
Avian Haemosporidian Diversity in South Texas: New Lineages and Variation in Prevalence between Sampling Sources and Sites



Abstract
:1. Introduction
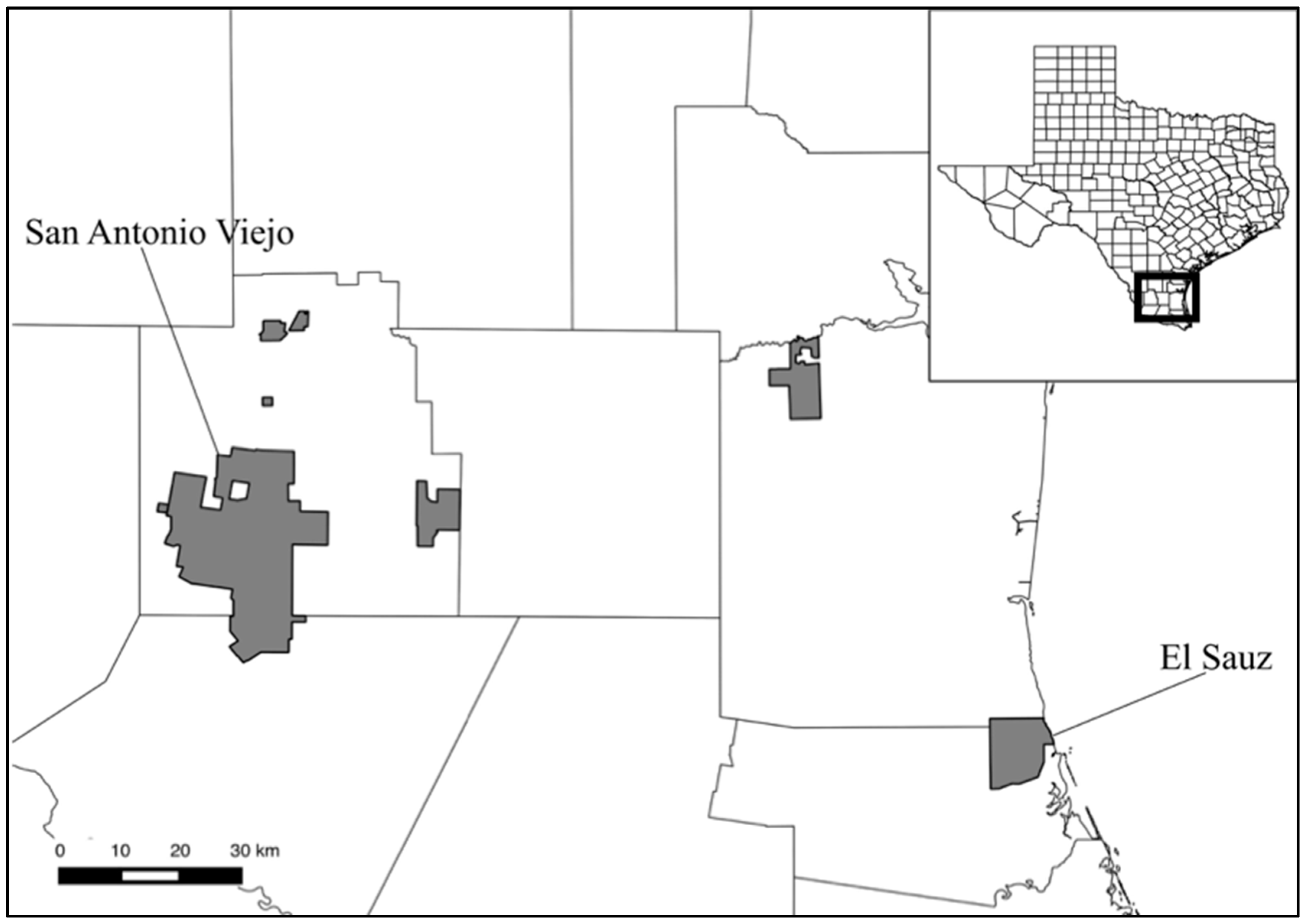
2. Materials and Methods
2.1. Prevalence Analyses
2.2. Lineage Identification and Molecular Analysis of Selected Host Families
2.3. Precipitation Data
3. Results
3.1. Prevalence
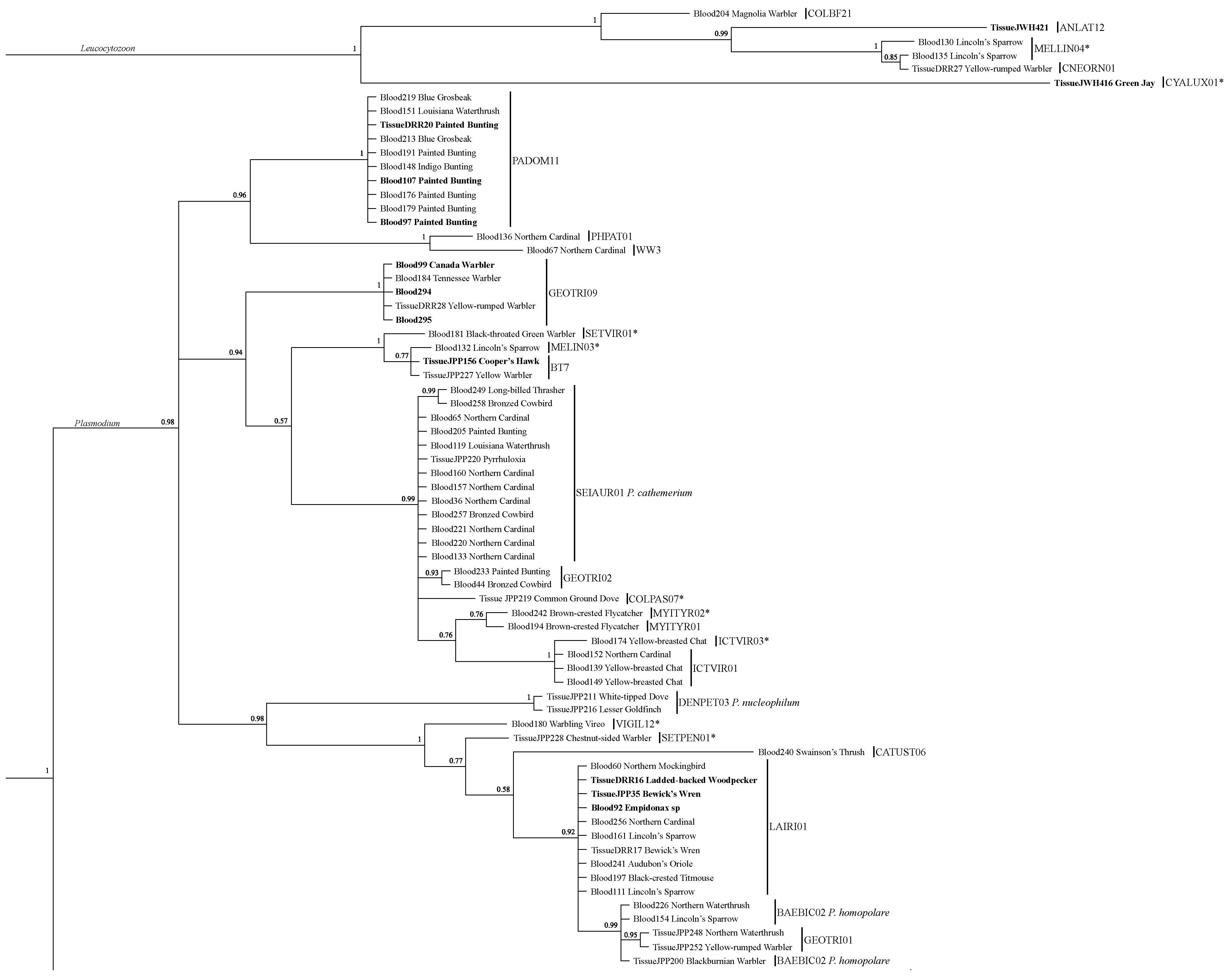
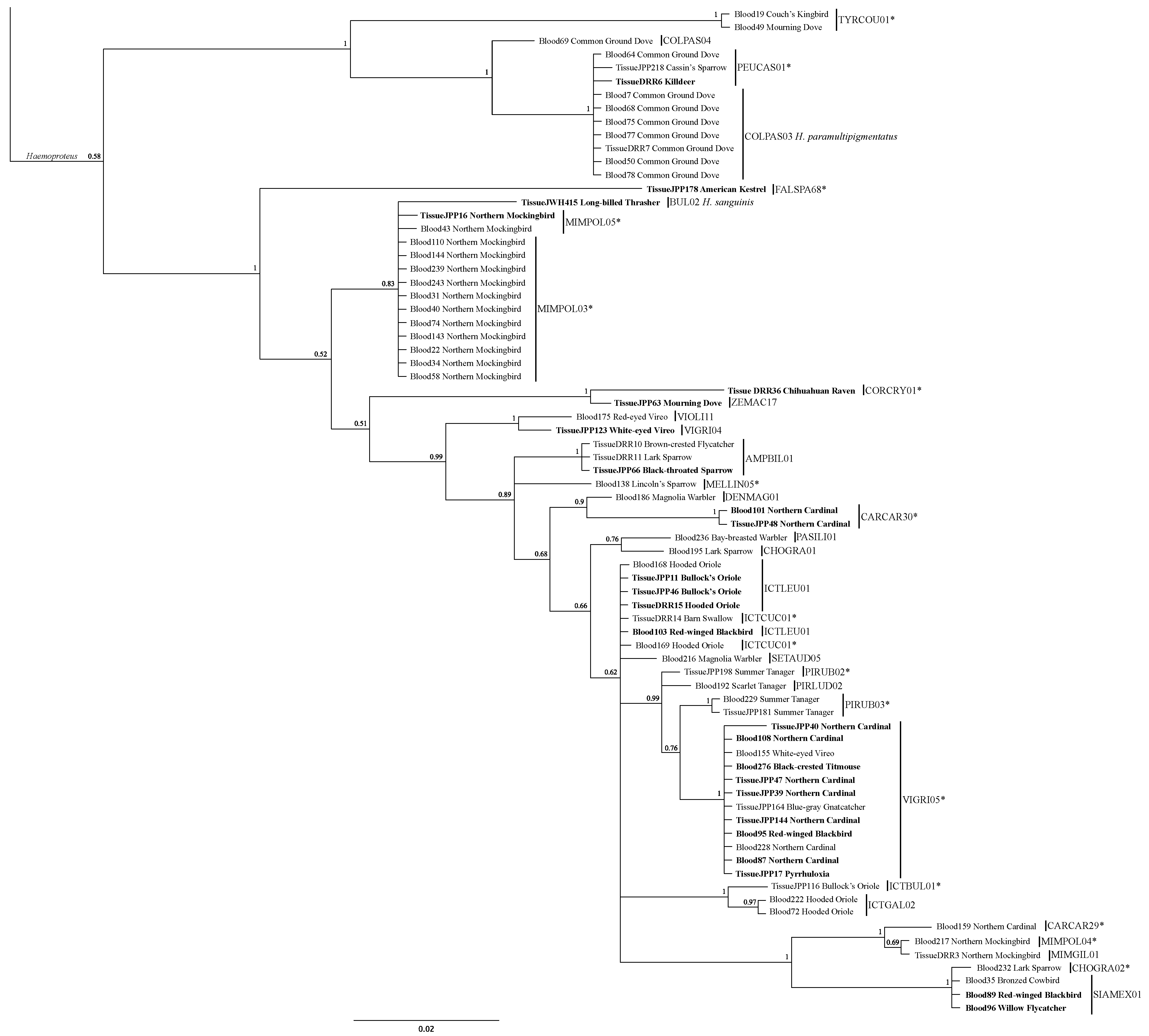
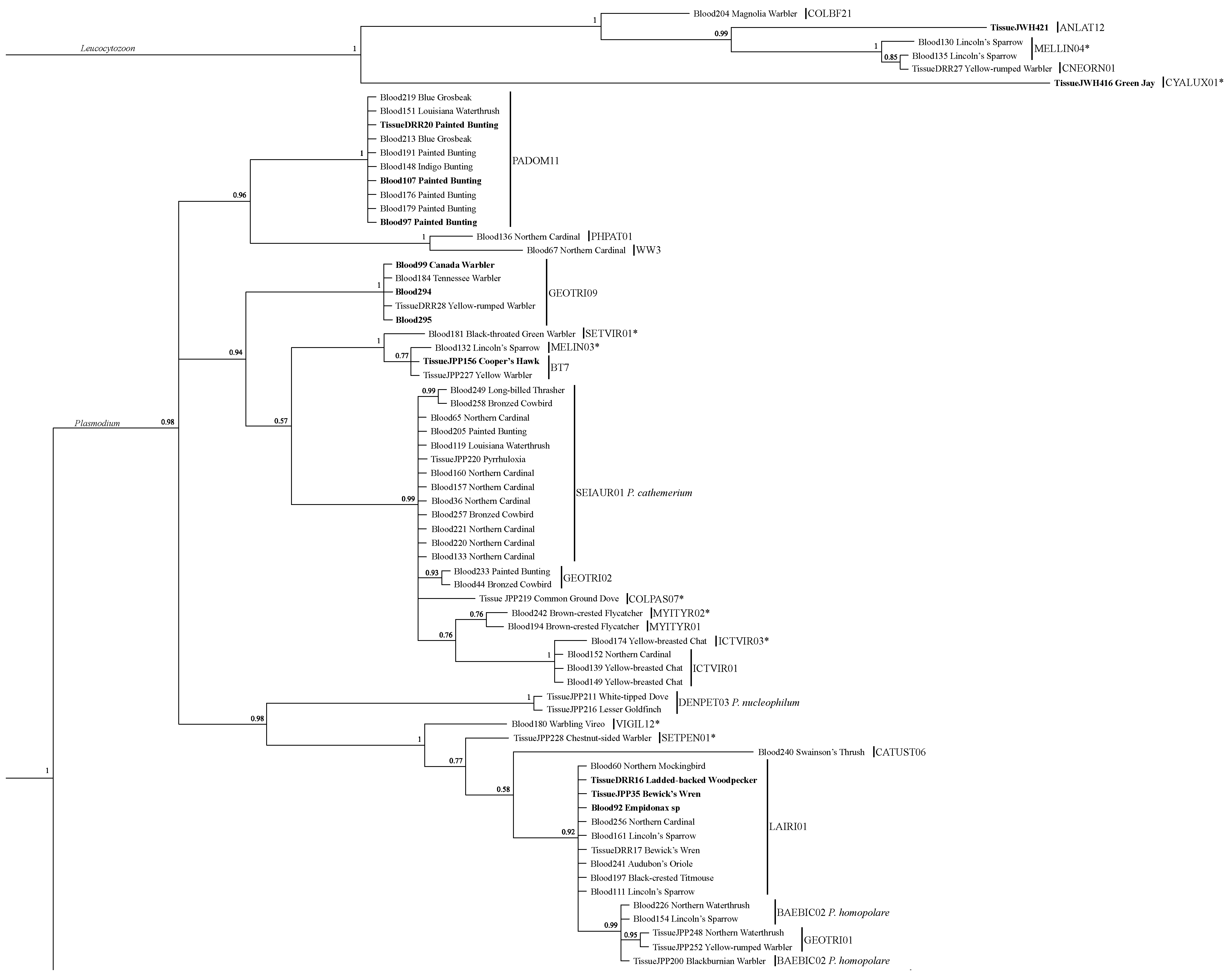
3.2. Lineage Analysis/Identification

{kind=link}
{kind=link}
{kind=link}
| MalAvi BLAST | % Match | BP Difference | Novel Lineage Designation | Location | Seasonality | |
|---|---|---|---|---|---|---|
| Haemoproteus | ||||||
| American Kestrel (Falco sparverius) | (BNOW03) | 99 | 5 | FALSPA68 | SAV | winter |
| Cassin’s Sparrow (Peucaea cassinii) | (COLPAS03) | 99 | 1 | PEUCAS01 | ES | year-round |
| Killdeer (Charadrius vociferus) | (COLPAS03) | 99 | 1 | PEUCAS01 | SAV | year-round |
| Chihuahuan Raven (Corvus cryptoleucus) | (CYGNUS01) | 99 | 6 | CORCRY01 | SAV | year-round |
| Northern Cardinal (Cardinalis cardinalis) (2) | (GRBRU02) | 98 | 11 | CARCAR30 | SAV (2) | year-round |
| Bullocks Oriole (Icterus bullockii) | (ICTGAL02) | 99 | 5 | ICTBUL01 | SAV | breeding |
| Barn Swallow (Hirundo rustica) | (ICTLEU01) | 99 | 1 | ICTCUC01 | ES | breeding |
| Hooded Oriole (Icterus cucullatus) | (ICTLEU01) | 99 | 1 | ICTCUC01 | ES | breeding |
| Northern Mockingbird (Mimus polyglottos) (11) | (MAFUS02) | 99 | 1 | MIMPOL03 | ES (11) | year-round |
| Northern Mockingbird (Mimus polyglottos) (2) | (MAFUS02) | 99 | 2 | MIMPOL05 | ES (1), SAV (1) | year-round |
| Northern Cardinal (Cardinalis cardinalis) | (MIMGIL01) | 99 | 4 | CARCAR29 | ES | year-round |
| Northern Mockingbird (Mimus polyglottos) | (MIMGIL01) | 99 | 1 | MIMPOL04 | ES | year-round |
| Summer Tanager (Piranga rubra) | (PACPEC02) | 99 | 1 | PIRUB02 | ES | breeding |
| Summer Tanager (Piranga rubra) (2) | (PIRFLA01) | 99 | 1 | PIRUB03 | ES (2) | breeding |
| Lincoln’s Sparrow (Melospiza lincolnii) | (PIRLUD01) | 99 | 7 | MELLIN05 | ES | winter |
| Lark Sparrow (Chondestes grammacus) | (SIAMEX01) | 99 | 1 | CHOGRA02 | ES | winter |
| Black-crested Titmouse (Baeolophus atricistatus) | (WILPUS01) | 99 | 3 | VIGRI05 | SAV | year-round |
| Blue-gray Gnatcatcher (Polioptila caerulea) | (WILPUS01) | 99 | 3 | VIGRI05 | ES | year-round |
| Northern Cardinal (Cardinalis cardinalis) (7) | (WILPUS01) | 99 | 3 | VIGRI05 | ES (1), SAV (6) | year-round |
| Pyrrhuloxia (Cardinalis sinuatus) | (WILPUS01) | 99 | 3 | VIGRI05 | SAV | year-round |
| Red-winged Blackbird (Agelaius phoeniceus) | (WILPUS01) | 99 | 3 | VIGRI05 | SAV | year-round |
| White-eyed Vireo (Vireo griseus) | (WILPUS01) | 99 | 3 | VIGRI05 | ES | year-round |
| Couch’s Kingbird (Tyrannus couchii) | (ZEAUR05) | 99 | 2 | TYRCOU01 | ES | year-round |
