
First Record of Phlebotomus (Larroussius) perfiliewi (Diptera: Psychodidae), Vector of Leishmania infantum and Phleboviruses, in Spain

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
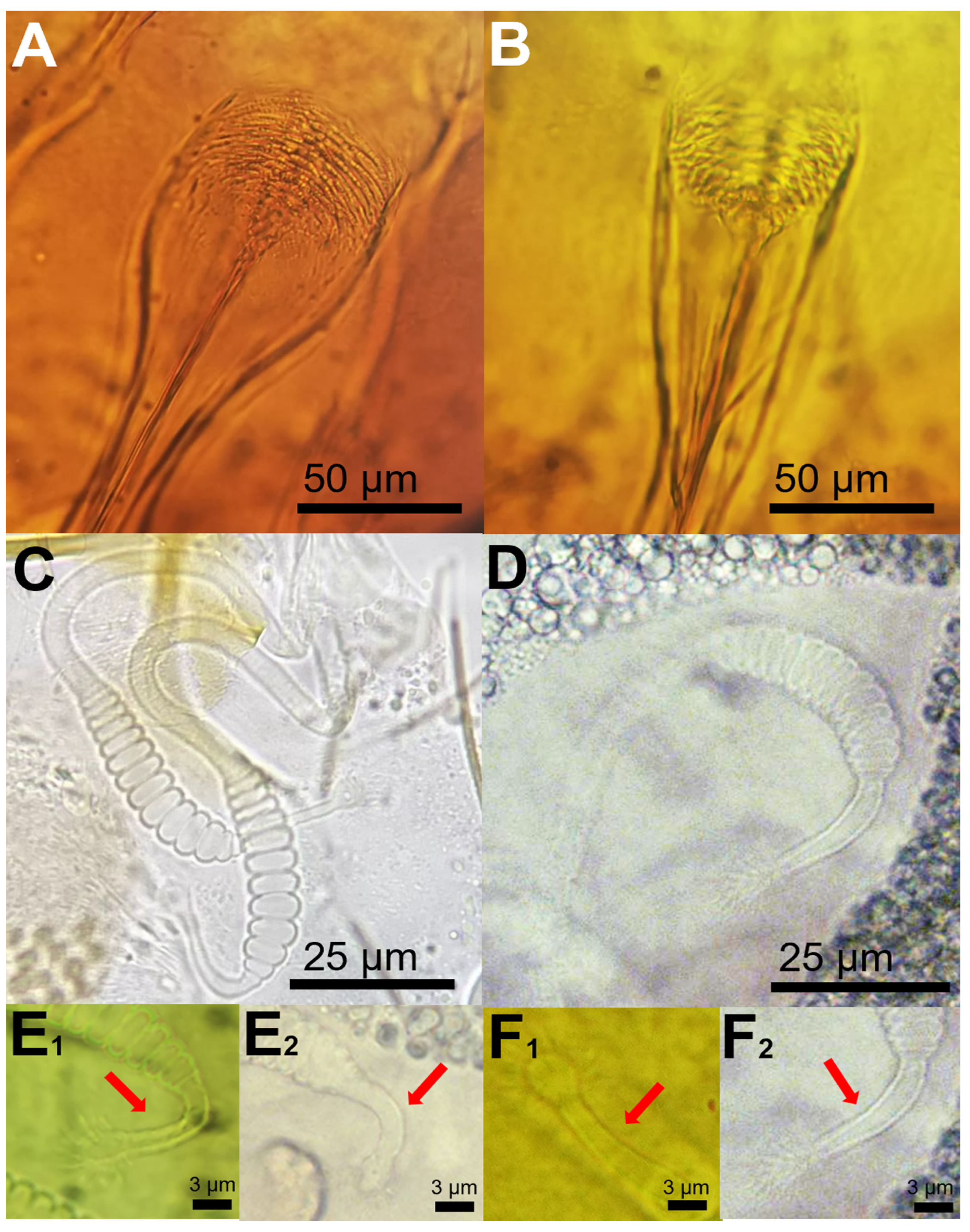
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of Public Health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Herrador, Z.; Gherasim, A.; Jimenez, B.C.; Granados, M.; San Martín, J.V.; Aparicio, P. Epidemiological changes in leishmaniasis in Spain according to hospitalization-based records, 1997–2011: Raising awareness towards leishmaniasis in non-HIV patients. PLoS Negl. Trop. Dis. 2015, 9, e0003594. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Van Bortel, W.; Zeller, H.; Alten, B. A Summary of the evidence for the change in European distribution of Phlebotomine sand flies (Diptera: Psychodidae) of Public Health importance. J. Vector Ecol. 2014, 39, 72–77. [Google Scholar] [CrossRef]
- González, E.; Jiménez, M.; Hernández, S.; Martín-Martín, I.; Molina, R. Phlebotomine sand fly survey in the focus of Leishmaniasis in Madrid, Spain (2012–2014): Seasonal dynamics, Leishmania Infantum infection rates and blood meal preferences. Parasites Vectors 2017, 10, 368. [Google Scholar] [CrossRef]
- Wagner, R.; Djong, H. Fauna Europaea: Psychodidae. In Fauna Europaea: Nematocera; De Jong, H., Ed.; version 1.3; Museum für Naturkunde Leibniz-Institut für Evolutions und Biodiversitätsforschung: Berlin, Germany, 2004; Available online: http://www.faunaeur.org (accessed on 15 November 2022).
- Bravo-Barriga, D.; Ruiz-Arrondo, I.; Peña, R.E.; Lucientes, J.; Delacour-Estrella, S. Phlebotomine sand flies (Diptera, Psychodidae) from Spain: An updated checklist and extended distributions. Zookeys 2022, 1106, 81–99. [Google Scholar] [CrossRef] [PubMed]
- Alcover, M.M.; Ballart, C.; Martín-Sánchez, J.; Serra, T.; Castillejo, S.; Portús, M.; Gállego, M. Factors influencing the presence of sand flies in Majorca (Balearic Islands, Spain) with special reference to Phlebotomus pernicious, vector of Leishmania Infantum. Parasites Vectors 2014, 7, 421. [Google Scholar] [CrossRef]
