

The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians


Abstract
:1. Introduction
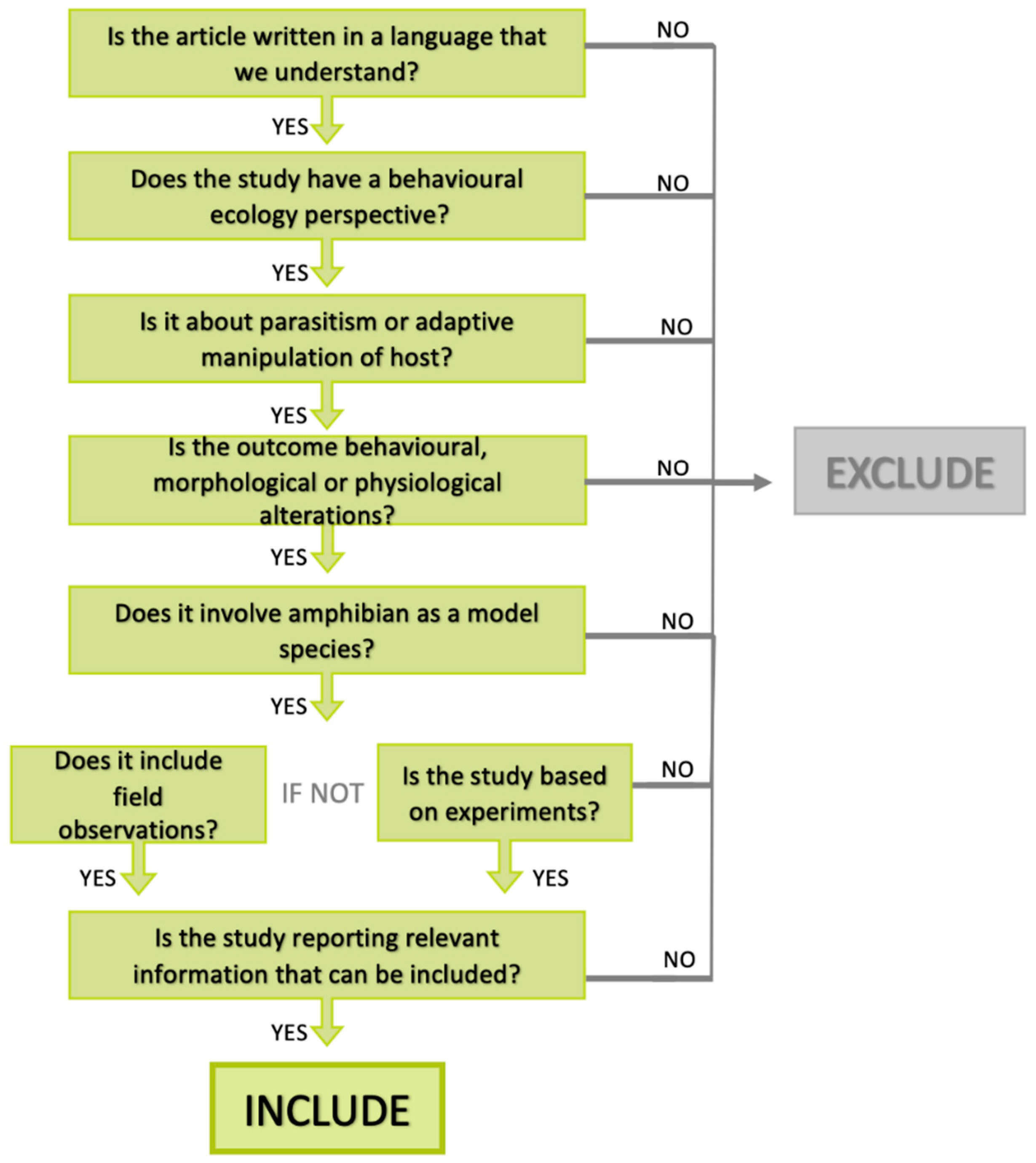
2. Material and Methods
Study Selection Criteria
3. Results
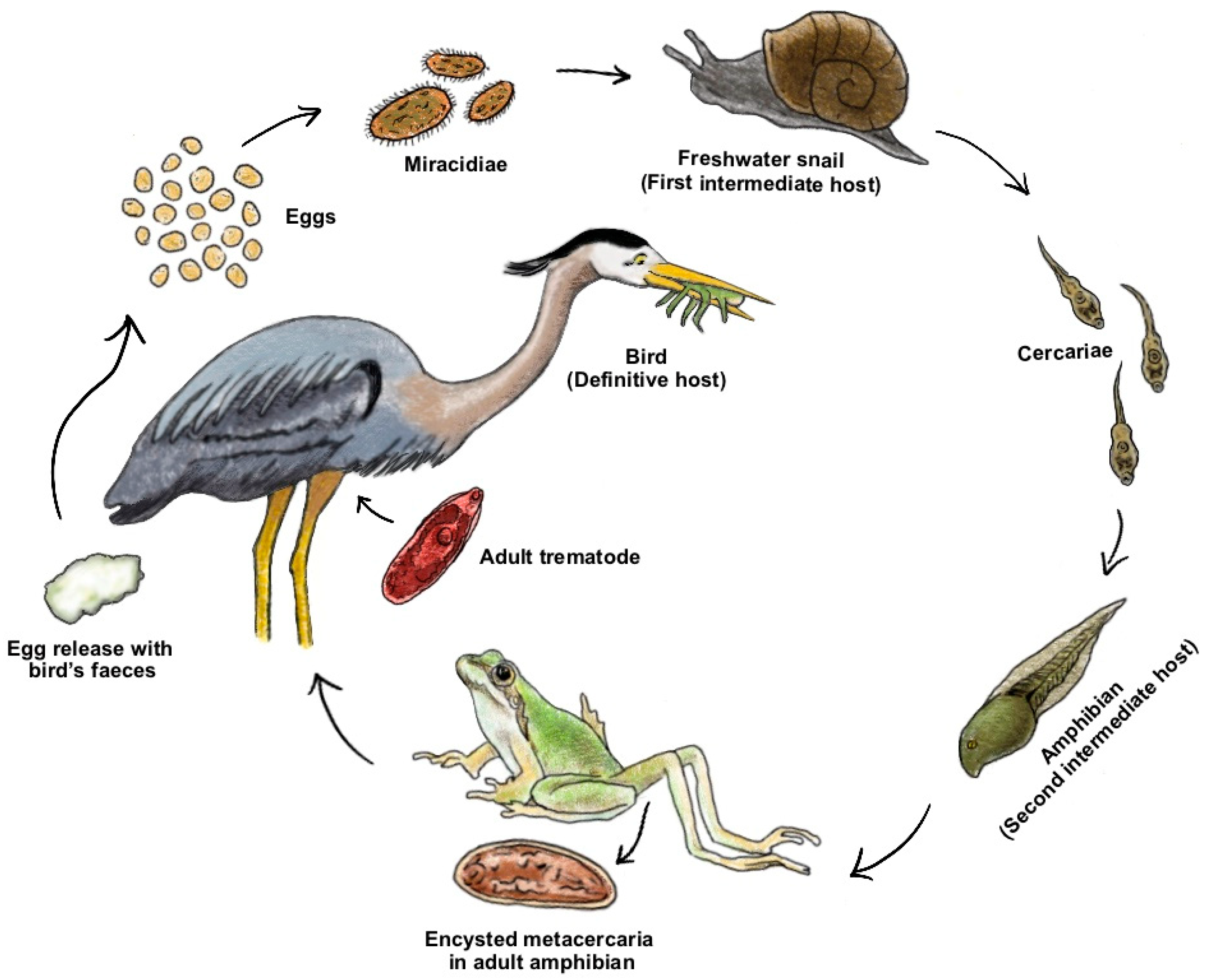
3.1. Behavioural Manipulation

{kind=link}
{kind=link}
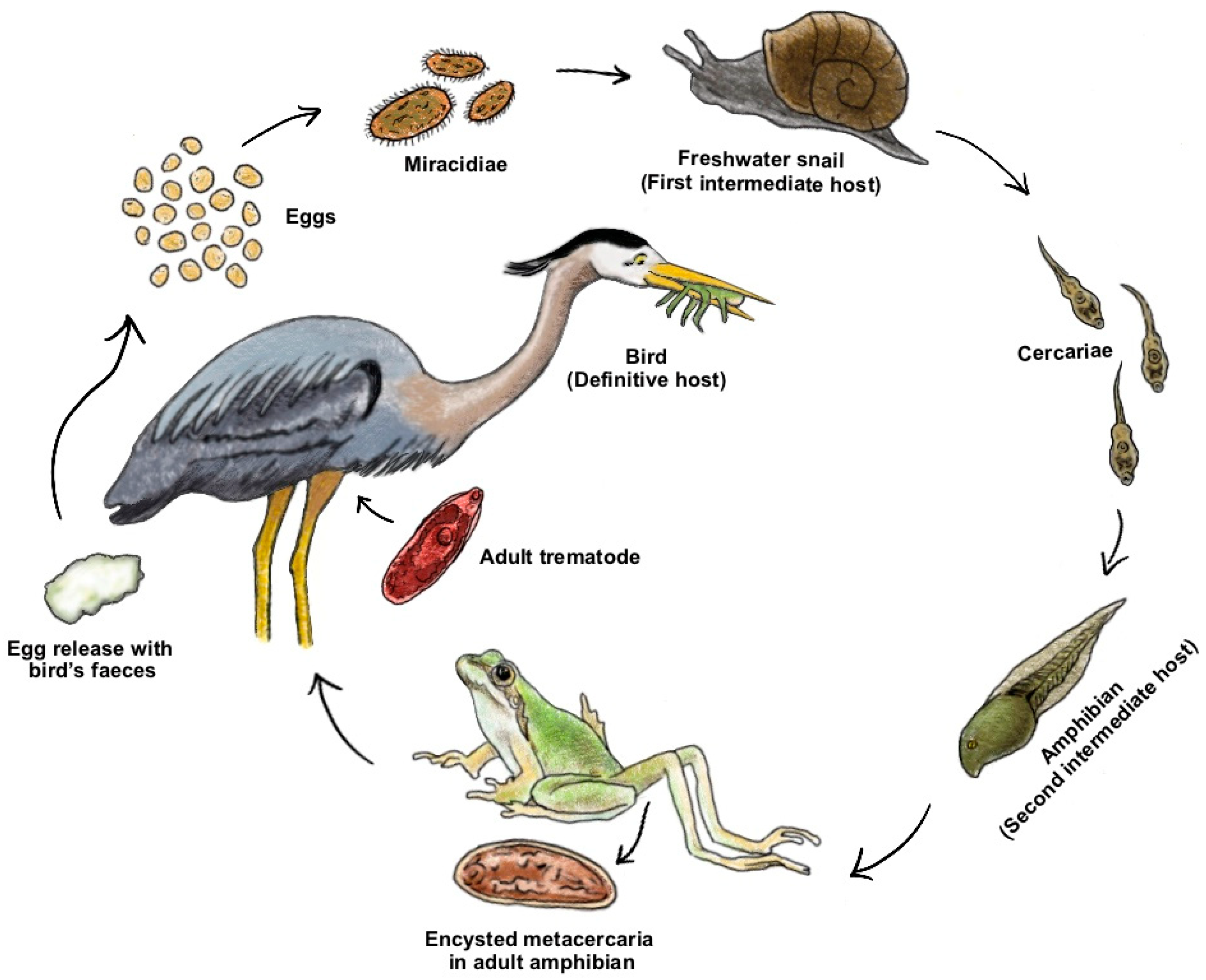{kind=link}
| Parasite Species (Taxonomic Class) | Host Species | Parasite Life Cycle | Outcome of Host–Parasite Interaction Increasing Parasite Transmission | Effect on Infected Host | References |
|---|---|---|---|---|---|
| Ribeiroia ondatrae (Trematoda) | Pseudacris regilla | Indirect | Enhanced predation risk of infected host | Loss of fear to predator Microhabitat altered choice Altered thermoregulatory behaviour | [51] |
| Codonocephalus urnigerus (Trematoda) | Pelophylax ridibundus | Indirect | Enhanced predation risk of infected host | Altered sexual, territorial, and foraging behaviour Failed anti-predator behaviour | [55] |
| Rhabdias pseudosphaerocephala (Secernentea) | Rhinella marina | Direct | Toads with higher temperatures showed higher lungworm parasitemias and increased lungworm larval production Increased parasite larvae survival in moist environments | Selection for warmer environments Preference for moist habitat for defecation | [53] |
| Hannemania eltoni (Arachnida) | Plethodon angusticlavius | Indirect | Increased contact between host and parasite | Increased agonistic behaviour in males Failed anti-parasitic behaviour in females | [58,60] |
| Hannemania dunni (Arachnida) | Plethodon ouachitae | Indirect | Increased contact between host and parasite | Reduced ability to detect prey and pheromones from conspecifics | [57] |
| Batrachochytrium dendrobatidis (Chytridiomycetes) | Hyla japonica Anaxyrus boreas Taricha glutinosa Ambystoma macrodactylyum | Direct | Increased contact between infected host during mating Enhanced predation risk of infected host | Enhanced calling behaviour in males Erratic swimming behaviour Failed detection of predator cues | [63,64] |
| Candida humicola (Saccharomycetes) | Rana aurora | Direct | Increased parasite transmission through coprophagy Enhanced predation risk of infected host | Preference for environments with high concentration of conspecifics and higher accumulation of faeces Failed detection of predator chemical cues | [65] |
