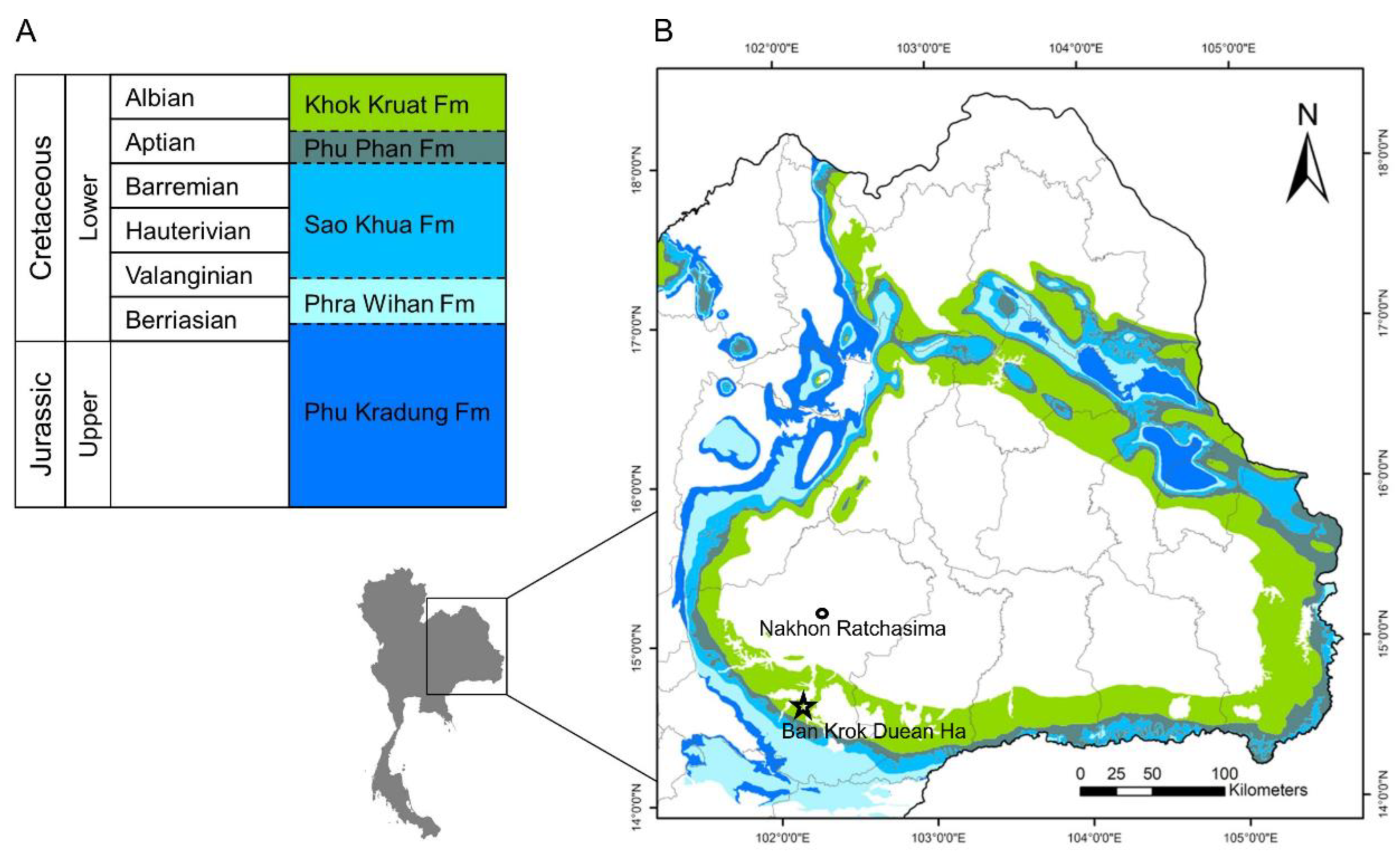
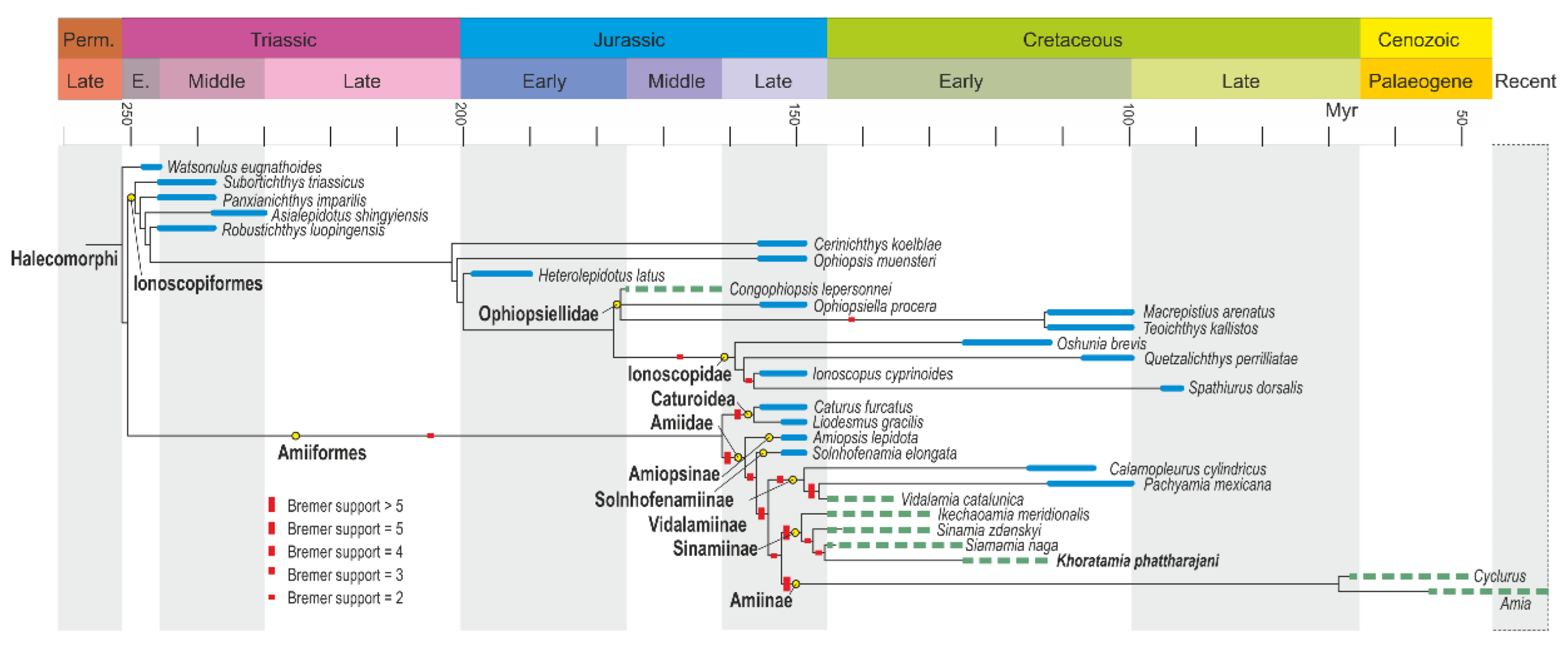
3.2.2. Cranium
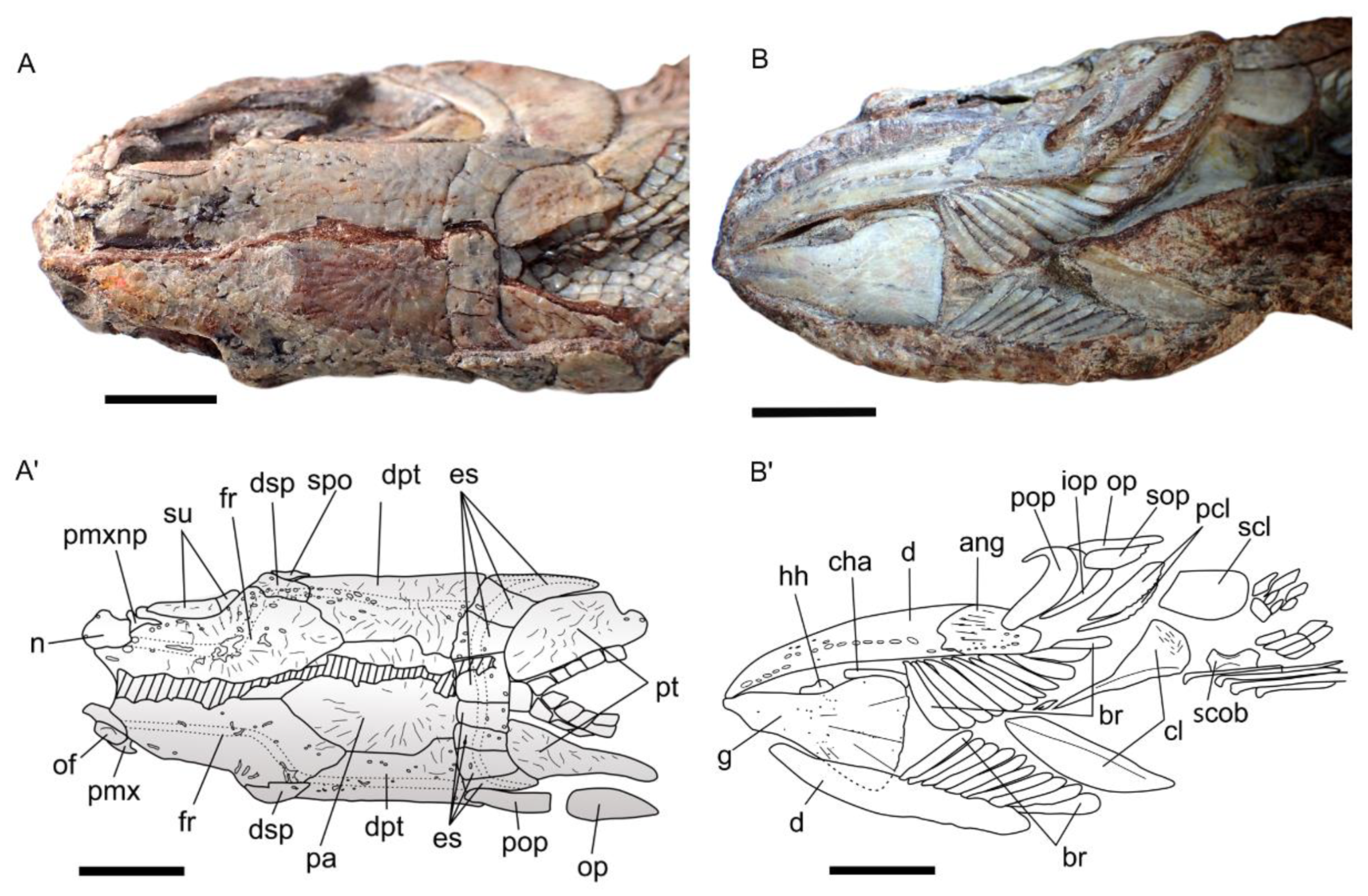
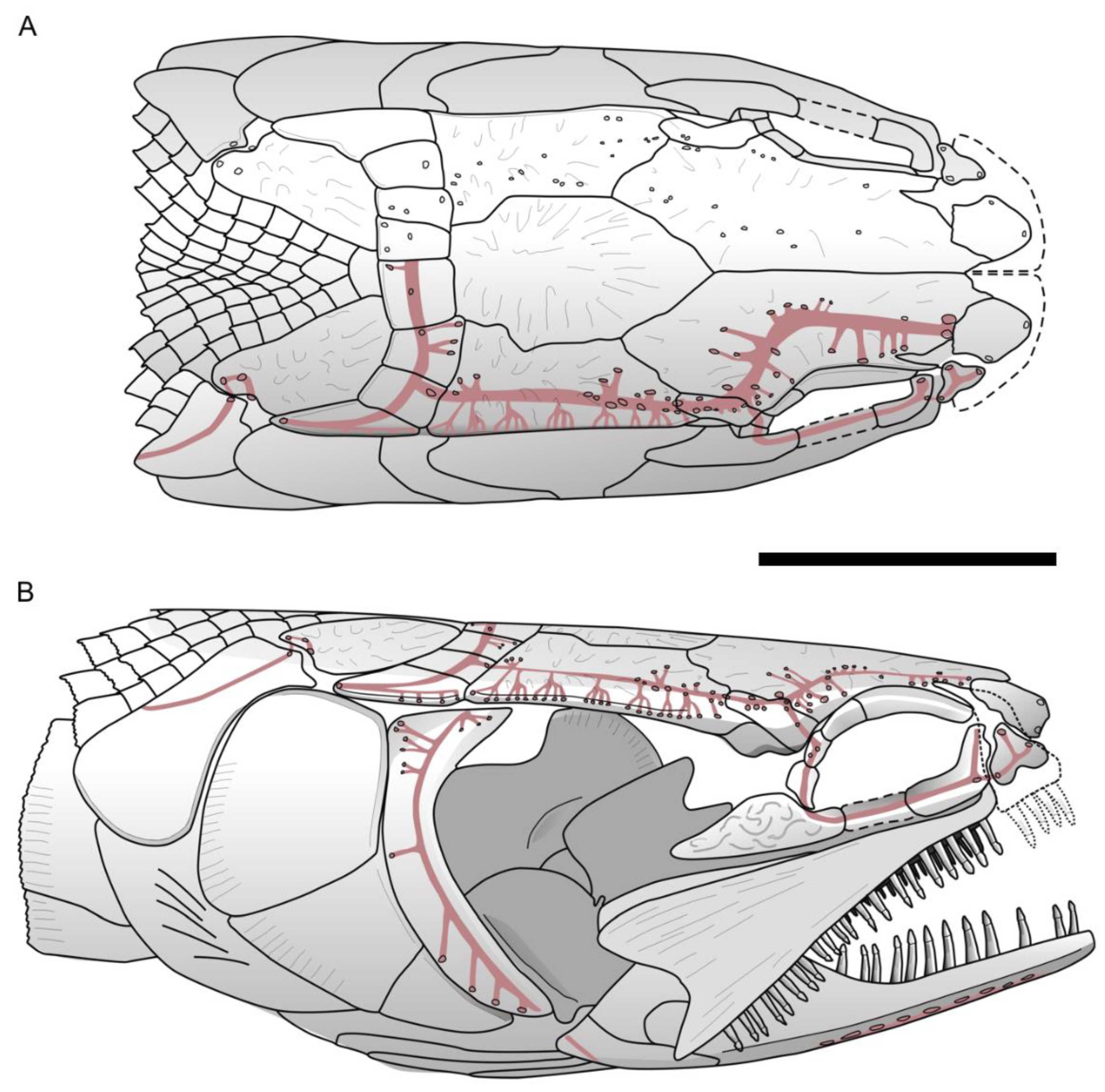
Skull roof. The frontals are the longest ossifications of the dermal skull roof. Each frontal is slightly narrower anteriorly than posteriorly, and marks a regular curvature at the level of the orbit. The posterior margin of the frontal is W-shaped and is firmly sutured with the paired dermopterotics and a median parietal. The anterior margin of the right frontal is notched. This notch is filled with a bone identified as the tip of the nasal process of the premaxilla. The lateral branch of the notch extends ventrally as a small process, with a concave surface that probably accommodated the dorsal extremity of the lacrimal (slightly shifted on the holotype). We cannot observe the suture between the left and right frontals on most of their length because of the presence of a crack along the mid-way of the skull roof. Only a short posterior portion of the suture