
Updated Distribution of the Mysid Antromysis cenotensis (Crustacea: Peracarida), a Protected Key Species in Yucatan Peninsula Cenotes



Abstract

1. Introduction
2. Materials and Methods
2.1. Bibliographic Review
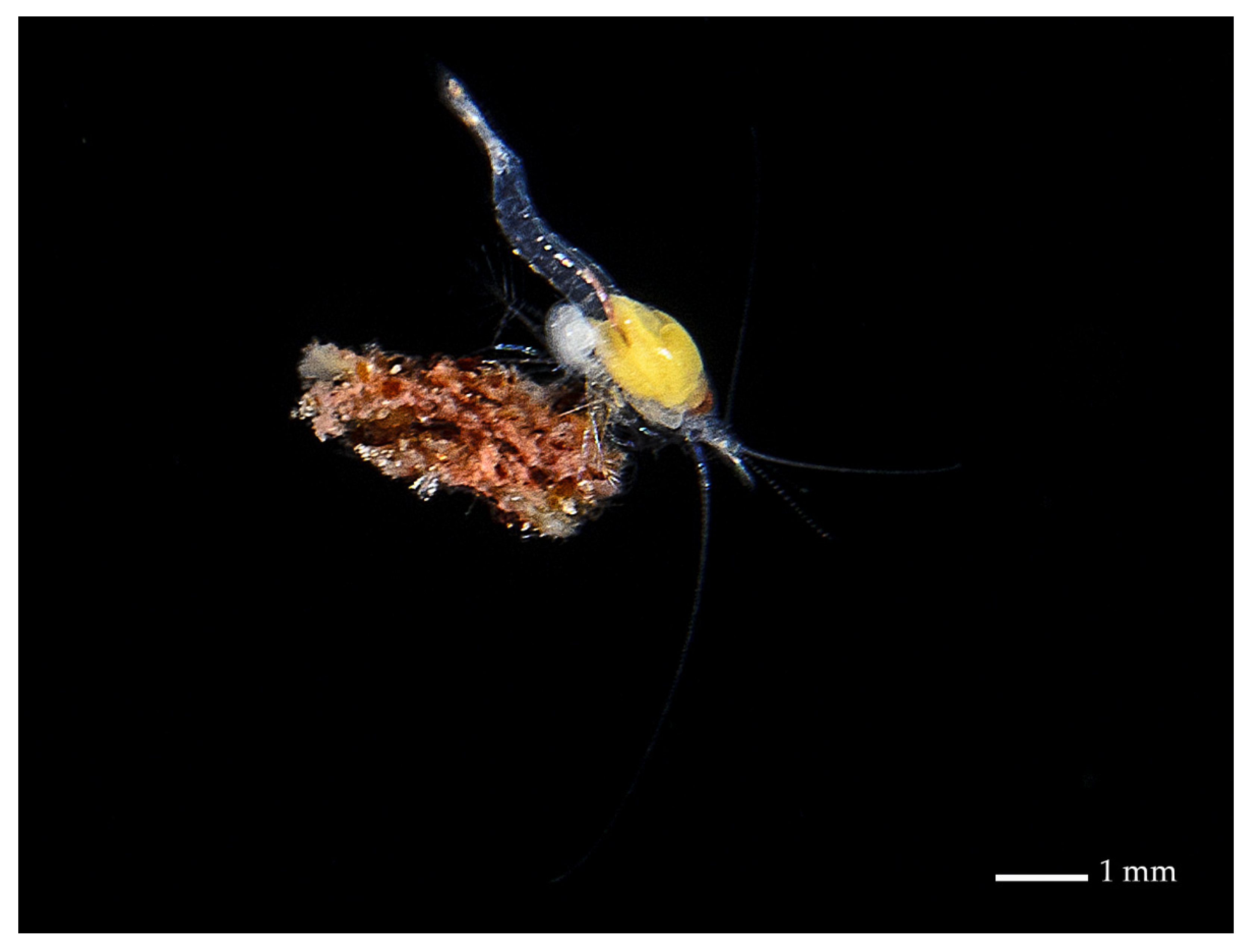
2.2. Specimen Collection
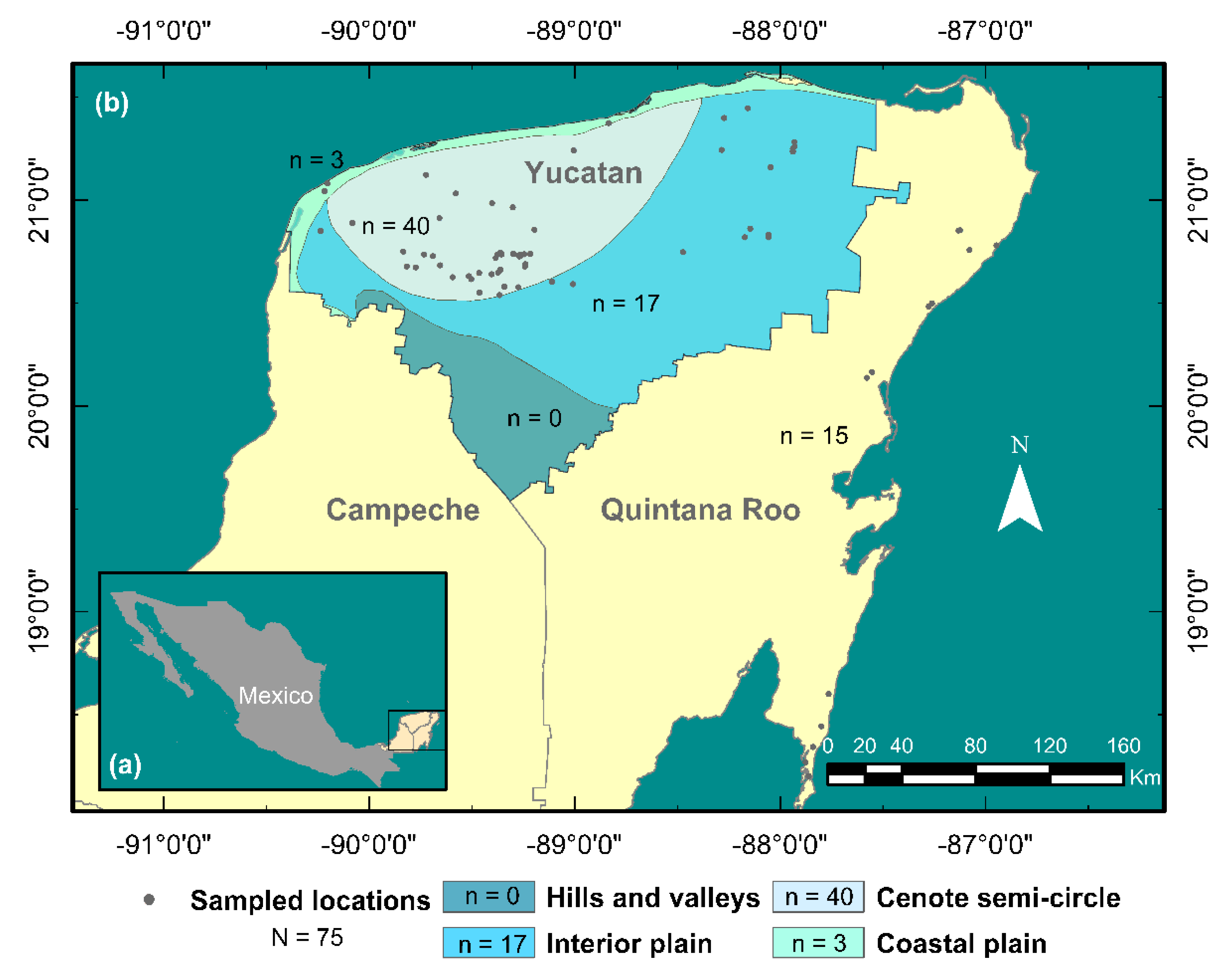
2.3. Map Ellaboration
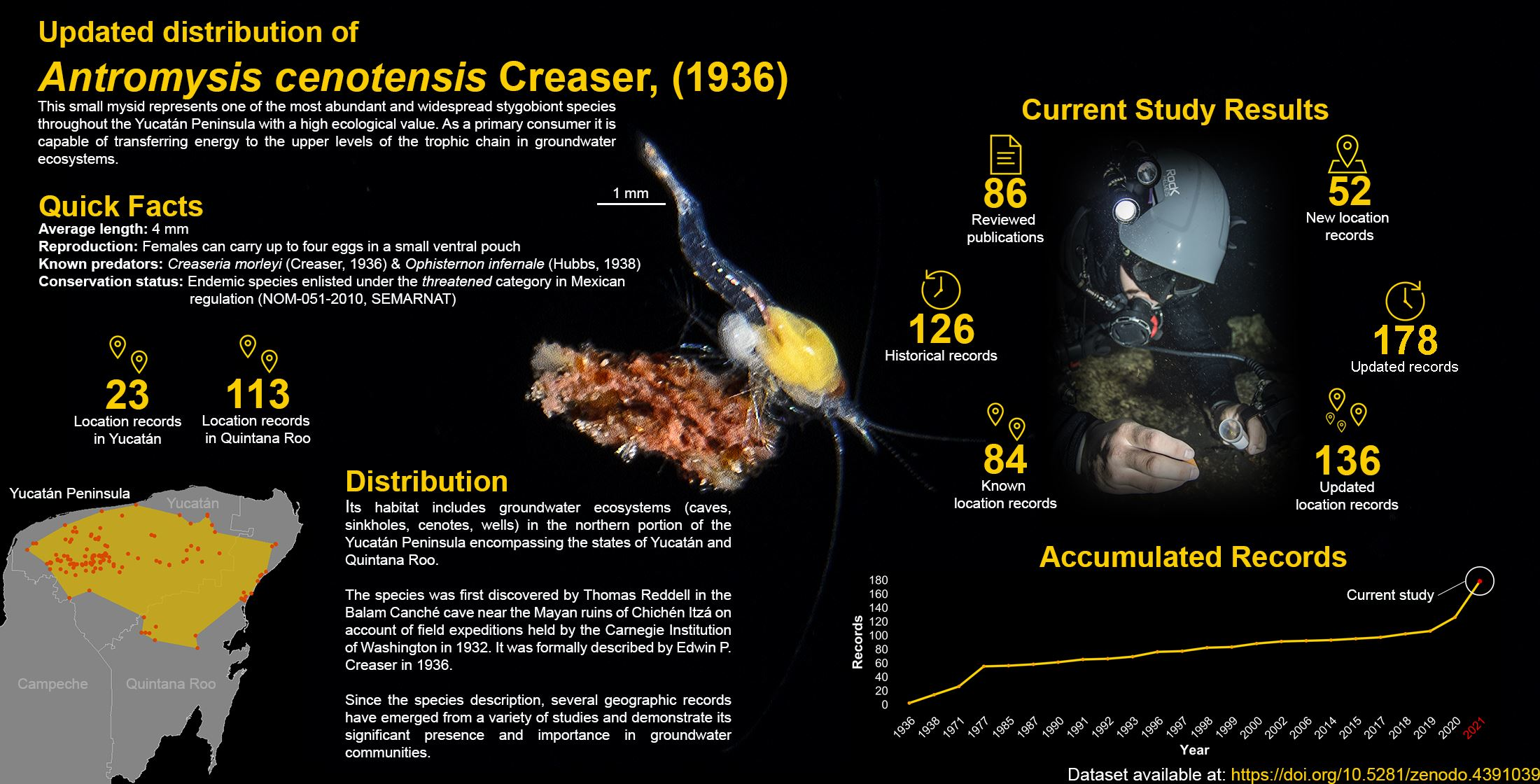
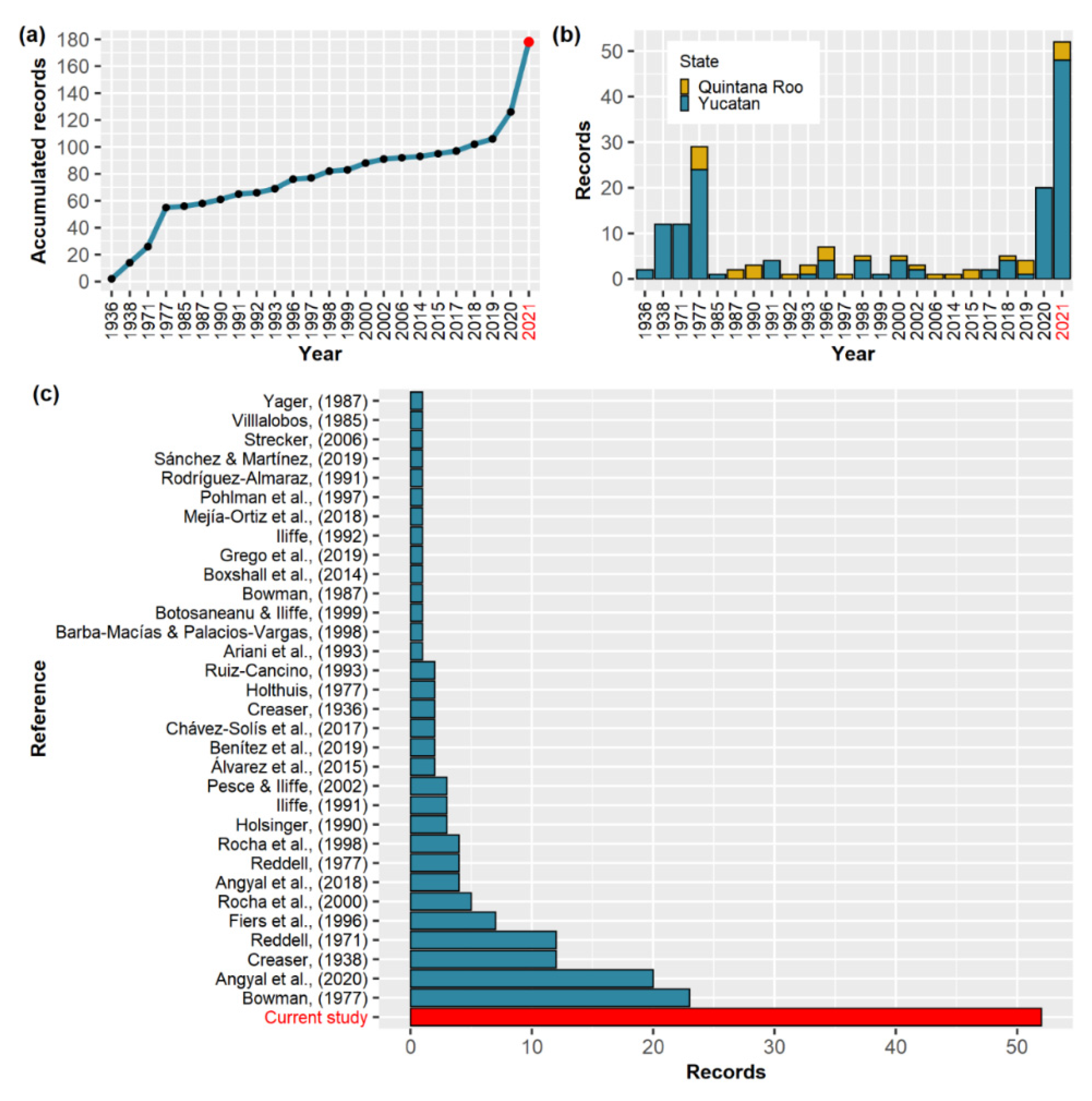
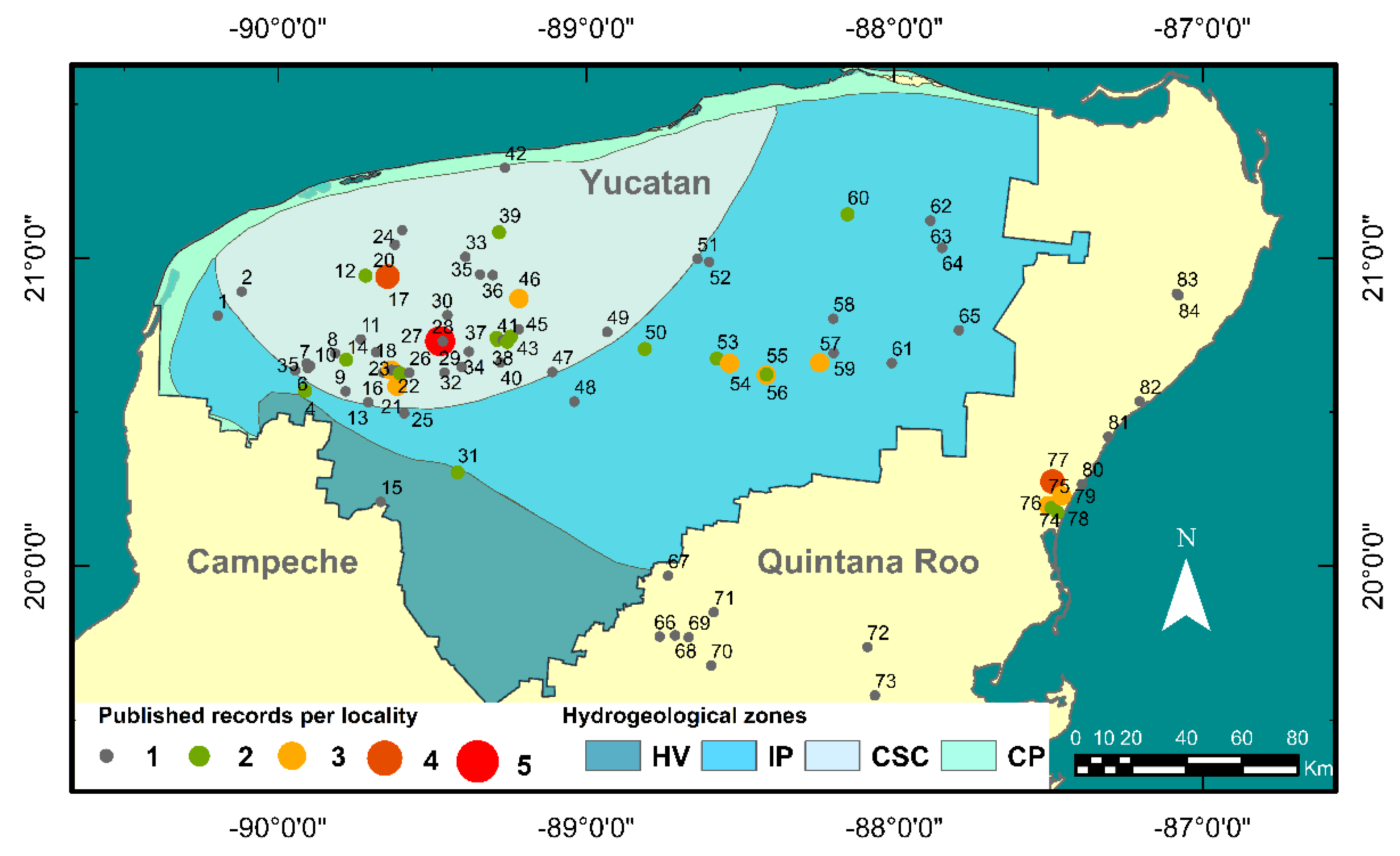
3. Results
3.1. An Account of A. cenotensis Historical Distribution Records
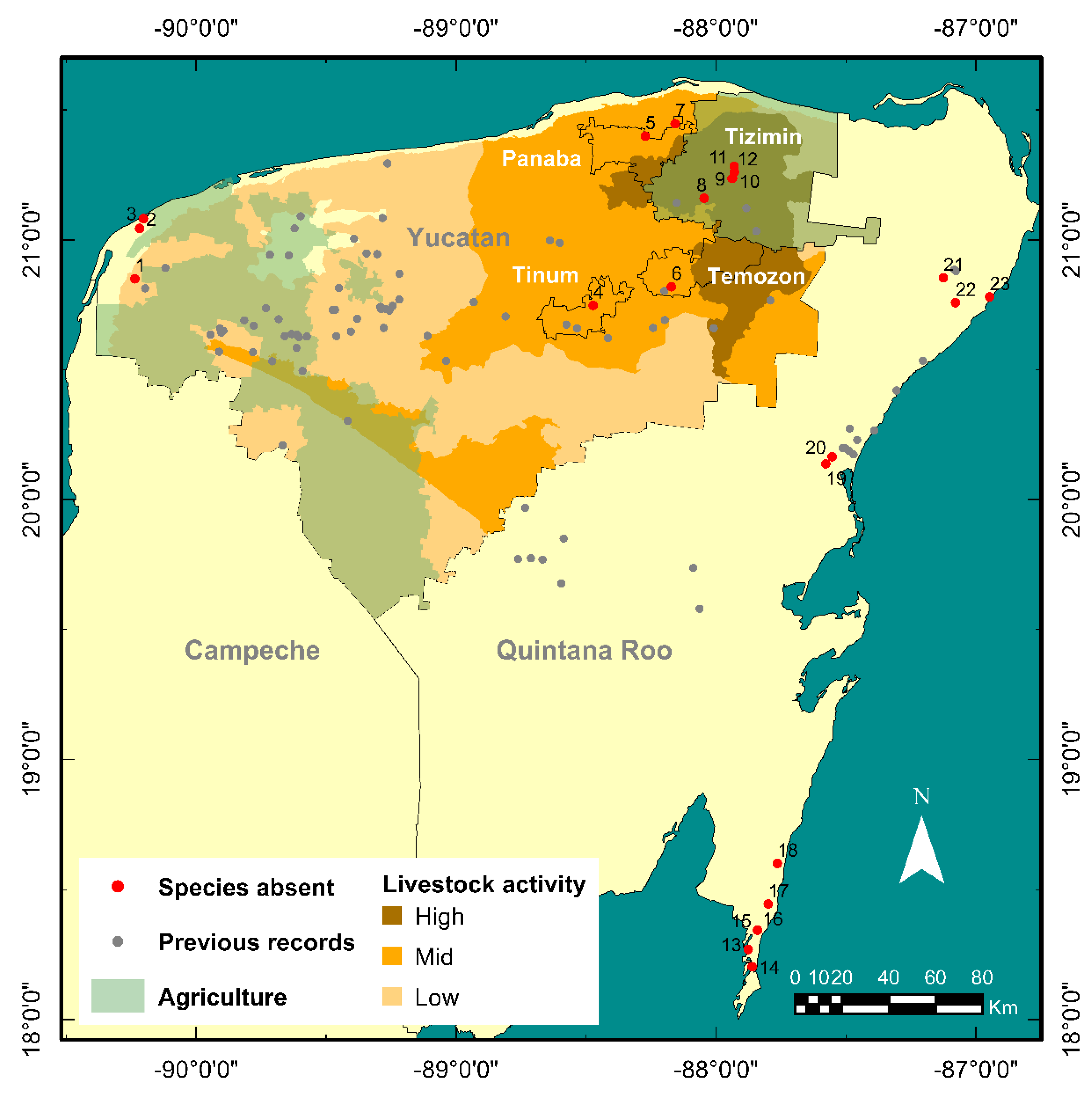
3.2. New Records
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mees, J.; Meland, K. World List of Lophogastrida, Stygiomysida and Mysida. Available online: http://www.marinespecies.org/mysidacea (accessed on 24 November 2020).
- Porter, M.L.; Meland, K.; Price, W. Global diversity of mysids (Crustacea-Mysida) in freshwater. Hydrobiologia 2008, 595, 213–218. [Google Scholar] [CrossRef]
