

A Snapshot into the Lives of Elephants: Camera Traps and Conservation in Etosha National Park, Namibia

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Camera Trap Photo Collection
2.3. Data Collection from Photos
2.4. Statistical Analysis
3. Results
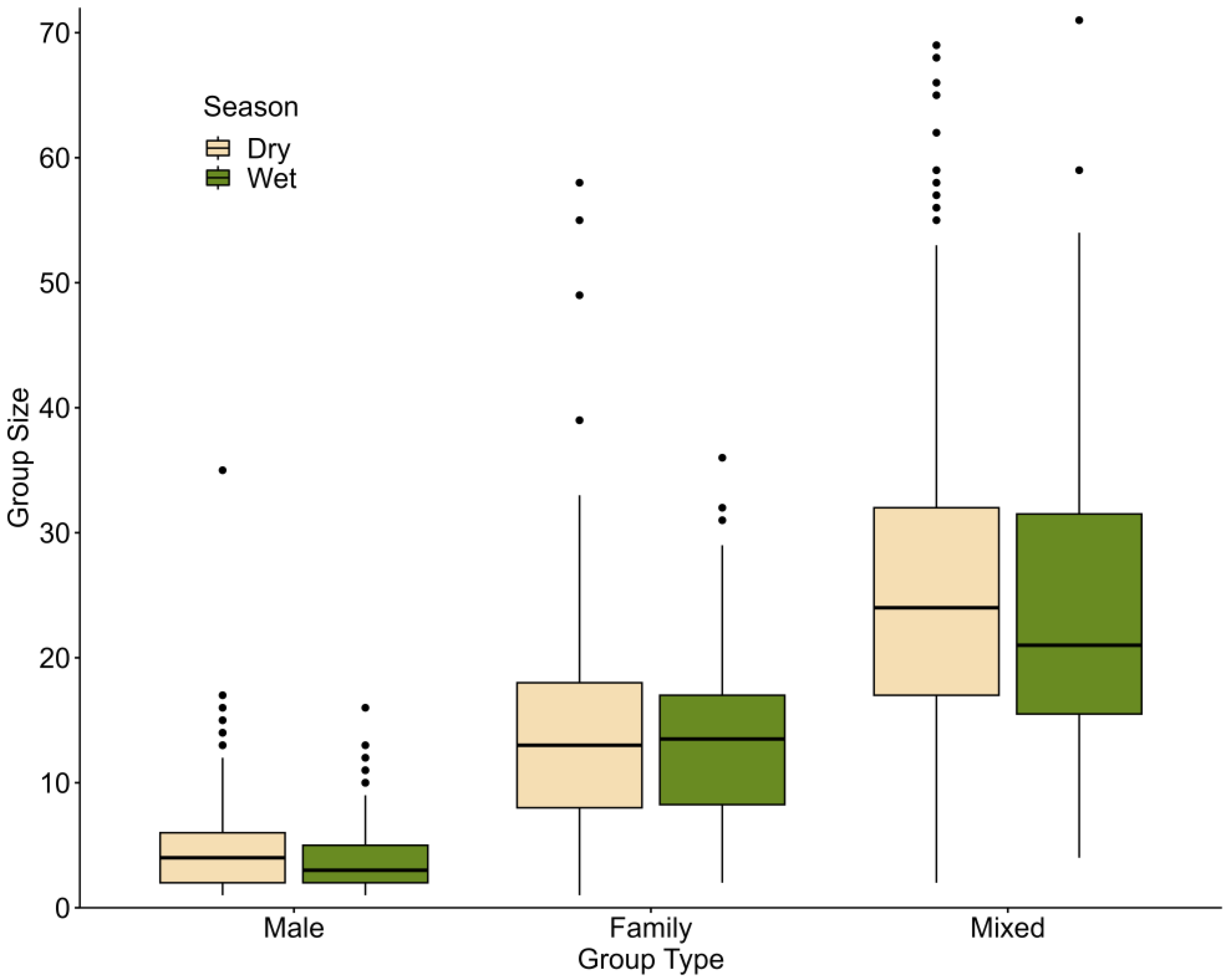
3.1. Seasonal Variation in Group Occurrence and Size
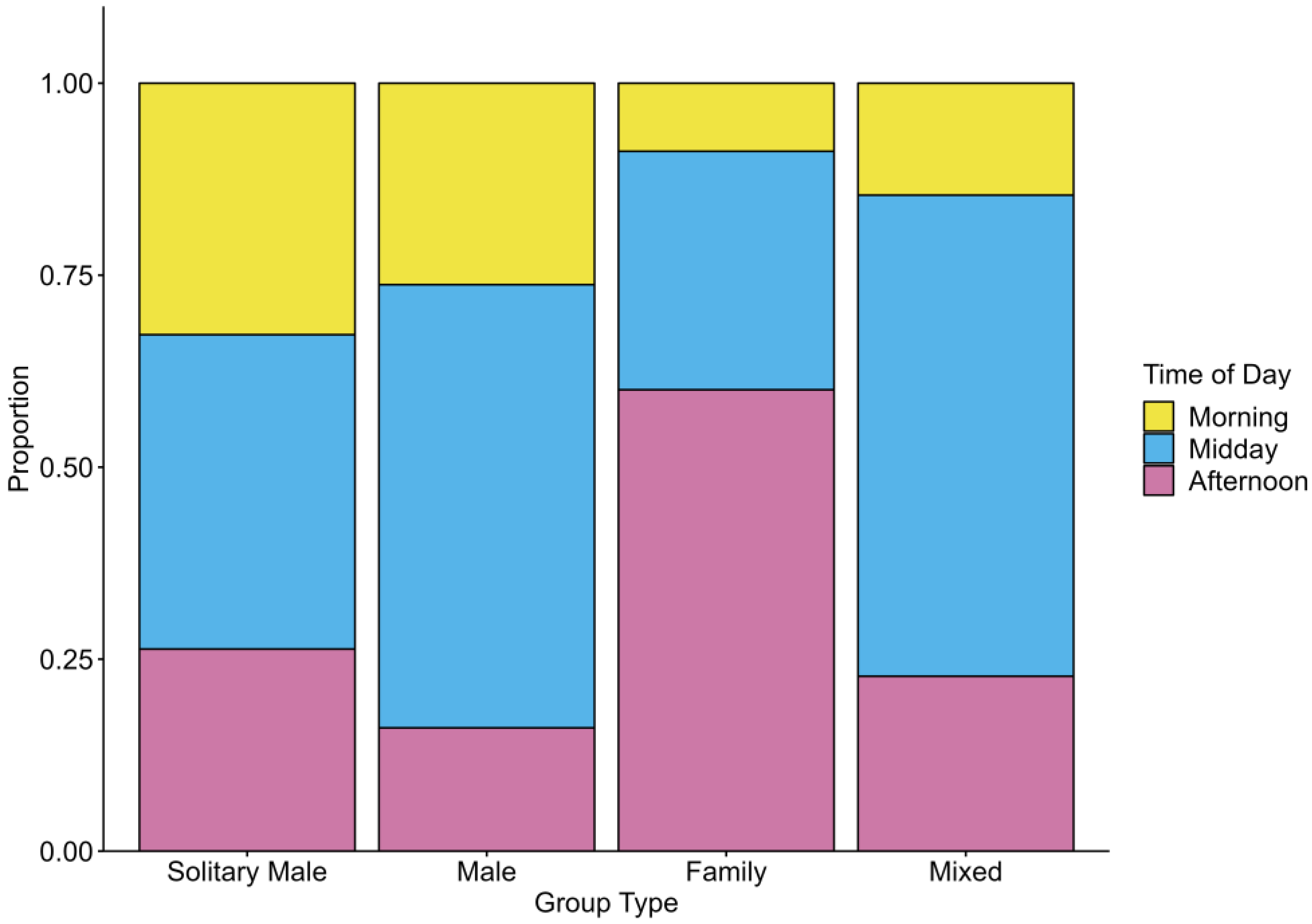
3.2. Diurnal Variation in Group Occurrence
3.3. Observer Reliability
4. Discussion
4.1. Seasonal Grouping Patterns
4.2. Diurnal Grouping Patterns
4.3. Undergraduate Students as Reliable Observers in Camera Trap Photo Data Collection
4.4. Additional Considerations of Camera Trap Methodology
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Type | Season | Time of Day | Total Photos per Season | ||
|---|---|---|---|---|---|
| Morning | Midday | Afternoon | |||
| Solitary male (n = 1197) | Wet | 208 | 228 | 91 | 527 |
| Dry | 184 | 262 | 224 | 670 | |
| Male (n = 2043) | Wet | 258 | 337 | 77 | 672 |
| Dry | 278 | 842 | 251 | 1371 | |
| Family (n = 248) | Wet | 6 | 11 | 41 | 58 |
| Dry | 16 | 66 | 108 | 190 | |
| Mixed (n = 878) | Wet | 42 | 63 | 34 | 139 |
| Dry | 86 | 487 | 166 | 739 | |
References
- Ahlering, M.A.; Maldonado, J.E.; Fleischer, R.C.; Western, D.; Eggert, L.S. Fine-Scale Group Structure and Demography of African Savanna Elephants Recolonizing Lands Outside Protected Areas: Demography of Elephants Outside Parks. Divers. Distrib. 2012, 18, 952–961. [Google Scholar] [CrossRef]