- Gil Collado, J.; Morillas-Marquez, F.; Sanchis Marin, M.C. Los flebotomos de España. Rev. Sanid. Hig. Publica 1989, 63, 15–34. [Google Scholar]
- Depaquit, J.Ô.; Bounamous, A.; Akhoundi, M.; Augot, D.; Sauvage, F.; Dvorak, V.; Chaibullinova, A.; Pesson, B.; Volf, P.; Léger, N. A Taxonomic study of Phlebotomus (Larroussius) perfiliewi s.L. Infect. Genet. Evol. 2013, 20, 500–508. [Google Scholar] [CrossRef]
- Gatt, P.; Williams, J.; Mifsud, D. New distributional data on sandflies from rubble walls in the Maltese Islands with an illustrated key to the Maltese species (Diptera: Phlebotominae). Bull. Entomol. Soc. Malta. 2009, 2, 95–110. [Google Scholar]
- Dantas-Torres, F.; Tarallo, V.D.; Otranto, D. Morphological keys for the identification of Italian phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae). Parasites Vectors 2014, 7, 479. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for amplification of mitochondrial Cytochrome c Oxidase Subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ruiz-Arrondo, I.; Hernández-Triana, L.M.; Ignjatović-Ćupina, A.; Nikolova, N.; Garza-Hernández, J.A.; Rodríguez-Pérez, M.A.; Oteo, J.A.; Fooks, A.R.; Lucientes Curdi, J. DNA barcoding of blackflies (Diptera: Simuliidae) as a tool for species identification and detection of hidden diversity in the eastern regions of Spain. Parasites Vectors 2018, 11, 463. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, L.; Prina, E.; Lang, T.; Milon, G. Real-Time PCR for detection and quantitation of Leishmania in mouse tissues. J. Clin. Microbiol. 2002, 40, 1666–1669. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- ECDC (European Centre for Disease Prevention and Control). Phlebotomus perfiliewi—Current Known Distribution. March 2022. Available online: https://www.ecdc.europa.eu/en/publications-data/phlebotomus-perfiliewi-current-known-distribution-march-2022 (accessed on 15 November 2022).
