



Climate Cycles, Habitat Stability, and Lineage Diversification in an African Biodiversity Hotspot


Abstract

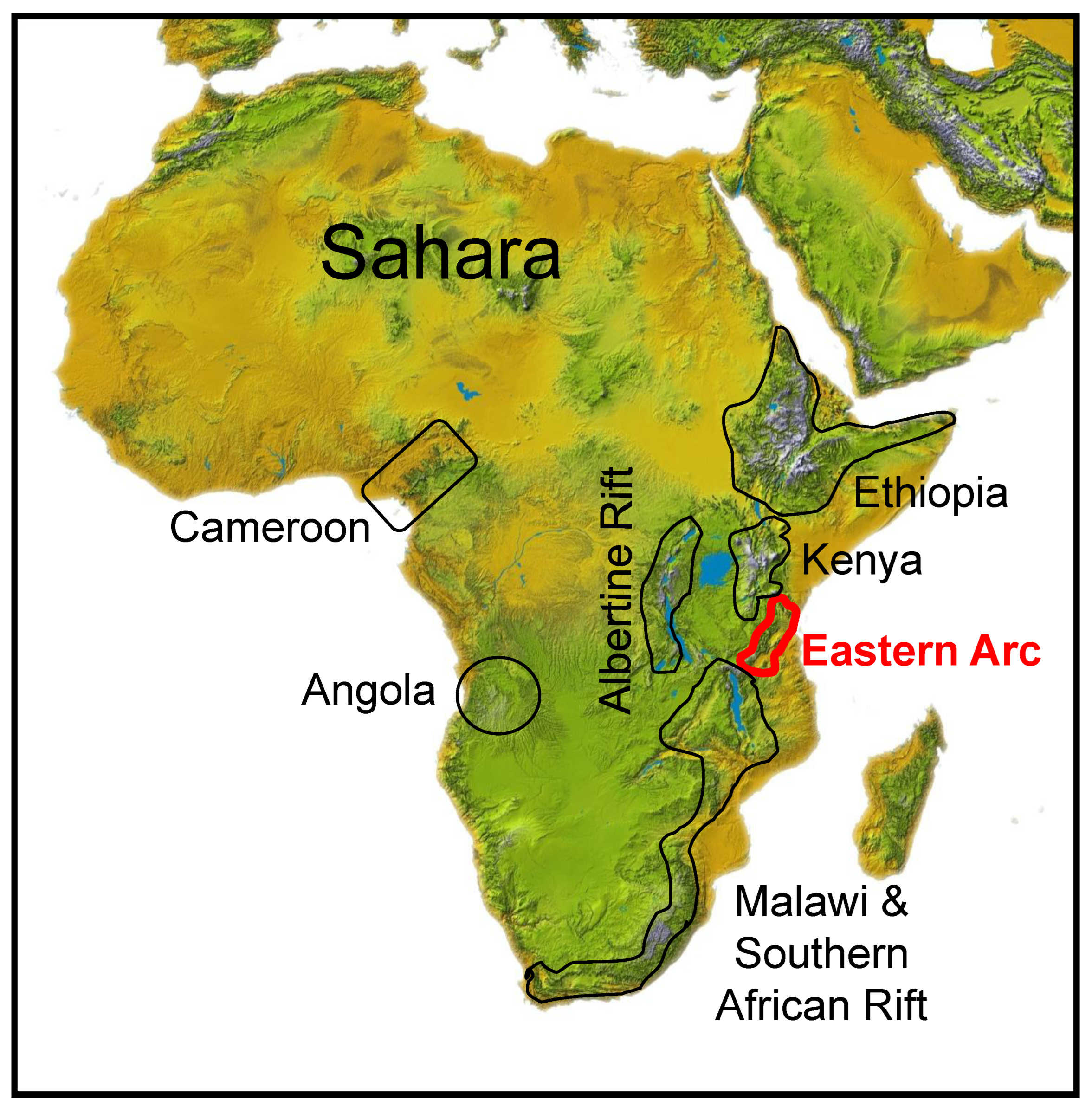
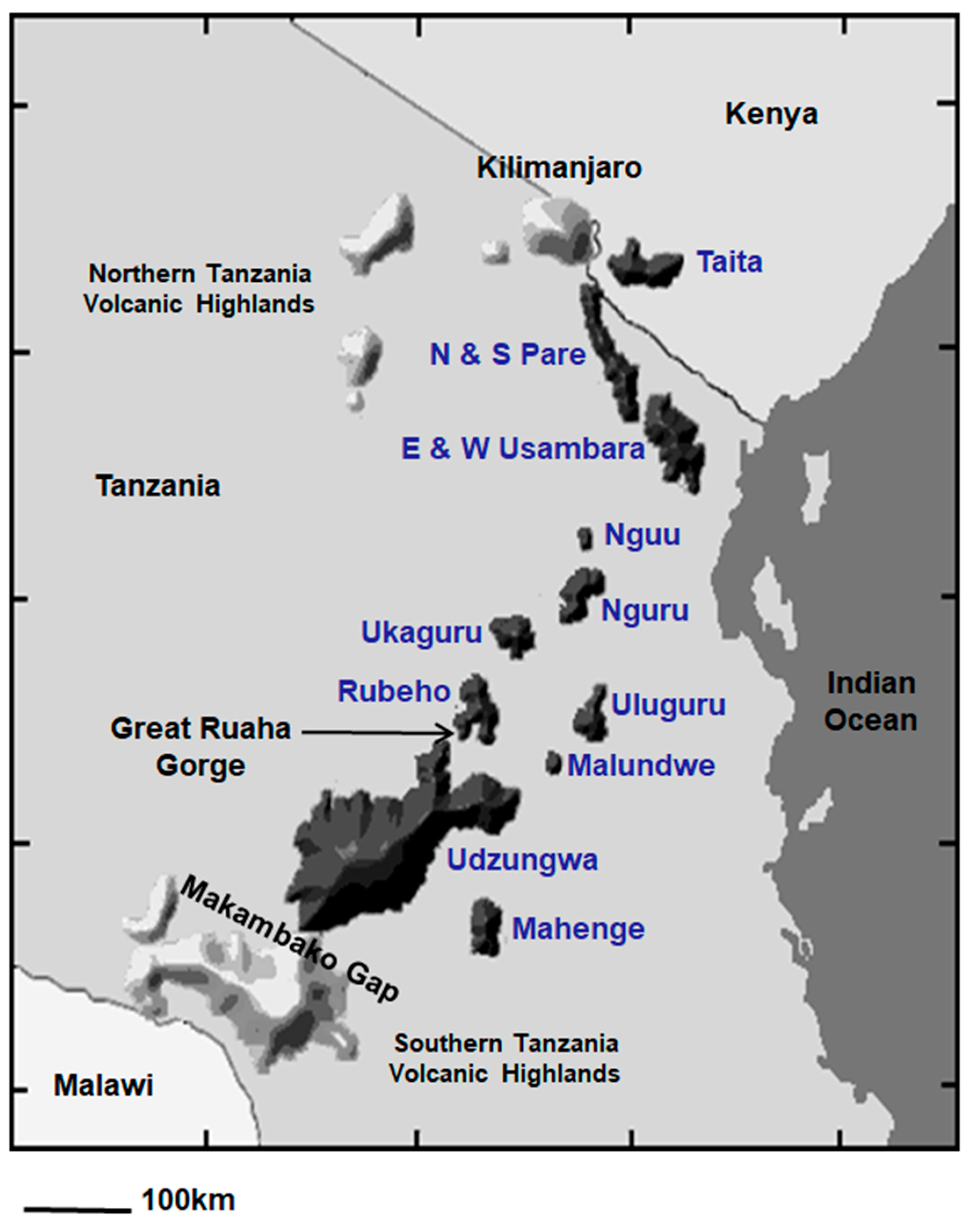
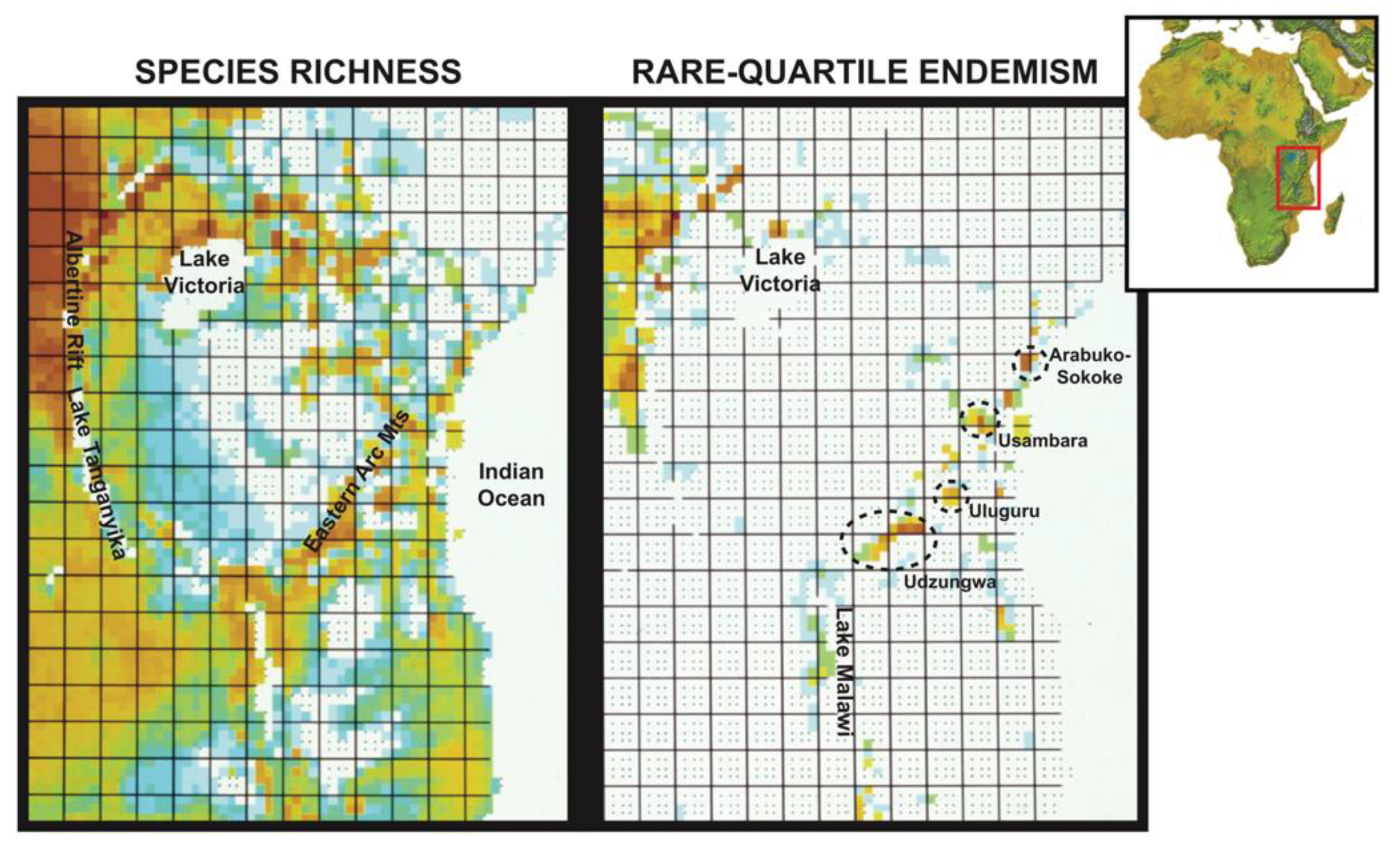
1. Introduction
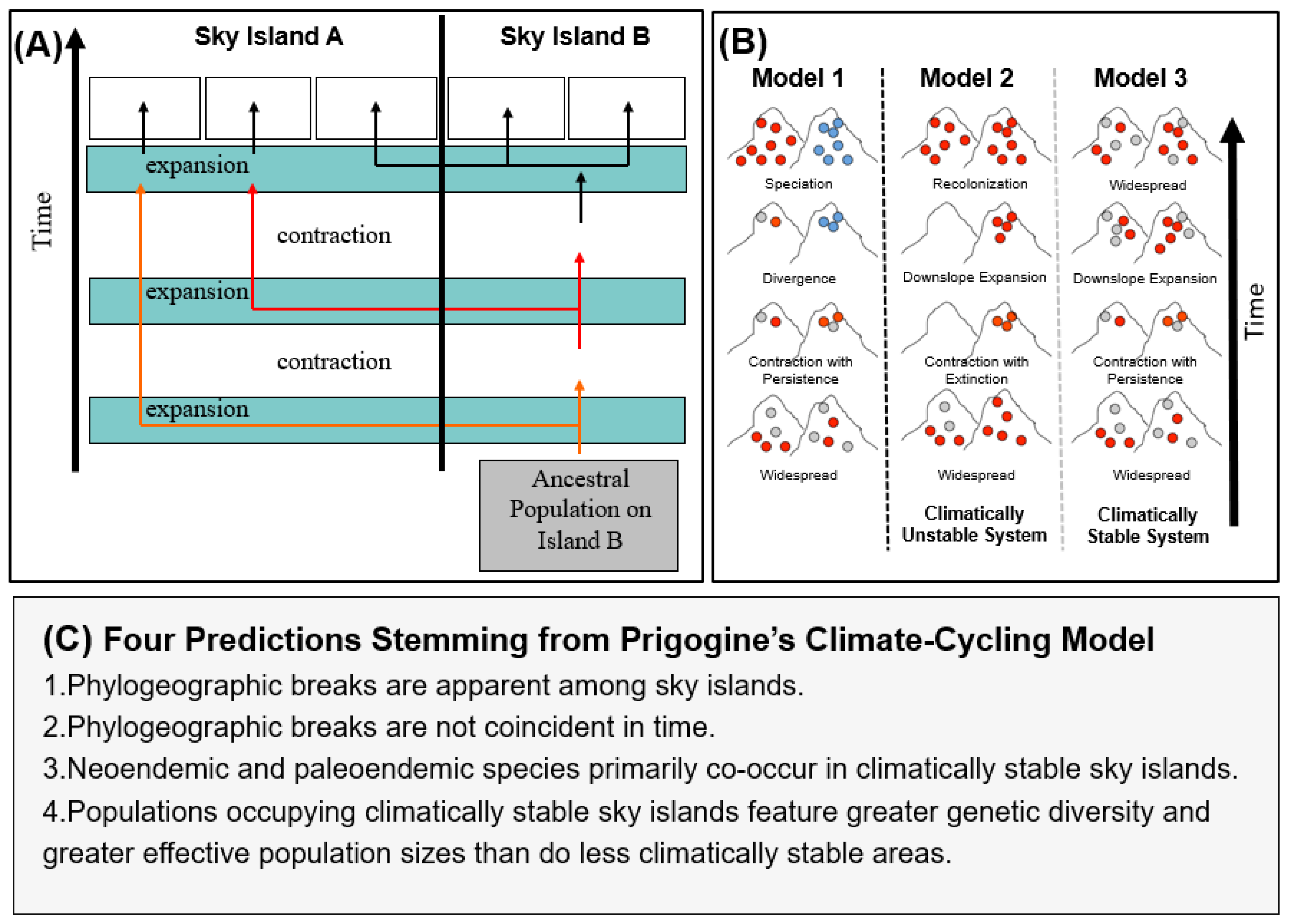
2. Materials and Methods
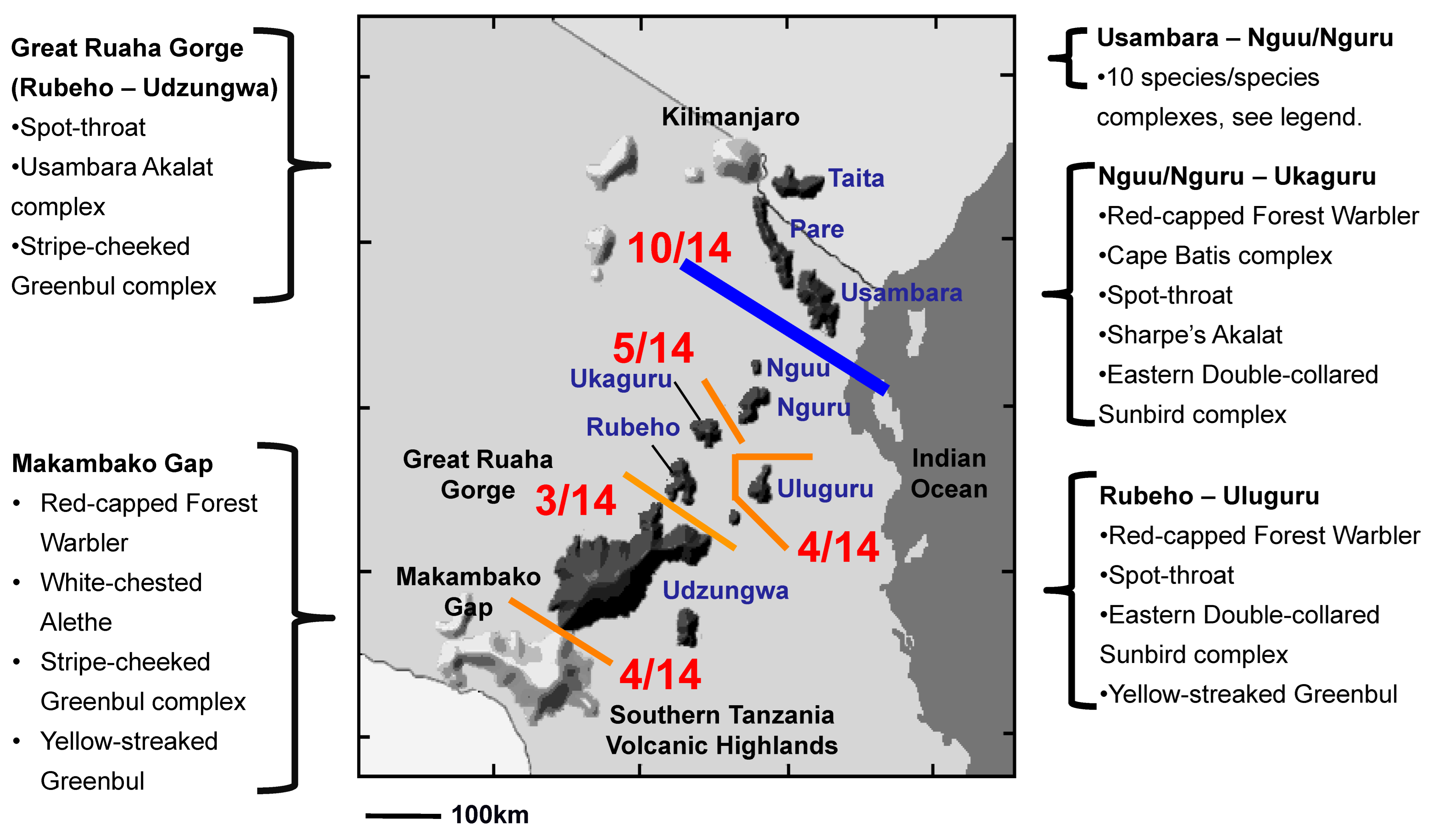
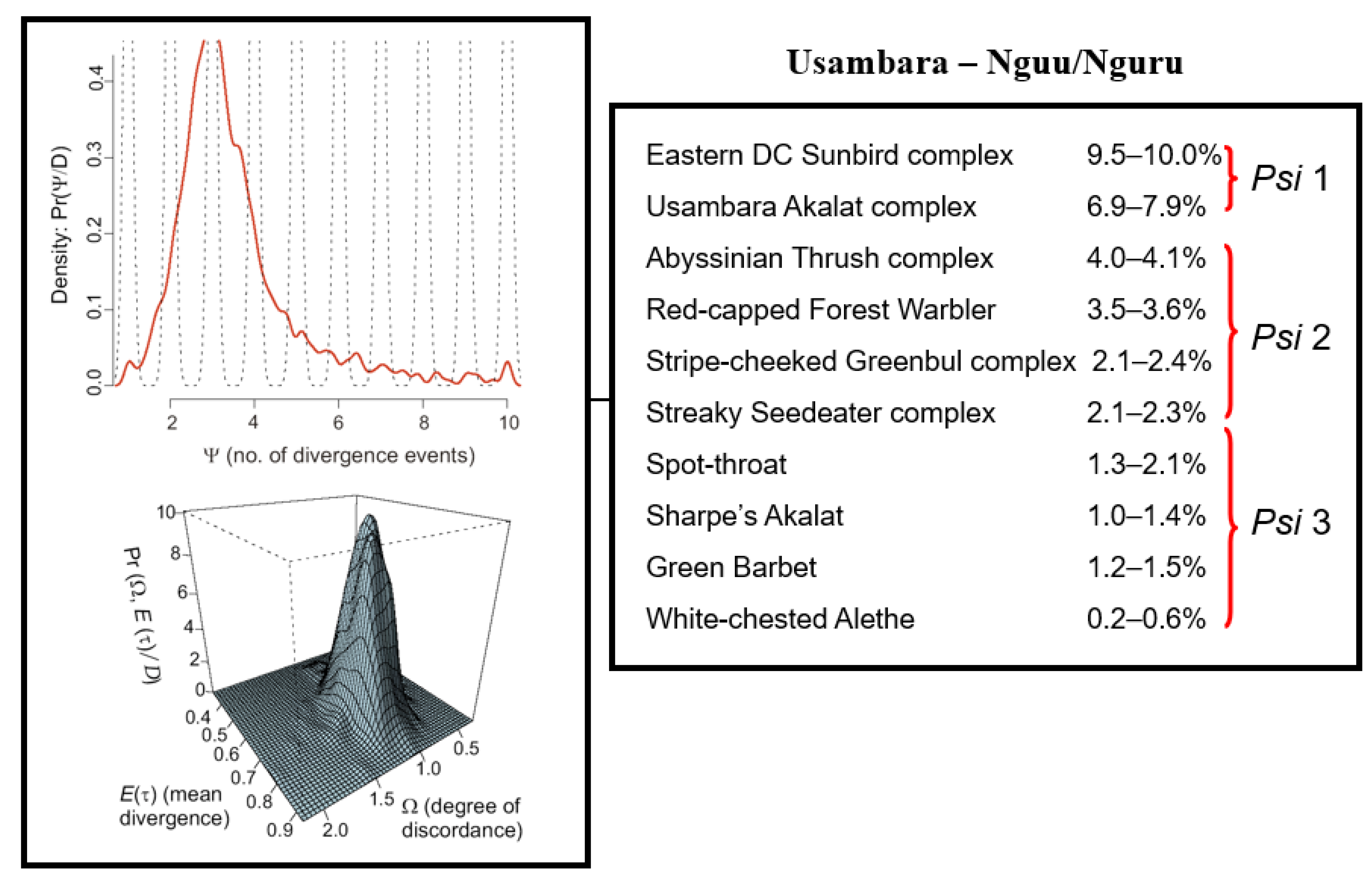
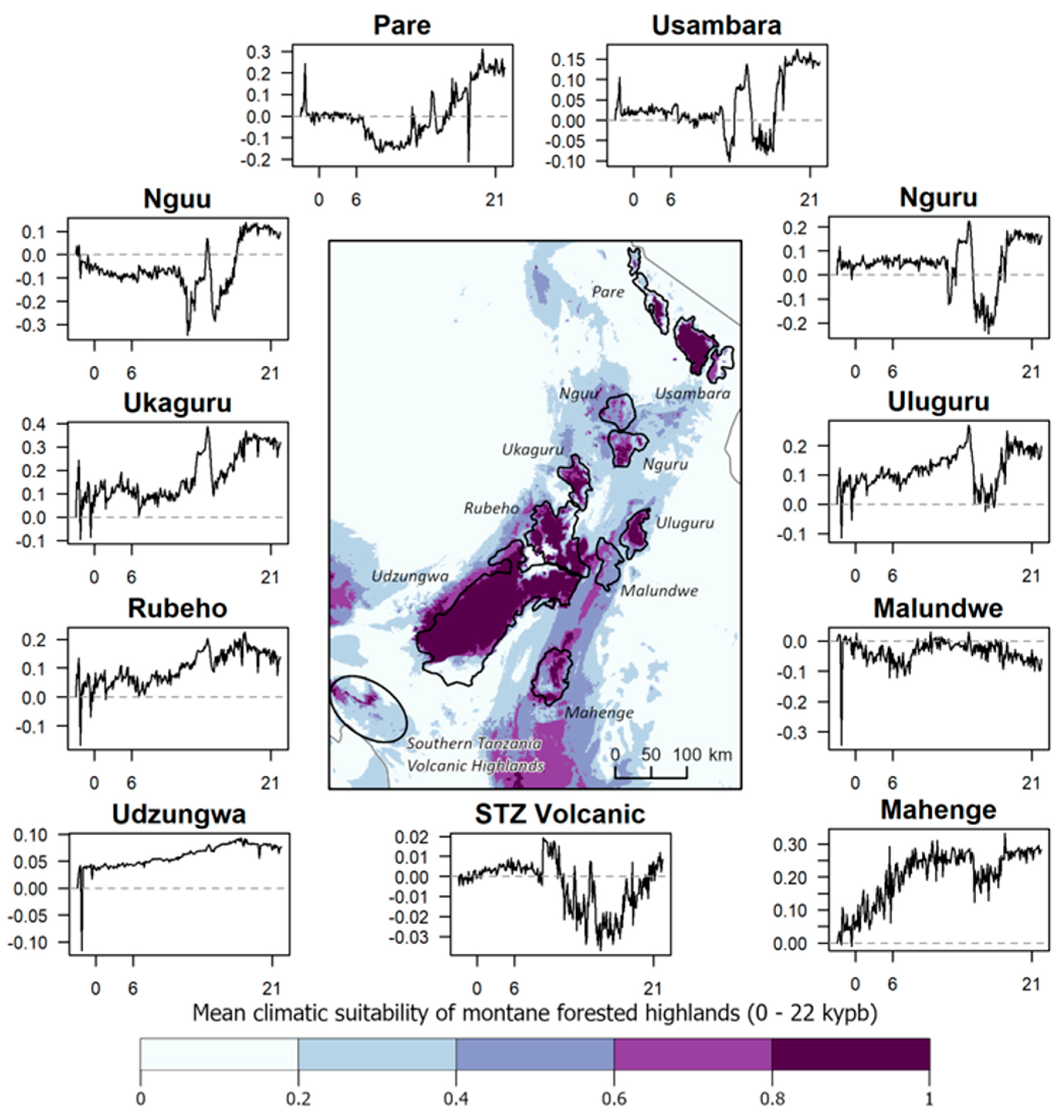
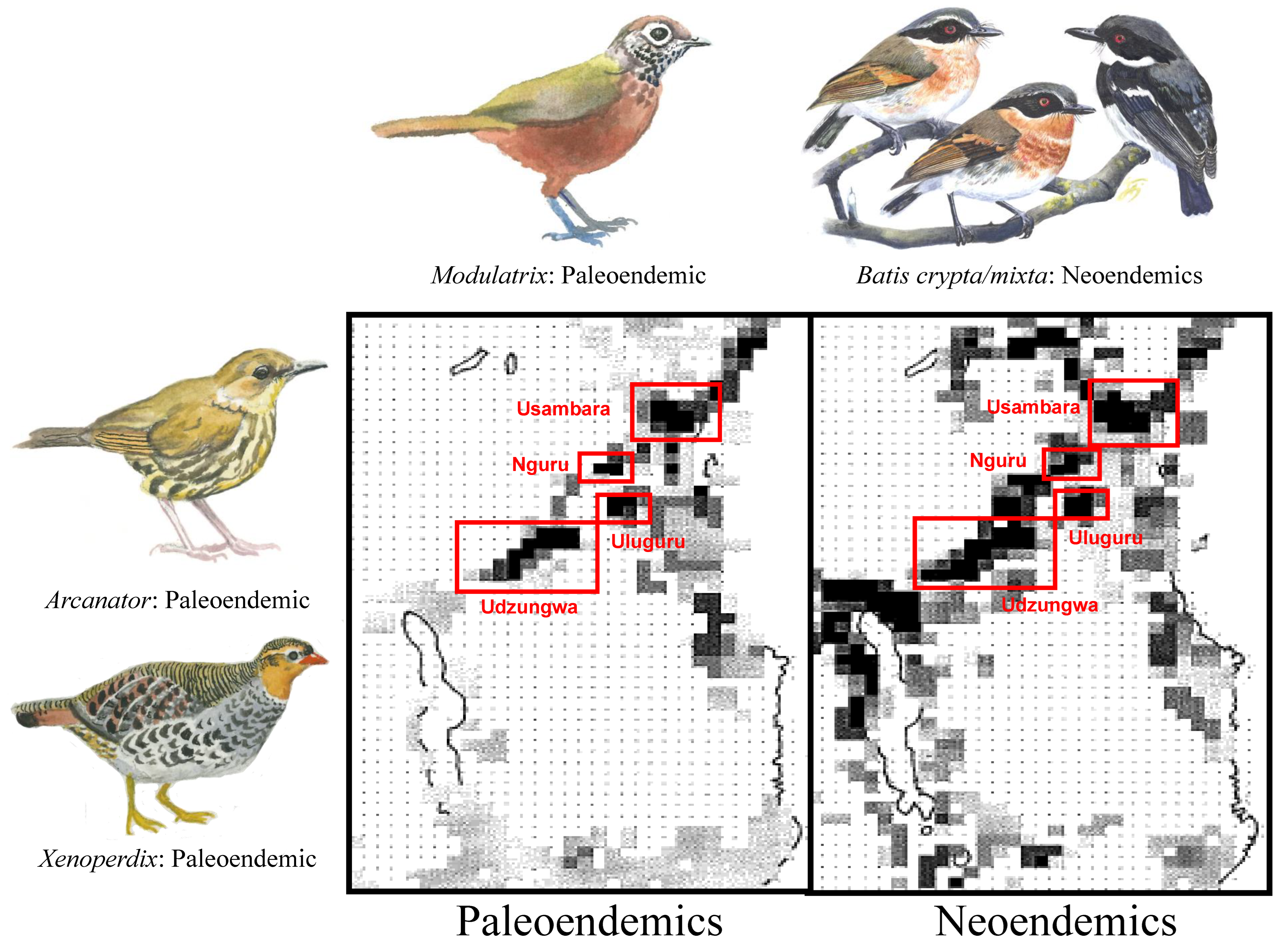
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dowsett, R.J. Origins of the high-altitude avifaunas of Tropical Africa. In High Altitude Tropical Biogeography; Vuilleumier, F.F., Monasterio, M., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 557–585. [Google Scholar]
- Bowie, R.C.K. Birds, Molecules and Evolutionary Processes among Africa’s Islands in the Sky. Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2003. [Google Scholar]
- Cooper, J.C. Hierarchical analyses of community biogeography in the Afromontane highlands. Front. Biogeogr. 2021, 13, e51310. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa; UNESCO: Paris, France, 1983. [Google Scholar]
- Hedberg, O. Origins of Afroalpine flora. In High Altitude Tropical Biogeography; Vuilleumier, F.F., Monasterio, M., Eds.; Oxford University Press: Oxford, UK, 1986; pp. 443–465. [Google Scholar]
- Prigogine, A. Disjunctions of montane birds in the Afrotropical region. Bonn. Zool. Beitr. 1987, 38, S195–S207. [Google Scholar]
- Fjeldså, J.; Bowie, R.C.K. New perspectives on Africa’s ancient forest avifauna. Afr. J. Ecol. 2008, 46, 235–247. [Google Scholar] [CrossRef]
- Fjeldså, J.; Bowie, R.C.K. Evolutionary and ecological explanations for the elevational flexibility of several East African bird species complexes. Front. Ecol. Evol. 2021, 9, 768062. [Google Scholar] [CrossRef]
- Biswas, A.; Karanth, K.P. Role of geographical gaps in the Western Ghats in shaping intra- and interspecific genetic diversity. J. Indian Inst. Sci. 2021, 101, 151–164. [Google Scholar] [CrossRef]
- Manthey, J.D.; Moyle, R.G. Isolation by environment in White-breasted Nuthatches (Sitta carolinensis) of the Madrean Archipelago sky islands: A landscape genomics approach. Mol. Ecol. 2015, 24, 3628–3638. [Google Scholar] [CrossRef]
- Moreau, R.E. The Bird Faunas of Africa and Its Islands; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Hall, B.P.; Moreau, R.E. An Atlas of Speciation in African Passerine Birds; British Museum: London, UK, 1970; (Natural History). [Google Scholar]
- Dowsett, R.J. Site-fidelity and survival rates of some montane forest birds in Malawi, south-central Africa. Biotropica. 1985, 17, 145–154. [Google Scholar] [CrossRef]
- Callens, T.; Galbusera, P.; Matthysens, E.; Durand, E.Y.; Githiru, M.; Huyghe, J.R.; Lens, L.U.C. Genetic signature of population fragmentation variers with mobility in seven bird species of a fragmented Kenyan Cloud Forest. Mol. Ecol. 2011, 20, 1829–1844. [Google Scholar] [CrossRef]