presents a straight line towards the point extremity of the median parietal. The dorsal surface of the frontal bears a faint rugose ornamentation forming a reticulated pattern from the center of ossification. The frontal is twice as long as broad at its broadest posterior level. The parietal is a flat large median ossification as in other sinamiids. It is longer than wide (12 vs. 8 mm), with a sharp and pointed extremity wedged in the reversed V of the W-shaped margin of both frontals anteriorly. The right lateral margin is slightly undulate whereas the left lateral margin is straight. Both lateral margins mark a narrowing at the level of the posterior fourth of their length, thus forming a spearhead-shaped parietal. The dorsal surface of the parietal bears ridges radiating from the center of the bone. The dermopterotic borders laterally the parietal and extends anteriorly as a lateral pointed process with a slightly undulate suture along the posterolateral margin of the frontal. The dermopterotic is nearly three times longer than broad (14 vs. 5 mm), and as long, but narrower, as the parietal. Unlike the frontal and parietal, the dermopterotic is slightly convex in lateral view. Ridges radiating from the center of bone are present. The dermosphenotic is a small roughly rectangular bone wedged laterally between the right anterior extremity of the dermopterotic and the posterolateral margin of the frontal. It forms part of the skull roof. Its surface bears ornamentation similar to the rest of the skull roof bones. Posterior to the parietal and dermopterotics is a series of four pairs of extrascapulars. The six medial extrascapulars are rectangular in shape and the two most lateral ones are triangular with a blunt spine extending posteriorly. The external surface of the extrascapulars has no ornamentation, but small pores for the extrascapular commissure. The anteriormost preserved bone of the skull roof is the nasal, visible on the right side. The nasal is a plate-like ossification lying on the ascending nasal process of the premaxilla. It articulates with frontal posteriorly.