- Benitez, L.; Kilian, J.W.; Wittemyer, G.; Hughey, L.F.; Fleming, C.H.; Leimgruber, P.; du Preez, P.; Stabach, J.A. Precipitation, Vegetation Productivity, and Human Impacts Control Home Range Size of Elephants in Dryland Systems in Northern Namibia. Ecol. Evol. 2022, 12, e9288. [Google Scholar] [CrossRef] [PubMed]
- Bohrer, G.; Beck, P.S.; Ngene, S.M.; Skidmore, A.K.; Douglas-Hamilton, I. Elephant Movement Closely Tracks Precipitation-Driven Vegetation Dynamics in a Kenyan Forest-Savanna Landscape. Mov. Ecol. 2014, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Craig, G.C.; Gibson, D.S.C.; Uiseb, K.H. Namibia’s Elephants—Population, Distribution and Trends. Pachyderm 2021, 62, 35–52. Available online: https://pachydermjournal.org/index.php/pachyderm/article/view/460 (accessed on 12 November 2023).
- de Beer, Y.; van Aarde, R.J. Do Landscape Heterogeneity and Water Distribution Explain Aspects of Elephant Home Range in Southern Africa’s Arid Savannas? J. Arid Environ. 2008, 72, 2017–2025. [Google Scholar] [CrossRef]
- Gill, B.A.; Wittemyer, G.; Cerling, T.E.; Musili, P.M.; Kartzinel, T.R. Foraging History of Individual Elephants Using DNA Metabarcoding. R. Soc. Open Sci. 2023, 10, 230337. [Google Scholar] [CrossRef]
- Harris, G.M.; Russell, G.J.; van Aarde, R.I.; Pimm, S.L. Rules of Habitat Use by Elephants Loxodonta africana in Southern Africa: Insights for Regional Management. Oryx 2008, 42, 66–75. [Google Scholar] [CrossRef]
- Kioko, J.; Zink, E.; Sawdy, M.; Kiffner, C. Elephant (Loxodonta africana) Demography and Behaviour in the Tarangire-Manyara Ecosystem, Tanzania. S. Afr. J. Wildl. Res. 2013, 43, 44–51. [Google Scholar] [CrossRef]