- Diamond, A.W.; Hamilton, A.C. The distribution of forest passerine birds and Quaternary climatic change in tropical Africa. J. Zool. 1981, 191, 379–402. [Google Scholar] [CrossRef]
- deMenocal, P.B. African climate change and faunal evolution during the Pliocene-Pleistocene. Earth Planet. Sci. Lett. 2004, 220, 3–24. [Google Scholar] [CrossRef]
- Trauth, M.H.; Maslin, M.A.; Deino, A.; Strecker, M.R. Late Cenozoic moisture history of East Africa. Science 2005, 309, 2051–2053. [Google Scholar] [CrossRef]
- Vrba, E.S. Turnover-pulses, the Red Queen, and related topics. Am. J. Sci. 1993, 293, 418–452. [Google Scholar] [CrossRef]
- Fjeldså, J. Geographical patterns of relict and young species of birds in Africa and South America and implications for conservation priorities. Biodiv. Conserv. 1994, 3, 107–126. [Google Scholar] [CrossRef]
- Fjeldså, J.; Lovett, J.C. Geographical patterns of old and young species in African forest biota: The significance of specific montane areas as evolutionary centres. Biodiv. Conserv. 1997, 6, 325–346. [Google Scholar] [CrossRef]
- Fjeldså, J.; Bowie, R.C.K.; Rahbek, R. The role of mountain ranges in the diversification of birds. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 249–265. [Google Scholar] [CrossRef]
- Azevedo, J.A.R.; Guedes, T.B.; Nogueira, C.D.C.; Passos, P.; Sawaya, R.J.; Prudente, A.L.C. Museums and cradles of diversity are geographically coincident for narrowly distributed Neotropical snakes. Ecography 2020, 43, 328–339. [Google Scholar] [CrossRef]
- Cooper, J.C.; Crouch, N.M.A.; Ferguson, A.W.; Bates, J.M. Climate refugia and reduced extinction correlate with underdispersion in mammals and birds in Africa. Ecol. Evol. 2022, 12, e8752. [Google Scholar]
- Päckert, M.; Martens, J.; Sun, Y.-H.; Severinghaus, I.I.; Nazarenko, A.A.; Ting, J.; Töpfer, T.; Tietze, D.T. Horizontal and elevational phylogenographic patterns of Himalayan and Southeast Asian forest passerines (Aves: Passeriformes). J. Biogeogr. 2012, 39, 556–573. [Google Scholar] [CrossRef]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogues-Bravo, D.; Rasmussen, C.M.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Päckert, M.; Favre, A.; Schnitzler, J.; Martens, J.; Sun, Y.-H.; Tietze, D.T.; Hailer, F.; Michalak, I.; Strutzenberger, P. Into and out of the Quinghai-Tibet Plateau and the Himalayas: Centers of origin and diversification across five clades of Eurasian montane and alpine passerine birds. Ecol. Evol. 2020, 10, 9283–9300. [Google Scholar] [CrossRef]
- Sonne, J.; Dalsgaard, B.; Borregaard, M.K.; Kennedy, J.; Fjeldså, J.; Rahbek, C. Biodiversity cradles and museums segregating within hotspots of endemism. Proc. R. Soc. B 2022, 289, 20221102. [Google Scholar]