| Aeromonas hydrophila (Gammaproteobacteria) | Rana catesbaiana | Direct | Enhanced predation risk of infected host | Reduced refuge-seeking behaviour in presence of predator | [68] |
3.2. Morphological Manipulation
| Parasite Species | Host Species | Parasite Life Cycle | Outcome of Host–Parasite Interaction Increasing Parasite Transmission | Effect on Infected Host | References |
|---|---|---|---|---|---|
| Ribeiroia ondatrae (Trematoda) | Taricha torosa Tarica granulosa Ambystoma macrodactylum Bufo boreas Pseudacris regilla Rana aurora Rana luteiventris Rana catesbaiana Rana cascadae | Indirect | Increased predation risk of infected host by impaired locomotor activity | Missing limbs and digits, extra limbs, extra appendices, skin webbings fusion | [51,69,70,71,72,77,78] |
| Clinostomumspp. (Trematoda) | Ambystoma tigrinum | Indirect | Increased predation risk of infected host by impaired locomotor activity | Scoliosis | [79] |
| Acanthostomum burminis (Trematoda) | Polypedates cruciger Duttaphrynus melanostictus | Indirect | Increased predation risk of infected host by impaired locomotor activity | Extension of the spine Scoliosis Kyphosis | [80,82] |
| Strigea robusta (Trematoda) | Pelophylax ridibundus Bufo bufo Bufotes viridis Bufotes baturae | Indirect | Increased predation risk of infected host by impaired locomotor activity | Mild and severe limb malformations | [74,76] |
3.3. Physiological Manipulation
| Parasite Species | Host Species | Parasite Life Cycle | Outcome of Host–Parasite Interaction Increasing Parasite Transmission | Effect on Infected Host | References |
|---|---|---|---|---|---|
| Rhabdias pseudosphaerocephala (Secernentea) | Rhinella marina | Direct | Higher fecundity and larval survival of lungworms in moister feces | Defecated feaces with increased water content | [53] |
| Placobdellaspp. (Clitellata) | Cryptobranchus alleganiensis | Direct | Avoid parasite detection by host; hence, parasites can feed for a longer time | Reduced corticosterone response of hosts to stressful situations | [87] |
3.4. Factors Influencing Amphibian–Parasite Interactions
3.4.1. Intrinsic Amphibian Traits That Influence Infection
