
The Evolutionary History, Diversity, and Ecology of Willows (Salix L.) in the European Alps


Abstract
1. Introduction
2. Phylogenetic Relationships and Biogeography
Sectional Classification and Character Evolution
3. Evolution and Ecology of Species
3.1. Evolution of Crossing Barriers
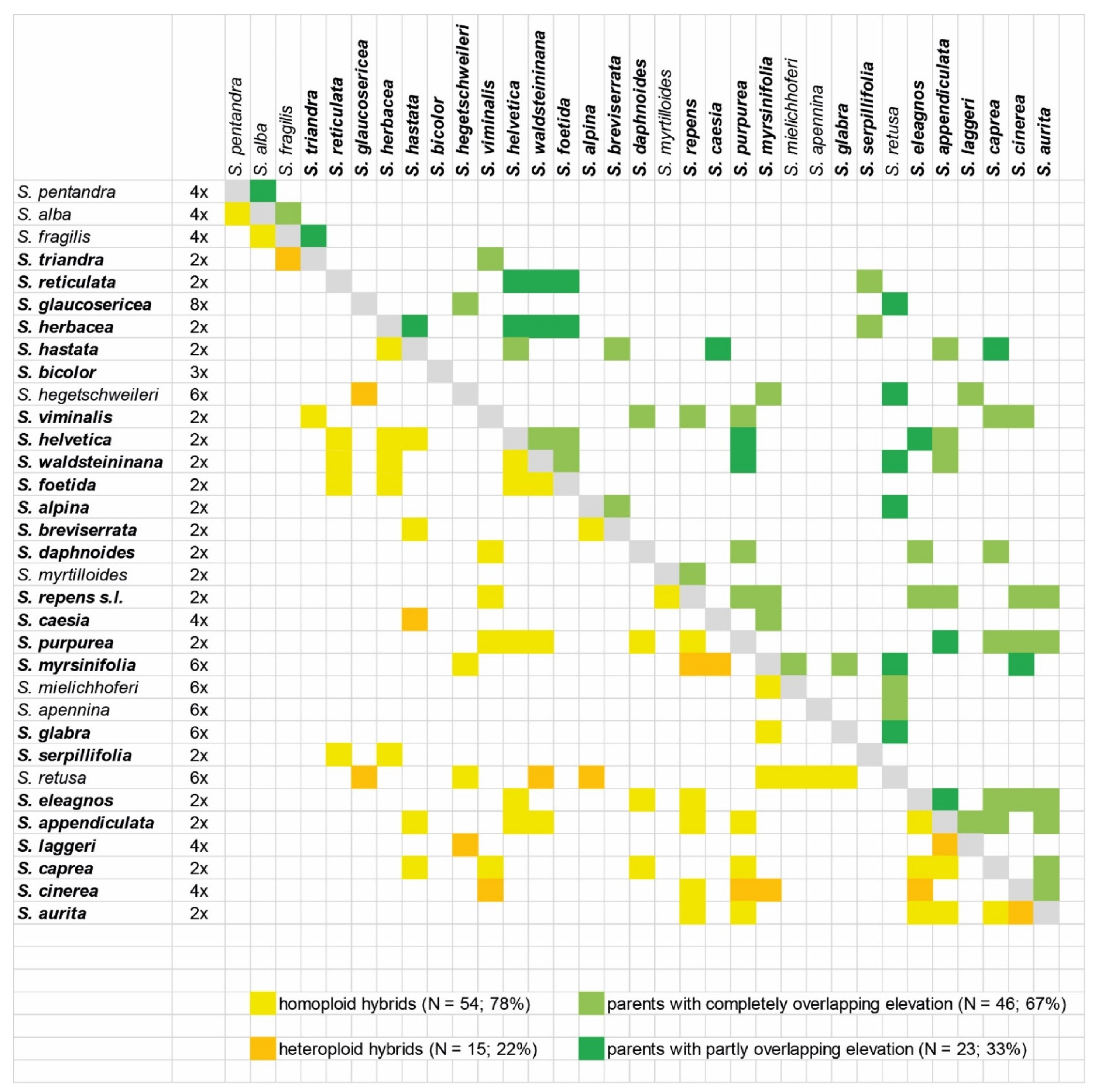
3.2. Hybridization in Secondary Contact Zones as a Consequence of Climatic Changes
3.3. Polyploids: Allopolyploid Speciation
4. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aeschimann, D.; Lauber, K.; Moser, D.; Theurillat, J.-P. Flora Alpina. Ein Atlas Sämtlicher 4500 Gefäßpflanzen der Alpen; Haupt: Bern, Switzerland, 2004. [Google Scholar]
- Buser, R. Kritische Beiträge zur Kenntnis der schweizerischen Weiden. Ber. Schweiz. Bot. Ges. 1940, 50, 567–788. [Google Scholar] [CrossRef]
- Toepffer, A. Salices Bavariae. Versuch einer Monographie der bayerischen Weiden unter Berücksichtigung der Arten der mitteleuropäischen Flora. Ber. Bayer. Bot. Ges. 1915, 15, 17–233. [Google Scholar]
- Rechinger, K.H. Salix. In Illustrierte Flora von Mitteleuropa III/1, 2nd ed.; Hegi, G., Ed.; Hanser: Munich, Germany, 1957; pp. 44–135. [Google Scholar]
- Skvortsov, A. Willows of Russia and Adjacent Countries; University of Joensuu: Joensuu, Finland, 1999. [Google Scholar]
- Chen, J.H.; Sun, H.; Wen, J.; Yang, Y.P. Molecular phylogeny of Salix L. (Salicaceae) inferred from three chloroplast datasets and its systematic implications. Taxon 2010, 59, 29–37. [Google Scholar] [CrossRef]
- Wagner, N.D.; He, L.; Hörandl, E. Phylogenomic Relationships and Evolution of Polyploid Salix Species Revealed by RAD Sequencing Data. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- IPNI. 2020 International Plant Names Index. The Royal Botanic Gardens Kew, Harvard University Herbaria & Libraries and Australian National Botanic Gardens. Available online: http://www.ipni.org (accessed on 2 March 2021).
- Meneghetti, S.; Barcaccia, G.; Paiero, P.; Lucchin, M. Genetic characterization of Salix alba L. and Salix fragilis L. by means of different PCR-derived marker systems. Plant Biosyst. 2007, 141, 283–291. [Google Scholar] [CrossRef]
- Oberprieler, C.; Dietz, L.; Harlander, C.; Heilmann, J. Molecular and phytochemical evidence for the taxonomic integrity of Salix alba, S-fragilis, and their hybrid S. x rubens (Salicaceae) in mixed stands in SE Germany. Plant Syst. Evol. 2013, 299, 1107–1118. [Google Scholar] [CrossRef]
- Hörandl, E.; Florineth, F.; Hadacek, F. Weiden in Österreich und Angrenzenden Gebieten [Willows in Austria and Adjacent Regions], 2nd ed.; University of Agriculture: Vienna, Austria, 2012; 164p. [Google Scholar]
- Hardig, T.M.; Brunsfeld, S.J.; Fritz, R.S.; Morgan, M.; Orians, C.M. Morphological and molecular evidence for hybridization and introgression in a willow (Salix) hybrid zone. Mol. Ecol. 2000, 9, 9–24. [Google Scholar] [CrossRef]