- van Els, P.; Herrera-Alsine, L.; Pigot, A.L.; Etienne, R. Dynamical analysis of the global diversity gradient in passerine birds reveals a prominent role for highlands as species pumps. Nat. Ecol. Evol. 2021, 5, 1259–1265. [Google Scholar] [CrossRef]
- Marchant, R.; Mumbi, C.; Behera, S.; Yamagata, T. The Indian Ocean dipole—The unsung driver of climatic variability in East Africa. Afr. J. Evol. 2007, 45, 4–16. [Google Scholar] [CrossRef]
- Mumbi, C.T.; Marchant, R.; Hooghiemstra, H.; Woeller, M.J. Late Quaternary vegetation reconstruction from the Eastern Arc Mountains, Tanzania. Quat. Res. 2008, 69, 326–341. [Google Scholar] [CrossRef]
- Finch, J.; Leng, M.J.; Marchant, R.A. Late Quaternary vegetation dynamics in a biodiversity hotspot, the Uluguru Mountains of Tanzania. Quat. Res. 2009, 72, 111–122. [Google Scholar] [CrossRef]
- Macgregor, D. History of the development of the East African Rift System: A series of interpreted maps through time. J. Afr. Earth Sci. 2015, 101, 232–252. [Google Scholar] [CrossRef]
- Antonelli, A.; Kissling, W.D.; Flantua, S.G.A.; Bermúdez, M.A.; Mulch, A.; Muellner-Riehl, A.N.; Kreft, H.; Linder, H.P.; Badgley, C.; Fjeldså, J.; et al. Geological and climatic influences on mountain diversity. Nat. Geosci. 2018, 11, 718–725. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef]
- Linder, P.; de Klerk, H.; Born, J.; Burgess, N.; Fjeldså, J.; Rahbek, C. The partitioning of Africa: Statistically defined biochorological zones in sub-Saharan Africa. J. Biogeogr. 2012, 39, 1189–1205. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C.; Colwell, R.K. The coincidence of rarity and richness and the potential signature of history in centers of endemism. Ecol. Lett. 2004, 7, 1180–1191. [Google Scholar] [CrossRef]
- Burgess, N.D.; Butynski, T.M.; Cordeiro, N.J.; Doggart, N.H.; Fjeldså, J.; Howell, K.M.; Kilahama, F.B.; Loader, S.P.; Lovett, J.C.; Mbilinyi, B.; et al. The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. Biol. Conserv. 2007, 134, 209–231. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Robles-Gill, P.; Hoffmann, M.; Pilgrim, J.D.; Brooks, T.B.; Mittermeier, C.G. Hotspots Revisited. Earth’s Biologically Richest and Most Endangered Ecoregions; CEMEX: Mexico City, Mexico, 2004. [Google Scholar]
- Bowie, R.C.K.; Pasquet, E.; McEntee, J.P.; Njilima, F.; Fjeldså, J. The systematics and biogeography of African Tailorbirds (Cisticolidae: Artisornis) with comments on the choice of Bayesian branch-length prior when analyzing heterogeneous data. Mol. Phylogenet. Evol. 2018, 118, 172–183. [Google Scholar] [CrossRef]
- Mostert, M.E. Unravelling species boundaries and high altitude phylogeography of the Streaky Canary of east and central Africa (Serinus striolatus). M.Sc. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2005. [Google Scholar]