- Meland, K.; Mees, J.; Porter, M.; Wittmann, K.J. Taxonomic Review of the Orders Mysida and Stygiomysida (Crustacea, Peracarida). PLoS ONE 2015, 10, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Meland, K.; Willassen, E. The disunity of “Mysidacea” (Crustacea). Mol. Phylogenet. Evol. 2007, 44, 1083–1104. [Google Scholar] [CrossRef] [PubMed]
- Rudstam, L.G.; Johansson, O.E. Advances in the ecology of freshwater mysids. Aquat. Biol. 2009, 5, 246–248. [Google Scholar] [CrossRef]
- Coma, R.; Carola, M.; Riera, T.; Zabala, M. Horizontal transfer of matter by a cave-dwelling mysid. Mar. Ecol. 1997, 18, 211–226. [Google Scholar] [CrossRef]
- Takahashi, K. Feeding ecology of mysids in freshwater and coastal marine habitats: A review. Bull. Plankt. Soc. Jpn. 2004, 51, 46–72. [Google Scholar]
- Rastorgueff, P.A.; Harmelin-Vivien, M.; Richard, P.; Chevaldonné, P. Feeding strategies and resource partitioning mitigate the effects of oligotrophy for marine cave mysids. Mar. Ecol. Prog. Ser. 2011, 440, 163–176. [Google Scholar] [CrossRef]