- Barcaccia, G.; Meneghetti, S.; Albertini, E.; Triest, L.; Lucchin, M. Linkage mapping in tetraploid willows: Segregation of molecular markers and estimation of linkage phases support an allotetraploid structure for Salix alba x Salix fragilis interspecific hybrids. Heredity 2003, 90, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Triest, L.; De Greef, B.; De Bondt, R.; Van Slycken, J. RAPD of controlled crosses and clones from the field suggests that hybrids are rare in the Salix alba-Salix fragilis complex. Heredity 2000, 84, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Gramlich, S.; Sagmeister, P.; Dullinger, S.; Hadacek, F.; Hörandl, E. Evolution in situ: Hybrid origin and establishment of willows (Salix L.) on alpine glacier forefields. Heredity 2016, 116, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Karrenberg, S.; Kollmann, J.; Edwards, P.J. Pollen vectors and inflorescence morphology in four species of Salix. Plant Syst. Evol. 2002, 235, 181–188. [Google Scholar] [CrossRef]
- Abbott, R.; Albach, D.; Ansell, S.; Arntzen, J.W.; Baird, S.J.E.; Bierne, N.; Boughman, J.W.; Brelsford, A.; Buerkle, C.A.; Buggs, R.; et al. Hybridization and speciation. J. Evol. Biol. 2013, 26, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. Quaternary phylogeography: The roots of hybrid zones. Genetica 2011, 139, 617–638. [Google Scholar] [CrossRef] [PubMed]
- Kadereit, J.W. The geography of hybrid speciation in plants. Taxon 2015, 64, 673–687. [Google Scholar] [CrossRef]
- Abbott, R.; Brennan, A.C. Altitudinal gradients, plant hybrid zones and evolutionary novelty. Philos. Trans. R. Soc. B 2014, 369, 20130346. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A. Die mitteleuropäischen Salix-Arten. Mitt. Forstl. Bundes-Vers. Wien 1981, 134, 1–152. [Google Scholar]
- Suda, Y.; Argus, G.W. Chromosome numbers of some North American Salix. Brittonia 1968, 20, 191–197. [Google Scholar] [CrossRef]
- Suda, Y.; Argus, G.W. Chromosome numbers of some North American arctic and boreal Salix. Can. J. Bot. 1969, 47, 859. [Google Scholar] [CrossRef]
- Martini, F.; Paiero, P. I Salici d’Italia; Edizione Lint: Trient, Italy, 1988. [Google Scholar]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen, 6th ed.; Eugen Ulmer: Stuttgart, Germany, 2010. [Google Scholar]
- Grabherr, G.; Mucina, L. Die Pflanzengesellschaften Österreichs. Teil II; G. Fischer: Jena, Germany, 1993. [Google Scholar]
- Schiechtl, H.M. Weiden in der Praxis—Die Weiden Mitteleuropas, ihre Verwendung und Bestimmung; Patzer: Berlin, Germany; Hannover, Germany, 1992; 130p. [Google Scholar]
- Wagner, N.D.; Gramlich, S.; Hörandl, E. RAD sequencing resolved phylogenetic relationships in European shrub willows (Salix L. subg. Chamaetia and subg. Vetrix) and revealed multiple evolution of dwarf shrubs. Ecol. Evol. 2018, 8, 8243–8255. [Google Scholar] [CrossRef] [PubMed]