- Beresford, P.; Fjeldså, J.; Kiure, J. A new species of akalat (Sheppardia) narrowly endemic in the Eastern Arc of Tanzania. Auk 2004, 121, 23–34. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J.; Hackett, S.J.; Bates, J.M.; Crowe, R.M. Coalescent models reveal the relative roles of dispersal, vicariance and ancestral polymorphism in shaping phylogeographical structure of an African montane forest robin. Mol. Phylogenet. Evol. 2006, 38, 171–188. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J.; Hackett, S.J.; Crowe, T.M. Systematics and biogeography of the double-collared sunbirds (Nectariniidae) of the Eastern Arc Mountains of Tanzania. Auk 2004, 121, 660–681. [Google Scholar] [CrossRef]
- McEntee, J.P.; Peñalba, J.V.; Werema, C.; Mulungi, E.; Mbilinyi, M.; Moyer, D.; Hansen, L.; Fjeldså, J.; Bowie, R.C.K. Social selection parapatry in Afrotropical sunbirds. Evolution 2016, 70, 1307–1321. [Google Scholar] [CrossRef]
- McEntee, J.P.; Zhelezov, G.; Werema, C.; Najar, N.; Peñalba, J.V.; Mulungi, E.; Mbilinyi, M.; Karimi, S.; Chumakova, L.; Burleigh, J.G.; et al. Inferring punctuated evolution in the learned songs of African sunbirds. Proc. R. Soc. B Biol. Sci. 2021, 288, 20212062. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J.; Hackett, S.J.; Crowe, T.M. Molecular evolution in space and though time: mtDNA phylogeography of the Olive Sunbird (Nectarinia olivacea/obscura) throughout continental Africa. Mol. Phylogenet. Evol. 2004, 33, 56–76. [Google Scholar] [CrossRef]
- Fjeldså, J.; Bowie, R.C.K.; Kiure, J. The Forest Batis Batis mixta is two species: Description of a new, narrowly distributed Batis species in the Eastern Arc biodiversity hotspot. J. Ornithol. 2006, 147, 578–590. [Google Scholar] [CrossRef]
- Kaliba, P.M. Faunal turnover between east and southern African terrestrial vertebrates: Is Malawi the geographical break? Ph.D. Thesis, University of Cape Town, Cape Town, South Africa, 2014. [Google Scholar]
- Johansson, U.S.; Fjeldså, J.; Lokugalappatti, L.G.S.; Bowie, R.C.K. A nuclear DNA phylogeny and proposed taxonomic revision of African greenbuls (Aves, Passeriformes, Pycnonotidae). Zool. Scripta 2007, 36, 417–427. [Google Scholar] [CrossRef]
- Lokugalappatti, L.G.S. Climatic Perturbations and Speciation of Southern and Eastern African Greenbuls (Passeriformes, Pycnonotidae). Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2011. [Google Scholar]
- Bowie, R.C.K.; Voelker, G.; Fjeldså, J.; Lens, L.; Hackett, S.J.; Crowe, T.M. Systematics of the Olive Thrush Turdus olivaceus species complex with reference to the taxonomic status of the endangered Taita Thrush, T. helleri. J. Avian Biol. 2005, 36, 391–404. [Google Scholar] [CrossRef]
- Voelker, G.; Rohwer, S.; Bowie, R.C.K.; Outlaw, D. Molecular systematics of a speciose, cosmopolitan songbird genus: Defining the limits of, and relationships among, the Turdus thrushes. Mol. Phylogenet. Evol 2007, 42, 422–434. [Google Scholar] [CrossRef]
