
Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon


Abstract
1. Introduction
2. Materials and Methods
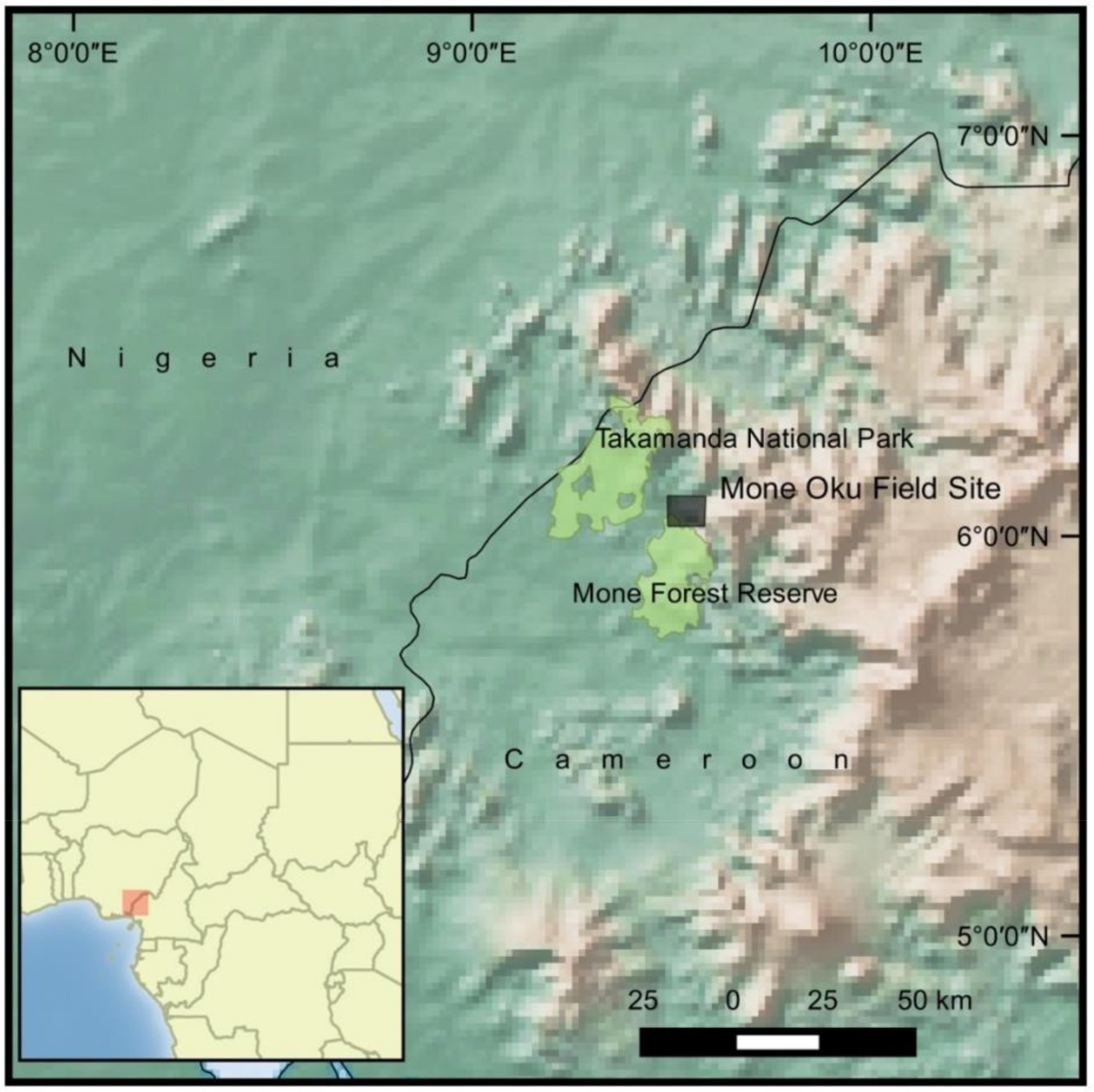
2.1. Study Site
2.2. Ethnographic Data Collection and Analysis
2.3. Ecological Data Collection and Analysis
2.3.1. Surveying
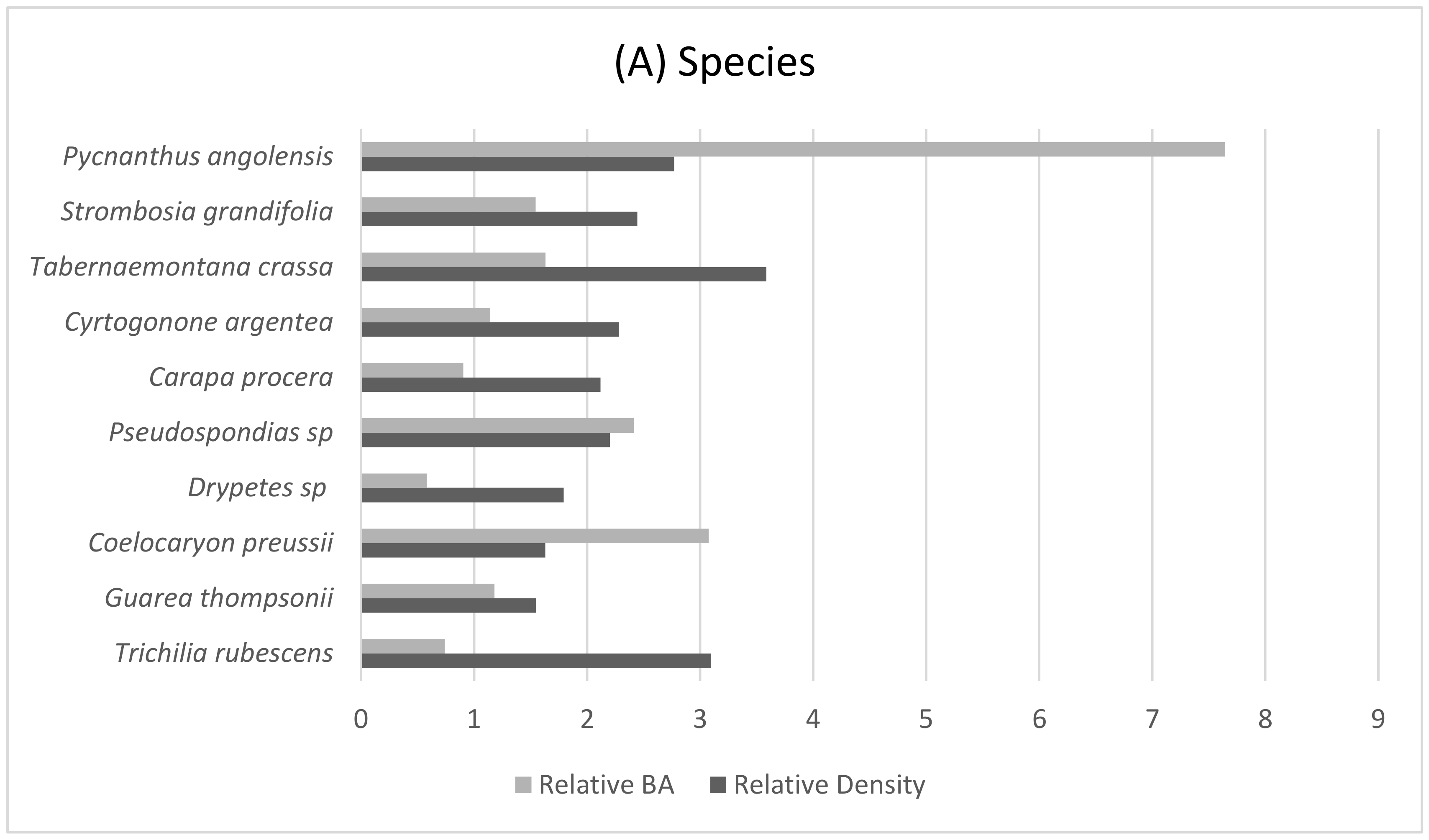
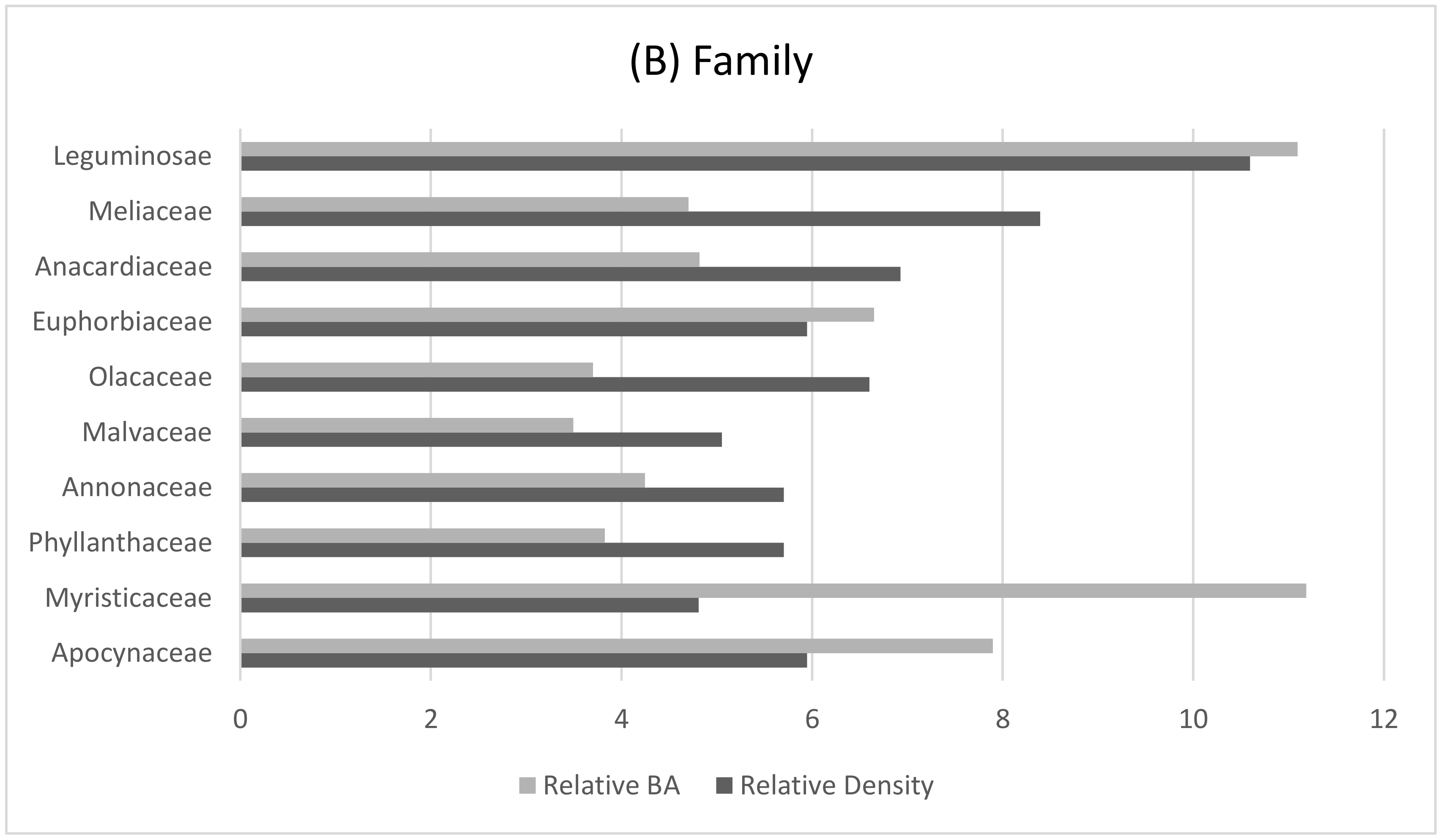
2.3.2. Botanical Sampling
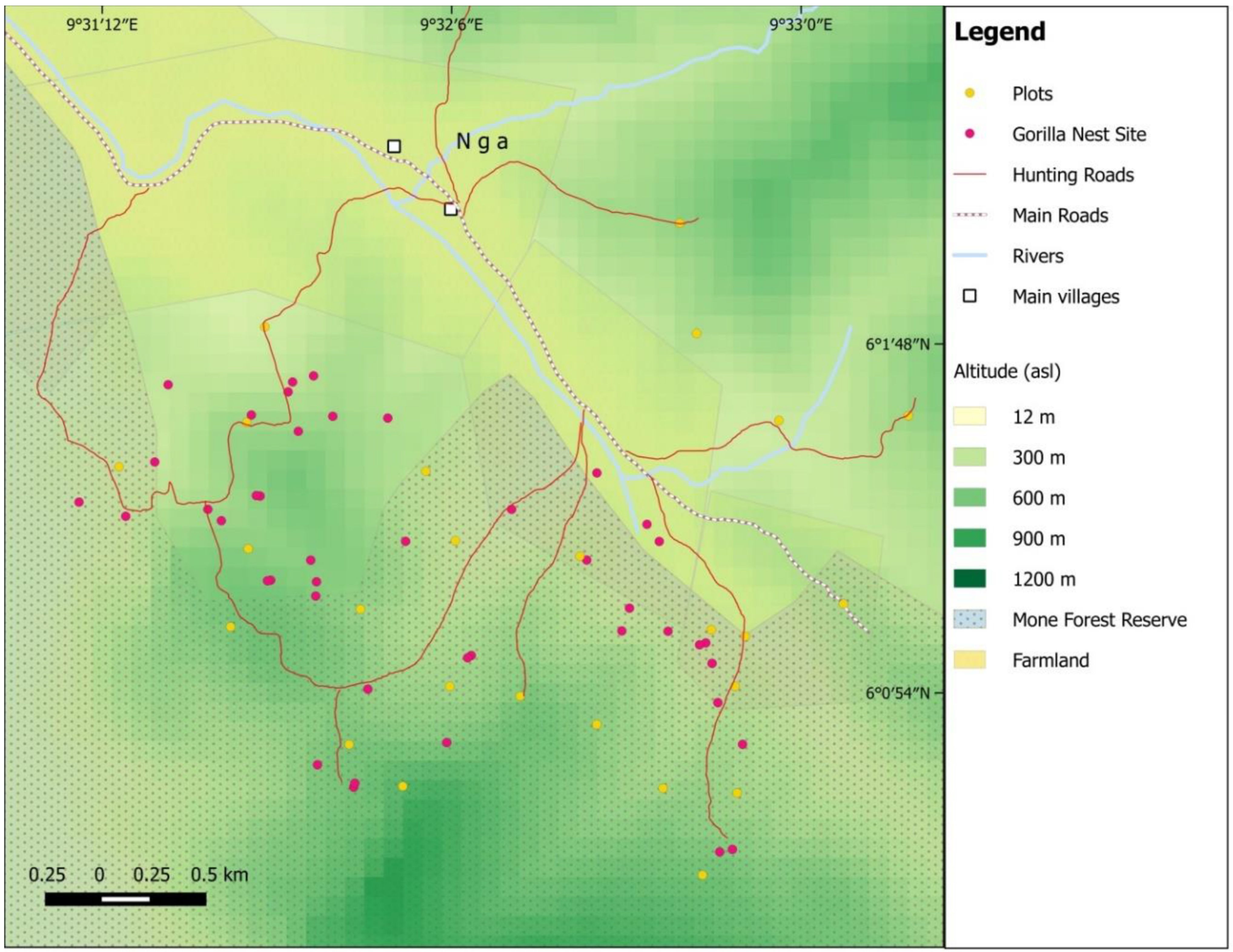
2.3.3. Nest Sites
2.3.4. Nest Site Analysis
2.3.5. Nesting Plant Preferences
2.3.6. Human Forest Use
2.4. Ethical Note
3. Results and Discussion
3.1. Characteristics of the Gorilla Nesting Environment
3.2. How Humans Have Contributed to Shaping the Current Gorilla Nesting Range
… the conviction remained that much of the rainforest consisted of dense natural vegetation that was unaffected by human settlement and which had few resources to offer. Nothing could be further from the truth and it is now realized that most of the forest has been subject to shifting agricultural exploitation at one time or another, and in some areas many times over [1] (pp. 112–113).