- Smit, J.B.; Searle, C.E.; Buchanan-Smith, H.M.; Strampelli, P.; Mkuburo, L.; Kakengi, V.A.; Kohi, E.M.; Dickman, A.J.; Lee, P.C. Anthropogenic Risk Increases Night-time Activities and Associations in African Elephants (Loxodonta africana) in the Ruaha-Rungwa Ecosystem, Tanzania. Afr. J. Ecol. 2022, 61, 64–76. [Google Scholar] [CrossRef]
- Tsalyuk, M.; Kilian, W.; Reineking, B.; Getz, W.M. Temporal Variation in Resource Selection of African Elephants Follows Long-term Variability in Resource Availability. Ecol. Monogr. 2019, 89, e01348. [Google Scholar] [CrossRef]
- Wittemyer, G.; Polansky, L.; Douglas-Hamilton, I.; Getz, W.M. Disentangling the Effects of Forage, Social Rank, and Risk on Movement Autocorrelation of Elephants Using Fourier and Wavelet Analyses. Proc. Natl. Acad. Sci. USA 2008, 105, 19108–19113. [Google Scholar] [CrossRef] [PubMed]
- Wittemyer, G.; Getz, W.M.; Vollrath, F.; Douglas-Hamilton, I. Social Dominance, Seasonal Movements, and Spatial Segregation in African Elephants: A Contribution to Conservation Behavior. Behav. Ecol. Sociobiol. 2007, 61, 1919–1931. [Google Scholar] [CrossRef]
- Beirne, C.; Houslay, T.M.; Morkel, P.; Clark, C.J.; Fay, M.; Okouyi, J.; White, L.J.T.; Poulsen, J.R. African Forest Elephant Movements Depend on Time Scale and Individual Behavior. Sci. Rep. 2021, 11, 12634. [Google Scholar] [CrossRef]
- King, L.; Raja, N.; Kumar, M.; Heath, N.; Pope, F. Part I: Development of a New Human–Elephant Coexistence Toolbox for Communities Living with African Savannah Elephants (Loxodonta africana). Pachyderm 2022, 63, 153–157. Available online: https://pachydermjournal.org/index.php/pachyderm/article/view/489 (accessed on 12 November 2023).