- Dowsett-Lemaire, F.; Dowsett, R.J. The Birds of Malawi; Tauraco Press: Liege, Belgium, 2006. [Google Scholar]
- Dowsett, R.J.; Aspinwall, D.R.; Dowsett-Lemaire, F. The Birds of Zambia: An Atlas and Handbook; Tauraco Press: Liege, Belgium, 2008. [Google Scholar]
- Fjeldså, J.; Kiure, J.; Doggart, N.; Hansen, L.A.; Perkin, A. Distribution of highland forest birds across a potential dispersal barrier in the Eastern Arc Mountains of Tanzania. Steenstrupia 2010, 32, 1–43. [Google Scholar]
- Rovero, F.; Menegon, M.; Fjeldså, J.; Collett, L.; Doggart, N.; Leonard, C.; Norton, G.; Owen, N.; Perkin, A.; Spitale, D.; et al. Targetted vertebrate surveys enhance the faunal importance and improve explanatory models within the Eastern Arc Mountains of Kenya and Tanzania. Divers. Distrib. 2014, 20, 1438–1449. [Google Scholar] [CrossRef]
- Sullivan, B.L.; Wood, C.L.; Iliff, M.J.; Bonney, R.E.; Fink, D.; Kelling, S. eBird: A citizen-based bird observation network in the biological sciences. Biol. Conserv. 2009, 142, 2282–2292. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J.; Kiure, J.; Kristensen, J. A new member of the greater double-collared sunbird complex (Passeriformes: Nectariniidae) from the Eastern Arc Mountains of Africa. Zootaxa 2016, 4175, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Voelker, G.; Outlaw, R.K.; Reddy, S.; Tobler, M.; Bates, J.M.; Hackett, S.J.; Kahindo, C.; Marks, B.D.; Kerbis Peterhans, J.C.; Gnoske, T.P. A new species of boubou (Malaconotidae: Laniarius) from the Albertine Rift. Auk 2010, 127, 678–689. [Google Scholar] [CrossRef]
- Fjeldså, J.; Dinesen, L.; Davies, O.R.; Irestedt, M.; Krabbe, N.K.; Hansen, L.A.; Bowie, R.C.K. Description of two new Cisticola species endemic to the marshes of the Kilombero floodplain of southwestern Tanzania. Ibis 2021, 163, 1330–1354. [Google Scholar] [CrossRef]
- Lane, M.A.; Edwards, J.L. The Global Biodiversity Information Facility (GBIF). Biodivers. Databases 2007, 73, 1. [Google Scholar]
- Hansen, L.A.; Fjeldså, J.; Burgess, N.D.; Rahbek, C. One degree resolution databases of the distribution of 1789 resident birds in Sub-Saharan Africa. Online Data Source-Version 1.00; Zoological Museum, University of Copenhagen: Copenhagen, Denmark, 2007. [Google Scholar]
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List (v12.1). 2022. Available online: https://www.worldbirdnames.org/ioc-lists/crossref/ (accessed on 1 July 2022).
- Dowsett-Lemaire, F. Ecological and territorial requirements of montane forest birds on the Nyika Plateau, south-central Africa. Gerfaut 1983, 73, 345–378. [Google Scholar]
- Fry, C.H.; Urban, E.K.; Keith, S.; Safford, R.; Hawkins, F. The Birds of Africa; Christopher Helm: London, UK, 1986–2004. [Google Scholar]
- Fuchs, J.; Bowie, R.C.K.; Fjeldså, J.; Pasquet, E. Phylogenetic relationships of the African bush-shrikes and helmet-shrikes (Passeriformes: Malaconotidae). Mol. Phylogenet. Evol. 2004, 33, 428–439. [Google Scholar] [CrossRef]