3.4.2. Extrinsic Factors That Influence Infection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How Many Parasites? How Many Hosts? Proc. Natl. Acad. Sci. USA 2008, 105, 11482–11489. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Dallas, T.A.; Alexander, L.W.; Phelan, A.L.; Phillips, A.J. What Would It Take to Describe the Global Diversity of Parasites? Proc. R. Soc. B Biol. Sci. 2020, 287, 20201841. [Google Scholar] [CrossRef] [PubMed]
- Kelehear, C.; Saltonstall, K.; Torchin, M.E. Negative Effects of Parasitic Lung Nematodes on the Fitness of a Neotropical Toad (Rhinella horribilis). Parasitology 2019, 146, 928–936. [Google Scholar] [CrossRef] [PubMed]
- Stegen, G.; Pasmans, F.; Schmidt, B.R.; Rouffaer, L.O.; van Praet, S.; Schaub, M.; Canessa, S.; Laudelout, A.; Kinet, T.; Adriaensen, C.; et al. Drivers of Salamander Extirpation Mediated by Batrachochytrium Salamandrivorans. Nature 2017, 544, 353–356. [Google Scholar] [CrossRef]
- Van Valen, L. A new Evolutionary Law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Poulin, R. Parasite Manipulation of Host Behavior: An Update and Frequently Asked Questions. Adv. Study Behav. 2010, 41, 151–186. [Google Scholar]
- Cozzarolo, C.S.; Glaizot, O.; Christe, P.; Pigeault, R. Enhanced Attraction of Arthropod Vectors to Infected Vertebrates: A Review of Empirical Evidence. Front. Ecol. Evol. 2020, 8, 568140. [Google Scholar] [CrossRef]
- Busula, A.O.; Verhulst, N.O.; Bousema, T.; Takken, W.; Boer, J.G. Mechanisms of Plasmodium-Enhanced Attraction of Mosquito Vectors. Trends Parasitol. 2017, 33, 961–973. [Google Scholar] [CrossRef]
- Moore, J. Parasites and the Behavior of Animals; Oxford University Press, Inc.: New York, NY, USA, 2002. [Google Scholar]
- Dass, S.A.H.; Vasudevan, A.; Dutta, D.; Soh, L.J.T.; Sapolsky, R.M.; Vyas, A. Protozoan Parasite Toxoplasma Gondii Manipulates Mate Choice in Rats by Enhancing Attractiveness of Males. PLoS ONE 2011, 6, e27229. [Google Scholar] [CrossRef]
- Poirotte, C.; Kappeler, P.M.; Ngoubangoye, B.; Bourgeois, S.; Moussodji, M.; Charpentier, M.J.E. Morbid Attraction to Leopard Urine in Toxoplasma-Infected Chimpanzees. Curr. Biol. 2016, 26, 98–99. [Google Scholar] [CrossRef]