As everywhere the bulk of agricultural work is done by the women. The men clear new bush and set their Plantains after which they do little else. The area is cultivated by one woman, including her husband’s plantain patch, is approximately one acre, of which two fifths is plantains, two fifths is cocoyams, and one fifth luxury crops (corn, beans, pepper, groundnuts, pumpkins and gourds or tobacco) [92] (p. 40).
Ku [gorilla] sleep for ground, broken sticks put them for ground. As you get close, they call and you stay quiet. I kill gorilla when eating chop [food], then carried them to use for country juju [medicine]. When die, put Ku for seat and tie up place where shot … After tied to chair, carry Ku to that back place where women would dance and dance before butchering.
… we came to here to clear black bush [primary forest], burn all the bush. For night elephant comes for church house, tigers [leopards] came, carry goats to bush, I was the greatest hunter, I go shoot dem tigers through the door with dem powder guns. Corn was fine for here, as was mushroom from trees that had been felled. That time no palm, so we went bush to cut those tall ones, aye, we suffered those times… Some go back, as said the Tito place is bad. People shooting others with guns, me shot here [ribs] sutee!! After death where one shot, they scattered. Some decided for here and some for up.
3.3. Cacao and Wider Connections
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Connah, G. Forgotten Africa: An Introduction to Its Archaeology; Taylor and Francis: Oxfordshire, UK, 2004. [Google Scholar]
- Oates, J.F. Workshop on the conservation of Cross River Gorillas. Gorilla J. 2001, 22, 26–27. [Google Scholar]
- Sarmiento, E.E.; Oates, J.F. The Cross River gorillas: A distinct subspecies Gorilla gorilla diehli Matschie 1904. Am. Mus. Novit. 2000, 3304, 1–55. [Google Scholar] [CrossRef]
- Gorilla gorilla ssp. diehli. The IUCN Red List of Threatened Species 2016. Available online: https://www.iucnredlist.org/species/39998/102326240 (accessed on 3 January 2020).
- Wade, A.H. Shared Landscapes: The Human-Ape Interface within the Mone-Oku Forest, Cameroon. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2020. [Google Scholar]
- Setchell, J.M.; Fairet, E.; Shutt, K.; Waters, S.; Bell, S. Biosocial conservation: Integrating biological and ethnographic methods to study human–primate interactions. Int. J. Primatol. 2017, 38, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Dore, K.M. Ethnoprimatology without conservation: The political ecology of farmer–green monkey (Chlorocebus sabaeus) relations in St. Kitts, West Indies. Int. J. Primatol. 2018, 39, 918–944. [Google Scholar] [CrossRef]
- Riley, E.P.; Fuentes, A.; Dore, K.M. Introduction: Doing Ethnoprimatology in the Anthropocene. In Ethnoprimatology: A Practical Guide to Research at the Human-Nonhuman Primate Interface; Dore, K.M., Riley, E.P., Fuentes, A., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 1–6. [Google Scholar]
- Malone, N.; Palmer, A.; Wade, A.H. Incorporating the ethnographic perspective: The value, process, and responsibility of working with human participants. In Ethnoprimatology: A Practical Guide to Research at the Human-Nonhuman Primate Interface; Dore, K.M., Riley, E.P., Fuentes, A., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 176–189. [Google Scholar]
- Parathian, H.E.; McLennan, M.R.; Hill, C.M.; Frazão-Moreira, A.; Hockings, K.J. Breaking through disciplinary barriers: Human–wildlife interactions and multispecies ethnography. Int. J. Primatol. 2018, 39, 749–775. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.P. The Maturation of ethnoprimatology: Theoretical and methodological pluralism. Int. J. Primatol. 2018, 39, 705–729. [Google Scholar] [CrossRef]
- Riley, E.P.; Ellwanger, A.L. Methods in ethnoprimatology: Exploring the human-nonhuman primate interface. In Primate Ecology and Conservation: A Handbook of Techniques; Sterling, E., Bynum, N., Blair, M., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 128–150. [Google Scholar]
- Ellwanger, A.L.; Lambert, J.E. Investigating niche construction in dynamic human-animal landscapes: Bridging ecological and evolutionary timescales. Int. J. Primatol. 2018, 39, 797–816. [Google Scholar] [CrossRef]
- Jost Robinson, C.A.; Remis, M.J. Engaging holism: Exploring multispecies approaches in ethnoprimatology. Int. J. Primatol. 2018, 39, 776–796. [Google Scholar] [CrossRef]
- Malone, N.; Wade, A.H.; Fuentes, A.; Riley, E.P.; Remis, M.; Robinson, C.J. Ethnoprimatology: Critical interdisciplinarity and multispecies approaches in anthropology. Crit. Anthropol. 2014, 34, 8–29. [Google Scholar] [CrossRef]
- Escobar, A. Whose knowledge, whose nature? Biodiversity, conservation, and the political ecology of social movements. J. Political Ecol. 1998, 5, 53–82. [Google Scholar] [CrossRef]
- Sousa, J.; Hill, C.M.; Ainslie, A. Chimpanzees, sorcery and contestation in a protected area in Guinea-Bissau. Soc. Anthropol. 2017, 25, 364–379. [Google Scholar] [CrossRef]
- Escobar, A. After nature: Steps to an antiessentialist political ecology. Curr. Anthropol. 1999, 40, 1–30. [Google Scholar] [CrossRef]
- Blaikie, P.; Brookfield, H. Land Degradation and Society; Methuen: London, UK, 1987. [Google Scholar]
- Forsyth, T. Critical Political Ecology: The Politics of Environmental Science; Routledge: London, UK, 2004. [Google Scholar]
- Nygren, A.; Rikoon, S. Political ecology revisited: Integration of politics and ecology does matter. Soc. Nat. Resour. 2008, 21, 767–782. [Google Scholar] [CrossRef]
- Stott, P.; Sullivan, S. Political Ecology: Science, Myth and Power; Arnold Publishers: London, UK, 2000. [Google Scholar]
- Wolf, E. Ownership and political ecology. Anthropol. Q. 1972, 45, 201–205. [Google Scholar] [CrossRef]
- Fairhead, J.; Leach, M. Reframing Deforestation: Global Analyses and Local Realities: Studies in West Africa; Routledge: London, UK, 1998. [Google Scholar]