| Mourning Dove (Zenaida macroura) | (ZEAUR05) | 99 | 2 | TYRCOU01 | ES | year-round |
| Black-throated Sparrow (Amphispiza bilineata) | AMPBIL01 | 100 | 0 | SAV | year-round | |
| Brown-crested Flycatcher (Myiarchus tyrannulus) | AMPBIL01 | 100 | 0 | ES | breeding | |
| Lark Sparrow (Chondestes grammacus) | AMPBIL01 | 100 | 0 | ES | winter | |
| Long-billed Thrasher (Toxostoma longirostre) | BUL2 | 100 | 0 | SAV | year-round | |
| Lark Sparrow (Chondestes grammacus) | CHOGRA01 | 100 | 0 | ES | winter | |
| Common Ground Dove (Columbina passerina) (8) | COLPAS03 | 100 | 0 | ES (8) | year-round | |
| Common Ground Dove (Columbina passerina) | COLPAS04 | 100 | 0 | ES | year-round | |
| Magnolia Warbler (Setophaga magnolia) | DENMAG01 | 100 | 0 | ES | migrant | |
| Hooded Oriole (Icterus cucullatus) (2) | ICTGAL02 | 100 | 0 | ES (2) | breeding | |
| Bullocks Oriole (Icterus bullockii) (2) | ICTLEU01 | 100 | 0 | SAV (2) | breeding | |
| Hooded Oriole (Icterus cucullatus) (2) | ICTLEU01 | 100 | 0 | ES (1), SAV (1) | breeding | |
| Red-winged Blackbird (Agelaius phoeniceus) | ICTLEU01 | 100 | 0 | SAV | year-round | |
| Northern Mockingbird (Mimus polyglottos) | MIMGIL01 | 100 | 0 | ES | year-round | |
| Bay-breasted Warbler (Setophaga castanea) | PASILI01 | 100 | 0 | ES | migrant | |
| Scarlet Tanager (Piranga olivacea) | PIRLUD02 | 100 | 0 | ES | migrant | |
| Magnolia Warbler (Setophaga magnolia) | SETAUD05 | 100 | 0 | ES | migrant | |
| Bronzed Cowbird (Molothrus aeneus) | SIAMEX01 | 100 | 0 | ES | breeding | |
| Red-winged Blackbird (Agelaius phoeniceus) | SIAMEX01 | 100 | 0 | SAV | year-round | |
| Willow Flycatcher (Empidonax traillii) | SIAMEX01 | 100 | 0 | SAV | migrant | |
| White-eyed Vireo (Vireo griseus) | VIGRI04 | 100 | 0 | SAV | year-round | |
| Red-eyed Vireo (Vireo olivaceus) | VIOLI11 | 100 | 0 | ES | migrant | |
| Mourning Dove (Zenaida macroura) | ZEMAC17 | 100 | 0 | SAV | year-round | |
| Leucocytozoon | ||||||
| Green Jay (Cyanocorax yncas) | (BGR3) | 89 | 52 | CYALUX01 | SAV | year-round |
| Lincoln’s Sparrow (Melospiza lincolnii) | (CNEORN01) | 99 | 2 | MELLIN04 | ES | winter |
| Varied Bunting (Passerina versicolor) | ANLAT12 | 100 | 0 | SAV | breeding | |
| Lincoln’s Sparrow (Melospiza lincolnii) | CNEORN01 | 100 | 0 | ES | winter | |
| Yellow-rumped Warbler (Setophaga coronata) | CNEORN01 | 100 | 0 | ES | winter | |
| Magnolia Warbler (Setophaga magnolia) | COLBF21 | 100 | 0 | ES | migrant | |
| Plasmodium | ||||||
| Black-throated Green Warbler (Steophaga virens) | (BT7) | 99 | 4 | SETVIR01 | ES | migrant |
| Chesnut-sided Warbler (Setophaga pensylvanica) | (CXRES06) | 98 | 9 | SETPEN01 | ES | migrant |
| Warbling Vireo (Vireo gilvus) | (CYCGUJ01) | 99 | 1 | VIGIL12 | ES | migrant |
| Yellow-breasted Chat (Icteria virens) | (ICTVIR01) | 98 | 11 | ICTVIR03 | ES | migrant |
| Lincoln’s Sparrow (Melospiza lincolnii) | (ICTVIR01) | 99 | 1 | MELIN03 | ES | winter |