- Viherluoto, M.; Kuosa, H.; Flinkman, J.; Viitasalo, M. Food utilisation of pelagic mysids, Mysis mixta and M. relicta, during their growing season in the northern Baltic Sea. Mar. Biol. 2000, 136, 553–559. [Google Scholar] [CrossRef]
- Punchihewa, N.N.; Krishnarajah, S.R. Trophic Position of Two Mysid Species (Crustacea: Mysidacea) in an Estuarine Ecosystem in Auckland, New Zealand, Using Stable Isotopic Analysis. Am. J. Mar. Sci. 2013, 1, 22–27. [Google Scholar] [CrossRef]
- González-Ortegón, E.; Drake, P. Effects of freshwater inputs on the lower trophic levels of a temperate estuary: Physical, physiological or trophic forcing? Aquat. Sci. 2012, 74, 455–469. [Google Scholar] [CrossRef]
- Mauchline, J. The Biology of Mysids and Euphausiids. In Advances in Marine Biology; Blaxter, J.H.S., Russel, F.S., Yonge, M., Eds.; Academic Press Inc.: Cambridge, MA, USA, 1980; Volume 18, p. 681. ISBN 0065-2881. [Google Scholar]
- Roast, S.D.; Thompson, R.S.; Widdows, J.; Jones, M.B. Mysids and environmental monitoring: A case for their use in estuaries. Mar. Freshw. Res. 1998, 49, 827–832. [Google Scholar] [CrossRef]
- Verslycke, T.A.; Fockedey, N.; McKenney, C.L.; Roast, S.D.; Jones, M.B.; Mees, J.; Janssen, C.R. Mysid crustaceans as potential test organisms for the evaluation of environmental endocrine disruption: A review. Environ. Toxicol. Chem. 2004, 23, 1219–1234. [Google Scholar] [CrossRef]
- Anderson, B.; Phillips, B. Saltwater Toxicity Tests. In Marine Ecotoxicology; Elsevier: Amsterdam, The Netherlands, 2016; pp. 167–197. [Google Scholar]
- Sket, B. The nature of biodiversity in hypogean waters and how it is endangered. Biodivers. Conserv. 1999. [Google Scholar] [CrossRef]
- The National Speleological Society Survey and Cartography of The Underwater Caves of Quintana Roo Mexico. Available online: https://caves.org/project/qrss/qrss.htm (accessed on 24 November 2020).