- Poulin, R.; Maure, F. Host Manipulation by Parasites: A Look Back Before Moving Forward. Trends Parasitol. 2015, 31, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Doherty, J.-F. When Fiction Becomes Fact: Exaggerating Host Manipulation by Parasites. Proc. R. Soc. B Biol. Sci. 2020, 287, 20201081. [Google Scholar] [CrossRef] [PubMed]
- Minchella, D.J. Host Life-History Variation in Response to Parasitism. Parasitology 1985, 90, 205–216. [Google Scholar] [CrossRef]
- Poulin, R. “Adaptive” Changes in the Behaviour of Parasitized Animals: A Critical Review. Int. J. Parasitol. 1995, 25, 1371–1383. [Google Scholar] [CrossRef]
- Berdoy, M.; Webster, J.P.; Mcdonald, D.W. Fatal Attraction in Rats Infected with Toxoplasma Gondii. Proc. R. Soc. B Biol. Sci. 2000, 267, 1591–1594. [Google Scholar] [CrossRef]
- Hari Dass, S.A.; Vyas, A. Toxoplasma Gondii Infection Reduces Predator Aversion in Rats through Epigenetic Modulation in the Host Medial Amygdala. Mol. Ecol. 2014, 23, 6114–6122. [Google Scholar] [CrossRef]
- Lafferty, K.D. The Evolution of Trophic Transmission. Parasitol. Today 1999, 15, 111–115. [Google Scholar] [CrossRef]
- Poulin, R.; Thomas, F. Phenotypic Variability Induced by Parasites: Extent and Evolutionary Implications. Parasitol. Today 1999, 15, 28–32. [Google Scholar] [CrossRef]
- Yanoviak, S.P.; Kaspari, M.; Dudley, R.; Poinar, G. Parasite-Induced Fruit Mimicry in a Tropical Canopy Ant. Am. Nat. 2008, 171, 536–544. [Google Scholar] [CrossRef]
- Thomas, F.; Adamo, S.; Moore, J. Parasitic Manipulation: Where Are We and Where Should We Go? Behav. Process. 2005, 68, 185–199. [Google Scholar] [CrossRef]
- Biron, D.G.; Marché, L.; Ponton, F.; Loxdale, H.D.; Galéotti, N.; Renault, L.; Joly, C.; Thomas, F. Behavioural Manipulation in a Grasshopper Harbouring Hairworm: A Proteomics Approach. Proc. R. Soc. B Biol. Sci. 2005, 272, 2117–2126. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Poulin, R.; Brodeur, J. Host Manipulation by Parasites: A Multidimensional Phenomenon. Oikos 2010, 119, 1217–1223. [Google Scholar] [CrossRef]
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.1 (20 June 2022). Electronic Database. American Museum of Natural History, New York, USA, 2021. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 12 August 2022).