| Brown-crested Flycatcher (Myiarchus tyrannulus) | (PADOM09) | 99 | 1 | MYITYR02 | ES | breeding |
| Common Ground Dove (Columbina passerina) | (SEIAUR01) | 98 | 1 | COLPAS07 | ES | year-round |
| Blackburnian Warbler (Setophaga fusca) | BAEBIC02 | 100 | 0 | ES | migrant | |
| Lincoln’s Sparrow (Melospiza lincolnii) | BAEBIC02 | 100 | 0 | ES | winter | |
| Northern Waterthrush (Parkesia noveboracensis) | BAEBIC02 | 100 | 0 | ES | migrant | |
| Cooper’s Hawk (Accipiter cooperii) | BT7 | 100 | 0 | SAV | year-round | |
| Yellow Warbler (Setophaga petechia) | BT7 | 100 | 0 | ES | migrant | |
| Swainson’s Thrush (Catharus ustulatus) | CATUST06 | 100 | 0 | ES | migrant | |
| Lesser Goldfinch (Spinus psaltria) | DENPET03 | 100 | 0 | ES | year-round | |
| White-tipped Dove (Leptotila verreauxi) | DENPET03 | 100 | 0 | ES | year-round | |
| Northern Waterthrush (Parkesia noveboracensis) | GEOTRI01 | 100 | 0 | ES | migrant | |
| Yellow-rumped Warbler (Setophaga coronata) | GEOTRI01 | 100 | 0 | ES | winter | |
| Bronzed Cowbird (Molothrus aeneus) | GEOTRI02 | 100 | 0 | ES | breeding | |
| Painted Bunting (Passerina ciris) | GEOTRI02 | 100 | 0 | ES | breeding | |
| Blood294 | GEOTRI09 | 100 | 0 | SAV | ||
| Blood295 | GEOTRI09 | 100 | 0 | SAV | ||
| Canada Warbler (Cardellina canadensis) | GEOTRI09 | 100 | 0 | SAV | migrant | |
| Tennessee Warbler (Leiothlypis peregrina) | GEOTRI09 | 100 | 0 | ES | migrant | |
| Yellow-rumped Warbler (Setophaga coronata) | GEOTRI09 | 100 | 0 | ES | winter | |
| Northern Cardinal (Cardinalis cardinalis) | ICTVIR01 | 100 | 0 | ES | year-round | |
| Yellow-breasted Chat (Icteria virens) (2) | ICTVIR01 | 100 | 0 | ES (2) | migrant | |
| Audubons Oriole (Icterus graduacauda) | LAIRI01 | 100 | 0 | ES | year-round | |
| Bewick’s Wren (Thyromanes bewickii) (2) | LAIRI01 | 100 | 0 | ES (1) SAV (1) | year-round | |
| Black-crested Titmouse (Baeolophus atricistatus) | LAIRI01 | 100 | 0 | ES | year-round | |
| Empidonax_sp. | LAIRI01 | 100 | 0 | SAV | likely migrant | |
| Ladder-backed Woodpecker (Dryobates scalaris) | LAIRI01 | 100 | 0 | SAV | year-round | |
| Lincoln’s Sparrow (Melospiza lincolnii) (2) | LAIRI01 | 100 | 0 | ES (2) | winter | |
| Northern Cardinal (Cardinalis cardinalis) | LAIRI01 | 100 | 0 | ES | year-round | |
| Northern Mockingbird (Mimus polyglottos) | LAIRI01 | 100 | 0 | ES | year-round | |
| Brown-crested Flycatcher (Myiarchus tyrannulus) | MYITYR01 | 100 | 0 | ES | breeding | |
| Blue Grosbeak (Passerina caerulea) (2) | PADOM11 | 100 | 0 | ES (2) | Breeding | |
| Indigo Bunting (Passerina cyanea) | PADOM11 | 100 | 0 | ES | breeding | |
| Louisiana Watherthrush (Parkesia motacilla) | PADOM11 | 100 | 0 | ES | migrant | |
| Painted Bunting (Passerina ciris) (6) | PADOM11 | 100 | 0 | ES (3), SAV (3) | breeding | |
| Northern Cardinal (Cardinalis cardinalis) | PHPAT01 | 100 | 0 | ES | year-round | |