- Karrenberg, S.; Suter, M. Phenotypic trade-offs in the sexual reproduction of Salicaceae from flood plains. Am. J. Bot. 2003, 90, 749–754. [Google Scholar] [CrossRef]
- Percy, D.M.; Argus, G.W.; Cronk, Q.C.; Fazekas, A.J.; Kesanakurti, P.R.; Burgess, K.S.; Husband, B.C.; Newmaster, S.G.; Barrett, S.C.H.; Graham, S.W. Understanding the spectacular failure of DNA barcoding in willows (Salix): Does this result from a trans-specific selective sweep? Mol. Ecol. 2014, 23, 4737–4756. [Google Scholar] [CrossRef] [PubMed]
- Leskinen, E.; Alstrom-Rapaport, C. Molecular phylogeny of Salicaceae and closely related Flacourtiaceae: Evidence from 5.8 S, ITS 1 and ITS 2 of the rDNA. Plant Syst. Evol. 1999, 215, 209–227. [Google Scholar] [CrossRef]
- Azuma, T.; Kajita, T.; Yokoyama, J.; Ohashi, H. Phylogenetic relationships of Salix (Salicaceae) based on rbcL sequence data. Am. J. Bot. 2000, 87, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lauron-Moreau, A.; Pitre, F.E.; Argus, G.W.; Labrecque, M.; Brouillet, L. Phylogenetic Relationships of American Willows (Salix L.; Salicaceae). PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Savage, J.A.; Cavender-Bares, J. Habitat specialization and the role of trait lability in structuring diverse willow (genus Salix) communities. Ecology 2012, 93, S138–S150. [Google Scholar] [CrossRef]
- Wu, J.; Nyman, T.; Wang, D.C.; Argus, G.W.; Yang, Y.P.; Chen, J.H. Phylogeny of Salix subgenus Salix s.l. (Salicaceae): Delimitation, biogeography, and reticulate evolution. BMC Evol. Biol. 2015, 15. [Google Scholar] [CrossRef]
- Zhang, L.; Xi, Z.X.; Wang, M.C.; Guo, X.Y.; Ma, T. Plastome phylogeny and lineage diversification of Salicaceae with focus on poplars and willows. Ecol. Evol. 2018, 8, 7817–7823. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wagner, N.D.; Hörandl, E. RAD sequencing data reveal a radiation of willow species (Salix L.; Salicaceae) in the Hengduan Mountains and adjacent areas. J. Syst. Evol. 2021, 59, 44–57. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.S.; Wang, D.S.; Zhang, J.G. Phylogeny of Populus-Salix (Salicaceae) and their relative genera using molecular datasets. Biochem. Syst. Ecol. 2016, 68, 210–215. [Google Scholar] [CrossRef]
- Collinson, M.E. The early fossil history of Salicaceae—A brief review. Proc. R. Soc. Edinb. Sect. B Biol. Sci. 1992, 98, 155–167. [Google Scholar] [CrossRef]
- Alsos, I.G.; Eidesen, P.B.; Ehrich, D.; Skrede, I.; Westergaard, K.; Jacobsen, G.H.; Landvik, J.Y.; Taberlet, P.; Brochmann, C. Frequent long-distance plant colonization in the changing Arctic. Science 2007, 316, 1606–1609. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P. Salix. In Flora Iberica; Castroviejo, S., Cirujano, S., Montserrat, P., Munoz Garmendia, F., Paiva, J., Eds.; Real Jardin Botanico: Madrid, Spain, 1993; pp. 477–517. [Google Scholar]
