

Chlamydiaceae-Like Bacterium in Wild Magellanic Penguins (Spheniscus magellanicus)

 , , , , and
, , , , and
Abstract
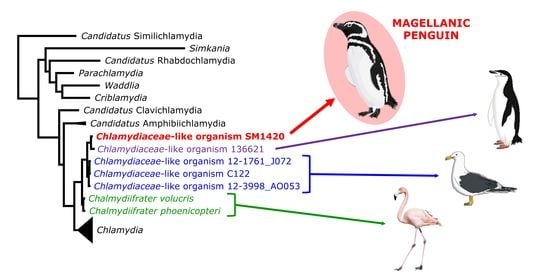
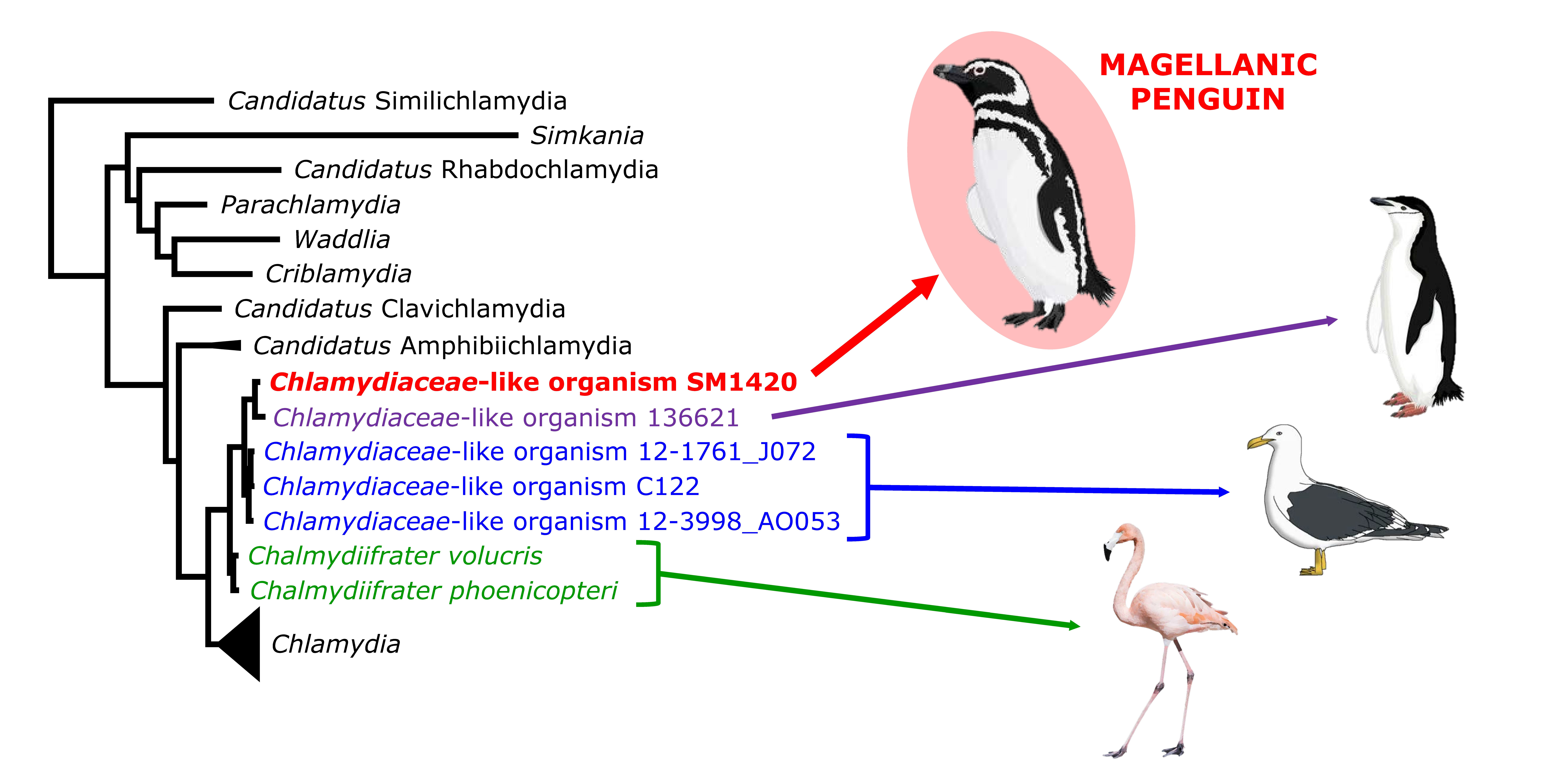{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Culture, DNA Extraction, qPCR and Sequencing
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, M.; Nylund, A.; Watanabe, K.; Helvik, J.V.; Nylund, S.; Plarre, H. Characterization of “Candidatus Clavochlamydia salmonicola”: An intracellular bacterium infecting salmonid fish. Environ. Microbiol. 2008, 10, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Martel, A.; Adriaensen, C.; Bogaerts, S.; Ducatelle, R.; Favoreel, H.; Crameri, S.; Hyatt, A.D.; Haesebrouck, F.; Pasmans, F. Novel Chlamydiaceae Disease in Captive Salamanders. Emerg. Infect. Dis. 2012, 18, 1020–1022. [Google Scholar] [CrossRef]
- Vorimore, F.; Hölzer, M.; Liebler-Tenorio, E.; Barf, L.-M.; Delannoy, S.; Vittecoq, M.; Wedlarski, R.; Lécu, A.; Scharf, S.; Blanchard, Y.; et al. Evidence for the existence of a new genus Chlamydiifrater gen. nov. inside the family Chlamydiaceae with two new species isolated from flamingo (Phoenicopterus roseus): Chlamydiifrater phoenicopteri sp. nov. and Chlamydiifrater volucris sp. nov. Syst. Appl. Microbiol. 2021, 44, 126200. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.M.; Hutton, S.; Myers, G.; Brunham, R.; Timms, P. Chlamydia pneumoniae Is Genetically Diverse in Animals and Appears to Have Crossed the Host Barrier to Humans on (At Least) Two Occasions. PLOS Pathog. 2010, 6, e1000903. [Google Scholar] [CrossRef]
- Sachse, K.; Kuehlewind, S.; Ruettger, A.; Schubert, E.; Rohde, G. More than classical Chlamydia psittaci in urban pigeons. Vet. Microbiol. 2012, 157, 476–480. [Google Scholar] [CrossRef]
- Harkinezhad, T.; Geens, T.; Vanrompay, D. Chlamydophila psittaci infections in birds: A review with emphasis on zoonotic consequences. Vet. Microbiol. 2009, 135, 68–77. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Taday, E.M.A. Avian host range of Chlamydophila spp. based on isolation, antigen detection and serology. Avian Pathol. 2003, 32, 435–462. [Google Scholar] [CrossRef]
- Sukon, P.; Nam, N.H.; Kittipreeya, P.; Sara-In, A.; Wawilai, P.; Inchuai, R.; Weerakhun, S. Global prevalence of chlamydial infections in birds: A systematic review and meta-analysis. Prev. Vet. Med. 2021, 192, 105370. [Google Scholar] [CrossRef]
- Andersen, A.; Vanrompay, D. Avian chlamydiosis (pisttacosis, ornithosis). In Diseases of Poultry; Iowa State University Press: Ames, IA, USA, 2003. [Google Scholar]