- Jones, S. A political ecology of wildlife conservation in Africa. Rev. Afr. Political Econ. 2006, 33, 483–495. [Google Scholar] [CrossRef]
- Mulder, M.B.; Coppolillo, P. Conservation: Linking Ecology, Economics, and Culture; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Neumann, R.P. Making Political Ecology; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Nelson, J.G.; Serafin, R. Assessing biodiversity: A human ecological approach. Ambio 1992, 21, 212–218. [Google Scholar]
- Gezon, L.L. Global Visions, Local Landscapes: A Political Ecology of Conservation, Conflict, and Control in Northern Madagascar; Altamira Press: Lanham, MA, USA, 2006. [Google Scholar]
- Dunn, A.; Bergl, R.; Byler, D.; Eben-Ebai, S.; Etiendem, D.N.; Fotso, R.; Ikfuingei, R.; Imong, I.; Jameson, C.; Macfie, E.J.; et al. Revised Regional Action Plan for the Conservation of the Cross River Gorilla (Gorilla gorilla diehli) 2014–2019; IUCN/SSC Primate Specialist Group and Wildlife Conservation Society: New York, NY, USA, 2014. [Google Scholar]
- McFarland, K.L. Ecology of Cross River gorillas (Gorilla gorilla diehli) on Afi Mountain, Cross River State, Nigeria. Ph.D. Thesis, City University of New York, New York, NY, USA, 2007. [Google Scholar]
- Oates, J.F.; Bergl, R.A.; Linder, J.M. Africa’s Gulf of Guinea Forests: Biodiversity Patterns and Conservation Priorities; Conservation International Center for Applied Biodiversity Science: Washington, DC, USA, 2004. [Google Scholar]
- Bergl, R.A.; Vigilant, L. Genetic analysis reveals population structure and recent migration within the highly fragmented range of the Cross River gorilla (Gorilla gorilla diehli). Mol. Ecol. 2007, 16, 501–516. [Google Scholar] [CrossRef]
- Bergl, R.A.; Bradley, B.J.; Nsubuga, A.; Vigilant, L. Effects of habitat fragmentation, population size and demographic history on genetic diversity: The Cross River gorilla in a comparative context. Int. J. Primatol. 2008, 70, 848–859. [Google Scholar] [CrossRef]
- Oates, J.F.; McFarland, K.L.; Groves, J.L.; Bergl, R.A.; Linder, J.M.; Disotell, T.R. The Cross River gorilla: The natural history and status of a neglected and critically endangered subspecies. In Gorilla Biology: A Multidisciplinary Perspective; Taylor, A., Goldsmith, M., Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 472–497. [Google Scholar]
- Sawyer, S.C. The Ecology and Conservation of the Critically Endangered Cross River Gorilla in Cameroon. Ph.D. Thesis, University of California, Oakland, CA, USA, 2012. [Google Scholar]
- Sunderland-Groves, J.L.; Ekinde, A.; Mboh, H. Cross River gorilla (Gorilla gorilla diehli) nesting behaviour at Kagwene Mountain, Cameroon: Implications for assessing group size and density. Int. J. Primatol. 2009, 30, 253–266. [Google Scholar] [CrossRef]
- Imong, I.; Robbins, M.M.; Mundry, R.; Bergl, R.; Kühl, H.S. Distinguishing ecological constraints from human activity in species range fragmentation: The case of Cross River gorillas. Anim. Conserv. 2014, 17, 323–331. [Google Scholar] [CrossRef]
- Etiendem, D.N. Ecological and Anthropogenic Factors of Cross River Gorilla (Gorilla gorilla diehli) Survival at Mawambi Hills, Southwest Cameroon. Ph.D. Thesis, Vrije Universiteit, Brussels, Belgium, 2013. [Google Scholar]
- Etiendem, D.N.; Tagg, N. Feeding ecology of Cross River gorillas (Gorilla gorilla diehli) at Mawambi Hills: The influence of resource seasonality. Int. J. Primatol. 2013, 34, 1261–1280. [Google Scholar] [CrossRef]
- Etiendem, D.N.; Funwi-Gabga, N.; Tagg, N.; Hens, L.; Indah, E.K. The Cross River gorillas (Gorilla gorilla diehli) at Mawambi Hills, south-west Cameroon: Habitat suitability and vulnerability to anthropogenic disturbance. Folia Primatol. 2013, 84, 18–31. [Google Scholar] [CrossRef]
- De Vere, R.A.; Warren, Y.; Nicholas, A.; Mackenzie, M.E.; Higham, J.P. Nest site ecology of the Cross River gorilla at the Kagwene Gorilla Sanctuary, Cameroon, with special reference to anthropogenic influence. Am. J. Primatol. 2011, 73, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, S.C.; Brashares, J.S. Applying resource selection functions at multiple scales to prioritize habitat use by the endangered Cross River gorilla. Divers. Distrib. 2013, 19, 943–954. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa, A Descriptive Memoir to Accompany the UNESCO/AETFAT/UNSO Vegetation Map of Africa (3 Plates, Northwestern African, Northeastern Africa, and Southern Africa, 1:5,000,000); UNESCO: Paris, France, 1983. [Google Scholar]
- Mboh, H.; Warren, Y. Large Mammal Survey of the Proposed Takamanda National Park; Unpublished Report; Wildlife Conservation Society: Limbe, Cameroon, 2007. [Google Scholar]
- Jameson, C. Gorilla Guardian update: Expansion of the community-based monitoring network. Gorilla J. 2012, 45, 13–15. [Google Scholar]
- Blommaert, J.; Jie, D. Ethnographic Fieldwork: A Beginner’s Guide; Multilingual Matters: Bristol, UK, 2010. [Google Scholar]
- Dewalt, K.M.; Dewalt, B.R. Participant Observation: A Guide for Fieldworkers, 1st ed.; AltaMira Press: Walnut Creek, CA, USA, 2002. [Google Scholar]
- Spradley, J.P. Participant Observation; Holt, Rinehart and Winston: New York, NY, USA, 1980. [Google Scholar]
- Davies, C.A. Reflexive Ethnography: A Guide to Researching Selves and Others; Routledge: London, UK, 1999. [Google Scholar]
- Kühl, H.; Maisels, F.; Ancrenaz, M.; Williamson, E.A. Best Practice Guidelines for Surveys and Monitoring of Great Ape Populations; IUCN SSC Primate Specialist Group: Gland, Switzerland, 2008. [Google Scholar]
- Brownlow, A.R.; Plumptre, A.J.; Reynolds, V.; Ward, R. Sources of variation in the nesting behavior of chimpanzees (Pan troglodytes schweinfurthii) in the Budongo Forest, Uganda. Am. J. Primatol. 2001, 55, 49–55. [Google Scholar] [CrossRef]
- The Plant List: A Working List of all Plant Species. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 14 July 2015).