- Rice, A.; Smarda, P.; Novosolov, M.; Drori, M.; Glick, L.; Sabath, N.; Meiri, S.; Belmaker, J.; Mayrose, I. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Körner, C. Alpine Plant Life. Functional Plant Ecology of High Mountain Systems; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Abbott, R.J.; Hegarty, M.J.; Hiscock, S.J.; Brennan, A.C. Homoploid hybrid speciation in action. Taxon 2010, 59, 1375–1386. [Google Scholar] [CrossRef]
- Fritz, R.S.; Hochwender, C.G.; Brunsfeld, S.J.; Roche, B.M. Genetic architecture of susceptibility to herbivores in hybrid willows. J. Evol. Biol. 2003, 16, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Arraiano-Castilho, R.; Bidartondo, M.I.; Niskanen, T.; Clarkson, J.J.; Brunner, I.; Zimmermann, S.; Senn-Irlet, B.; Frey, B.; Peintner, U.; Mrak, T.; et al. Habitat specialisation controls ectomycorrhizal fungi above the treeline in the European Alps. N. Phytol. 2021, 229, 2901–2916. [Google Scholar] [CrossRef] [PubMed]
- Arraiano-Castilho, R.; Bidartondo, M.I.; Niskanen, T.; Zimmermann, S.; Frey, B.; Brunner, I.; Senn-Irlet, B.; Hörandl, E.; Gramlich, S.; Suz, L.M. Plant-fungal interactions in hybrid zones: Ectomycorrhizal communities of willows (Salix) in an alpine glacier forefield. Fungal Ecol. 2020, 45. [Google Scholar] [CrossRef]
- He, L.; Jia, K.-H.; Zhang, R.-G.; Wang, Y.; Shi, T.-L.; Li, Z.-C.; Zeng, S.-W.; Cai, X.-J.; Wagner, N.D.; Hörandl, E.; et al. Chromosome-scale assembly of the genome of Salix dunnii reveals a male-heterogametic sex determination system on chromosome 7. Mol. Ecol. Resour. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Gramlich, S.; Wagner, N.D.; Hörandl, E. RAD-seq reveals genetic structure of the F-2-generation of natural willow hybrids (Salix L.) and a great potential for interspecific introgression. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Change 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Lautenschlager-Fleury, D.; Lautenschlager-Fleury, E. Der Gletschboden, ein Weidenparadies. Bauhinia 1985, 8, 89–98. [Google Scholar]
- Gramlich, S.; Hörandl, E. Fitness of natural willow hybrids in a pioneer mosaic hybrid zone. Ecol. Evol. 2016, 6, 7645–7655. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Willis, J.H. Plant speciation. Science 2007, 317, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, C. Willows. The Genus Salix; Timber Press: Portland, OR, USA, 1992. [Google Scholar]
- Kadereit, J.W. The role of in situ species diversification for the evolution of high vascular plant species diversity in the European Alps—A review and interpretation of phylogenetic studies of the endemic flora of the Alps. Perspect. Plant Ecol. Evol. Syst. 2017, 26, 28–38. [Google Scholar] [CrossRef]
- Ramsey, J.; Schemske, D.W. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annu. Rev. Ecol. Syst. 1998, 29, 467–501. [Google Scholar] [CrossRef]