| Bronzed Cowbird (Molothrus aeneus) (2) | SEIAUR01 | 100 | 0 | ES (2) | breeding | |
| Long-billed Thrasher (Toxostoma longirostre) | SEIAUR01 | 100 | 0 | ES | year-round | |
| Louisiana Watherthrush (Parkesia motacilla) | SEIAUR01 | 100 | 0 | ES | migrant | |
| Northern Cardinal (Cardinalis cardinalis) (7) | SEIAUR01 | 100 | 0 | ES (7) | year-round | |
| Painted Bunting (Passerina ciris) | SEIAUR01 | 100 | 0 | ES | breeding | |
| Pyrrhuloxia (Cardinalis sinuatus) | SEIAUR01 | 100 | 0 | ES | year-round | |
| Northern Cardinal (Cardinalis cardinalis) | WW3 | 100 | 0 | ES | year-round |
3.3. Precipitation
4. Discussion
4.1. Prevalence
4.2. Host Associations
4.3. Precipitaion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hernández-Lara, C.; Monteros, A.E.D.L.; Ibarra-Cerdeña, C.N.; García-Feria, L.; Santiago-Alarcon, D. Combining morphological and molecular data to reconstruct the phylogeny of avian Haemosporida. Int. J. Parasitol. 2018, 48, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Lachish, S.; Knowles, S.C.L.; Alves, R.; Sepil, I.; Davies, A.; Lee, S.; Wood, M.; Sheldon, B.C. Spatial determinants of infection risk in a multi-species avian malaria system. Ecography 2013, 36, 587–598. [Google Scholar] [CrossRef]
- Fecchio, A.; Clark, N.J.; Bell, J.A.; Skeen, H.R.; Lutz, H.L.; De La Torre, G.M.; Vaughan, J.A.; Tkach, V.V.; Schunck, F.; Ferreira, F.C.; et al. Global drivers of avian haemosporidian infections vary across zoogeographical regions. Glob. Ecol. Biogeogr. 2021, 30, 2393–2406. [Google Scholar] [CrossRef]
- Ellis, V.A.; Medeiros, M.C.I.; Collins, M.D.; Sari, E.H.R.; Coffey, E.D.; Dickerson, R.C.; Lugarini, C.; Stratford, J.A.; Henry, D.R.; Merrill, L.; et al. Prevalence of avian haemosporidian parasites is positively related to the abundance of host species at multiple sites within a region. Parasitol. Res. 2017, 116, 73–80. [Google Scholar] [CrossRef]
- Olsson-Pons, S.; Clark, N.; Ishtiaq, F.; Clegg, S.M. Differences in host species relationships and biogeographic influences produce contrasting patterns of prevalence, community composition and genetic structure in two genera of avian malaria parasites in southern Melanesia. J. Anim. Ecol. 2015, 84, 985–998. [Google Scholar] [CrossRef]
- Clark, N.J.; Drovetski, S.V.; Voelker, G. Robust geographical determinants of infection prevalence and a contrasting latitudinal diversity gradient for haemosporidian parasites in Western Palearctic birds. Mol. Ecol. 2020, 29, 3131–3143. [Google Scholar] [CrossRef]
- Pellegrino, I.; Ilahiane, L.; Boano, G.; Cucco, M.; Pavia, M.; Prestridge, H.L.; Voelker, G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity 2021, 13, 75. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, K.; Álvarez-Mendizábal, P.; Chapa-Vargas, L.; Escobar, F.; González-García, F.; Santiago-Alarcon, D. Haemosporidian prevalence, parasitaemia and aggregation in relation to avian assemblage life history traits at different elevations. Int. J. Parasitol. 2021, 51, 365–378. [Google Scholar] [CrossRef]