- Sunderland-Groves, J.L.; Maisels, F.; Ekinde, A. Surveys of the Cross River Gorilla and chimpanzee populations in Takamanda Forest Reserve, Cameroon. In Takamanda—The Biodiversity of an African Rainforest; Comiskey, J.A., Sunderland, T.C.H., Sunderland-Groves, J.L., Eds.; Smithsonian Institution: Washington, DC, USA, 2003; pp. 129–140. [Google Scholar]
- White, L.J.T.; Edwards, A. Conservation Research in the African Rain Forest: A Technical Handbook; Wildlife Conservation Society: New York, NY, USA, 2000. [Google Scholar]
- Fruth, B.; Hohmann, G. Nest building behavior in the great apes: The great leap forward? In Great Ape Societies; McGrew, W.C., Merchant, L.F., Nishida, T., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 225–240. [Google Scholar]
- Stewart, F.A.; Pruetz, J.D.; Hansell, M.H. Do chimpanzees build comfortable nests? Am. J. Primatol. 2007, 69, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Koops, K.; McGrew, W.C.; de Vries, H.; Matsuzawa, T. Nest-building by chimpanzees (Pan troglodytes verus) at Seringbara, Nimba Mountains: Antipredation, thermoregulation, and antivector hypotheses. Int. J. Primatol. 2012, 33, 356–380. [Google Scholar] [CrossRef]
- Tutin, C.E.; Parnell, R.J.; White, L.J.; Fernandez, M. Nest building by lowland gorillas in the Lopé Reserve, Gabon: Environmental influences and implications for censusing. Int. J. Primatol. 1995, 16, 53–76. [Google Scholar] [CrossRef]
- Samson, D.R.; Hunt, K.D. A thermodynamic comparison of arboreal and terrestrial sleeping sites for dry-habitat chimpanzees (Pan troglodytes schweinfurthii) at the Toro-Semliki Wildlife Reserve, Uganda. Am. J. Primatol. 2012, 74, 811–818. [Google Scholar] [CrossRef]
- Van Casteren, A.; Sellers, W.I.; Thorpe, S.K.; Coward, S.; Crompton, R.H.; Myatt, J.P.; Ennos, A.R. Nest-building orangutans demonstrate engineering know-how to produce safe, comfortable beds. Proc. Natl. Acad. Sci. USA 2012, 109, 6873–6877. [Google Scholar] [CrossRef]
- Rothman, J.M.; Pell, A.N.; Dierenfeld, E.S.; Mccann, C.M. Plant choice in the construction of night nests by gorillas in the Bwindi Impenetrable National Park, Uganda. Am. J. Primatol. 2006, 68, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons: New York, NY, USA, 1974. [Google Scholar]
- Fedigan, L.M. Ethical issues faced by field primatologists: Asking the relevant questions. Am. J. Primatol. 2010, 72, 754–771. [Google Scholar] [CrossRef] [PubMed]
- Bezanson, M.; McNamara, A. The what and where of primate field research may be failing primate conservation. Evol. Anthropol. Issues News Rev. 2019, 28, 166–178. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, K.C.; Riley, E.P. Contemporary ethical issues in field primatology. In Ethics in the Field: Contemporary Challenges; MacClancy, J., Fuentes, A., Eds.; Berghahn Books: New York, NY, USA, 2013; pp. 98–107. [Google Scholar]
- Riley, E.P.; Bezanson, M. Ethics of primate fieldwork: Toward an ethically engaged primatology. Annu. Rev. Anthropol. 2018, 47, 493–512. [Google Scholar] [CrossRef]
- Strier, K.B. Are observational field studies of wild primates really noninvasive? In Ethics in the Field: Contemporary Challenges; MacClancy, J., Fuentes, A., Eds.; Berghahn Books: New York, NY, USA, 2013; pp. 67–83. [Google Scholar]
- Mehlman, P.T.; Doran, D.M. Influencing western gorilla nest construction at Mondika Research Center. Int. J. Primatol. 2002, 23, 1257–1285. [Google Scholar] [CrossRef]
- Remis, M. The positional behavior of lowland gorillas (Gorilla gorilla gorilla) in the Dzanga-Sangha Reserve, Central African Republic. Am. J. Primatol. 1993, 30, 344. [Google Scholar]
- Willie, J.; Petre, C.A.; Tagg, N.; Lens, L. Density of herbaceous plants and distribution of western gorillas in different habitat types in south-east Cameroon. Afr. J. Ecol. 2013, 51, 111–121. [Google Scholar] [CrossRef]
- Etiendem, D.N. Tree nesting in Mawambi Hills gorillas. Gorilla J. 2013, 46, 10–12. [Google Scholar]
- Bergl, R.A.; Warren, Y.; Nicholas, A.; Dunn, A.; Imong, I.; Sunderland-Groves, J.L.; Oates, J.F. Remote sensing analysis reveals habitat, dispersal corridors and expanded distribution for the Critically Endangered Cross River gorilla Gorilla gorilla diehli. Oryx 2012, 46, 278–289. [Google Scholar] [CrossRef]
- Imong, I.; Robbins, M.M.; Mundry, R.; Bergl, R.; Kühl, H.S. Informing conservation management about structural versus functional connectivity: A case study of Cross River gorillas. Am. J. Primatol. 2014, 76, 978–988. [Google Scholar] [CrossRef]
- Boivin, N.L.; Zeder, M.A.; Fuller, D.Q.; Crowther, A.; Larson, G.; Erlandson, J.M.; Denham, T.; Petraglia, M.D. Ecological consequences of human niche construction: Examining long-term anthropogenic shaping of global species distributions. Proc. Natl. Acad. Sci. USA 2016, 113, 6388–6396. [Google Scholar] [CrossRef]
- Duvall, C.S. Human settlement ecology and chimpanzee habitat selection in Mali. Landsc. Ecol. 2008, 23, 699–716. [Google Scholar] [CrossRef]
- Malone, N.; Selby, M.; Longo, S. Political-ecological dimensions of silvery gibbon conservation efforts: An endangered ape in (and on) the verge. Int. J. Sociol. 2014, 44, 34–53. [Google Scholar] [CrossRef]
- Tsing, A.L. Friction; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Goldewijk, K.K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef]
- Grayson, D.K. The archaeological record of human impacts on animal populations. J. World Prehist. 2001, 15, 1–68. [Google Scholar] [CrossRef]
- Wright, D.K.; MacEachern, S.; Ambrose, S.H.; Choi, J.; Choi, J.-H.; Lang, C.; Wang, H. Iron Age landscape changes in the Benoué River Valley, Cameroon. Quat. Res. 2019, 92, 323–339. [Google Scholar] [CrossRef]
- De Maret, P.; Clist, B.; Van Neer, W. Résultats des premiéres fouilles dans les abris de Shum Laka et d’Abeke au Nord-Ouest du Cameroun. L’anthropologie 1987, 91, 559–584. [Google Scholar]