- Duellman, W.E.; Trueb, L. Biology of Amphibians; The John Hopkins University Press: Baltimore, MD, USA, 1986. [Google Scholar]
- Dunson, W.A.; Wyman, R.L.; Corbett, E.S. A Symposium on Amphibian Declines and Habitat Acidification. J. Herpetol. 1992, 26, 349–352. [Google Scholar] [CrossRef]
- Welsh, H.H., Jr.; Ollivier, L.M. Stream amphibians as indicators of ecosystem stress: A case study from California’s redwoods. Ecol. Appl. 1998, 8, 1118–1132. [Google Scholar] [CrossRef]
- Venturino, A.; Rosenbaum, E.; Caballero De Castro, A.; Anguiano, O.L.; Gauna, L.; Fonovich de Schroeder, T.; Pechen de D’Angelo, A.M. Biomarkers of Effect in Toads and Frogs. Biomarkers 2003, 8, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Peluso, J.; Aronzon, C.M.; Acquaroni, M.; Pérez Coll, C.S. Biomarkers of Genotoxicity and Health Status of Rhinella Fernandezae Populations from the Lower Paraná River Basin, Argentina. Ecol. Indic. 2020, 117, 106588. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. 2021. Available online: https://www.iucnredlist.org (accessed on 28 June 2022).
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological Annihilation via the Ongoing Sixth Mass Extinction Signaled by Vertebrate Population Losses and Declines. Proc. Natl. Acad. Sci. USA 2017, 114, 6089–6096. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and Trends of Amphibian Declines and Extinctions Worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.J.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello De Alba, A.; Bosch, J. Collapse of Amphibian Communities Due to an Introduced Ranavirus. Curr. Biol. 2014, 24, 2586–2591. [Google Scholar] [CrossRef]
- Berger, L.; Roberts, A.A.; Voyles, J.; Longcore, J.E.; Murray, K.A.; Skerratt, L.F. History and Recent Progress on Chytridiomycosis in Amphibians. Fungal Ecol. 2016, 19, 89–99. [Google Scholar] [CrossRef]
- Frainer, A.; McKie, B.G.; Amundsen, P.A.; Knudsen, R.; Lafferty, K.D. Parasitism and the Biodiversity-Functioning Relationship. Trends Ecol. Evol. 2018, 33, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Koprivnikar, J.; Marcogliese, D.J.; Rohr, J.R.; Orlofske, S.A.; Raffel, T.R.; Johnson, P.T.J. Macroparasite Infections of Amphibians: What Can They Tell Us? Ecohealth 2012, 9, 342–360. [Google Scholar] [CrossRef] [PubMed]
- Denver, R.J. Neuroendocrinology of Amphibian Metamorphosis. In Current Topics in Developmental Biology; Academic Press Inc.: Cambridge, MA, USA, 2013; Volume 103, pp. 195–227. [Google Scholar]
- Kulkarni, S.S.; Denver, R.J.; Gomez-Mestre, I.; Buchholz, D.R. Genetic Accommodation via Modified Endocrine Signalling Explains Phenotypic Divergence among Spadefoot Toad Species. Nat. Commun. 2017, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Reiehenbach-Klinke, H.; Elkan, E. The Principal Diseases of Lower Vertebrates; Oxford University Press: Oxford, UK, 1965. [Google Scholar]
- Barta, J.R.; Desser, S.S. Blood Parasites of Amphibians from Algonquin Park, Ontario. J. Wildl. Dis. 1984, 20, 180–189. [Google Scholar] [CrossRef]
- Boulianne, B.; Evans, R.C.; Smith, T.G. Phylogenetic Analysis of Hepatozoon Species (Apicomplexa: Adeleorina) Infecting Frogs of Nova Scotia, Canada, Determined by ITS-1 Sequences. J. Parasitol. 2007, 93, 1435–1441. [Google Scholar] [CrossRef]
- Stanford, M. Avian and Exotic Animal Hematology and Cytology. J. Avian Med. Surg. 2007, 21, 335. [Google Scholar] [CrossRef]
- Readel, A.M.; Goldberg, T.L. Blood Parasites of Frogs from an Equatorial African Montane Forest in Western Uganda. J. Parasitol. 2010, 96, 448–450. [Google Scholar] [CrossRef]
- Muriel, J.; González-Blázquez, M.; Matta Cahacho, N.E.; Vargas-León, C.M.; Marzal, A. Parasitas Sanguíneos de Anfíbios; Editora da Universidade Federal do Piauí: Teresina, Brazil, 2021. [Google Scholar]
- Bardsley, J.E.; Harmsen, R. The Trypanosomes of Anura. Adv. Parasitol. 1973, 11, 1–73. [Google Scholar]
- Arneberg, P. Host Population Density and Body Mass as Determinants of Species Richness in Parasite Communities: Comparative Analyses of Directly Transmitted Nematodes of Mammals. Ecography 2002, 25, 88–94. [Google Scholar] [CrossRef]
- Garcia-Longoria, L.; Marzal, A.; de Lope, F.; Garamszegi, L. Host-Parasite Interaction Explains Variation in the Prevalence of Avian Haemosporidians at the Community Level. PLoS ONE 2019, 14, e0205624. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. J. Chin. Integr. Med. 2009, 7, 889–896. [Google Scholar] [CrossRef]