- Krama, T.; Krams, R.; Cīrule, D.; Moore, F.R.; Rantala, M.J.; Krams, I.A. Intensity of haemosporidian infection of parids positively correlates with proximity to water bodies, but negatively with host survival. J. Ornithol. 2015, 156, 1075–1084. [Google Scholar] [CrossRef]
- Wood, M.J.; Cosgrove, C.L.; Wilkin, T.A.; Knowles, S.C.L.; Day, K.P.; Sheldon, B.C. Within-population variation in prevalence and lineage distribution of avian malaria in blue tits, Cyanistes caeruleus. Mol. Ecol. 2007, 16, 3263–3273. [Google Scholar] [CrossRef]
- Muriel, J.; Marzal, A.; Magallanes, S.; García-Longoria, L.; Suarez-Rubio, M.; Bates, P.J.J.; Lin, H.H.; Soe, A.N.; Oo, K.S.; Aye, A.A.; et al. Prevalence and Diversity of Avian Haemosporidians May Vary with Anthropogenic Disturbance in Tropical Habitats in Myanmar. Diversity 2021, 13, 111. [Google Scholar] [CrossRef]
- Reis, S.; Melo, M.; Covas, R.; Doutrelant, C.; Pereira, H.; de Lima, R.; Loiseau, C. Influence of land use and host species on parasite richness, prevalence and co-infection patterns. Int. J. Parasitol. 2021, 51, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Soares, L.; Latta, S.C.; Ricklefs, R.E. Neotropical migratory and resident birds occurring in sympatry during winter have distinct haemosporidian parasite assemblages. J. Biogeogr. 2020, 47, 748–759. [Google Scholar] [CrossRef]
- Sibley, D. The Sibley Guide to Birds, 2nd ed.; Alfred A. Knopf: New York, NY, USA, 2014. [Google Scholar]
- Adler, P.H.; Roach, D.; Reeves, W.K.; Flanagan, J.P.; Morrow, M.E.; Toepfer, J.E. Attacks on the Endangered Attwater’s Prairie-Chicken (Tympanuchus Cupido Attwateri) by Black Flies (Diptera: Simuliidae) Infected with an Avian Blood Parasite. J. Vector Ecol. 2007, 32, 309. [Google Scholar] [CrossRef]
- Castle, M.D.; Christensen, B.M.; Rocke, T.E. Hematozoan parasites of rio grande wild turkeys from southern texas. J. Wildl. Dis. 1988, 24, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Bertram, M.R.; Hamer, G.L.; Hartup, B.K.; Snowden, K.F.; Medeiros, M.C.; Hamer, S.A. Haemosporida prevalence and diversity are similar in endangered wild whooping cranes (Grus americana) and sympatric sandhill cranes (Grus canadensis). Parasitology 2016, 144, 629–640. [Google Scholar] [CrossRef] [Green Version]
- Ramey, A.M.; Reed, J.A.; Walther, P.; Link, P.; Schmutz, J.A.; Douglas, D.C.; Stallknecht, D.E.; Soos, C. Evidence for the exchange of blood parasites between North America and the Neotropics in blue-winged teal (Anas discors). Parasitol. Res. 2016, 115, 3923–3939. [Google Scholar] [CrossRef]
- Walstrom, V.W.; Outlaw, D.C. Distribution and Prevalence of Haemosporidian Parasites in the Northern Cardinal (Cardinalis cardinalis). J. Parasitol. 2017, 103, 63. [Google Scholar] [CrossRef]