- Bostoen, K.; Clist, B.; Doumenge, C.; Grollemund, R.; Hombert, J.M.; Muluwa, J.K.; Maley, J.; Blench, R.; Di Carlo, P.; Good, J.; et al. Middle to late Holocene Paleoclimatic change and the early Bantu expansion in the rain forests of Western Central Africa. Curr. Anthropol. 2015, 56, 367–368. [Google Scholar] [CrossRef]
- Neumann, K.; Bostoen, K.; Höhn, A.; Kahlheber, S.; Ngomanda, A.; Tchiengué, B. First farmers in the Central African rainforest: A view from southern Cameroon. Quat. Int. 2012, 249, 53–62. [Google Scholar] [CrossRef]
- Giresse, P.; Maley, J.; Chepstow-Lusty, A. Understanding the 2500 yr BP rainforest crisis in West and Central Africa in the framework of the Late Holocene: Pluridisciplinary analysis and multi-archive reconstruction. Glob. Planet. Chang. 2020, 192, 103257. [Google Scholar] [CrossRef]
- Röschenthaler, U.M. Translocal cultures: The slave trade and cultural transfer in the Cross River region. Soc. Anthropol. 2006, 14, 71–91. [Google Scholar] [CrossRef]
- Michels, S. Imagined Power Contested: Germans and Africans in the Upper Cross River Area of Cameroon, c. 1887–c. 1915; ’Lit Verlag: Münster, Germany, 2004; Volume 2. [Google Scholar]
- Röschenthaler, U.M. Purchasing Culture: The Dissemination of Associations in the Cross River Region of Cameroon and Nigeria; Africa World Press: Trenton, NJ, USA, 2011. [Google Scholar]
- Biwolé, A.B.; Morin-Rivat, J.; Fayolle, A.; Bitondo, D.; Dedry, L.; Dainou, K.; Hardy, O.J.; Doucet, J.L. New data on the recent history of the littoral forests of southern Cameroon: An insight into the role of historical human disturbances on the current forest composition. Plant Ecol. Evol. 2015, 148, 19–28. [Google Scholar] [CrossRef]
- Garcin, Y.; Deschamps, P.; Ménot, G.; De Saulieu, G.; Schefuß, E.; Sebag, D.; Dupont, L.M.; Oslisly, R.; Brademann, B.; Mbusnum, K.G.; et al. Early anthropogenic impact on Western Central African rainforests 2600 y ago. Proc. Natl. Acad. Sci. USA 2018, 115, 3261–3266. [Google Scholar] [CrossRef]
- Morin-Rivat, J.; Fayolle, A.; Favier, C.; Bremond, L.; Gourlet-Fleury, S.; Bayol, N.; Lejeune, P.; Beeckman, H.; Doucet, J.L. Present-day central African forest is a legacy of the 19th century human history. eLife 2017, 6, e20343. [Google Scholar] [CrossRef]
- Sharwood-Smith, B.E.; Cantle, L.L. An Assessment Report on the Anyang and Manta Tribes of Mamfe Division, Cameroons Province; Buea National Archive AF4; Buea National Archive: Buea, Cameroon, 1924. [Google Scholar]
- Courade, G.; Ngo-Botok, V.; Ngum-Kwe, F. Village Dictionary of Manyu Division; Office de la Recherche Scientifique et Technique Outre-mer: Yaounde, Cameroun, 1973. [Google Scholar]
- Hull, G.; Cooper, J.E. Catalogue of preserved gorilla materials. In Gorilla Pathology and Health: With a Catalogue of Preserved Materials; Hull, G., Cooper, J.E., Eds.; Elsevier Inc.: London, UK, 2017; pp. 379–602. [Google Scholar]
- Blake, S. The Ecology of Forest Elephant Distribution and Its Implications for Conservation. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 2002. [Google Scholar]
- Bekhuis, P.D.; De Jong, C.B.; Prins, H.H. Diet selection and density estimates of forest buffalo in Campo-Ma’an National Park, Cameroon. Afr. J. Ecol. 2008, 46, 668–675. [Google Scholar] [CrossRef]
- Eniang, E.A.; Ebin, C.O.; Nchor, A.A.; Agba, G.I.; Gideon, I.K.; Amori, G.; Luiselli, L. Distribution and status of the African forest buffalo Syncerus caffer nanus in south-eastern Nigeria. Oryx 2017, 51, 538–541. [Google Scholar] [CrossRef]
- Head, J.S.; Boesch, C.; Makaga, L.; Robbins, M.M. Sympatric chimpanzees (Pan troglodytes troglodytes) and gorillas (Gorilla gorilla gorilla) in Loango National Park, Gabon: Dietary composition, seasonality, and intersite comparisons. Int. J. Primatol. 2011, 32, 755–775. [Google Scholar] [CrossRef]
- Bergl, R.A. Conservation Biology of the Cross River gorilla (Gorilla gorilla diehli). Ph.D. Thesis, The City University of New York, New York, NY, USA, 2006. [Google Scholar]
- Nuesiri, E.O. Conservation and resource use in Cross River—Takamanda Forest Complex of Cameroon and Nigeria; Fauna and Flora International: Cambridge, UK, 2003. [Google Scholar]
- Bitondo, D.; Schmidt-Soltau, K.; Ntep, J.; Onya, H.; Wadt Zela, F.F.; Frida, N.A.; Tambe, J.B.; Ebot, R. The Mamfe—Akwaya Road Project: An Environmental Impact Assessment; German Development Cooperation: Buea, Cameroon, 1999. [Google Scholar]
- Oates, J.F. Myth and Reality in the Rain Forest: How Conservation Strategies Are Failing in West Africa; University of California Press: Berkeley, CA, USA, 1999. [Google Scholar]
- Laird, S.A.; Awung, G.L.; Lysinge, R.J. Cocoa farms in the Mount Cameroon region: Biological and cultural diversity in local livelihoods. Biodivers. Conserv. 2007, 16, 2401–2427. [Google Scholar] [CrossRef]
- Klarer, A.J. The Evolution and Expansion of Cacao Farming in South West Cameroon and Its Effects on Local Livelihoods. Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 2014. [Google Scholar]
- Dada, L.A. The African Export Industry: What Happened and How Can It Be Revived. Case Study on the Cameroonian Cocoa Sector; Agricultural Management, Marketing and Finance, Working Paper; Food and Agricultural Organisation (FAO): Geneva, Switzerland, 2007. [Google Scholar]
- Abei, L. An Analysis of the Competitive Performance of the Cameroonian Cocoa Industry. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2017. [Google Scholar]
- Business in Cameroon. Available online: https://www.businessincameroon.com/agriculture/0204-9006-cameroon-cutscocoa-production-forecasts-for-2020-from-600-000-to-285-000-tons (accessed on 29 April 2019).
- Cameroon Concord. Available online: http://www.cameroonconcordnews.com/cameroon-was-never-one-theagbaw-ebai-debate/ (accessed on 17 July 2019).