- O’Dea, R.E.; Lagisz, M.; Jennions, M.D.; Koricheva, J.; Noble, D.W.A.; Parker, T.H.; Gurevitch, J.; Page, M.J.; Stewart, G.; Moher, D.; et al. Preferred Reporting Items for Systematic Reviews and Meta-Analyses in Ecology and Evolutionary Biology: A PRISMA Extension. Biol. Rev. 2021, 96, 1695–1722. [Google Scholar] [CrossRef] [PubMed]
- Foo, Y.Z.; O’Dea, R.E.; Koricheva, J.; Nakagawa, S.; Lagisz, M. A Practical Guide to Question Formation, Systematic Searching and Study Screening for Literature Reviews in Ecology and Evolution. Methods Ecol. Evol. 2021, 12, 1705–1720. [Google Scholar] [CrossRef]
- Goodman, B.A.; Johnson, P.T.J. Disease and the Extended Phenotype: Parasites Control Host Performance and Survival through Induced Changes in Body Plan. PLoS ONE 2011, 6, e20193. [Google Scholar] [CrossRef]
- Reading, C. The Effect of Winter Temperatures on the Timing of breeding Activity in the Common Toad, Bufo Bufo. Oecologia 2007, 117, 469–475. [Google Scholar] [CrossRef]
- Patrick, B. Finnerty; Richard Shine; Gregory, P. Brown Survival of the Feces: Does a Nematode Lungworm Adaptively Manipulate the Behavior of Its Cane Toad Host? Ecol. Evol. 2018, 8, 4606–4618. [Google Scholar]
- Chikhlyaev, I.V.; Ruchin, A.B. An Overview of the Helminths of Moor Frog Rana Arvalis Nilsson, 1842 (Amphibia: Anura) in the Volga Basin. Diversity 2021, 13, 61. [Google Scholar] [CrossRef]
- Ivanov, V.M.; Kalmykov, A.P.; Semyonova, N.N.; Fedorovich, V.V.; Parshina, O.Y. Lake Frog Behavior and Viability Changes under the Influence of Helminthic Invasion. Curr. Stud. Herpet 2012, 12, 49–55. [Google Scholar]
- Wells, K.D.; Schwartz, J.J. The Behavioral Ecology of Anuran Communication. In Hearing and Sound Communication in Amphibians; Springer: New York, NY, USA, 2007; pp. 44–86. [Google Scholar]
- Anthony, C.D.; Mendelson, J.R., III; Simons, R.R. Differential Parasitism by Sex on Plethodontid Salamanders and Histological Evidence for Structural Damage to the Nasolabial Groove. Am. Midl. Nat. 1994, 132, 302–307. [Google Scholar] [CrossRef]
- Maksimowich, D.S.; Mathis, A. Parasitized Salamanders Are Inferior Competitors for Territories and Food Resources. Ethology 2000, 106, 319–329. [Google Scholar] [CrossRef]
- Hillgarth, N. Ectoparasite transfer during mating in ring-necked pheasants Phasianus colchicus. J. Avian Biol. 1996, 27, 260–262. [Google Scholar] [CrossRef]
- Maksimowich, D.S.; Mathis, A. Pheromonal Markers as Indicators of Parasite Load: Parasite-Mediated Behavior in Salamanders (Plethodon angusticlavius). Acta Ethologica 2001, 3, 83–87. [Google Scholar] [CrossRef]
- Grafe, T.U. Anuran Choruses as Communication; Animal Communication Networks; University of Wurzburg: Wurzburg, Germany, 2005; p. 277. [Google Scholar]
- Zahavi, A. Mate selection—A selection for a handicap. J. Theor. Biol. 1975, 53, 205–214. [Google Scholar] [CrossRef]
- Deuknam, A.; Waldman, B. Enhanced Call Effort in Japanese Tree Frogs Infected by Amphibian Chytrid Fungus. Biol. Lett. 2016, 12, 20160018. [Google Scholar]
- Han, B.A.; Searle, C.L.; Blaustein, A.R. Effects of an Infectious Fungus, Batrachochytrium dendrobatidis, on Amphibian Predator-Prey Interactions. PLoS ONE 2011, 6, e16675. [Google Scholar] [CrossRef]
- Lefcort, H.; Blaustein, A.R. Disease, predator avoidance, and vulnerability to predation in tadpoles. Oikos 1995, 74, 469–474. [Google Scholar] [CrossRef]
- Steinwascher, K. Host-Parasite Interaction as a Potential Population-Regulating Mechanism. Ecology 1979, 60, 884–890. [Google Scholar] [CrossRef]
- Densmore, C.L.; Green, D.L. Diseases of Amphibians. ILAR J. 2007, 48, 235–254. [Google Scholar] [CrossRef]
- Lefcort, H.; Eiger, S.M. Antipredatory behaviour of feverish tadpoles: Implications for pathogen transmission. Behaviour 1993, 126, 13–27. [Google Scholar] [CrossRef]
- Sessions, S.K.; Franssen, R.A.; Horner, V.L. Morphological clues from multilegged frogs: Are retinoids to blame? Science 1999, 284, 800–802. [Google Scholar] [CrossRef]
- Stopper, G.F.; Franssen, R.A.; Sessions, S.K. How Trematodes Cause Limb Deformities in Amphibians. J. Exp. Zool. 2002, 294, 252–263. [Google Scholar] [CrossRef]
- Johnson, P.T.; Lunde, K.B.; Thurman, E.M.; Ritchie, E.G.; Wray, S.N.; Sutherland, D.R.; Blaustein, A.R. Parasite (Ribeiroia ondatrae) Infection Linked to Amphibian Malformations in the Western United States. Ecol. Monogr. 2002, 72, 151–168. [Google Scholar] [CrossRef]