- Box, E.D. Blood and Tissue Protozoa of the English Sparrow (Passer domesticus domesticus) in Galveston, Texas*. J. Protozool. 1966, 13, 204–208. [Google Scholar] [CrossRef]
- Harvey, J.A.; Voelker, G. Avian haemosporidian detection across source materials: Prevalence and genetic diversity. Parasitol. Res. 2017, 116, 3361–3371. [Google Scholar] [CrossRef]
- Paul-Murphy, J.; Engilis, A.; Pascoe, P.; Williams, C. Comparsion of pentobarbital and thoracic (cardiac) compression to euthanize anesthetized sparrows (Passer domesticus) and starlings (Sturnus vulgaris). In ExoticsCon Conference Proceedings; ExoticsCon: Atlanta, GA, USA, 2016. [Google Scholar]
- Seutin, G.; White, B.N.; Boag, P. Preservation of avian blood and tissue samples for DNA analyses. Can. J. Zool. 1991, 69, 82–90. [Google Scholar] [CrossRef]
- Drovetski, S.V.; Aghayan, S.A.; Mata, V.A.; Lopes, R.J.; Mode, N.A.; Harvey, J.A.; Voelker, G. Does the niche breadth or trade-off hypothesis explain the abundance-occupancy relationship in avian Haemosporidia? Mol. Ecol. 2014, 23, 3322–3329. [Google Scholar] [CrossRef] [PubMed]
- “Kite-Eating Tree” Copyright (C) 2017, R version 3.4.3; Platform: x86_64-apple-darwin15.6.0 (64-bit); The R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 20 March 2022).
- RStudio Team. RStudio: Integrated Development Environment for R.; RStudio, Inc.: Boston, MA, USA, 2015; Available online: https://www.rstudio.com/ (accessed on 20 March 2022).
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Outlaw, D.C.; Ricklefs, R.E. Species limits in avian malaria parasites (Haemosporida): How to move forward in the molecular era. Parasitology 2014, 141, 1223–1232. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In 2010 Gateway Computing Environments Workshop (GCE); IEEE: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1. 6. 2014. Available online: https://bioweb.pasteur.fr/packages/pack@Tracer@v1.6 (accessed on 20 March 2022).
- Rambaut, A. FigTree v1. 4.; University of Edinburgh: Edinburgh, UK, 2014. [Google Scholar]
- Sutcliffe, J.F. Black fly host location: A review. Can. J. Zool. 1986, 64, 1041–1053. [Google Scholar] [CrossRef]
- Njabo, K.Y.; Cornel, A.J.; Sehgal, R.N.M.; Loiseau, C.; Buermann, W.; Harrigan, R.J.; Pollinger, J.; Valkiūnas, G.; Smith, T.B. Coquillettidia (Culicidae, Diptera) mosquitoes are natural vectors of avian malaria in Africa. Malar. J. 2009, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Lamerton, J.F. A Key to the Genera of Hippoboscidae (Diptera: Pupipara), and to the Species of Hippobosca, in Africa. East Afr. Agric. For. J. 1965, 31, 1–7. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides Biting Midges: Their Role as Arbovirus Vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef] [PubMed]
- ArcGIS Release 10; Environmental Systems Research Institute (ESRI): Redlands, CA, USA, 2012.