- Kimball, R.T.; Braun, E.L.; Barker, F.K.; Bowie, R.C.K.; Braun, M.J.; Chojnowski, J.L.; Hackett, S.J.; Han, K.-L.; Harshman, J.; Heimer-Torres, V.; et al. A well-tested set of primers to amplify regions spread across the avian genome. Mol. Phylogenet. Evol. 2009, 50, 654–660. [Google Scholar] [CrossRef]
- Voelker, G.; Outlaw, R.K.; Bowie, R.C.K. Pliocene forest dynamics as a primary driver of African bird speciation. Glob. Ecol. Biogeogr. 2010, 19, 111–121. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In 2010 Gatewway Computing Environments Workshop (GCE); IEEE: New Orleans, LA, USA, 2010. [Google Scholar] [CrossRef]
- Oliveros, C.H.; Field, D.J.; Ksepka, D.T.; Barker, F.K.; Aleixo, A.; Anderson, M.J.; Hosner, P.A.; Joseph, L.; Mack, A.L.; Robbins, M.B. Earth history and the passerine superradiation. Proc. Nat. Acad. Sci. USA. 2019, 116, 7916–7925. [Google Scholar] [CrossRef]
- Bowie, R.C.K.; Fjeldså, J. Superfamily Passeroidea. In troduction and the early lineages. In The Largest Avian Radiation. The Evolution of Perching Birds, or the Order Passeriformes; Fjeldså, J., Christidis, L., Ericson, P.G.P., Eds.; Lynx: Barcelona, Spain, 2020; pp. 263–275. [Google Scholar]
- Hickerson, M.J.; Stahl, E.; Lessios, H.A. Test for simultaneous divergence using approximate Bayesian computation. Evolution 2006, 60, 2435–2453. [Google Scholar] [CrossRef]
- Hickerson, M.J.; Meyer, C.P. Testing comparative phylogeographic models of marine vicariance and dispersal using a hierarchical Bayesian approach. BMC Evol. Biol. 2008, 8, 322. [Google Scholar] [CrossRef]
- Hickerson, M.J.; Stahl, E.; Takebayashi, N. MSBAYES: Pipeline for testing comparative phylogeo-graphic histories using hierarchical approximate Bayesiancomputation. BMC Bioinform. 2007, 8, 268. [Google Scholar] [CrossRef]
- Huang, W.; Takebayashi, N.; Hickerson, M.J. MTML-msBayes: Approximate Bayesian comparative phylogeographic inference from multiple taxa and multiple loci with rate heterogeneity. BMC Bioinform. 2011, 12, 1. [Google Scholar] [CrossRef]
- Beaumont, M.A.; Zhang, W.; Blading, D.J. Approximate Bayesian computation in population genetics. Genetics 2002, 16, 2025–2035. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 1 March 2022).
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.3-7. 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 11 March 2022).
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the Earth land surface areas. Sci. Data. 2017, 4, 170122. [Google Scholar] [CrossRef] [PubMed]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Data from: Climatologies at High Resolution for the Earth’s Land Surface Areas, Dryad, Dataset; Dryad: Lewis, WA, USA, 2018. [Google Scholar] [CrossRef]
- EAMCEF. Eastern Arc Mountains GIS Data. 2020. Available online: http://www.easternarc.or.tz/downloads/ (accessed on 1 March 2022).