- Johnson, P.T.; Preu, E.R.; Sutherland, D.R.; Romansic, J.M.; Han, B.; Blaustein, A.R. Adding infection to injury: Synergistic effects of predation and parasitism on amphibian malformations. Ecology 2006, 87, 2227–2235. [Google Scholar] [CrossRef]
- Johnson, R.L.; Tabin, C.J. Molecular Models for Vertebrate Limb Development. Cell 1997, 90, 979–990. [Google Scholar] [CrossRef]
- Svinin, A.O.; Matushkina, K.A.; Dedukh, D.V.; Bashinskiy, I.V.; Ermakov, O.A.; Litvinchuk, S.N. Strigea robusta (Digenea: Strigeidae) Infection Effects on the Gonadal Structure and Limb Malformation in Toad Early Development. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2022, 337, 675–686. [Google Scholar] [CrossRef]
- Heneberg, P.; Sitko, J.; Těšínský, M.; Rząd, I.; Bizos, J. Central European Strigeidae Railliet, 1919 (Trematoda: Strigeidida): Molecular and Comparative Morphological Analysis Suggests the Reclassification of Parastrigea robusta Szidat, 1928 into Strigea Abildgaard, 1790. Parasitol. Int. 2018, 67, 688–701. [Google Scholar] [CrossRef]
- Svinin, A.O.; Bashinskiy, I.V.; Litvinchuk, S.N.; Ermakov, O.A.; Ivanov, A.Y.; Neymark, L.A.; Vedernikov, A.A.; Osipov, V.V.; Drobot, G.P.; Dubois, A. Strigea robusta causes polydactyly and severe forms of Rostand’s anomaly P in water frogs. Parasites Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Sessions, S.K.; Ruth, S.B. Explanation for naturally occurring supernumerary limbs in amphibians. J. Exp. Zool. 1990, 254, 38–47. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Lunde, K.B.; Ritchie, E.G.; Launer, A.E. The Effect of Trematode Infection on Amphibian Limb Development and Survivorship. Science 1999, 284, 802–804. [Google Scholar] [CrossRef]
- McAllister, C.T.; Bursey, C.R.; Niemiller, M.L.; Miller, B.T. A Noteworthy Infection of Clinostomum Complanatum (Digenea: Clinostomidae) in a Cave Salamander, Eurycea Lucifuga (Caudata: Plethodontidae), from Northcentral Tennessee. Tex. J. Sci. 2007, 59, 321–327. [Google Scholar]
- Perpiñán, D.; Garner, M.M.; Trupkiewicz, J.G.; Malarchik, J.; Armstrong, D.L.; Lucio-Forster, A.; Bowman, D.D. Scoliosis in a tiger salamander (Ambystoma tigrinum) associated with encysted digenetic trematodes of the genus Clinostomum. J. Wildl. Dis. 2010, 46, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Rajakaruna, R.S.; Piyatissa, P.M.J.R.; Jayawardena, U.A.; Navaratne, A.N.; Amerasinghe, P.H. Trematode Infection Induced Malformations in the Common Hourglass Treefrogs. J. Zool. 2008, 275, 89–95. [Google Scholar] [CrossRef]
- Pathirana, N.U.K.; Rajakaruna, R.S. Trematode Infections in Frogs: Do All Cercarial Morphotypes Infect and Induce Effects on the Common Hourglass Tree Frog, Polypedates Cruciger? Ceylon J. Sci. 2018, 47, 319–330. [Google Scholar] [CrossRef]
- Jayawardena, U.A.; Tkach, V.V.; Navaratne, A.N.; Amerasinghe, P.H.; Rajakaruna, R.S. Malformations and Mortality in the Asian Common Toad Induced by Exposure to Pleurolophocercous Cercariae (Trematoda: Cryptogonimidae). Parasitol. Int. 2013, 62, 246–252. [Google Scholar] [CrossRef]
- Kupferberg, S.J.; Catenazzi, A.; Lunde, K.; Lind, A.J.; Palen, W.J. Parasitic Copepod (Lernaea cyprinacea) Outbreaks in Foothill Yellow-Legged Frogs (Rana boylii) Linked to Unusually Warm Summers and Amphibian Malformations in Northern California. Copeia 2009, 2009, 529–537. [Google Scholar] [CrossRef]
- Hildebrandt, J.P.; Lemke, S. Small Bite, Large Impact-Saliva and Salivary Molecules in the Medicinal Leech, Hirudo Medicinalis. Naturwissenschaften 2011, 98, 995–1008. [Google Scholar] [CrossRef]
- Salzet, M.; Chopin, V.; Baert, J.L.; Matias, I.; Malecha, J. Theromin, a Novel Leech Thrombin Inhibitor. J. Biol. Chem. 2000, 275, 30774–30780. [Google Scholar] [CrossRef] [Green Version]
- DuRant, S.E.; Hopkins, W.A.; Davis, A.K.; Romero, L.M. Evidence of Ectoparasite-Induced Endocrine Disruption in an Imperiled Giant Salamander, the Eastern Hellbender (Cryptobranchus alleganiensis). J. Exp. Biol. 2015, 218, 2297–2304. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar]