- Artemiev, M.M.; Neronov, V.M. Distribution and Ecology of Sandflies of the Old World (Genus Phlebotomus); Institut Ėvolyutsionnoĭ Morfologii i Ėkologii: Moscow, Russia, 1984; p. 207. [Google Scholar]
- Lewis, D.J. A Taxonomic Review of the Genus Phlebotomus (Diptera: Psychodidae); British Museum (Natural History): London, UK, 1982; pp. 121–209. [Google Scholar]
- García San Miguel, L.; Sierra, M.J.; Vazquez, A.; Fernandez-Martínez, B.; Molina, R.; Sanchez-Seco, M.P.; Lucientes, J.; Figuerola, J.; de Ory, F.; Monge, S.; et al. Phlebovirus-associated diseases transmitted by phlebotominae in Spain: Are we at risk? Enferm. Infecc. Microbiol. Clin. 2021, 39, 345–351. [Google Scholar] [CrossRef]
- Duport, M.; Lupaşcu, G.H.; Cristescu, A. Contribution à l’étude des phlébotomes des biotopes naturels de Roumanie. Arch. Roum. Path. Exp. Microbiol. 1971, 30, 387–398. [Google Scholar]
- Izri, M.A.; Belazzoug, S. Phlebotomus (Larroussius) perfiliewi naturally infected with dermotropic Leishmania infantum at Tenes, Algeria. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 399. [Google Scholar] [CrossRef]
- Oshaghi, M.A.; Ravasan, N.M.; Hide, M.; Javadian, E.A.; Rassi, Y.; Sadraei, J.; Mohebali, M.; Mehdi Sedaghat, M.; Hajjaran, H.; Zarei, Z.; et al. Phlebotomus perfiliewi transcaucasicus is circulating both Leishmania donovani and L. infantum in northwest Iran. Exp. Parasitol. 2009, 123, 218–225. [Google Scholar] [CrossRef]
- Zivkovic, V. Recherches récentes sur les phlébotomes (Diptera, Psychodidae) dans un foyer endémique de leishmaniose viscérale en Serbie (Yougoslavie). Acta Parasitol. Lugoslavica 1975, 6, 37–43. [Google Scholar]
- Farkas, R.; Tánczos, B.; Bongiorno, G.; Maroli, M.; Dereure, J.; Ready, P.D. First surveys to investigate the presence of canine leishmaniasis and its phlebotomine vectors in Hungary. Vector-Borne Zoonotic Dis. 2011, 11, 823–834. [Google Scholar] [CrossRef]
- Calzolari, M.; Carra, E.; Rugna, G.; Bonilauri, P.; Bergamini, F.; Bellini, R.; Varani, S.; Dottori, M. Isolation and molecular typing of Leishmania Infantum from Phlebotomus perfiliewi in a reemerging focus of leishmaniasis, Northeastern Italy. Microorganisms 2019, 7, 644. [Google Scholar] [CrossRef]
- Millán, J.; Ferroglio, E.; Solano-Gallego, L. Role of wildlife in the epidemiology of Leishmania infantum infection in Europe. Parasitol. Res. 2014, 113, 2005–2014. [Google Scholar] [CrossRef]
- Maresca, C.; Scoccia, E.; Barizzone, F.; Catalano, A.; Mancini, S.; Pagliacci, T.; Porrini, M.; Principato, M.; Venditti, G.; Grelloni, V. A survey on canine leishmaniasis and phlebotomine sand flies in Central Italy. Res. Vet. Sci. 2009, 87, 36–38. [Google Scholar] [CrossRef]
- Calzolari, M.; Angelini, P.; Finarelli, A.C.; Cagarelli, R.; Bellini, R.; Albieri, A.; Bonilauri, P.; Cavrini, F.; Tamba, M.; Dottori, M.; et al. Human and entomological surveillance of Toscana virus in the Emilia-Romagna region, Italy, 2010 to 2012. Eurosurveillance 2014, 19, 20978. [Google Scholar] [CrossRef]