- Karger, D.N.; Nobis, M.P.; Normand, S.; Graham, C.H.; Zimmermann, N.E. CHELSA-TraCE21k v1. 0. Downscaled transient temperature and precipitation data since the last glacial maximum. Clim. Past Discuss. 2021, 19, 439–456. [Google Scholar] [CrossRef]
- Fjeldså, J.; Christidis, L.; Ericson, P.G.P. The Largest Avian Radiation. In The Evolution of Perching Birds, or the Order Passeriformes; Lynx: Barcelona, Spain, 2020. [Google Scholar]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Stephens, M.; Smith, N.J.; Donnelly, P. A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Genet. 2001, 68, 978–989. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Fu, Y.-X. Statistical tests of neutrality of mutations against population growth, hitchhicking, and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Watterson, G.A. On the number of segregating sites in genetical models without recombination. Theor. Popul. Biol. 1975, 7, 256–276. [Google Scholar] [CrossRef]
- Burgess, N.D.; Clarke, G.P. Coastal Forests of Eastern Africa; IUCN The World Conservation Union: Gland, Switzerland, 2000. [Google Scholar]
- Lovett, J.C. Climatic history and forest distribution in eastern Africa. In Biogeography & Ecology of the Rain Forests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge Univ. Press: Cambridge, UK, 1993; pp. 23–29. [Google Scholar]
- Bowie, R.C.K.; Fjeldså, J. Genetic and morphological evidence for two species in the Udzungwa Forest Partridge Xenoperdix udzungwensis. J. East Afr. Nat. Hist. 2005, 94, 191–201. [Google Scholar] [CrossRef]
- Sepulchre, P.; Ramstein, G.; Fluteau, F.; Schuster, M.; Tiercelin, J.-J.; Brunet, M. Tectonic uplift and eastern Africa aridification. Science 2006, 313, 1419–1423. [Google Scholar] [CrossRef]
- Crowe, T.M.; Bowie, R.C.K.; Bloomer, P.; Mandiwana, T.; Hedderson, T.; Randi, E.; Pereira, S.L.; Wakeling, J. Phylogenetics and biogeography of, and character evolution in gamebirds (Aves: Galliformes): Effects of character exclusion, partitioning and missing data. Cladistics 2006, 22, 495–532. [Google Scholar] [CrossRef]
- Nicholson, S.E. The nature of rainfall variability over Africa on time scales of decades to millennia. Glob. Planet. Chang. 2000, 26, 137–158. [Google Scholar] [CrossRef]
- Lyons, R.P.; Scholz, C.A.; Cohen, A.S.; King, J.W.; Brown, E.T.; Ivory, S.J.; Johnson, T.C.; Deino, A.L.; Reinthal, P.N.; McGlue, M.M.; et al. Continuous 1.3-million-year record of East African hydroclimate, and implications for patterns of evolution and biodiversity. Proc. Natl. Acad. Sci. USA 2015, 112, 15568–15573. [Google Scholar] [CrossRef] [PubMed]
- Prell, W.I.; Hutson, W.H.; Williams, D.F.; Bé, A.W.H.; Geitzenauer, K.; Molfino, B. Surface circulation of the Indian Ocean during the Last Glacial Maximum, approximately 18.000 yr BP. Quat. Res. 1980, 14, 309–336. [Google Scholar] [CrossRef]
- Burney, C.W.; Brumfield, R.T. Ecology predicts levels of genetic differentiation in Neotropical birds. Am. Nat. 2009, 174, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Flantua, S.G.A.; O’Dea, A.; Onstein, R.E.; Giraldo, C.; Hooghiemstra, H. The flickering connectivity system of the northern Andean páramos. J. Biogeogr. 2018, 46, 1808–1825. [Google Scholar] [CrossRef]
- Bell, R.C.; MacKenzie, J.B.; Hickerson, M.J.; Chavarría, K.L.; Cunningham, M.; Williams, S.; Moritz, C. Comparative multi-locus phylogeography confirms multiple vicariance events in co-distributed rainforest frogs. Proc. R. Soc. B Biol. Sci. 2012, 279, 991–999. [Google Scholar] [CrossRef] [PubMed]
- Carnaval, A.C.; Hickerson, M.J.; Haddad, C.F.B.; Rodrigues, M.T.; Moritz, C. Stability predicts genetic diversity in the Brazilian Atlantic forest hotspot. Science 2009, 323, 785–789. [Google Scholar] [CrossRef]
- Dolman, G.; Joseph, L. A species assemblage approach to comparative phylogeography of birds in southern Australia. Ecol. Evol. 2012, 2, 354–369. [Google Scholar] [CrossRef]
- Leaché, A.D.; Crews, S.C.; Hickerson, M.J. Two waves of diversification in mammals and reptiles of Baja California revealed by hierarchical Bayesian analysis. Biol. Lett. 2007, 3, 646–650. [Google Scholar] [CrossRef]
- Benkman, C.W.; Smith, J.W.; Keenan, P.C.; Parchman, T.L.; Santisteban, L. A new species of the Red Crossbill (Fringillidae: Loxia) from Idaho. Condor 2019, 111, 169–176. [Google Scholar] [CrossRef]
- Cheek, R.G.; Forester, B.R.; Salerno, P.E.; Trumbo, D.R.; Langin, K.M.; Chen, N.; Sillett, T.S.; Morrison, S.A.; Ghalambor, C.K.; Funk, W.C. Habitat-linked genetic variation supports microgeographic adaptive divergence in an islands-endemic bird species. Mol. Ecol. 2022, 31, 2830–2846. [Google Scholar] [CrossRef] [PubMed]
- Sorenson, M.D.; Sefc, K.M.; Payne, R.B. Speciation by host switch in brood parasitic indigobirds. Nature 2003, 424, 928–931. [Google Scholar] [CrossRef]
- Stanley, W.T.; Olson, E. Phylogeny, Phylogeography, and geographic variation of Sylvisorex howelli (Soricidae), an endemic shrew of the Eastern Arc Mountains, Tanzania. J. Zool. 2005, 266, 341–354. [Google Scholar] [CrossRef]
- Taylor, P.J.; Maree, S.; Van Sandwyk, J.; Kerbis Peterhans, J.C.; Stanley, W.T.; Verheyen, E.; Kaliba, P.; Verheyen, W.; Kaleme, P.; Nigel, C. Speciation mirrors geomorphology and palaeoclimatic history in African laminate-toothed rats (Muridae: Otomyini) of the Otomys denti and Otomys lacustris species-complexes in the ‘Montane Circle’ of East Africa. Biol. J. Linn. Soc. 2009, 96, 913–941. [Google Scholar] [CrossRef]
- Voelker, G.; Huntley, J.W.; Bryja, J.; Deny, C.; Ŝumbera, R.; Demos, T.; Lavrenchenko, L.; Nicolas, V.; Gnoske, T.; Peterhans, J.K. Molecular systematics and biogeographic history of the African climbing-mouse complex (Dendromus). Mol. Phylogenet. Syst. 2021, 161, 107166. [Google Scholar] [CrossRef]
- Blackburn, D.C.; Measey, G.J. Dispersal to or from an African biodiversity hotspot. Mol. Ecol. 2009, 9, 1904–1915. [Google Scholar] [CrossRef]
- Lawson, L.P. The discordance of diversification: Evolution in the tropical-montane frogs of the Eastern Arc Mountains of Tanzania. Mol. Ecol. 2010, 19, 4046–4060. [Google Scholar] [CrossRef]
- Tolley, K.A.; Tilbury, C.R.; Measey, G.J.; Menegon, M.; Branch, W.R.; Matthee, C.A. Ancient forest fragmentation or recent radiation? Testing refugial speciation models in chameleons within an African biodiversity hotspot. J. Biogeogr. 2011, 38, 1748–1760. [Google Scholar] [CrossRef]
- Voje, K.L.; Hemp, C.; Flagstad, Ø.; Sætre, G.-P.; Stenseth, N.C. Climate change as an engine for speciation in flightless Orthoptera inhabiting African mountains. Mol. Ecol. 2009, 18, 93–108. [Google Scholar]