- Joseph, S.J.; Marti, H.; Didelot, X.; Castillo-Ramirez, S.; Read, T.D.; Dean, D. Chlamydiaceae Genomics Reveals Interspecies Admixture and the Recent Evolution of Chlamydia abortus Infecting Lower Mammalian Species and Humans. Genome Biol. Evol. 2015, 7, 3070–3084. [Google Scholar] [CrossRef]
- Herrmann, B.; Persson, H.; Jensen, J.-K.; Joensen, H.D.; Klint, M.; Olsen, B. Chlamydophila psittaci in Fulmars, the Faroe Islands. Emerg. Infect. Dis. 2006, 12, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Jencek, J.E.; Beaufrere, H.; Tully, T.N.; Garner, M.M.; Dunker, F.H.; Baszler, T.V. An Outbreak of Chlamydophila psittaci in an Outdoor Colony of Magellanic Penguins (Spheniscus magellanicus). J. Avian Med. Surg. 2012, 26, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Sieburth, J.M. Respiratory flora and diseases of Antarctic birds. Avian Dis. 1958, 2, 402–408. [Google Scholar] [CrossRef]
- Sladen, W. Studies of respiratory pathogens in Antarctica. Polar Rec. 1962, 11, 318. [Google Scholar]
- Moore, B.; Cameron, A. Chlamydia antibodies in Antarctic fauna. Avian Dis. 1969, 13, 681–684. [Google Scholar] [CrossRef]
- Karesh, W.B.; Uhart, M.M.; Frere, E.; Gandini, P.; Braselton, W.E.; Puche, H.; Cook, R.A. Health evaluation of free-ranging rockhopper penguins (Eudyptes chrysocomes) in Argentina. J. Zoo Wildl. Med. 1999, 30, 25–31. [Google Scholar]
- Travis, E.K.; Vargas, F.H.; Merkel, J.; Gottdenker, N.; Miller, R.E.; Parker, P.G. Hematology, Serum Chemistry, and Serology of Galápagos Penguins (Spheniscus mendiculus) in the Galápagos Islands, Ecuador. J. Wildl. Dis. 2006, 42, 625–632. [Google Scholar] [CrossRef]
- Smith, K.M.; Karesh, W.; Majluf, P.; Paredes, R.; Zavalaga, C.; Reul, A.H.; Stetter, M.; Braselton, W.E.; Puche, H.; Cook, R.A. Health Evaluation of Free-Ranging Humboldt Penguins (Spheniscus humboldti) in Peru. Avian Dis. 2008, 52, 130–135. [Google Scholar] [CrossRef]
- Uhart, M.; Vanstreels, R.E.T.; Gallo, L. Serological survey for select infectious agents in wild Magellanic penguins (Spheniscus magellanicus) in Argentina, 1994–2008. J. Wildl. Dis. 2020, 56, 66–81. [Google Scholar] [CrossRef]
- Isaksson, J.; Christerson, L.; Blomqvist, M.; Wille, M.; Alladio, L.A.; Sachse, K.; Olsen, B.; González-Acuña, D.; Herrmann, B. Chlamydiaceae-like bacterium, but no Chlamydia psittaci, in sea birds from Antarctica. Polar Biol. 2015, 38, 1931–1936. [Google Scholar] [CrossRef]
- Cameron, A. The isolation of a Psittacosis-Lymphogranuloma venereum (PL) agent from an emperor penguin (Aptenodytes forsteri) chick. Aust. J. Exp. Biol. Med. Sci. 1968, 46, 647–649. [Google Scholar] [CrossRef] [PubMed]
- González-Acuña, D.; Hernandez, J.; Moreno, L.; Herrmann, B.; Palma, R.; Latorre, A.; Medina-Vogel, G.; Kinsella, M.J.; Martin, N.; Araya, K.; et al. Health evaluation of wild gentoo penguins (Pygoscelis papua) in the Antarctic Peninsula. Polar Biol. 2013, 36, 1749–1760. [Google Scholar] [CrossRef]