- Calzolari, M.; Chiapponi, C.; Bellini, R.; Bonilauri, P.; Lelli, D.; Moreno, A.; Barbieri, I.; Pongolini, S.; Lavazza, A.; Dottori, M. Isolation of three novel reassortant phleboviruses, Ponticelli I, II, III, and of Toscana virus from field-collected sand flies in Italy. Parasites Vectors 2018, 11, 84. [Google Scholar] [CrossRef]
- Calzolari, M.; Rugna, G.; Clementi, E.; Carra, E.; Pinna, M.; Bergamini, F.; Fabbi, M.; Dottori, M.; Sacchi, L.; Votýpka, J. Isolation of a trypanosome related to Trypanosoma theileri (Kinetoplastea: Trypanosomatidae) from Phlebotomus perfiliewi (Diptera: Psychodidae). Biomed Res. Int. 2018, 2018, 2597074. [Google Scholar] [CrossRef]
- Rassi, Y.; Javadian, E.; Nadim, A.; Rafizadeh, S.; Zahraii, A.; Azizi, K.; Mohebali, M. Phlebotomus perfiliewi transcaucasicus, a vector of Leishmania infantum in Northwestern Iran. J. Med. Entomol. 2009, 46, 1094–1098. [Google Scholar] [CrossRef]
- Rassi, Y.; Sanei Dehkordi, A.; Oshaghi, M.A.; Abai, M.R.; Mohtarami, F.; Enayati, A.; Zarei, Z.; Javadian, E. First Report on Natural Infection of the Phlebotomus tobbi by Leishmania infantum in Northwestern Iran. Exp. Parasitol. 2012, 131, 344–349. [Google Scholar] [CrossRef]
- Epidemiología. Informes y Estadísticas de La Red de Vigilancia Epidemiológica de Les Illes Balears. February 2022. Available online: https://www.caib.es/sites/epidemiologia/es/informes_anuales-11681/ (accessed on 10 December 2022).
- Cazan, C.D.; Păstrav, I.R.; Györke, A.; Oguz, G.; Alten, B.; Mihalca, A.D. Seasonal dynamics of a population of Phlebotomus (Larroussius) perfiliewi Parrot, 1930 (Diptera: Psychodidae) in North-Eastern Romania. Parasitol. Res. 2019, 118, 1371–1384. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ph. perfiliewi s.s. | Ph. galilaeus | Ph. transcaucasicus | Ph. perniciosus | |
|---|---|---|---|---|
| Ph. perfiliewi s.s. | ||||
| Ph. galilaeus | 0.038 | |||
| Ph. transcaucasicus | 0.037 | 0.045 | ||
| Ph. perniciosus | 0.073 | 0.081 | 0.083 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, M.A.; Ruiz-Arrondo, I.; Gutiérrez-López, R.; Barceló, C.; Miranda, M.Á. First Record of Phlebotomus (Larroussius) perfiliewi (Diptera: Psychodidae), Vector of Leishmania infantum and Phleboviruses, in Spain. Diversity 2023, 15, 400. https://doi.org/10.3390/d15030400
González MA, Ruiz-Arrondo I, Gutiérrez-López R, Barceló C, Miranda MÁ. First Record of Phlebotomus (Larroussius) perfiliewi (Diptera: Psychodidae), Vector of Leishmania infantum and Phleboviruses, in Spain. Diversity. 2023; 15(3):400. https://doi.org/10.3390/d15030400
Chicago/Turabian StyleGonzález, Mikel A., Ignacio Ruiz-Arrondo, Rafael Gutiérrez-López, Carlos Barceló, and Miguel Á. Miranda. 2023. "First Record of Phlebotomus (Larroussius) perfiliewi (Diptera: Psychodidae), Vector of Leishmania infantum and Phleboviruses, in Spain" Diversity 15, no. 3: 400. https://doi.org/10.3390/d15030400
APA StyleGonzález, M. A., Ruiz-Arrondo, I., Gutiérrez-López, R., Barceló, C., & Miranda, M. Á. (2023). First Record of Phlebotomus (Larroussius) perfiliewi (Diptera: Psychodidae), Vector of Leishmania infantum and Phleboviruses, in Spain. Diversity, 15(3), 400. https://doi.org/10.3390/d15030400