Braincase. Nothing is visible of the braincase, except the lateral extremity of the sphenotic wedged between the dermosphenotic and the dermopterotic, and fragments of ethmoid bones. The lateral ethmoid is visible on the right side as a vertical piece of bone held vertically under the anterolateral corner of the frontal, which forms a small ventral process (see above). On the left side, this region is not well preserved, and only a fragment of the lateral ethmoid is visible posterior to the nasal process of the premaxilla, which is almost vertical.
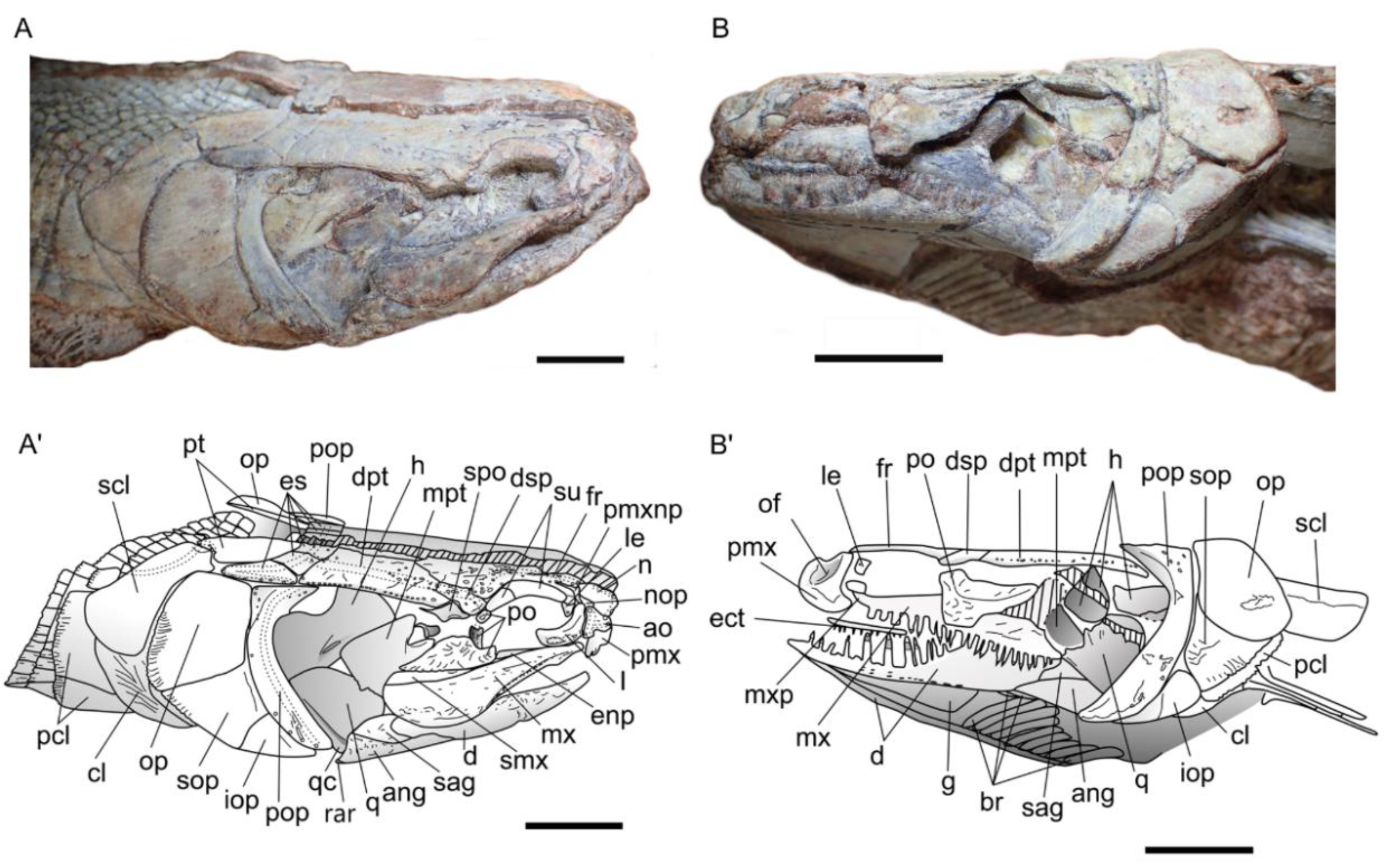
Circumorbital Ring. The circumorbital ring is well preserved on the right side. It consists of a dermosphenotic, two supraorbitals and three postinfraorbitals with the largest one located posteroventrally to the orbit, a lacrimal and an antorbital. Ventral to the orbit is a gap between the ventral most postinfraorbital and the lacrimal, in which lay probably one or two subinfraorbitals. The dermosphenotic ossification has no contact with the orbit, because anteriorly there is a tiny element corresponding likely to the third postinfraorbital. The shape of dermosphenotic is roughly rectangular with a notch at the anterodorsal corner of the bone. The external surface of the bone is not smooth and bears several pores located mostly along the dorsal edge. Two supraorbitals are present above the orbit. They are firmly sutured to each other and form a narrow crescent shape with their convex dorsal margins fitting well the deep curvature of the frontal leaving no space to the frontal to line the orbital space. Anterior to the orbit is the lacrimal, which is slightly shifted unveiling the lateral ethmoid underneath. It is boomerang-shaped, with the length of dorsal arm equal to ventral arm. Its concave dorsal margin forms the anteroventral border of the orbit. The antorbital, without contact with the orbit, is roughly triangular in shape with pore openings for the sensory canal at each summit of the triangle. A space delimitated by the antorbital, lacrimal, frontal and nasal corresponds to the posterior nostril. The lacrimal is separated by a small gap from the massive ventral postinfraorbital indicating that one or two small subinfraorbital were probably present on the living fish. The ventral postinfraorbital, situated at the posteroventral corner of the orbit, is the largest and more massive bone of the circumorbital ring. It bears strong ornamentation with strong knobs and grooves, even stronger than on the dermal roof bones. The shape is very peculiar, somewhat conch shell shaped, with the anterior portion protruding as a blunt process, with a broad mid-length area and with the posterior extremity tapering. The margins of the bone are smooth except the ventral one, which is undulate in correspondence to the ridges and grooves lying on the lateral surface. There are two small postinfraorbitals dorsally to the massive postinfraorbital. The second postinfraorbital is deeply sunk from its normal position. It is trapezoidal in shape with the ventral edge forming an oblique line contacting a curvature of the massive postinfraorbital. Its dorsal edge is indented at the mid-length for the path of the sensory canal. The third postinfraorbital is the smallest. It is rectangular in shape, and its surface bears a pore opening.