- Schinkel, C.C.F.; Kirchheimer, B.; Dullinger, S.; Geelen, D.; De Storme, N.; Hörandl, E. Pathways to polyploidy: Indications of a female triploid bridge in the alpine species Ranunculus kuepferi (Ranunculaceae). Plant Syst. Evol. 2017, 303, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Schoenfelder, K.P.; Fox, D.T. The expanding implications of polyploidy. J. Cell Biol. 2015, 209, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Syngelaki, E.; Schinkel, C.C.F.; Klatt, S.; Hörandl, E. Effects of temperature treatments on cytosine-methylation profiles of diploid and autotetraploid plants of the alpine species Ranunculus kuepferi (Ranunculaceae). Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Feliner, G.N.; Casacuberta, J.; Wendel, J.F. Genomics of Evolutionary Novelty in Hybrids and Polyploids. Front. Genet. 2020, 11, 21. [Google Scholar] [CrossRef]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.; Wang, D.Y.; Zhang, L.; Kang, M.H.; Lu, Z.Q.; Zhu, R.B.; Mao, X.X.; Xi, Z.X.; Ma, T. Intergeneric Relationships within the Family Salicaceae s.l. Based on Plastid Phylogenomics. Int. J. Mol. Sci. 2019, 20, 3788. [Google Scholar] [CrossRef]
- Zhou, R.; Macaya-Sanz, D.; Rodgers-Melnick, E.; Carlson, C.H.; Gouker, F.E.; Evans, L.M.; Schmutz, J.; Jenkins, J.W.; Yan, J.Y.; Tuskan, G.A.; et al. Characterization of a large sex determination region in Salix purpurea L. (Salicaceae). Mol. Genet. Genom. 2018, 293, 1437–1452. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wei, S.Y.; Pan, H.X.; Zhuge, Q.; Yin, T.M. Uneven selection pressure accelerating divergence of Populus and Salix. Hortic. Res. 2019, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.H.; Gouker, F.E.; Crowell, C.R.; Evans, L.; DiFazio, S.P.; Smart, C.D.; Smart, L.B. Joint linkage and association mapping of complex traits in shrub willow (Salix purpurea L.). Ann. Bot. 2019, 124, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Kosinski, P.; Sliwinska, E.; Hilpold, A.; Boratynski, A. DNA ploidy in Salix retusa agg. only partly in line with its morphology and taxonomy. Nord. J. Bot. 2019, 37. [Google Scholar] [CrossRef]
- Little, C.J.; Wheeler, J.A.; Sedlacek, J.; Cortes, A.J.; Rixen, C. Small-scale drivers: The importance of nutrient availability and snowmelt timing on performance of the alpine shrub Salix herbacea. Oecologia 2016, 180, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Hojsgaard, D.; Hörandl, E. The Rise of Apomixis in Natural Plant Populations. Front. Plant Sci. 2019, 10, 358. [Google Scholar] [CrossRef]
- Ikeno, S. On hybridization of some species of Salix. Ann. Bot. 1922, 36, 175–191. [Google Scholar] [CrossRef]
- Hojsgaard, D.; Klatt, S.; Baier, R.; Carman, J.G.; Hörandl, E. Taxonomy and biogeography of apomixis in angiosperms and associated biodiversity characteristics. Crit. Rev. Plant. Sci. 2014, 33, 414–427. [Google Scholar] [CrossRef]