- Mumby, H.S.; Plotnik, J.M. Taking the Elephants’ Perspective: Remembering Elephant Behavior, Cognition and Ecology in Human-Elephant Conflict Mitigation. Front. Ecol. Evol. 2018, 6, 122. [Google Scholar] [CrossRef]
- Caravaggi, A.; Banks, P.B.; Burton, A.C.; Finlay, C.M.V.; Haswell, P.M.; Hayward, M.W.; Rowcliffe, M.J.; Wood, M.D. A Review of Camera Trapping for Conservation Behaviour Research. Remote Sens. Ecol. Conserv. 2017, 3, 109–122. [Google Scholar] [CrossRef]
- Fazio, J.M.; Barthel, T.; Freeman, E.W.; Garlick-Ott, K.; Scholle, A.; Brown, J.L. Utilizing Camera Traps, Closed Circuit Cameras and Behavior Observation Software to Monitor Activity Budgets, Habitat Use, and Social Interactions of Zoo-Housed Asian Elephants (Elephas Maximus). Animals 2020, 10, 2026. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.; Clark, N.; Rendle-Worthington, J.; Yon, L. Behaviour and Welfare Impacts of Releasing Elephants from Overnight Tethers: A Zimbabwean Case Study. Animals 2022, 12, 1933. [Google Scholar] [CrossRef]
- Cardoso, A.W.; Malhi, Y.; Oliveras, I.; Lehmann, D.; Ndong, J.E.; Dimoto, E.; Bush, E.; Jeffery, K.; Labriere, N.; Lewis, S.L.; et al. The Role of Forest Elephants in Shaping Tropical Forest–Savanna Coexistence. Ecosystems 2020, 23, 602–616. [Google Scholar] [CrossRef]
- von Hagen, R.L. An Evaluation of Deterrent Methods Utilized to Prevent Crop Raiding by African Elephants (Loxodonta africana) in the Kasigau Wildlife Corridor, Kenya. Master’s Thesis, Western Kentucky University, Bowling Green, KY, USA, 2018. [Google Scholar]
- Varma, S.; Pittet, A.; Jamadagni, H.S. Experimenting Usage of Camera-Traps for Population Dynamics Study of the Asian Elephant Elephas Maximus in Southern India. Mapana J. Sci. 2007, 6, 81–95. [Google Scholar] [CrossRef]
- Zeppelzauer, M.; Stoeger, A.S. Establishing the Fundamentals for an Elephant Early Warning and Monitoring System. BMC Res. Notes 2015, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.R.B.; Brent, L.J.N.; Motsentwa, T.; Weiss, M.N.; Croft, D.P. Importance of Old Bulls: Leaders and Followers in Collective Movements of All-Male Groups in African Savannah Elephants (Loxodonta africana). Sci. Rep. 2020, 10, 13996. [Google Scholar] [CrossRef] [PubMed]
- Karczmarski, L.; Chan, S.C.Y.; Rubenstein, D.I.; Chui, S.Y.S.; Cameron, E.Z. Individual Identification and Photographic Techniques in Mammalian Ecological and Behavioural Research—Part 1: Methods and Concepts. Mamm. Biol. 2022, 102, 545–549. [Google Scholar] [CrossRef]
- de Silva, E.M.K.; Kumarasinghe, P.; Indrajith, K.K.D.A.K.; Pushpakumara, T.V.; Vimukthi, R.D.Y.; de Zoysa, K.; Gunawardana, K.; de Silva, S. Feasibility of Using Convolutional Neural Networks for Individual-Identification of Wild Asian Elephants. Mamm. Biol. 2022, 102, 931–941. [Google Scholar] [CrossRef]
- Webb, J.L.; Crawley, J.A.H.; Seltmann, M.W.; Liehrmann, O.; Hemmings, N.; Nyein, U.K.; Aung, H.H.; Htut, W.; Lummaa, V.; Lahdenperä, M. Evaluating the Reliability of Non-Specialist Observers in the Behavioural Assessment of Semi-Captive Asian Elephant Welfare. Animals 2020, 10, 167. [Google Scholar] [CrossRef]