Jaws. The premaxilla is incomplete, being located in the most destroyed region of the skull. Therefore, its outline cannot be precisely described but the nasal process is apparently triangular in shape and held almost vertically. The extremity of the nasal process extends posteriorly as a spiny process fitting a notch on the frontal. The exposure of the nasal process on the skull roof is, as far as we know, unique within halecomorphs but present in lepisosteoidei among holosteans. On the right premaxilla, one broken tooth is visible on the oral margin of the bone, but the total number of teeth cannot be estimated. A large excavation marking the path of the olfactory nerve, with the nonvisible olfactory foramen located in the bottom is present on the left premaxilla. The maxillae are visible on both sides of the head. On the left side, the posterodorsal part of the bone is visible as an imprint only, but it clearly shows the concave margin that forms a deep notch. On the right side the posterior extremity of the maxilla appears distinctive convex, but the posteroventral part of the ossification is not preserved.
The oral margin of the maxilla is better preserved on the left side. It is slightly concave. There are about thirty-three teeth in total; each has a long conical base. The acrodine cap is very peculiar, being proportionally small with an arrow-head shape with cutting carinae. There are some pits on the lateral surface of the right maxilla. A supramaxilla is preserved on the right side just above the posterior part of the maxilla. It is narrow and elongate, ca. 8.5 mm in length and 2 mm in depth at the posterior part, and gradually tapering towards its anterior extremity. The length of the bone is about half length of the maxilla.
The mandibles are low and their ventral margins expand horizontally. The visible part of the lower jaw comprises a dentary that bears a single row of conical teeth, an angular, a supraangular and a retroarticular visible as a tiny ossification at the posteroventral tip of the right mandible. The angular forms most of the posterior part of the mandible. It contacts the dentary along a zigzag suture anteriorly and the supraangular dorsally, which is partly visible on both sides. The dentary is long and anteriorly very shallow in lateral view with the posterior part of the bone increasing in depth to form the coronoid process. The dentary bears about 20 conical teeth with pointed arrowhead carinate acrodine caps. The teeth vary in size: the largest ones are located at the mid-length while the ones on the posterior and anterior portions of the bone are smaller. Each tooth is composed of a high cylindrical base and extends with conical enamel stalk marked by fine ridges. Its acrodine cap forms a rounded base and gradually tapers at the tip. Along the lateral edges of acrodine tip are strongly sharp carinae. All teeth are curved inwardly into the mouth.