- Lautenschlager-Fleury, D.; Lautenschlager-Fleury, E. Salix laggeri Wimmer—Monographie einer wenig bekannten Weide. Bauhinia 1984, 8, 149–156. [Google Scholar]
- Li, W.; Wu, H.T.; Li, X.P.; Chen, Y.N.; Yin, T.M. Fine mapping of the sex locus in Salix triandra confirms a consistent sex determination mechanism in genus Salix. Hortic. Res. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Brereton, N.J.B.; Gonzalez, E.; Marleau, J.; Guidi Nissim, W.; Labrecque, M.; Joly, S.; Pitre, F.E. Comparative Transcriptomic Approaches Exploring Contamination Stress Tolerance in Salix sp Reveal the Importance for a Metaorganismal de Novo Assembly Approach for Nonmodel Plants. Plant. Physiol. 2016, 171, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yuan, H.W.; Li, Y.J.; Chen, Y.H.; Liu, G.Y.; Ye, M.X.; Yu, C.M.; Lian, B.L.; Zhong, F.; Jiang, Y.N.; et al. Genome sequencing and phylogenetic analysis of allotetraploid Salix matsudana Koidz. Hortic. Res. 2020, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Kosinski, P.; Sekiewicz, K.; Walas, L.; Boratynski, A.; Dering, M. Spatial genetic structure of the endemic alpine plant Salix serpillifolia: Genetic swamping on nunataks due to secondary colonization? Alp. Bot 2019, 129, 107–121. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Taxon | Author (Year) * | Distribution | Elevation and Habitat | Ploidy Level | Allopolyploid Origins | Phylogenetic Position |
|---|---|---|---|---|---|---|
| Subg. Salix | ||||||
| S. alba ** | L. (1753) | Europe to W. Asia | colline riversides | 4x | [6] | |
| S. fragilis ** | L. (1753) | Europe to W. Asia | colline riversides | 4x | Wagner et al. subm. | |
| S. pentandra | L. (1753) | Europe to W. Siberia | montane–subalpine riversides, shrubs | 4x | [6] | |
| S. triandra | L. (1753) | Eurasia, N-Africa | colline–montane riversides, lakes | 2x | [7] | |
| Subg. Vetrix s.l. (Chamaetia/Vetrix clade) | ||||||
| S. alpina | Scop. (1772) | Alps, Carpathians, Dinarids, Balcan | subalpine screes, rocks, snowbeds | 2x | [7] | |
| S. apennina | Skvorts. (1965) | Apennines, Alps | colline–montane shrubs | 6x | ||
| S. appendiculata | Vill. (1789) | Alps, Central European mts., Dinarids | montane–subalpine shrubs | 2x | [7] | |
| S. aurita | L. (1753) | Europe to Siberia | colline-montane wetlands, bogs | 2x | [7] | |
| S. bicolor | Ehrh. ex Willd. (1796) | Pyrenees, Massif Central, Vosges, Alps, Sudeten and Carpathians | montane–subalpine shrubs | 3x | [7] | |
| S. breviserrata | Flod. (1940) | Cantabrian mts., Alps, Apennines | Subalpine screes, rocks | 2x | [7] | |
| S. caesia | Vill. (1789) | Alps, and Central Asia | Subalpine wetlands | 4x | S. purpurea × repens | |
| S. caprea | L. (1753) | Eurasia, lowland | colline–subalpine pioneer forests | 2x | [7] | |