- Boersma, P.D.; Frere, E.; Kane, O.J. Magellanic penguin (Spheniscus magellanicus). In Penguins: Natural History and Conservation; García-Borboroglu, P., Bo-ersma, P.D., Eds.; University of Washington Press: Seattle, DC, USA, 2013; pp. 285–302. [Google Scholar]
- Gallo, L.; Vanstreels, R.E.T.; Cook, R.A.; Karesh, W.B.; Uhart, M. Hematology, plasma biochemistry, and trace element reference values for free-ranging adult Magellanic Penguins (Spheniscus magellanicus). Polar Biol. 2019, 42, 733–742. [Google Scholar] [CrossRef]
- Warford, A.L.; Rekrut, K.A.; Levy, R.A.; Drill, A.E. Sucrose Phosphate Glutamate for Combined Transport of Chlamydial and Viral Specimens. Am. J. Clin. Pathol. 1984, 81, 762–764. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schulte-Hostedde, A.I.; Zinner, B.; Millar, J.S.; Hickling, G.J. Restitution of Mass–Size Residuals: Validating Body Condition Indices. Ecology 2005, 86, 155–163. [Google Scholar] [CrossRef]
- Sachse, K.; Laroucau, K.; Riege, K.; Wehner, S.; Dilcher, M.; Creasy, H.H.; Weidmann, M.; Myers, G.; Vorimore, F.; Vicari, N.; et al. Evidence for the existence of two new members of the family Chlamydiaceae and proposal of Chlamydia avium sp. nov. and Chlamydia gallinacea sp. nov. Syst. Appl. Microbiol. 2014, 37, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Vretou, E.; Livingstone, M.; Borel, N.; Pospischil, A.; Longbottom, D. Recent developments in the laboratory diagnosis of chlamydial infections. Vet. Microbiol. 2009, 135, 2–21. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 2–3. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Christerson, L.; Blomqvist, M.; Grannas, K.; Thollesson, M.; Laroucau, K.; Waldenström, J.; Eliasson, I.; Olsen, B.; Herrmann, B. A novel Chlamydiaceae-like bacterium found in faecal specimens from sea birds from the Bering Sea. Environ. Microbiol. Rep. 2010, 2, 605–610. [Google Scholar] [CrossRef]
- Aaziz, R.; Gourlay, P.; Vorimore, F.; Sachse, K.; Siarkou, V.I.; Laroucau, K. Chlamydiaceae in North Atlantic Seabirds Admitted to a Wildlife Rescue Center in Western France. Appl. Environ. Microbiol. 2015, 81, 4581–4590. [Google Scholar] [CrossRef] [PubMed]
- Everett, K.D.E.; Bush, R.M.; Andersen, A.A. Emended description of the order Chlamydiales, proposal of Parachlamydiaceae fam. nov. and Simkaniaceae fam. nov., each containing one monotypic genus, revised taxonomy of the family Chlamydiaceae, including a new genus and five new species, and standards for the identification of organisms. Int. J. Syst. Evol. Microbiol. 1999, 49, 415–440. [Google Scholar] [CrossRef]
