–14.9) | 12.3 (10.1–15) | 15.4 (7.3–23.6) | 11.5 (1–22.9) | 11.8 (7.2–16.2) | 11.9 (8.6–15.7) | 16.4 (14.6–20) |
| pH | 7.6 (6.5–8.4) | 7.6 (7.3–7.9) | 7.7 (7.4–7.8) | 7.5 (7–7.8) | 7.8 (7.1–8.4) | 7.7 (6.7–7.9) | 7.6 (6.8–7.9) | 7.6 (7.3–7.9) |
| Electric conductivity (µS/cm) | 340 (250–430) | 561 (439–728) | 516 (440–562) | 540 (462–774) | 350 (260–440) | 580 (448–783) | 554 (481–632) | 620 (505–712) |
| Redox potential (mV) | 97 (5–270) | 222 (151–294) | 591 (315–656) | 489 (258–580) | 93 (3.8–218) | 134 (4.5–245) | 178 (128–275) | 222 (157–317) |
| NH3 (mg/L) | 0.1 (<LOD–0.2) | <LOD | <LOD | <LOD | 0.1 (<LOD–0.3) | <LOD (<LOD–0.1) | <LOD (<LOD–0.1) | <LOD (<LOD–0.1) |
| NO2− (mg/L) | <LOD (<LOD–0.1) | <LOD | <LOD | <LOD | <LOD (<LOD–0.1) | <LOD (<LOD–0.1) | <LOD | <LOD |
| NO3− (mg/L) | 6.9 (3.4–12) | 13 (3–29) | 11.1 (4.7–16) | 7.8 (5–12) | 7.0 (3.8–12) | 3.5 (0.6–16) | 4.0 (1.8–7.7) | 9.5 (3.7–16) |
| Cl− (mg/L) | 18 (10–35) | 25 (15–38) | 23 (16–28) | 21 (15–31) | 19 (11–36) | 23 (15–32) | 22 (16–29) | 28 (19–35) |
| SO42− (mg/L) | 28 (17–38) | 48 (23–91) | 39 (25–49) | 35 (24–46) | 29 (18–39) | 61 (30–150) | 51 (31–68) | 68 (37–89) |
| Turbidity (NTU) | 27.7 (3.5–170) | 0.1 (0.1–1.3) | 0.3 (0.1–3.4) | 0.1 (0.1–0.8) | 27.6 (4–230) | 2.3 (0.1–12) | 0.2 (0.1–1) | 0.4 (0.1–3.4) |
| TOC (mg/L) | 2.6 (1.3–4.1) | 1.1 (0.6–2) | 1.3 (0.8–2.8) | 1.4 (0.9–2.3) | 2.7 (1.5–4.5) | 1.6 (1–2.6) | 1.4 (1.1–2.4) | 1.3 (1–2.2) |
| DOC (mg/L) | 2.4 (1.4–3.7) | 1.1 (0.6–1.9) | 1.2 (0.8–1.9) | 1.3 (0.9–1.8) | 2.5 (1.5–3.9) | 1.5 (1–2.2) | 1.3 (1–1.7) | 1.2 (0.9–1.6) |
| Fe (µg/L) | 347 (29–3347) | 20 (7.1–190) | 19 (7.1–110) | 15 (7.1–27) | 363 (12–2900) | 362 (14.1–2200) | 27 (4.6–140) | 108 (47–750) |
| Mn (µg/L) | 28 (3.5–234) | 9.1 (0.1–79) | 13.5 (0.1–180) | 4.8 (0.1–15) | 34.5 (3.5–210) | 297 (20–650) | 6.1 (3.5–22) | 21 (3.5–110) |
| Combined chlorine (mg/L) | NA | NA | 0.1 (<LOD–0.6) | 0.1 (<LOD–0.2) | NA | NA | <LOD (<LOD–0.1) | 0.1 (0.1–0.2) |
| Free chlorine (mg/L) | NA | NA | 0.3 (0.1–0.5) | 0.2 (<LOD–0.6) | NA | NA | <LOD (<LOD–0.1) | 0.1 (<LOD–0.2) |
| Colony count 22 °C | 3700 (100–28,000) | 120 (0–3800) | 160 (0–4300) | 36 (0–450) | 6600 (375–67,000) | 220 (0–4500) | 170 (0–2900) | 160 (0–2800) |
| E. coli | 149 (5–1720) | 0 (0–0) | 0 (0–0) | 0 (0–0) | 420 (5–1920) | 0 (0–0) | 0 (0–0) | 0 (0–0) |
| Enterococci | 43 (0–400) | 0 (0–1) 1 | 0 (0–0) | 0 (0–0) | 95 (1–500) | 0 (0–0) | 0 (0–1) 1 | 0 (0–1) 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargha, M.; Róka, E.; Erdélyi, N.; Németh, K.; Nagy-Kovács, Z.; Kós, P.B.; Engloner, A.I. From Source to Tap: Tracking Microbial Diversity in a Riverbank Filtration-Based Drinking Water Supply System under Changing Hydrological Regimes. Diversity 2023, 15, 621. https://doi.org/10.3390/d15050621
Vargha M, Róka E, Erdélyi N, Németh K, Nagy-Kovács Z, Kós PB, Engloner AI. From Source to Tap: Tracking Microbial Diversity in a Riverbank Filtration-Based Drinking Water Supply System under Changing Hydrological Regimes. Diversity. 2023; 15(5):621. https://doi.org/10.3390/d15050621
Chicago/Turabian StyleVargha, Márta, Eszter Róka, Norbert Erdélyi, Kitti Németh, Zsuzsanna Nagy-Kovács, Péter B. Kós, and Attila I. Engloner. 2023. "From Source to Tap: Tracking Microbial Diversity in a Riverbank Filtration-Based Drinking Water Supply System under Changing Hydrological Regimes" Diversity 15, no. 5: 621. https://doi.org/10.3390/d15050621
APA StyleVargha, M., Róka, E., Erdélyi, N., Németh, K., Nagy-Kovács, Z., Kós, P. B., & Engloner, A. I. (2023). From Source to Tap: Tracking Microbial Diversity in a Riverbank Filtration-Based Drinking Water Supply System under Changing Hydrological Regimes. Diversity, 15(5), 621. https://doi.org/10.3390/d15050621