- Loarie, S.R.; Aarde, R.J.V.; Pimm, S.L. Fences and Artificial Water Affect African Savannah Elephant Movement Patterns. Biol. Conserv. 2009, 142, 3086–3098. [Google Scholar] [CrossRef]
- Wiśniewska, M.; O’Connell-Rodwell, C.E.; Kilian, W.; Garnier, S.; Russell, G.J. Interactions between Physical and Social Drivers of Movement in Male African Savanna Elephants. bioRxiv, 2023; submitted. [Google Scholar]
- van Aarde, R.J.; Jackson, T.P.; Ferreira, S. Conservation Science and Elephant Management in Southern Africa. S. Afr. J. Sci. 2006, 102, 385–388. [Google Scholar]
- Thouless, C.R.; Dublin, H.T.; Blanc, J.J.; Skinner, D.P.; Daniels, T.E.; Taylor, R.D.; Maisels, F.; Fredrick, H.L.; Bouché, P. African Elephant Status Report 2016: An Update from the African Elephant Database; IUCN/SSC African Elephant Specialist Group: Gland, Switzerland, 2016. [Google Scholar]
- Thurber, M.I.; O’Connell-Rodwell, C.E.; Turner, W.C.; Nambandi, K.; Kinzley, C.; Rodwell, T.C.; Faulkner, C.T.; Felt, S.A.; Bouley, D.M. Effects of Rainfall, Host Demography, and Musth on Strongyle Fecal Egg Counts in African Elephants (Loxodonta africana) in Namibia. J. Wildl. Dis. 2011, 47, 172–181. [Google Scholar] [CrossRef]
- O’Connell-Rodwell, C.E.; Freeman, P.T.; Kinzley, C.; Sandri, M.N.; Berezin, J.L.; Wiśniewska, M.; Jessup, K.; Rodwell, T.C. A Novel Technique for Aging Male African Elephants (Loxodonta africana) Using Craniofacial Photogrammetry and Geometric Morphometrics. Mamm. Biol. 2022, 102, 591–613. [Google Scholar] [CrossRef]
- O’Connell-Rodwell, C.E.; Sandri, M.N.; Berezin, J.L.; Munevar, J.M.; Kinzley, C.; Wood, J.D.; Wiśniewska, M.; Kilian, J.W. Male African Elephant (Loxodonta africana) Behavioral Responses to Estrous Call Playbacks May Inform Conservation Management Tools. Animals 2022, 12, 1162. [Google Scholar] [CrossRef]
- O’Connell-Rodwell, C.E.; Wood, J.D.; Kinzley, C.; Rodwell, T.C.; Alarcon, C.; Wasser, S.K.; Sapolsky, R. Male African Elephants (Loxodonta africana) Queue When the Stakes Are High. Ethol. Ecol. Evol. 2011, 23, 388–397. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2023. Available online: https://www.R-project.org/ (accessed on 12 November 2023).
- McHugh, M.L. Interrater Reliability: The Kappa Statistic. Biochem. Med. 2012, 22, 276–282. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3900052/ (accessed on 12 November 2023). [CrossRef]
- Gamer, M.; Lemon, J.; Fellows, I.; Singh, P. Irr: Various Coefficients of Interrater Reliability and Agreement. R package version 0.84.1. 2012. Available online: https://CRAN.R-project.org/package=irr (accessed on 12 November 2023).
- Koo, T.; Li, M. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Fleiss, J.L. Measuring Nominal Scale Agreement among Many Raters. Psychol. Bull. 1971, 76, 378–382. [Google Scholar] [CrossRef]
- Landis, J.R.; Koch, G.G. The Measurement of Observer Agreement for Categorical Data. Biometrics 1977, 33, 159. [Google Scholar] [CrossRef]
- RStudio Posit Team. RStudio: Integrated Development Environment for R. Posit Software, PBC, Boston, Massachusetts. 2023. Available online: https://posit.co/products/open-source/rstudio/ (accessed on 12 November 2023).