- Mukete, N.; Li, Z.; Beckline, M.; Patricia, B. Cocoa production in Cameroon: A socioeconomic and technical efficiency perspective. Int. J. Agric. Econ. 2018, 3, 1–8. [Google Scholar] [CrossRef]
- Asase, A.; Tetteh, D.A. The role of complex agroforestry systems in the conservation of forest tree diversity and structure in southeastern Ghana. Agrofor. Syst. 2010, 79, 355–368. [Google Scholar] [CrossRef]
- Degrande, A.; Schreckenberg, K.; Mbosso, C.; Anegbeh, P.; Okafor, V.; Kanmegne, J. Farmers’ fruit tree-growing strategies in the humid forest zone of Cameroon and Nigeria. Agrofor. Syst. 2006, 67, 159–175. [Google Scholar] [CrossRef]
- Herzog, F. Multipurpose shade trees in coffee and cocoa plantations in Côte d’Ivoire. Agrofor. Syst. 1994, 27, 259–267. [Google Scholar] [CrossRef]
- Jagoret, P.; Michel-Dounias, I.; Malézieux, E. Long-term dynamics of cocoa agroforests: A case study in central Cameroon. Agrofor. Syst. 2011, 81, 267–278. [Google Scholar] [CrossRef]
- Ruf, F.; Schroth, G. Chocolate forests and monocultures: A historical review of cocoa growing and its conflicting role in tropical deforestation and forest conservation. In Agroforestry and Biodiversity Conservation in Tropical Landscapes; Schroth, G., Da Fonseca, G.A.B., Harvey, C.A., Gascon, C., Vasconcelos, H.L., Izac, A.M.N., Eds.; Island Press: Washington, DC, USA, 1995; pp. 107–133. [Google Scholar]
- Voigt, M.; Wich, S.A.; Ancrenaz, M.; Meijaard, E.; Abram, N.; Banes, G.L.; Campbell-Smith, G.; d’Arcy, L.J.; Delgado, R.A.; Erman, A.; et al. Global demand for natural resources eliminated more than 100,000 Bornean orangutans. Curr. Biol. 2018, 28, 761–769. [Google Scholar] [CrossRef]
- Human Rights Watch. Available online: https://www.hrw.org/news/2019/05/23/cameroon-promised-investigation-shouldbe-independent (accessed on 3 June 2019).
- Foreign Policy. Available online: https://foreignpolicy.com/2019/05/13/cameroons-separatistmovement-is-going-international-ambazonia-military-forces-amf-anglophonecrisis/ (accessed on 3 June 2019).
- United Nations Office for the Coordination of Humanitarian Affairs. Available online: https://www.humanitarianresponse.info/sites/www.humanitarianresponse.info/files/documents/files/sitrep_nwsw_nov_2020_vf.pdf (accessed on 14 January 2020).
- Gaynor, K.M.; Fiorella, K.J.; Gregory, G.H.; Kurz, D.J.; Seto, K.L.; Withey, L.S.; Brashares, J.S. War and wildlife: Linking armed conflict to conservation. Front Ecol Environ. 2016, 14, 533–542. [Google Scholar] [CrossRef]
- Plumptre, A.J.; Nixon, S.; Critchlow, R.; Vieilledent, G.; Kirkby, A.; Williamson, E.A.; Hall, J.; Kujirakwinja, D. Status of Grauer’s Gorilla and Chimpanzee in Eastern Democratic Republic of Congo: Historical and Current Distribution and Abundance; Wildlife Conservation Society, Fauna & Flora International and Institut Congolais pour la Conservation de la Nature: New York, NY, USA, 2015. [Google Scholar]
- Shambaugh, J.; Oglethorpe, J.; Ham, R. The Trampled Grass: Mitigating the Impacts of Armed Conflict on the Environment; Biodiversity Support Program: Washington, DC, USA, 2001. [Google Scholar]
- Hockings, K.J.; McLennan, M.R.; Carvalho, S.; Ancrenaz, M.; Bobe, R.; Byrne, R.W.; Dunbar, R.I.; Matsuzawa, T.; McGrew, W.C.; Williamson, E.A.; et al. Apes in the Anthropocene: Flexibility and survival. Trends Ecol. Evol. 2015, 30, 215–222. [Google Scholar] [CrossRef] [PubMed]
- McLennan, M.R.; Spagnoletti, N.; Hockings, K.J. The implications of primate behavioral flexibility for sustainable human–primate coexistence in anthropogenic habitats. Int. J. Primatol. 2017, 38, 105–121. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Class | Description | Indicator Plants |
|---|---|---|
| Old secondary forest | Forest recovering from disturbance; trees are smaller and canopy height is lower than in primary forest, either from human modification or natural disturbances. | Elaeis guineensis, Musanga cecropioides, Piptadenianstrum africanum, Tabernaemontana crassa, Pycnanthus angolensis |
| Lowland forest < 500 m a.s.l. | Mature forest with tall and large trees; a high canopy and a distinctive middle story. | Afrostyrax kameroensis, Desbordesia spp., Carapa procera, Parkia bicolor, Strombosia grandifolia |
| Lowland ridge forest | Mature forest growing on the sides of finger-like ridges; at some places the forest is interspersed with rocky cliffs. | Allenblackia floribunda, Tapura africana, Chrysophyllum spp., Lophira alata, Pseudospondias microcarpa |
| Mid-elevation forest 500–800 m a.s.l. | Mature forest with a lower canopy and denser understory. Diversity is reduced but also contains elements of both lowland and montane forests. | Homalium spp., Syzygium spp., Xylopia staudtii, Carpolobia lutea, Rinorea dentata, Dichapetalum spp. |
| Montane forest 800–1500 m a.s.l. | Mature forest with a further reduction in canopy height which is often disjunct; a decrease in species richness. | Dactyladeina staudii, Dracenea arborea, Eugenia spp. |
| Category | Description | |
|---|---|---|
| Density of understory | Very dense | (Stems < 50 cm apart) |
| Dense | (Stems < 1 m apart) | |
| Sparse | (Stems > 1 m apart) | |
| Slope | Precipitous | (Over 50%, >26.6°) |
| Steep | (10–50%, 5.7–26.6°) | |
| Gentle | (0–10%, 0–5.7°) | |
| Flat | ||
| Canopy cover | Very closed | (>75%) |
| Closed | (51–75%) | |
| Open | (26–50%) | |
| Very open | (0–25%) |
| Age Category | Approximate Age | Definition |
|---|---|---|
| New | Less than two days | Nest has green unwilted leaves. Presence of dung and urine in or around the nest. |
| Recent | Less than one week | Nest has wilted green leaves. |
| Old and intact | Between one week to one month | Nest remains intact but leaves have turned brown. |