| S. cinerea | L. (1753) | Eurasia, lowland | colline–montane wetlands, bogs | 4x | S. appendiculata × aurita | [7] |
| S. daphnoides | Vill. (1779) | European lowland and mountains | Montane shrubs | 2x | [7] | |
| S. eleagnos | Scop. (1772) | Central and Southern Europe | colline-montane | 2x | [7] | |
| S. foetida | Schleich. ex DC. (1805) | Pyrenees, Alps, Apennines | subalpine wetlands | 2x | [7] | |
| S. glabra | Scop. (1772) | Alps, Dinarids | montane–subalpine screes | 6x | 2 parents? | [7] |
| S. glaucosericea | Flod. (1943) | Alps | subalpine shrubs | 8x | 3–4 parents? | [7] |
| S. hastata | L. (1753) | circumboreal-European mountain system | subalpine shrubs | 2x | [7] | |
| S. hegetschweileri *** | Heer (1840) | Alps | montane–subalpine shrubs, glacier moraines | 4x–6x | S. bicolor × myrsinifolia? | |
| S. helvetica | Vill. (1789) | Alps, Carpathians | subalpine shrubs | 2x | [7] | |
| S. herbacea | L. (1753) | circumarctic-European mountain system | alpine snowbeds | 2x | [7] | |
| S. laggeri | Wimm. (1854) | Alps | subalpine shrubs | 4x | S. caprea × appendiculata | [7] |
| S. mielichhoferi | Saut. (1849) | Alps | subalpine shrubs | 6x | ||
| S. myrtilloides | L. (1753) | Eurasia | montane bogs | 2x | ||
| S. myrsinifolia | Salisb. (1796) | European mountains | montane wetlands, riversides | 6x | 3 parents? | [7] |
| S. purpurea | L. (1753) | European lowland | colline–subalpine alluvials, moraines | 2x | [7] | |
| S. repens s.l. **** | L. (1753) | Eurasia | colline–montane wet meadows | 2x | [7] | |
| S. retusa | L. (1759) | European mountain system | alpine snowbeds, screes, rocks | 6x | ||
| S. reticulata | L. (1753) | circumarctic-European mountain system | alpine snowbeds | 2x | [7] | |
| S. serpillifolia | Scop. (1772) | Alps, Dinarids | alpine screes | 2x | [7] | |
| S. viminalis | L. (1753) | European lowland | colline river and lakesides | 2x | [7] | |
| S. waldsteininana | Willd. (1806) | Alps, Dinarids, Balcan | subalpine shrubs | 2x | [7] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, N.D.; He, L.; Hörandl, E. The Evolutionary History, Diversity, and Ecology of Willows (Salix L.) in the European Alps. Diversity 2021, 13, 146. https://doi.org/10.3390/d13040146
Wagner ND, He L, Hörandl E. The Evolutionary History, Diversity, and Ecology of Willows (Salix L.) in the European Alps. Diversity. 2021; 13(4):146. https://doi.org/10.3390/d13040146
Chicago/Turabian StyleWagner, Natascha D., Li He, and Elvira Hörandl. 2021. "The Evolutionary History, Diversity, and Ecology of Willows (Salix L.) in the European Alps" Diversity 13, no. 4: 146. https://doi.org/10.3390/d13040146
APA StyleWagner, N. D., He, L., & Hörandl, E. (2021). The Evolutionary History, Diversity, and Ecology of Willows (Salix L.) in the European Alps. Diversity, 13(4), 146. https://doi.org/10.3390/d13040146