- du Plessis, K.; Ganswindt, S.B.; Bertschinger, H.; Crossey, B.; Henley, M.D.; Ramahlo, M.; Ganswindt, A. Social and Seasonal Factors Contribute to Shifts in Male African Elephant (Loxodonta africana) Foraging and Activity Patterns in Kruger National Park, South Africa. Animals 2021, 11, 3070. [Google Scholar] [CrossRef]
- Shannon, G.; Mackey, R.L.; Slotow, R. Diet Selection and Seasonal Dietary Switch of a Large Sexually Dimorphic Herbivore. Acta Oecolog. 2013, 46, 48–55. [Google Scholar] [CrossRef]
- Stokke, S.; Du Toit, J.T. Sexual Segregation in Habitat Use by Elephants in Chobe National Park, Botswana: Sexual Segregation in Habitat Use by Elephants. Afr. J. Ecol. 2002, 40, 360–371. [Google Scholar] [CrossRef]
- Moss, C.J.; Lee, P.C. Female Social Dynamics: Fidelity and Flexibility. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; The University of Chicago Press: Chicago, IL, USA, 2011; pp. 205–223. [Google Scholar]
- Turner, W.C.; Périquet, S.; Goelst, C.E.; Vera, K.B.; Cameron, E.Z.; Alexander, K.A.; Belant, J.L.; Cloete, C.C.; Du Preez, P.; Getz, W.M.; et al. Africa’s Drylands in a Changing World: Challenges for Wildlife Conservation under Climate and Land-Use Changes in the Greater Etosha Landscape. Glob. Ecol. Conserv. 2022, 38, e02221. [Google Scholar] [CrossRef]
- Maúre, G.; Pinto, I.; Ndebele-Murisa, M.; Muthige, M.; Lennard, C.; Nikulin, G.; Dosio, A.; Meque, A. The Southern African Climate under 1.5 °C and 2 °C of Global Warming as Simulated by CORDEX Regional Climate Models. Environ. Res. Lett. 2018, 13, 065002. [Google Scholar] [CrossRef]
- Garstang, M.; Larom, D.; Raspet, R.; Lindeque, M. Atmospheric Controls on Elephant Communication. J. Exp. Biol. 1995, 198, 939–951. [Google Scholar] [CrossRef] [PubMed]



| Season | Month | Time of Day | Total Photos per Month (n = 4366) | ||
|---|---|---|---|---|---|
| Morning (n = 1078) | Midday (n = 2296) | Afternoon (n = 992) | |||
| Wet (n = 1396) | January | 107 | 101 | 36 | 244 |
| February | 4 | 8 | 6 | 18 | |
| March | 0 | 0 | 0 | 0 | |
| April | 10 | 26 | 31 | 67 | |
| November | 180 | 236 | 83 | 499 | |
| December | 213 | 268 | 87 | 568 | |
| Dry (n = 2970) | May | 34 | 154 | 97 | 285 |
| June | 35 | 148 | 153 | 336 | |
| July | 23 | 146 | 122 | 291 | |
| August | 63 | 364 | 162 | 589 | |
| September | 161 | 417 | 109 | 687 | |
| October | 248 | 428 | 106 | 782 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berezin, J.L.; Odom, A.J.; Hayssen, V.; O’Connell-Rodwell, C.E. A Snapshot into the Lives of Elephants: Camera Traps and Conservation in Etosha National Park, Namibia. Diversity 2023, 15, 1146. https://doi.org/10.3390/d15111146
Berezin JL, Odom AJ, Hayssen V, O’Connell-Rodwell CE. A Snapshot into the Lives of Elephants: Camera Traps and Conservation in Etosha National Park, Namibia. Diversity. 2023; 15(11):1146. https://doi.org/10.3390/d15111146
Chicago/Turabian StyleBerezin, Jodie L., Amanda J. Odom, Virginia Hayssen, and Caitlin E. O’Connell-Rodwell. 2023. "A Snapshot into the Lives of Elephants: Camera Traps and Conservation in Etosha National Park, Namibia" Diversity 15, no. 11: 1146. https://doi.org/10.3390/d15111146
APA StyleBerezin, J. L., Odom, A. J., Hayssen, V., & O’Connell-Rodwell, C. E. (2023). A Snapshot into the Lives of Elephants: Camera Traps and Conservation in Etosha National Park, Namibia. Diversity, 15(11), 1146. https://doi.org/10.3390/d15111146