- Creaser, E.P. Crustaceans from Yucatan. In The Cenotes of Yucatan. A Zoological and Hydrographic Survey; Pearse, A.S., Creaser, E.P., Hall, F.., Eds.; Carnegie Institution of Washington: Washington, DC, USA, 1936; pp. 117–132. [Google Scholar]
- Reddell, J.R. A Preliminary Survey of the Caves of the Yucatan Peninsula; Reddell, J.R., Ed.; The Speleo Press: Austin, TX, USA, 1977; Volume 6. [Google Scholar]
- Iliffe, T.M. An annotated list of the troglobitic anchialine and freshwater fauna of Quintana Roo. In Diversidad Biológica en la Reserva de la Biosfera de Sian Ka’an, Quintana Roo, México; Navarro, D., Suárez-morales, E., Eds.; CIQRO/SEDESOL: Quintana Roo, México, 1992; Volume 2, pp. 196–215. ISBN 9789686780055. [Google Scholar]
- Pohlman, J.; Iliffe, T.; Cifuentes, L. A stable isotope study of organic cycling and the ecology of an anchialine cave ecosystem. Mar. Ecol. Prog. Ser. 1997, 155, 17–27. [Google Scholar] [CrossRef]
- CONAGUA Programa Hídrico Regional 2014-2018 de la Región HidrológicoAdministrativa XII Península de Yucatán; CONAGUA: Ciudad de México, México, 2015; 140p.
- Bauer-Gottwein, P.; Gondwe, B.R.N.; Charvet, G.; Marín, L.E.; Rebolledo-Vieyra, M.; Merediz-Alonso, G. Review: The Yucatán Peninsula karst aquifer, Mexico. Hydrogeol. J. 2011, 19, 507524. [Google Scholar] [CrossRef]
- INEGI. Estudio Hidrológico del Estado de Yucatán; INEGI: Aguascalientes, Mexico, 2002; ISBN 9788578110796. [Google Scholar]
- Schmitter-Soto, J.J.; Comín, F.A.; Escobar-Briones, E.; Herrera-Silveira, J.; Alcocer, J.; Suárez-Morales, E.; Elías-Gutiérrez, M.; Díaz-Arce, V.; Marín, L.E.; Steinich, B. Hydrogeochemical and biological characteristics of cenotes in the Yucatan Peninsula (SE Mexico). Hydrobiologia 2002, 467, 215–228. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gotzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: Explanation and elaboration. BMJ 2009, 339, b2700. [Google Scholar] [CrossRef]
- NMNH Smithsonian National Museum of Natural History Invertebrate Zoology Collections. Available online: https://collections.nmnh.si.edu/search/iz/ (accessed on 5 February 2021).
- UNAM. Colección Nacional de Crustáceos del Instituto de Biología, Universidad Nacional Autónoma de México. Available online: http://www.ibiologia.unam.mx/colecciones/crustaceos/centro_crusta.htm (accessed on 5 February 2021).
- Biodiversity Heritage Library. Available online: https://www.biodiversitylibrary.org/ (accessed on 5 February 2021).
- HathiTrust Digital Library. Available online: https://www.hathitrust.org/ (accessed on 5 February 2021).
- Scopus. Available online: https://www.scopus.com/home.uri (accessed on 5 February 2021).
- Web of Science Master Journal List—Search. Available online: https://mjl.clarivate.com/search-results (accessed on 5 February 2021).
- Google Scholar. Available online: https://scholar.google.com/ (accessed on 5 February 2021).
- Ocean Biodiversity Information System. Available online: https://obis.org/ (accessed on 25 February 2021).
- iNaturalist. Available online: https://www.inaturalist.org/ (accessed on 5 February 2021).
- GBIF. Available online: https://www.gbif.org/ (accessed on 5 February 2021).
- Bowman, T.E.; Mckenzie, H.; Murphy, S. A review of the genus Antromysis (Crustacea: Mysidacea), including new species from Jamaica and Oaxaca, Mexico, and a redescription and new records for A. cenotensis. In Studies on the Caves and Cave Fauna of the Yucatan Peninsula; Reddell, J.R., Ed.; The Speleo Press: Austin, TX, USA, 1977; Volume 6, pp. 27–38. [Google Scholar]
- Chapman, A.D. Current Best Practices for Generalizing Sensitive Species Occurrence Data. Copenhagen GBIF Secr. 2020. [Google Scholar] [CrossRef]
- CONABIO Geoportal del Sistema Nacional de Información sobre Biodiversidad. Available online: http://www.conabio.gob.mx/informacion/gis/ (accessed on 20 February 2021).
- Bitacora Ambiental del Programa de Ordenamiento Ecológico Territorial del Estado de Yucatán. Available online: https://bitacoraordenamiento.yucatan.gob.mx/index.php (accessed on 10 December 2020).
- Angyal, D.; Chávez-Solís, E.M.; Liévano-Beltrán, L.A.; Magaña, B.; Simões, N.; Mascaró, M. New distribution records of subterranean crustaceans from cenotes in Yucatan (Mexico). Zookeys 2020, 911, 21–49. [Google Scholar] [CrossRef]
- Bowman, T.E. Bahalana mayana, a new troglobitic cirolanid isopod from Cozumel Island and the Yucatan Peninsula, Mexico. Proc. Biol. Soc. Wash. 1987, 100, 659–663. [Google Scholar]
- Yager, J. Speleonectes tulumensis, n. sp. (Crustacea, Remipedia) from two anchialine cenotes of the Yucatan Peninsula, Mexico. Stygologia 1987, 3, 160–166. [Google Scholar]
- Holsinger, J. Tuluweckelia cernua, a new genus and species of stygobiont amphipod crustaceans (Hadzidae) from anchialine caves on the Yucatan peninsula in Mexico. Beaufortia 1990, 41, 97–107. [Google Scholar]
- Ruiz-Cancino, G.; Mejía-Ortíz, L.M.; Lozano-Álvarez, E. Dinámica poblacional de Creaseriella anops (Crustace: Isopoda) en cenotes dulceacuícolas de Quintana Roo. In La Carcinologia el México; Researchgate: Berlin, Germany, 2013. [Google Scholar]
- Fiers, F.; Reid, J.W.; Iliffe, T.M.; Suárez-Morales, E. New hypogean cyclopoid copepods (Crustacea) from the Yucatán Peninsula, Mexico. Contrib. Zool. 1996, 66, 65–102. [Google Scholar] [CrossRef]