- Moeller, M.; Cronk, Q. Phylogeny and disjunct distribution: Evolution of Saintpaulia (Gesneriaceae). Proc. R. Soc. B Biol. Sci. 1998, 264, 1827–1836. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, D.; Nogués-Bravo, D.; Scharff, N. Why do tropical mountains support exceptionally high biodiversity? The Eastern Arc Mountains and the drivers of Saintpaulina diversity. PLoS ONE 2012, 7, e48908. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.L.; Smith, M.F. MTDNA phylogeny of Andean mice: A test of diversification across ecological gradients. Evolution 1992, 46, 174–183. [Google Scholar] [CrossRef]
- Caro, L.M.; Caycedo-Rosales, P.C.; Bowie, R.C.K.; Slabbekoorn, H.; Cadena, C.D. Ecological speciation along an elevational gradient in a tropical passerine bird? J. Evol. Biol. 2013, 26, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Cadena, C.K.; Cespedes, L.N. Origin of elevational replacements in a clade of nearly flighless birds: Most diversity in tropical mountains accumulates via secondary contact following allopatric speciation. In Neotropical Speciation; Rull, V., Carnaval, A.C., Eds.; Springer: Berlin, Germany, 2020; pp. 635–639. [Google Scholar]
- Cadena, C.D.; Kozak, K.H.; Gómez, J.P.; Parra, J.L.; McCain, C.M.; Bowie, R.; Carnaval, A.; Moritz, C.; Rahbek, C.; Roberts, T.E.; et al. Latitude, elevational climatic zonation and speciation in New World vertebrates. Proc. R. Soc. B Biol. Sci. 2012, 279, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Fjeldså, J.; Alström, P.; Bowie, R.C.K. How new species evolve. In The Evolution of Perching Birds, or the Order Passeriformes; Fjeldså, J., Christidis, L., Ericson, P.G.P., Eds.; Lynx: Barcelona, Spain, 2020. [Google Scholar]
- Freeman, B.G. Lower elevation animal species do not tend to be better competitors than their higher elevation relatives. Glob. Ecol. Biogeogr. 2020, 29, 171–181. [Google Scholar] [CrossRef]
- Vianna, J.A.; Fernandes, F.A.N.; Frugone, M.J.; Figueiró, H.; Pertierra, L.R.; Noll, D.; Bi, K.; Lowther, A.; Parker, P.; Wienecke, B.; et al. Genome-wide analyses reveal drivers of penguin diversification. Proc. Nat. Acad. Sci. USA 2020, 117, 22303–22310. [Google Scholar] [CrossRef]
- Robinson, J.A.; Bowie, R.C.K.; Dudchenko, O.; Lieberman, A.E.; Hendrickson, S.L.; Steiner, C.C.; Ryder, O.A.; Mindell, D.P.; Wall, J.D. Genome-wide diversity of the California Condor tracks its prehistoric abundance and decline. Curr. Biol. 2021, 31, 2939–2946. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species Complex | Species | Habitat | Foraging | Diet | n | Source—Molecular Data |
|---|---|---|---|---|---|---|---|
| Lybiidae | Green Barbet | Stactolaema olivacea | Montane & Lowland Forest | Mid-stratum & Canopy | Fruit | 84 | [8] |
| Cisticolidae | Red-capped Forest Warbler | Artisornis metopias | Montane Forest | Understorey | Insects | 119 | [39] |
| Fringillidae | Streaky & Yellow-browed Seedeater | Crithagra striolata, C. whytii | Montane Forest | Understorey | Insects | 63 | [40] |
| Modulatricidae | Spot-throat | Modulatrix stictigula | Montane Forest | Understorey | Insects | 106 | MS |
| Muscicapidae | Sharpe’s Akalat | Sheppardia sharpei | Montane Forest | Understorey | Insects | 44 | [8] |
| Muscicapidae | Usambara Akalat complex | Sheppardia montana, S. lowei, S. aurantiithorax | Montane Forest | Understorey | Insects | 13 | [41] |
| Muscicapidae | White-chested Alethe | Chamaetylas fuelleborni | Montane & Lowland Forest | Understorey | Insects | 75 | [8] |
| Muscicapidae | White-starred Robin | Pogonocichla stellata | Montane & Lowland Forest | Understorey & Mid-Stratum | Insects | 283 | [42] |
| Nectariniidae | Eastern Double-collared Sunbird complex | Cinnyris mediocris, C. usambarica, C. moreaui, C. loveridgei, C. fuelleborni | Montane Forest | Understorey & Mid-Stratum | Nectar & Insects | 256 | [43,44,45] |
| Nectariniidae | Olive Sunbird | Cyanomitra olivacea | Montane & Lowland Forest | Understorey & Mid-Stratum | Nectar & Insects | 282 | [46] |
| Platysteridae | Cape Batis complex | Batis mixta, B. crypta, B. dimorpha | Montane & Lowland Forest | Understorey & Mid-Stratum | Insects | 238 | [8,47,48] |
| Pycnonotidae | Stripe-cheeked Greenbul complex | Arizelocichla milanjensis, A. striifacies, A. olivaceiceps | Montane Forest | Mid-stratum & Canopy | Insects & Fruit | 149 | [48,49] |
| Pycnonotidae | Yellow-streaked Greenbul | Phyllastrephus flavostriatus | Montane & Lowland Forest | Mid-stratum & Canopy | Insects & Occasionally Fruit | 248 | [8,50] |
| Turdidae | Abyssinian Thrush complex | Turdus abyssinicus, T. roehi, T. helleri | Montane Forest | Understorey & Mid-Stratum | Insects & Fruit | 39 | [51,52] |
| Marker | Sky Island | Stability * | Summary Statistics | |||
|---|---|---|---|---|---|---|
| Yellow-Streaked Greenbul (Phyllastrephus flavostriatus) | n | h | π × 103 | θ(s) | ||
| Autosomal | Usambara | Stable | 20 | 0.628 | 2.10 | 1.43 |
| (4 loci) | Nguru | Reduced | 24 | 0.531 | 1.84 | 1.32 |
| Udzungwa | Stable | 26 | 0.646 | 2.62 | 1.97 | |
| STZ Volcanic | Unstable | 18 | 0.270 | 0.67 | 0.42 | |
| Z-linked | Usambara | Stable | 20 | 0.509 | 4.50 | 2.21 |
| (2 loci) | Nguru | Reduced | 20 | 0.358 | 1.07 | 0.56 |
| Udzungwa | Stable | 24 | 0.641 | 2.15 | 1.09 | |
| STZ Volcanic | Unstable | 18 | 0.270 | 0.67 | 0.42 | |
| mtDNA | Usambara | Stable | 21 | 0.857 | 3.76 | 5.56 |
| Nguru | Reduced | 39 | 0.629 | 0.78 | 2.36 | |
| Udzungwa | Stable | 30 | 0.825 | 1.28 | 3.28 | |
| STZ Volcanic | Unstable | 10 | 0.533 | 0.51 | 0.35 | |
| Spot-throat (Modulatrix stictigula) | ||||||
| Autosomal | Usambara | Stable | 18 | 0.816 | 3.92 | 2.26 |
| (2 loci) | Nguru | Reduced | 10 | 0.752 | 4.82 | 2.01 |
| Rubeho | Stable | 26 | 0.819 | 5.23 | 3.34 | |
| Uluguru | Stable | 17 | 0.546 | 1.40 | 0.77 | |
| Udzungwa | Stable | 18 | 0.740 | 4.75 | 2.95 | |
| mtDNA | Usambara | Stable | 19 | 0.836 | 3.02 | 4.00 |
| Nguru | Reduced | 15 | 0.819 | 2.54 | 2.46 | |
| Rubeho | Stable | 26 | 0.905 | 2.57 | 2.62 | |
| Uluguru | Stable | 24 | 0.830 | 1.32 | 1.87 | |
| Udzungwa | Stable | 19 | 0.879 | 1.51 | 2.54 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bowie, R.C.K.; Monahan, W.B.; Fjeldså, J. Climate Cycles, Habitat Stability, and Lineage Diversification in an African Biodiversity Hotspot. Diversity 2023, 15, 394. https://doi.org/10.3390/d15030394
Bowie RCK, Monahan WB, Fjeldså J. Climate Cycles, Habitat Stability, and Lineage Diversification in an African Biodiversity Hotspot. Diversity. 2023; 15(3):394. https://doi.org/10.3390/d15030394
Chicago/Turabian StyleBowie, Rauri C. K., William B. Monahan, and Jon Fjeldså. 2023. "Climate Cycles, Habitat Stability, and Lineage Diversification in an African Biodiversity Hotspot" Diversity 15, no. 3: 394. https://doi.org/10.3390/d15030394
APA StyleBowie, R. C. K., Monahan, W. B., & Fjeldså, J. (2023). Climate Cycles, Habitat Stability, and Lineage Diversification in an African Biodiversity Hotspot. Diversity, 15(3), 394. https://doi.org/10.3390/d15030394