- Bodden, H.N.; Outlaw, D.C. Diversity of Haemosporidian Parasites in Mississippi Songbirds. Southeast. Nat. 2019, 18, 314–320. [Google Scholar] [CrossRef]
- Mata, V.C.A. A Comparison of Avian Haemosporidian Parasite Communities across the Strait of Gibraltar. Masters Thesis, University of Porto, Lisbon, Portugal, 2012. [Google Scholar]
- Fecchio, A.; Wells, K.; Bell, J.A.; Tkach, V.V.; Lutz, H.L.; Weckstein, J.D.; Clegg, S.M.; Clark, N.J. Climate variation influences host specificity in avian malaria parasites. Ecol. Lett. 2019, 22, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Longoria, L.; Muriel, J.; Magallanes, S.; Villa-Galarce, Z.H.; Ricopa, L.; Inga-Díaz, W.G.; Fong, E.; Vecco, D.; Guerra-Saldaña, C.; Salas-Rengifo, T.; et al. Diversity and host assemblage of avian haemosporidians in different terrestrial ecoregions of Peru. Curr. Zool. 2022, 68, 27–40. [Google Scholar] [CrossRef]
- Levin, I.I.; Valkiūnas, G.; Santiago-Alarcon, D.; Cruz, L.L.; Iezhova, T.A.; O’Brien, S.L.; Hailer, F.; Dearborn, D.; Schreiber, E.A.; Fleischer, R.C.; et al. Hippoboscid-transmitted Haemoproteus parasites (Haemosporida) infect Galapagos Pelecaniform birds: Evidence from molecular and morphological studies, with a description of Haemoproteus iwa. Int. J. Parasitol. 2011, 41, 1019–1027. [Google Scholar] [CrossRef]
- Jaramillo, M.; Rohrer, S.; Parker, P.G. From Galapagos doves to passerines: Spillover of Haemoproteus multipigmentatus. Int. J. Parasitol. Parasites Wildl. 2017, 6, 155–161. [Google Scholar] [CrossRef]
- Baker, J.R. A Review of the Role Played by the Hippoboscidae (Diptera) as Vectors of Endoparasites. J. Parasitol. 1967, 53, 412. [Google Scholar] [CrossRef]
- McDermott, E.G.; Mullens, B.A. Desiccation Tolerance in the Eggs of the Primary North American Bluetongue Virus Vector, Culicoides sonorensis (Diptera: Ceratopogonidae), and Implications for Vector Persistence. J. Med. Entomol. 2014, 51, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- Mukhin, A.; Palinauskas, V.; Platonova, E.; Kobylkov, D.; Vakoliuk, I.; Valkiūnas, G. The Strategy to Survive Primary Malaria Infection: An Experimental Study on Behavioural Changes in Parasitized Birds. PLoS ONE 2016, 11, e0159216. [Google Scholar] [CrossRef] [Green Version]
- Keith, K.D.; Weesner, R.; Voelker, G. Malavi Meta. Texas A&M University, College Station, TX 77843, USA. 2022; Unpublished Manuscript. [Google Scholar]

| Number | Detection Rate | ||||||
|---|---|---|---|---|---|---|---|
| Total | Positive | Negative | Overall | Haemoproteus | Leucocytozoon | Plasmodium | |
| Overall | 576 | 148 | 428 | 25.69% | 50.68% | 4.05% | 42.57% |
| Blood | 281 | 103 | 178 | 36.65% | 46.60% | 2.91% | 46.60% |
| Tissue | 295 | 45 | 250 | 15.25% | 60.00% | 6.67% | 33.33% |
| SAV | 260 | 38 | 222 | 14.62% | 44.55% | 3.64% | 48.18% |
| ES | 316 | 110 | 206 | 34.81% | 68.42% | 5.26% | 26.32% |
| Lineage | ES | SAV | Both | Total |
|---|---|---|---|---|