- Rocha, C.E.F.; Iuffe, T.M.; Reid, J.W.; Suarez-Morales, E.; Båmstedt, U. A new species of Halicyclops (Copepoda, Cyclopoida, Cyclopidae) from cenotes of the Yucatan Peninsula, Mexico, with an identification key for the species of the genus from the Caribbean region and adjacent areas. Sarsia 1998, 83, 387–399. [Google Scholar] [CrossRef]
- Rocha, C.E.F.; Iliffe, T.M.; Reid, J.W.; Suárez-Morales, E. Prehendocyclops, a new genus of the subfamily Halicyclopinae (Copepoda, Cyclopoida, Cyclopidae) from cenotes of the Yucatan Peninsula, Mexico. Sarsia 2000, 85, 119–140. [Google Scholar] [CrossRef]
- Pesce, G.L.; Iliffe, T.M. New records of cave-dwelling mysids from the Bahamas and Mexico with description of Palaumysis bahamensis n. sp. (Crustacea: Mysidacea). J. Nat. Hist. 2002, 36, 265–278. [Google Scholar] [CrossRef]
- Strecker, U. The impact of invasive fish on an endemic Cyprinodon species flock (Teleostei) from Laguna Chichancanab, Yucatan, Mexico. Ecol. Freshw. Fish 2006, 15, 408–418. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Zylinski, S.; Jaume, D.; Iliffe, T.M.; Suárez-Morales, E. A new genus of speleophriid copepod (Copepoda: Misophrioida) from a cenote in the Yucatan, Mexico with a phylogenetic analysis at the species level. Zootaxa 2014, 3821, 321–336. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, F.; Iliffe, T.M.; Benítez, S.; Brankovits, D.; Villalobos, J.L. New records of anchialine fauna from the Yucatan Peninsula, Mexico. Check List 2015, 11, 1–10. [Google Scholar] [CrossRef]
- Mejía-Ortíz, L.M.; Pipan, T.; Culver, D.C.; Sprouse, P. The blurred line between photic and aphotic environments: A large Mexican cave with almost no dark zone. Int. J. Speleol. 2018, 47, 69–80. [Google Scholar] [CrossRef]
- Benítez, S.; Iliffe, T.M.; Quiroz-Martínez, B.; Alvarez, F. How is the anchialine fauna distributed within a cave? A study of the Ox Bel Ha System, Yucatan Peninsula, Mexico. Subterr. Biol. 2019, 31, 15–28. [Google Scholar] [CrossRef]
- Sánchez, N.; Martínez, A. Dungeons and dragons: Two new species and records of Kinorhyncha from anchialine cenotes and marine lava tubes. Zool. Anz. 2019, 282, 161–175. [Google Scholar] [CrossRef]
- Creaser, E.P. Larger Cave Crustacea of the Yucatan Peninsula. Carnegie Inst. Wash. Publ. 1938, 491, 159–164. [Google Scholar]
- Reddell, J.R. A Preliminary Bibliography of Mexican Cave Biology with a Checklist of Published Records. Assoc. Mex. Cave Stud. Bull. 1971, 3, 1–184. [Google Scholar]
- Holthuis, L.B. Cave shrimps (Crustacea: Decapoda, Natantia) from Mexico. In Subterranean Fauna of Mexico; Accademia Nazionale dei Lincei Quaderno: Rome, Italy, 1977; pp. 173–195. [Google Scholar]
- Ariani, A.P.; Wittmann, K.J.; Franco, E. A Comparative Study of Static Bodies in Mysid Crustaceans: Evolutionary Implications of Crystallographic Characteristics. Biol. Bull. 1993, 185, 393–404. [Google Scholar] [CrossRef]
- Barba-Macías, E.; Palacios-Vargas, J.G. Fauna acuática cavernícola de la Península de Yucatán. Mundos Subterráneos UMAE 1998, 9, 44–50. [Google Scholar]
- Botosaneanu, L.; Iliffe, T.M. On four new stygobitic cirolanids (Isopoda: Cirolanidae) and several already described species from Mexico and the Bahamas. Bull. l’Institut R. Sci. Nat. Belg. Biol. 1999, 69, 111–141. [Google Scholar]
- Chávez-Solís, E.M.; Mejía-Ortíz, L.M.; Simões, N. Predatory behavior of the cave shrimp Creaseria morleyi (Creaser, 1936) (Caridea: Palaemonidae), the blind hunter of the Yucatán cenotes, Mexico. J. Crustac. Biol. 2018, 38, 1–7. [Google Scholar] [CrossRef]
- Angyal, D.; Solís, E.C.; Magaña, B.; Balázs, G.; Simoes, N. Mayaweckelia troglomorpha, a new subterranean amphipod species from Yucatán state, México (Amphipoda, Hadziidae). Zookeys 2018. [Google Scholar] [CrossRef]
- Grego, J.; Angyal, D.; Beltrán, L.A.L. First record of subterranean freshwater gastropods (Mollusca, Gastropoda, Cochliopidae) from the cenotes of Yucatán state. Subterr. Biol. 2019, 29, 79–88. [Google Scholar] [CrossRef]
- Pohlman, J.W.; Cifuentes, L.A.; Iliffe, T.M. Food Web Dynamics and Biogeochemistry of Anchialine Caves: A Stable Isotope Approach. In Ecosystems of the World; Wilkens, H., Culver, D., Humphreys, W., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 30, pp. 351–363. [Google Scholar]
- Schmitter-Soto, J.J. Catálogo de los Peces Continentales de Quintana Roo; El Colegio de la Frontera Sur: Chetumal, Mexico, 1996; ISBN 968 7555 041. [Google Scholar]
- Medina–González, R.; Proudlove, G.; Chumba–Segura, L.; Iliffe, T. Threatened Fishes of the World: Ophisternon infernale (Hubbs, 1938) (Synbranchidae). Environ. Biol. Fishes 2001, 62, 170. [Google Scholar] [CrossRef]
- Navarro-Mendoza, M. Inventario íctico y estudios ecológicos preliminares en los cuerpos de agua continentales en la reserva de la biósfera de Sian Ka’an y áreas circunvecinas en Quintana Roo, México; Centro de Investigaciones de Quintana Roo/Secretaría de Desarrollo Urbano y Ecología/United States Fish & Wildlife Service: Washington, DC, USA, 1988. [Google Scholar]
- Proudlove, G.; Medina–González, R.; Chumba–Segura, L.; Iliffe, T. Threatened Fishes of the World: Ogilbia pearsei (Hubbs, 1938) (Bythitidae). Environ. Biol. Fishes 2001, 62, 214. [Google Scholar] [CrossRef]
- Sket, B. The ecology of anchihaline caves. Trends Ecol. Evol. 1996, 11, 221–225. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2019; Volume 49, ISBN 9780198820765. [Google Scholar]