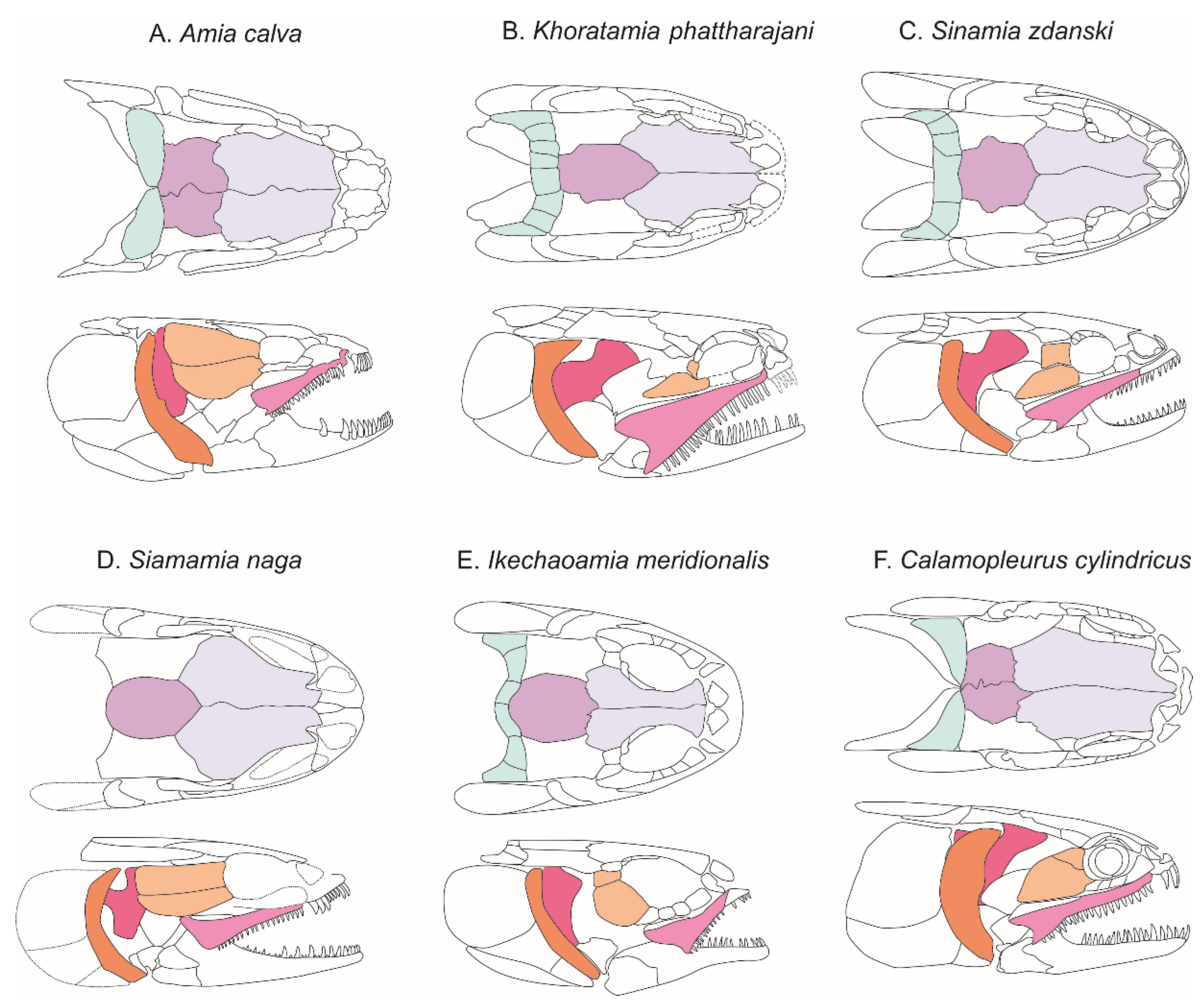
Suspensorium. The well-preserved suspensorium on the right side of the holotype shows the hyomandibula, the metapterygoid, the quadrate and a piece of the entopterygoid visible in the gap present in the circumorbital ring, whereas fragments of these bones plus a piece of ectopterygoid with small teeth is present on the left side. The right hyomandibula exposes its lateral side. The main axis of the bone is only slightly inclined posteriorly from the horizontal axis, and only part of the rounded articular head, the concave posterior margin and a fragment of the concave anterior margin are visible. The foramen and groove for the hyomandibular trunk, a mixed nerve containing fibers from cranial nerve VII plus the anteroventral lateral line nerve, opens near the anterior concave margin of the bone. The general shape of the hyomandibula is discussed in the ”comparisons” section. Anterior to the hyomandibula is the metapterygoid, which is roughly subrectangular, with a posterior portion slightly expanding over the hyomandibula and an anterior portion narrowing. Its anterior margin has a deep notch for the path of the trigeminal nerve, between the basal and otic processes, according to Stensiö’s (1935) nomenclature [
5]. Ventrally, it connects the quadrate through an interdigitate suture. The otic process of the metapterygoid is twisted while the basal process is blunt. The lateral surface of the ossification is totally concave. On the left side, the ectopterygoid is visible as a poorly preserved rod of bone with tiny recurved teeth. The quadrate is a large fan-shaped ossification. Its lateral surface shows a concavity extending the concavity on the metapterygoid. The convex ventral end forms the condyle for articulation with the lower jaw. Along the posteroventral border of the bone runs a strong crest starting from the condyle. The symplectic is not visible.