- Griffiths, E.; Ventresca, M.S.; Gupta, R.S. BLAST screening of chlamydial genomes to identify signature proteins that are unique for the Chlamydiales, Chlamydiaceae, Chlamydophila and Chlamydia groups of species. BMC Genom. 2006, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Bavoil, P.M.; Kaltenboeck, B.; Stephens, R.S.; Kuo, C.-C.; Rosselló-Móra, R.; Horn, M. Emendation of the family Chlamydiaceae: Proposal of a single genus, Chlamydia, to include all currently recognized species. Syst. Appl. Microbiol. 2015, 38, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Kern, D.G.; Neill, M.A.; Schachter, J. A Seroepidemiologic Study of Chlamydia pneumoniae in Rhode Island. Chest 1993, 104, 208–213. [Google Scholar] [CrossRef]
- Casson, N.; Entenza, J.M.; Greub, G. Serological Cross-Reactivity between Different Chlamydia-Like Organisms. J. Clin. Microbiol. 2007, 45, 234–236. [Google Scholar] [CrossRef]
- Andersen, A.; Franson, J. Avian chlamydiosis. In Infectious Diseases of Wild Birds; Thomas, N., Hunter, D., Atkinson, C.T., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 303–316. [Google Scholar]
- Knittler, M.R.; Berndt, A.; Böcker, S.; Dutow, P.; Hänel, F.; Heuer, D.; Kägebein, D.; Klos, A.; Koch, S.; Liebler-Tenorio, E.; et al. Chlamydia psittaci: New insights into genomic diversity, clinical pathology, host–pathogen interaction and anti-bacterial immunity. Int. J. Med. Microbiol. 2014, 304, 877–893. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallo Vaulet, L.; Vanstreels, R.E.T.; Gallo, L.; Entrocassi, A.C.; Peker, L.; Blanco, G.S.; Rago, M.V.; Rodriguez Fermepin, M.; Uhart, M.M. Chlamydiaceae-Like Bacterium in Wild Magellanic Penguins (Spheniscus magellanicus). Diversity 2022, 14, 746. https://doi.org/10.3390/d14090746
Gallo Vaulet L, Vanstreels RET, Gallo L, Entrocassi AC, Peker L, Blanco GS, Rago MV, Rodriguez Fermepin M, Uhart MM. Chlamydiaceae-Like Bacterium in Wild Magellanic Penguins (Spheniscus magellanicus). Diversity. 2022; 14(9):746. https://doi.org/10.3390/d14090746
Chicago/Turabian StyleGallo Vaulet, Lucía, Ralph Eric Thijl Vanstreels, Luciana Gallo, Andrea Carolina Entrocassi, Laura Peker, Gabriela S. Blanco, Maria Virginia Rago, Marcelo Rodriguez Fermepin, and Marcela M. Uhart. 2022. "Chlamydiaceae-Like Bacterium in Wild Magellanic Penguins (Spheniscus magellanicus)" Diversity 14, no. 9: 746. https://doi.org/10.3390/d14090746
APA StyleGallo Vaulet, L., Vanstreels, R. E. T., Gallo, L., Entrocassi, A. C., Peker, L., Blanco, G. S., Rago, M. V., Rodriguez Fermepin, M., & Uhart, M. M. (2022). Chlamydiaceae-Like Bacterium in Wild Magellanic Penguins (Spheniscus magellanicus). Diversity, 14(9), 746. https://doi.org/10.3390/d14090746