- Calderón-Gutiérrez, F.; Sánchez-Ortiz, C.A.; Huato-Soberanis, L. Ecological patterns in anchialine caves. PLoS ONE 2018, 13, e0202909. [Google Scholar] [CrossRef]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Brankovits, D.; Pohlman, J.W.; Niemann, H.; Leigh, M.B.; Leewis, M.C.; Becker, K.W.; Iliffe, T.M.; Alvarez, F.; Lehmann, M.F.; Phillips, B. Methane-and dissolved organic carbon-fueled microbial loop supports a tropical subterranean estuary ecosystem. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Steinich, B. Investigaciones geofísicas e hidrogeológicas en el noroeste de la Península de Yucatán, México. Ph.D. Thesis, Instituto de Geofísica, Universidad Nacional Autónoma de México, México, 1996. [Google Scholar]
- Alcocer, J.; Lugo, A.; Marín, L.E.; Escobar, E. Hydrochemistry of waters from five cenotes and evaluation of their suitability for drinking-water supplies, northeastern Yucatan, Mexico. Hydrogeol. J. 1998, 6, 293–301. [Google Scholar] [CrossRef]
- Coke, J.G. Underwater Caves of the Yucatán Peninsula. In Encyclopedia of Caves; Elsevier: Amsterdam, The Netherlands, 2012; pp. 833–838. ISBN 9780123838322. [Google Scholar]
- Iliffe, T.M.; Kornicker, L.S. Worldwide diving discoveries of living fossil animals from the depths of anchialine and marine caves. Smithson. Contrib. Mar. Sci. 2009, 38. [Google Scholar]
- Gonzalez, B.C.; Iliffe, T.M.; Macalady, J.L.; Schaperdoth, I.; Kakuk, B. Microbial hotspots in anchialine blue holes: Initial discoveries from the Bahamas. Hydrobiologia 2011, 677, 149–156. [Google Scholar] [CrossRef]
- Seymour, J.; Humphreys, W.; Mitchell, J. Stratification of the microbial community inhabiting an anchialine sinkhole. Aquat. Microb. Ecol. 2007, 50, 11–24. [Google Scholar] [CrossRef][Green Version]
- Van Beynen, P.; Townsend, K. A disturbance index for karst environments. Environ. Manag. 2005, 36, 101–116. [Google Scholar] [CrossRef]
- Pacheco-Ávila, J.; Cabrera-Sansores, A.; Pérez-Ceballos, R. Diagnóstico de la calidad del agua subterránea en los sistemas municipales de abastecimiento en el Estado de Yucatán, México. Ingeniería 2004, 8, 165–179. [Google Scholar]
- Aguilar-Duarte, Y.; Bautista, F.; Mendoza, M.E.; Frausto, O.; Ihl, T.; Delgado, C. Ivaky: Índice De La Vulnerabilidad Del Acuífero Kárstico Yucateco A La Contaminación. Rev. Mex. Ing. Química 2016, 15, 913–933. [Google Scholar]
- Bautista, F.; Aguilar Duarte, Y.; Batllori, E. Amenazas, vulnerabilidad y riesgo de contaminación de las aguas subterráneas en la Península de Yucatán. Teoríay Prax. 2010, 7, 9–31. [Google Scholar] [CrossRef]
- Casas-Beltrán, D.A.; Gallaher, C.M.; Hernandez Yac, E.; Febles Moreno, K.; Voglesonger, K.; Leal-Bautista, R.M.; Lenczewski, M. Seaweed Invasion! Temporal Changes in Beach Conditions Lead to Increasing Cenote Usage and Contamination in the Riviera Maya. Sustainability 2020, 12, 2474. [Google Scholar] [CrossRef]
- Mateo-Sagasta, J.; Zadeh, S.M.; Turral, H. Water Pollution from Agriculture: A global Review; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy; International Water Management Institute (IWMI): Colombo, Sri Lanka, 2017. [Google Scholar]
- Lopez-Maldonado, Y.; Berkes, F. Restoring the environment, revitalizing the culture: Cenote conservation in Yucatan, Mexico. Ecol. Soc. 2017, 22, art7. [Google Scholar] [CrossRef]
- Hernández-Terrones, L.; Rebolledo-Vieyra, M.; Merino-Ibarra, M.; Soto, M.; Le-Cossec, A.; Monroy-Ríos, E. Groundwater Pollution in a Karstic Region (NE Yucatan): Baseline Nutrient Content and Flux to Coastal Ecosystems. Water Air Soil Pollut. 2010, 218, 517–528. [Google Scholar] [CrossRef]
- INEGI. Anuario Estadístico y Geográfico de Yucatán 2014; INEGI: Mérida, Yucatán, Mexico, 2014; ISBN 9786077393658. [Google Scholar]
- INEGI. Anuario Estadístico y Geográfico de Yucatán 2015; INEGI: Mérida, Yucatán, Mexico, 2015. [Google Scholar]
- Lutz, W.; Prieto, L.; Anderson, W. Population, Development, and Environment on the Yucatán Peninsula: From Ancient Maya to 2030; ILASA: Laxenburg, Austria, 2000; ISBN 3-7045-0138-7. [Google Scholar]
- Perez Brito, E.A.; de Jesus Espinosa Atoche, T.; Patricia Quintal Gordillo, K. Factors of Competitiveness for the Bovine Livestock in Yucatan, Mexico. In Bovine Science—A Key to Sustainable Development; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar]
- Danielopol, D.L.; Griebler, C.; Gunatilaka, A.; Notenboom, J. Present state and future prospects for groundwater ecosystems. Environ. Conserv. 2003, 30, 104–130. [Google Scholar] [CrossRef]
- Arcega-Cabrera, F.; Garza-Pérez, R.; Noreña-Barroso, E.; Oceguera-Vargas, I. Impacts of Geochemical and Environmental Factors on Seasonal Variation of Heavy Metals in a Coastal Lagoon Yucatan, Mexico. Bull. Environ. Contam. Toxicol. 2015, 94, 58–65. [Google Scholar] [CrossRef]
- White, W.B. Contaminant Transport in Karst Aquifers: Systematics and Mechanisms; White, W.B., Herman, J.S., Herman, E.K., Rutigliano, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 55–81. [Google Scholar]
- White, W.B. Hydrogeology of Karst Aquifers. In Encyclopedia of Caves; Elsevier: Amsterdam, The Netherlands, 2012; pp. 383–391. ISBN 9780123838322. [Google Scholar]
- White, W.B. Karst Groundwater Contamination and Public Health; White, W.B., Herman, J.S., Herman, E.K., Rutigliano, M., Eds.; Advances in Karst Science; Springer International Publishing: Cham, Switzerland, 2018; Volume 13, ISBN 978-3-319-51069-9. [Google Scholar]
- Kløve, B.; Ala-Aho, P.; Bertrand, G.; Gurdak, J.J.; Kupfersberger, H.; Kværner, J.; Muotka, T.; Mykrä, H.; Preda, E.; Rossi, P.; et al. Climate change impacts on groundwater and dependent ecosystems. J. Hydrol. 2014, 518, 250–266. [Google Scholar] [CrossRef]