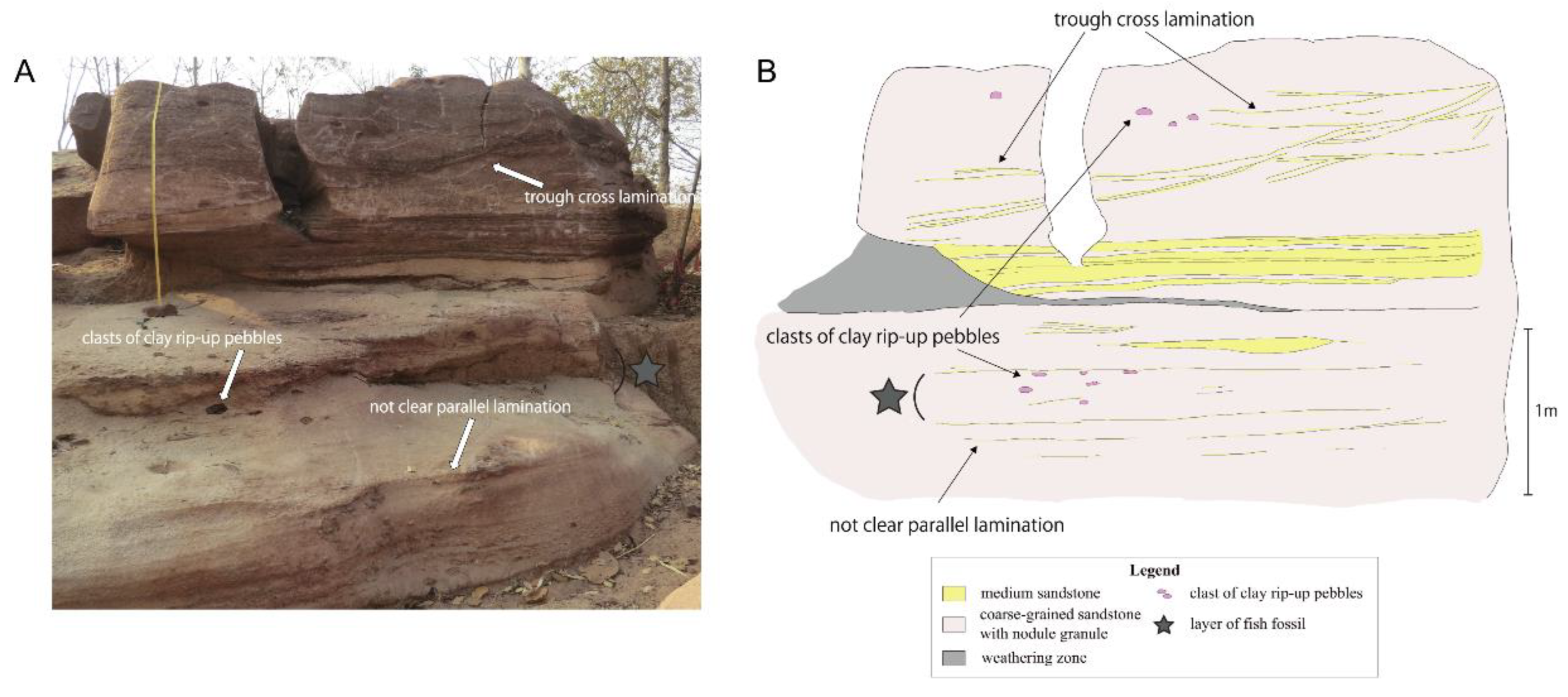
Opercular series, branchiostegal rays, and gular. The opercular series is complete, formed by the preopercle, operculum, subopercle and interopercle.
The preopercle is a narrow crescent-shaped bone with its extremities distinctly widening both ventrally and dorsally. The anterior and posterior margins are regularly curved, while the dorsal margin is straight. The preopercle is lying along the anterior edges of the opercle, subopercle and interopercle. The dorsoposterior edge of the bone forms a plan that marks an angle with the rest of the ossification and completely fits with the opercle. This feature seems to be peculiar to Khoratamia among halecomorphs. The anteroventral margin of the bone rests against the strong crest of quadrate, which runs along the upper arm. The opercle is roughly trapezoid in shape and deeper than long (12 × 8 mm). The dorsal and anterior borders are rather straight, while the posterior and ventral borders are slightly curved. The nearly upper half of the posterior edge is smooth whereas the lower part has strong serrations corresponding to the fine grooves present on its venteroposterior surface. The shape of the subopercle, lying between the opercle and interopercle, is somewhat the miniature of opercle in overturn position. It reaches the half depth of the opercle. The posterior edge of the subopercle is entirely serrated and no anterior ascending process is visible. The interopercle is visible as a triangular bone wedged between the subopercle and the posteroventral edge of the preopercle. Its anterior tip is pointed and it runs toward the ventral tip of the preopercle. The ventral margin of the bone is gently convex.
Eleven elongated branchiostegal rays are well preserved on the holotype, KT-181; the posterior most one is the shortest but the widest. The branchiostegal rays are rather closely packed to each other. Each ray is narrow proximally and much broader distally. The anterolateral surface of each ray bears a crest, which is pronounced at the articulated point and gradually decreases and becomes complete flat at the half length of the bone. Anterior to the branchiostegals is a median pyriform-shaped gular plate. The anterior portion is longer and narrower than the posterior portion. On the mid-line of the ventral surface is a ridge that nearly reaches the center of the bone. The ventral surface of the expended posterior portion shows faint ridges and grooves of regular size extending to the posterior margin, which forms a slightly undulate edge. In the central portion of the gular are randomly distributed small pits. The ceratohyal is partly exposed between the left dentary and the gular plate, but its shape is hard to describe precisely. A thin element anterior to the ceratohyal is regarded as the hypohyal.