- Dumont, M.; Moné, H.; Mouahid, G.; Idris, M.A.; Shaban, M.; Boissier, J. Influence of Pattern of Exposure, Parasite Genetic Diversity and Sex on the Degree of Protection against Reinfection with Schistosoma Mansoni. Parasitol. Res. 2007, 101, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Telfer, S.; Lambin, X.; Birtles, R.; Beldomenico, P.; Burthe, S.; Paterson, S.; Begon, M. Species Interactions in a Parasite Community Drive Infection Risk in a Wildlife Population. Science 2010, 330, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Rohr, J.R.; Hoverman, J.T.; Kellermanns, E.; Bowerman, J.; Lunde, K.B. Living Fast and Dying of Infection: Host Life History Drives Interspecific Variation in Infection and Disease Risk. Ecol. Lett. 2012, 15, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Raffel, T.R.; Sessions, S.K. Digenetic trematodes and their relationship to amphibian declines and deformities. Amphib. Biol. 2009, 8, 3067–3088. [Google Scholar]
- Koprivnikar, J.; Forbes, M.R.; Baker, R.L. Larval Amphibian Growth and Development under Varying Density: Are Parasitized Individuals Poor Competitors? Oecologia 2008, 155, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Kluger, M.J. Fever: Role of pyrogens and cryogens. Physiol. Rev. 1991, 71, 93–127. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Alford, R.A.; Marantelli, G. Emerging Disease of Amphibians Cured by Elevated Body Temperature. Dis. Aquat. Organ. 2003, 55, 65–67. [Google Scholar] [CrossRef]
- Taylor, C.N.; Oseen, K.L.; Wassersug, R.J. On the Behavioural Response of Rana and Bufo Tadpoles to Echinostomatoid Cercariae: Implications to Synergistic Factors Influencing Trematode Infections in Anurans. Can. J. Zool. 2004, 82, 701–706. [Google Scholar] [CrossRef]
- Daly, E.W.; Johnson, P.T.J. Beyond Immunity: Quantifying the Effects of Host Anti-Parasite Behavior on Parasite Transmission. Oecologia 2011, 165, 1043–1050. [Google Scholar] [CrossRef]
- Koprivnikar, J.; Gibson, C.H.; Redfern, J.C. Infectious Personalities: Behavioural Syndromes and Disease Risk in Larval Amphibians. Proc. R. Soc. B Biol. Sci. 2012, 279, 1544–1550. [Google Scholar] [CrossRef] [Green Version]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral Syndromes: An Ecological and Evolutionary Overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef]
- Knutie, S.A.; Wilkinson, C.L.; Kohl, K.D.; Rohr, J.R. Early-Life Disruption of Amphibian Microbiota Decreases Later-Life Resistance to Parasites. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.; Chase, J.M.; Dosch, K.L.; Hartson, R.B.; Gross, J.A.; Larson, D.J.; Sutherland, D.R.; Carpenter, S.R. Aquatic eutrophication promotes pathogenic infection in amphibians. Proc. Natl. Acad. Sci. USA 2007, 104, 15781–15786. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.J.; Sutherland, D.R. Amphibian Deformities and Ribeiroia Infection: An Emerging Helminthiasis. Trends Parasitoly 2003, 19, 332–335. [Google Scholar] [CrossRef]
- Kiesecker, J.M. Synergism between Trematode Infection and Pesticide Exposure: A Link to Amphibian Limb Deformities in Nature? Proc. Natl. Acad. Sci. USA 2002, 99, 9900–9904. [Google Scholar] [CrossRef] [PubMed]
- Koprivnikar, J. Interactions of environmental stressors impact survival and development of parasitized larval amphibians. Ecol. Appl. 2010, 20, 2263–2272. [Google Scholar] [CrossRef]
- Buss, N.; Hua, J. Parasite Susceptibility in an Amphibian Host Is Modified by Salinization and Predators. Environ. Pollut. 2018, 236, 754–763. [Google Scholar] [CrossRef]
- Milotic, D.; Milotic, M.; Koprivnikar, J. Effects of Road Salt on Larval Amphibian Susceptibility to Parasitism through Behavior and Immunocompetence. Aquat. Toxicol. 2017, 189, 42–49. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez-Caballero, I.; Garcia-Longoria, L.; Gomez-Mestre, I.; Marzal, A. The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians. Diversity 2022, 14, 739. https://doi.org/10.3390/d14090739
Hernandez-Caballero I, Garcia-Longoria L, Gomez-Mestre I, Marzal A. The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians. Diversity. 2022; 14(9):739. https://doi.org/10.3390/d14090739
Chicago/Turabian StyleHernandez-Caballero, Irene, Luz Garcia-Longoria, Ivan Gomez-Mestre, and Alfonso Marzal. 2022. "The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians" Diversity 14, no. 9: 739. https://doi.org/10.3390/d14090739
APA StyleHernandez-Caballero, I., Garcia-Longoria, L., Gomez-Mestre, I., & Marzal, A. (2022). The Adaptive Host Manipulation Hypothesis: Parasites Modify the Behaviour, Morphology, and Physiology of Amphibians. Diversity, 14(9), 739. https://doi.org/10.3390/d14090739