| Decomposing | More than one month | Nest consists of interwoven stems and branches that lack leaves. |
| Parameter | Estimate | Standard Error | Z-Score | p-Value | 95% Confidence Interval | |
|---|---|---|---|---|---|---|
| Lower | Upper | |||||
| Canopy cover | −0.357 | 0.253 | −1.412 | 0.158 | −0.853 | 0.139 |
| Elevation | −0.679 | 0.189 | −3.589 | <0.001 | −1.049 | −0.308 |
| Degree of slope | 1.029 | 0.387 | 2.660 | 0.008 | 0.271 | 1.788 |
| Vegetation Class | Plots (n) | Trees DBH > 10 cm (n) | Tree Density (Trees/ha) | Tree Species (n) | Understory Species (n) | Stem Density (Stems/m2) | Mean Height (m) | Mean DBH (cm) | Stand Basal Area (m2/ha) | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |||||||
| Old secondary | 9 | 472 | 520 | 119 | 129 | 24 | 11 | 16 | 8 | 23 | 17 | 34 |
| Lowland | 42 | 2083 | 500 | 203 | 212 | 29 | 18 | 16 | 7 | 23 | 15 | 29 |
| Lowland ridge | 7 | 365 | 520 | 116 | 80 | 21 | 9 | 17 | 8 | 24 | 19 | 39 |
| Mid-elevation | 53 | 2824 | 530 | 215 | 226 | 22 | 20 | 16 | 8 | 23 | 16 | 33 |
| Montane | 9 | 517 | 570 | 120 | 113 | 26 | 20 | 14 | 7 | 22 | 15 | 32 |
| Ape nesting range | ||||||||||||
| Chimpanzee | 23 | 1303 | 570 | 164 | 143 | 15 | 6 | 16 | 10 | 24 | 17 | 39 |
| Gorilla | 25 | 1227 | 490 | 183 | 171 | 24 | 20 | 16 | 8 | 23 | 16 | 29 |
| Genera | Form | # of Times Plant Used in Nest | Mean # of Stems in Nest | Median Proportion of Stems in Nest | # of Times Plant Was Available in Environment | Mean # of Stems in Environment | Median Proportion of Stems in Environment | Median of the Difference | p-Value | r-Value | Preference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Aframomum** | Herb | 406 | 13 | 0.50 | 644 | 22 | 0.28 | −0.1146 | * 0.016 | 0.41 | Preferred |
| Acanthaceae | Herb | 104 | 5 | 0.35 | 582 | 14 | 0.24 | 0.1163 | * 0.001 | 0.49 | Avoided |
| Urera ** | Liana | 87 | 5 | 0.14 | 150 | 7 | 0.03 | −0.0402 | 0.171 | - | Neutral |
| Palisota ** | Herb | 58 | 2 | 0.03 | 172 | 9 | 0.05 | 0.0213 | 0.317 | - | Neutral |
| Piper ** | Vine | 54 | 6 | 0.02 | 60 | 3 | 0.02 | 0.0206 | 0.198 | - | Neutral |
| Marantaceae | Herb | 53 | 13 | 0.08 | 92 | 9 | 0.05 | 0.0435 | 0.600 | - | Neutral |
| Alchornea | Shrub | 46 | 4 | 0.13 | 71 | 5 | 0.07 | −0.0563 | 0.053 | - | Neutral |
| Leea ** | Shrub | 40 | 4 | 0.05 | 34 | 2 | 0.02 | −0.0215 | 0.136 | - | Neutral |
| Protomegabaria ** | Tree | 39 | 6 | 0.13 | 20 | 7 | 0.02 | −0.1339 | * 0.025 | 0.79 | Preferred |
| Penianthus ** | Shrub | 38 | 3 | 0.03 | 108 | 4 | 0.05 | 0.0339 | 0.673 | - | Neutral |
| Thaumatococcus ** | Herb | 30 | 30 | 0.75 | 37 | 37 | 0.79 | 0.0372 | 0.317 | - | Neutral |
| Landolphia ** | Liana | 28 | 2 | 0.04 | 49 | 2 | 0.02 | 0.0222 | 0.248 | - | Neutral |
| Sorindeia ** | Shrub | 19 | 1 | 0.02 | 71 | 2 | 0.03 | 0.0233 | 0.512 | - | Neutral |
| Rinorea ** | Shrub | 18 | 2 | 0.03 | 56 | 2 | 0.03 | 0.0241 | 0.347 | Neutral | |
| Pavetta ** | Shrub | 17 | 2 | 0.01 | 59 | 2 | 0.03 | 0.0270 | 0.163 | - | Neutral |
| Genera | Form | # of Times Plant Used in Nest | Mean # of Stems in Nest | Median Proportion of Stems in Nest | # of Times Plant Was Available in Environment | Mean # of Stems in Environment | Median Proportion of Stems in Environment | Median of the Difference | p-Value | r-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Aframomum | Herb | 406 | 13 | 0.50 | 644 | 22 | 0.28 | −0.1146 | 0.016 | 0.41 |
| Protomegabaria | Tree | 39 | 6 | 0.13 | 20 | 7 | 0.02 | −0.1339 | 0.025 | 0.79 |
| Tiliaceae | Vine | 13 | 2 | 0.07 | 6 | 2 | 0.01 | −0.0606 | 0.028 | 0.69 |
| Tree Species | Observed # of Trees Used for Nesting (1) | Cumulative % | Proportion of (1) to All Trees Used for Nesting | # of Trees Sampled in Botanical Plots (2) | Proportion of (2) to All Trees Sampled | Expected # of Trees Used for Nesting | Manly’s α | Category by Significant Preference | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Sorindeia sp. 2 ** | 10 | 17 | 0.172 | 63 | 0.033 | 1.9 | 0.019 | Preferred | <0.001 |
| Pycnanthus angolensis ** | 6 | 28 | 0.103 | 49 | 0.026 | 1.5 | 0.015 | Preferred | 0.004 |
| Microdesmis sp. | 3 | 33 | 0.052 | 24 | 0.013 | 0.7 | 0.015 | Preferred | 0.04 |
| Angylocalyx pynaertii | 2 | 36 | 0.034 | 44 | 0.023 | 1.3 | 0.005 | Neutral | NS |
| Chrysophyllum sp. ** | 2 | 40 | 0.034 | 12 | 0.006 | 0.4 | 0.020 | Preferred | 0.048 |
| Cola lateritia | 2 | 43 | 0.034 | 8 | 0.004 | 0.2 | 0.030 | Preferred | 0.023 |
| Maesobotrya sp. | 2 | 47 | 0.034 | 18 | 0.009 | 0.5 | 0.013 | Preferred | NS |
| Napoleonaea vogelii ** | 2 | 50 | 0.034 | 24 | 0.013 | 0.7 | 0.010 | Preferred | NS |
| Protomegabaria sp. ** | 2 | 53 | 0.034 | 18 | 0.009 | 0.5 | 0.013 | Preferred | NS |
| Pseudospondias sp. ** | 2 | 57 | 0.034 | 29 | 0.015 | 0.9 | 0.008 | Preferred | NS |
| Tabernaemontana crassa | 2 | 60 | 0.034 | 49 | 0.026 | 1.5 | 0.005 | Neutral | NS |
| Trema sp. | 2 | 64 | 0.034 | 1 | 0.001 | 0.0 | 0.240 | Preferred | 0.002 |
| Trichilia rubescens ** | 2 | 67 | 0.034 | 46 | 0.024 | 1.4 | 0.005 | Neutral | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wade, A.H.; Malone, N. Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon. Diversity 2021, 13, 175. https://doi.org/10.3390/d13040175
Wade AH, Malone N. Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon. Diversity. 2021; 13(4):175. https://doi.org/10.3390/d13040175
Chicago/Turabian StyleWade, Alison H., and Nicholas Malone. 2021. "Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon" Diversity 13, no. 4: 175. https://doi.org/10.3390/d13040175
APA StyleWade, A. H., & Malone, N. (2021). Ecological, Historical, Economic, and Political Factors Shaping the Human–Gorilla Interface in the Mone-Oku Forest, Cameroon. Diversity, 13(4), 175. https://doi.org/10.3390/d13040175