Cephalic sensory canals. The numerous pores at the surface of the bones indicate the general pattern of the sensory canal system. In some parts of the head, the path of the canals can be seen through the very thin bone. The supraorbital sensory canal opens via numerous pores on the dermopterotic and frontal. The dermopterotic bears approximately 12 irregularly arranged pores on its dorsal surface. Moreover, tiny pores also open along the thin lateral margin of the dermopterotic, as previously observed in other sinamiids. The connection with the preopercular canal is not visible and it is possible that it occurred on the ventral surface of the bone. On the frontal, the pores are irregularly arranged and more concentrated above the posterior level of the orbit and in the anterior portion of the bone. The main supraorbital canal apparently forms an angle in the center of the frontal and gives off tubuli medially and laterally that reach the openings spread over the surface. The dermosphenotic, which is part of the skull roof, bears ca. six foramens irregularly arranged. The path of the canal in the nasal is unclear, but a large pore is present on the anterior margin of the bone. The supratemporal commissure is clearly visible by transparency along the mid-length of the extrascapulars. The lateral most extrascapular, which extends posteriorly, bears ca. six openings along its lateral border in alignment with the series on the dermopterotic. On the posttemporal, two openings for the sensory canal are present on the lateral margin of the posterior prominent knob of bone, and a third smaller foramen opens between the two on the dorsal surface. The pores on the lateral margin of the bone correspond to the entry and exit of the canal, which thus forms a short loop in the posttemporal. This pattern implies that a section of the canal is not bone-enclosed between the exit of the extrascapular and the entry in the posttemporal. The sensory canal runs posteroventrally from the knob on the posterior extremity of the posttemporal to the mid-depth of the bone posteriorly. The preopercular sensory canal runs along the center of the bone. It gives off posterior tubuli that open via a few pores in the depressed posterodorsal corner, at least one pore in the mid-depth of the bone and at least four pores along the ventral part of the preopercle. The infraorbital sensory canal crosses both small dorsal postinfraorbitals as indicates the presence of pores and notch. We cannot follow the path of the canal within the large ventral postinfraorbital, in particular the potential presence of tubuli extending posteriorly as in
Sinamia zdanskyi [
5] because of the presence of strong ornamentation. There are pores located at the center of the lacrimal indicating the path of the canal that reach the antorbital anteriorly. The three pores in the antorbital indicates the path of a canal that crosses the bone and reaches the rostral (not preserved), and an opening dorsally close to the posterior nostril. The mandibular sensory canal runs along the ventral horizontal lamina of bone formed by the angular and dentary. Small foramens along the medial margin of the angular, which are followed anteriorly by a larger pore at the limit between this bone and the dentary, and by another medium-size foramen aligned in the dentary. Along most of the length of the dentary, the ca. 10 large oval pores are shifted laterally compared to the posterior alignment, and located at the limit between the ventral and lateral sides of the mandible. At the anterior extremity of the mandible, five large oval foramens are located again along the medial margin.
3.2.3. Postcranium
Pectoral girdle and fin. The large posttemporal lies posteriorly to the extrascapulars. It is visible as a large trapezoidal to ovoid shaped ossification with a notch along its posterior margin for the path of the sensory canal. Its external surface bears a faint rugose ornamentation forming a reticulated pattern from the center of ossification. The anteroventral edge is strongly convex whereas its dorsal edge is almost straight with slight undulation. Because of the posterior extension of the lateral most extrascapulars, the posttemporals do not reach the lateral margin of the skull roof and have no direct contact with the opercle. The supracleithrum is proportionally very large, with an elongated general rectangular shape, an expended ventral extremity and a dorsal border firmly applied against the notch at the posterior margin of the posttemporal. Along the dorsal margin of the supracleithrum runs a short and curved ridge, which lies against a notch at the posterior margin of the posttemporal. The cleithrum is well visible on the right side. It is crescent in shape with the anterior part narrower than the posterior part. The lateral face of the cleithrum is marked by smooth ridges that converge to the center of the expanding posterior area. The ventral margin of the cleithrum is convex. Two large postcleithra are preserved in connection on the right side and slightly shifted on the left side. The upper postcleithrum is larger and deeper and is prominently convex dorsally and becomes thinner in its ventral part, which is flattened. The lowest postcleithrum is small and less convex, but it is not complete, making the estimate of its shape impossible. The posterior margins of the postcleithra are serrated with grooves extending on the surface of the bone. A saddle-shaped ossified scapulocoracoid is partly visible on the left side.
Eleven fin rays are visible, but more were likely present. The preserved portion of the rays, ca. 25 mm long, are unsegmented, indicating that the total length of the fin was proportionally longer because about half of the rays are unsegmented in the pectoral fin of Amia calva.
Vertebral column. A series of vertebrae is visible on the paratype (NRRU-F01020024). Five articulated centra are preserved wedged between the left and right scales covering of the body. The length of each centrum is shorter than its depth in lateral view (ca. 6 mm long and 11 mm deep, respectively). Owing to two broken posterior centra in their middle part, the sandglass shape of the centrum is visible and indicates that the anterior and posterior articular surfaces are deeply concave. The lateral surface present trace of grooves and ridges.
Dorsal fin. Although this area was destroyed, the origin of the dorsal fin can, however, be located. It is placed at the 5/8 distance of the pelvic fin.
Pelvic girdle and fins. The pelvic fins are apparently very small based on the fin rays partly preserved on the left side, which are small and short. The basipterygium is visible as a long rod with an enlarged posterior extremity.
Squamation. The whole body is covered by thick rhombic scales covered with a ganoin layer. The scales of the dorsal region are small, as long as deep, whereas the scales of the lateral and ventral regions of the trunk are distinctly longer than deep (about 3 mm long and 1.5 mm deep). In addition, their lateral surface has a ridge along the ventral edge in the holotype, but in KAB-30 (NRRU-F01020025) there are two ridges located near the dorsal and ventral margins. The posterior margin of all scales is serrated, with about 11 tiny denticulations in maximum (the number varies from 2 to 11 depending on the scale position on the body). The lateral line scales are not visible on the anterior part of the preserved body. At the level of the beginning of the dorsal fin, a scale every two or three scales is marked by a sensory pore, which has the shape of a vertical elongate furrow dug in the middle of the scale. No peg and socket articulation has been observed in the area where scales are altered. The imprints of scales indicate that the internal surface of the scales protrudes like a strong keel or ridge at its center. The keel is visible on scales preserved in internal view behind the dorsal fin of the holotype.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