| Haemoproteus | 13/12 | 6/7 | 3/3 | 16/16 (32) |
| Leucocytozoon | 2/1 | 1/1 | 0/0 | 3/2 (5) |
| Plasmodium | 14/7 | 4/0 | 4/0 | 14/7 (21) |
| Lineage | Blood Only | Tissue Only | Both | Total |
| Haemoproteus | (43.8%) 14 | (34.4%) 11 | 7 | 32 |
| Leucocytozoon | (40.0%) 2 | (40.0%) 2 | 1 | 5 |
| Plasmodium | (52.4%) 11 | (28.6%) 6 | 4 | 21 |
| Dataset | Positive | Negative | df | X2 | p-Value | FET | 95% CI | p-Value | |
|---|---|---|---|---|---|---|---|---|---|
| A | Blood | 103 | 178 | 1 | 33.41 | <0.001 | 3.208 | 2.121–4.910 | <0.001 |
| Tissue | 45 | 250 | |||||||
| ES | 110 | 206 | 1 | 29.42 | <0.001 | 3.114 | 2.028–4.857 | <0.001 | |
| B | SAV | 38 | 222 | ||||||
| ES | SAV | ||||||||
| Blood | 89 | 14 | 1 | 23.878 | <0.001 | 7.143 | 2.992–17.779 | <0.001 | |
| C | Tissue | 21 | 24 | ||||||
| Haemoproteus | Plasmodium | ||||||||
| Blood | 48 | 48 | 1 | 1.862 | 0.172 | 0.558 | 0.243–1.244 | 0.140 | |
| D | Tissue | 27 | 15 | ||||||
| ES | 49 | 53 | 1 | 3.335 | 0.021 | 2.792 | 1.159–7.188 | 0.019 | |
| E | SAV | 26 | 10 |
| Haemoproteus | Breeding | Migrant | Winter | Resident |
|---|---|---|---|---|
| El Sauz | 10 | 5 | 4 | 30 |
| San AV | 4 | 1 | 1 | 33 |
| total | 14 | 6 | 5 | 63 |
| Leucocytozoon | ||||
| El Sauz | 0 | 1 | 3 | 0 |
| San AV | 1 | 0 | 0 | 1 |
| total | 1 | 1 | 3 | 1 |
| Plasmodium | ||||
| El Sauz | 13 | 14 | 6 | 20 |
| San AV | 3 | 2 | 0 | 3 |
| total | 16 | 16 | 6 | 23 |
| Non-Resident | Resident | df | X2 | p-Value | |
|---|---|---|---|---|---|
| Haemoproteus | 25 | 63 | 1 | 16.95 | <0.001 |
| Plasmodium | 38 | 23 | |||
| ES | 56 | 50 | 1 | 10.93 | <0.001 |
| SAV | 12 | 37 |
| Locality | 2011 | 2012 | 2013 | 2014 | 2015 |
|---|---|---|---|---|---|
| El Sauz (coastal) | 38.44 | 41.32 | 60.06 | 97.38 | 100.4 |
| San Antonio Viejo (inland) | 26.51 | 40.33 | 54.57 | 47.5 | 74.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keith, K.D.; Pistone, J.P.; Campbell, T.A.; Voelker, G.A. Avian Haemosporidian Diversity in South Texas: New Lineages and Variation in Prevalence between Sampling Sources and Sites. Diversity 2022, 14, 378. https://doi.org/10.3390/d14050378
Keith KD, Pistone JP, Campbell TA, Voelker GA. Avian Haemosporidian Diversity in South Texas: New Lineages and Variation in Prevalence between Sampling Sources and Sites. Diversity. 2022; 14(5):378. https://doi.org/10.3390/d14050378
Chicago/Turabian StyleKeith, Katrina D., John P. Pistone, Tyler A. Campbell, and Gary A. Voelker. 2022. "Avian Haemosporidian Diversity in South Texas: New Lineages and Variation in Prevalence between Sampling Sources and Sites" Diversity 14, no. 5: 378. https://doi.org/10.3390/d14050378
APA StyleKeith, K. D., Pistone, J. P., Campbell, T. A., & Voelker, G. A. (2022). Avian Haemosporidian Diversity in South Texas: New Lineages and Variation in Prevalence between Sampling Sources and Sites. Diversity, 14(5), 378. https://doi.org/10.3390/d14050378