- Arcega-Cabrera, F.; Velázquez-Tavera, N.; Fargher, L.; Derrien, M.; Noreña-Barroso, E. Fecal sterols, seasonal variability, and probable sources along the ring of cenotes, Yucatan, Mexico. J. Contam. Hydrol. 2014, 168, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kantun Manzano, C.; Arcega-Cabrera, F.; Derrien, M.; Noreña-Barroso, E.; Herrera-Silveira, J. Submerged Groundwater Discharges as Source of Fecal Material in Protected Karstic Coastal Areas. Geofluids 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Boyd, A.; Stewart, C.B.; Philibert, D.A.; How, Z.T.; El-Din, M.G.; Tierney, K.B.; Blewett, T.A. A burning issue: The effect of organic ultraviolet filter exposure on the behaviour and physiology of Daphnia magna. Sci. Total Environ. 2021, 750, 1–11. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, T.; Zhou, C.; Cheng, P.; Bai, Q.; Ao, J.; Wang, W.; Zhang, H. Effects of four commonly used UV filters on the growth, cell viability and oxidative stress responses of the Tetrahymena thermophila. Chemosphere 2013, 93, 2507–2513. [Google Scholar] [CrossRef] [PubMed]
- Olmstead, A.W.; LeBlanc, G.A. Toxicity Assessment of Environmentally Relevant Pollutant Mixtures Using a Heuristic Model. Integr. Environ. Assess. Manag. 2005, 1, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Ruszkiewicz, J.A.; Pinkas, A.; Ferrer, B.; Peres, T.V.; Tsatsakis, A.; Aschner, M. Neurotoxic effect of active ingredients in sunscreen products, a contemporary review. Toxicol. Rep. 2017, 4, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Lee, Y.G.; Kim, S.D.; Cha, C.J.; Ahn, J.H.; Hur, H.G. Biodegradation of the insecticide N,N-diethyl-m-toluamide by fungi: Identification and toxicity of metabolites. Arch. Environ. Contam. Toxicol. 2005. [Google Scholar] [CrossRef] [PubMed]
- Aronson, R.B.; Swanson, D.W. Video surveys of coral reefs: Uni- and multivariate applications. In Proceedings of the 8th International Coral Reef Symposium; Lessios, H.A., Macintyre, I.G., Eds.; Smithsonian Tropical Research Institute: Panama City, Panama, 1997; Volume 2, pp. 1441–1446. [Google Scholar]
- Giokas, D.L.; Salvador, A.; Chisvert, A. UV filters: From sunscreens to human body and the environment. TrAC Trends Anal. Chem. 2007, 26, 360–374. [Google Scholar] [CrossRef]
- Brausch, J.M.; Rand, G.M. A review of personal care products in the aquatic environment: Environmental concentrations and toxicity. Chemosphere 2011, 82, 1518–1532. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | Year | Author | N | Locations | Reference |
|---|---|---|---|---|---|
| Q. Roo | 1977 | Bowman | 5 | 69, 70, 73, 77, 81 | [37] |
| 1987 | Bowman | 1 | 79 | [42] | |
| Yager | 1 | 77 | [43] | ||
| 1990 | Holsinger | 3 | 74, 77, 79 | [44] | |
| 1992 | Iliffe | 1 | 79 | [20] | |
| 1993 | Ruiz-Cancino | 2 | 83, 84 | [45] | |
| 1996 | Fiers et al. | 3 | 66, 68, 71 | [46] | |
| 1997 | Pohlman et al. | 1 | 75 | [21] | |
| 1998 | Rocha et al. | 1 | 75 | [47] | |
| 2000 | Rocha et al. | 1 | 72 | [48] | |
| 2002 | Pesce & Iliffe | 1 | 75 | [49] | |
| 2006 | Strecker | 1 | 67 | [50] | |
| 2014 | Boxshall et al. | 1 | 77 | [51] | |
| 2015 | Álvarez et al. | 2 | 76, 78 | [52] | |
| 2018 | Mejía-Ortiz et al. | 1 | 82 | [53] | |
| 2019 | Benítez et al. | 2 | 76, 78 | [54] | |
| Sánchez & Martínez | 1 | 80 | [55] | ||
| Yucatan | 1936 | Creaser | 2 | 17,54 | [18] |
| 1938 | Creaser | 12 | 4, 12, 17, 31, 39, 46, 50, 53, 54, 55, 56, 60 | [56] | |
| 1971 | Reddell | 11 | 4, 12, 17, 31, 39, 46, 50, 53, 54, 55, 56, 60 | [57] | |
| 1977 | Bowman | 18 | 3, 5, 13, 14, 17, 19, 20, 25, 27, 29, 36, 42, 46, 61, 62, 63, 64, 65 | [37] | |
| Holthuis | 2 | 57, 59 | [58] | ||
| Reddell | 4 | 6, 9, 15, 30 | [19] | ||
| 1985 | Villalobos | 1 | 57 | [28] | |
| 1991 | Iliffe | 3 | 38, 55, 57 | [27] | |
| Rodríguez-Almaraz | 1 | 49 | [27] | ||
| 1993 | Ariani et al. | 1 | 27 | [59] | |
| 1996 | Fiers et al. | 4 | 2, 21, 23, 58 | [46] | |
| 1998 | Barba-Macías & Palacios-Vargas | 1 | 27 | [60] | |
| Rocha et al. | 3 | 21, 27, 37 | [47] | ||
| 1999 | Botosaneanu & Iliffe | 1 | 24 | [61] | |
| 2000 | Rocha et al. | 4 | 21, 27, 37, 41 | [48] | |
| 2002 | Pesce & Iliffe | 2 | 23, 52 | [49] | |
| 2017 | Chávez-Solís et al. | 2 | 18, 38 | [62] | |
| 2018 | Angyal et al. | 4 | 10, 18, 43, 44 | [63] | |
| 2019 | Grego et al. | 1 | 51 | [64] | |
| 2020 | Angyal et al. | 20 | 1, 7, 8, 10, 11, 16, 18, 22, 26, 28, 32, 33, 34, 35, 40, 43, 44, 45, 47, 48 | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liévano-Beltrán, L.A.; Simões, N. Updated Distribution of the Mysid Antromysis cenotensis (Crustacea: Peracarida), a Protected Key Species in Yucatan Peninsula Cenotes. Diversity 2021, 13, 154. https://doi.org/10.3390/d13040154
Liévano-Beltrán LA, Simões N. Updated Distribution of the Mysid Antromysis cenotensis (Crustacea: Peracarida), a Protected Key Species in Yucatan Peninsula Cenotes. Diversity. 2021; 13(4):154. https://doi.org/10.3390/d13040154
Chicago/Turabian StyleLiévano-Beltrán, Luis Arturo, and Nuno Simões. 2021. "Updated Distribution of the Mysid Antromysis cenotensis (Crustacea: Peracarida), a Protected Key Species in Yucatan Peninsula Cenotes" Diversity 13, no. 4: 154. https://doi.org/10.3390/d13040154
APA StyleLiévano-Beltrán, L. A., & Simões, N. (2021). Updated Distribution of the Mysid Antromysis cenotensis (Crustacea: Peracarida), a Protected Key Species in Yucatan Peninsula Cenotes. Diversity, 13(4), 154. https://doi.org/10.3390/d13040154