
Experimental Evaluation of δ2H, δ13C and δ15N Variability in Blood and Feathers of Wild and Captive Birds: Implications for Interspecific Food Web Studies

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Monitoring of Breeding Birds in Nest Boxes
2.3. Cross-Fostering in Swallows and Kestrels
2.4. Dietary Samples—Swallows and Kestrels
2.5. Captive Mallards
2.6. Stable Isotope Analyses
2.7. Statistical Analyses
3. Results
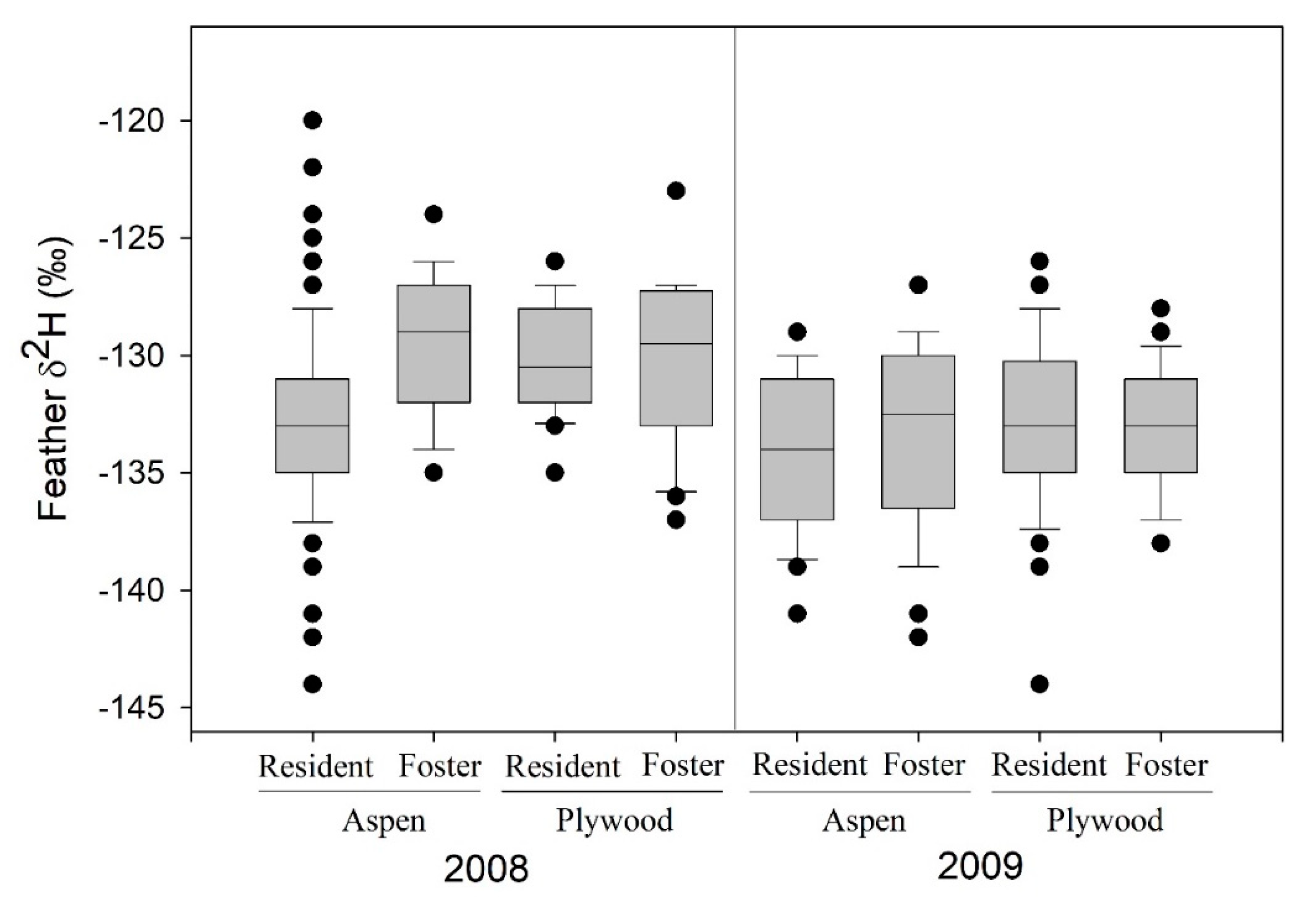
3.1. Cross-Fostering Experiments
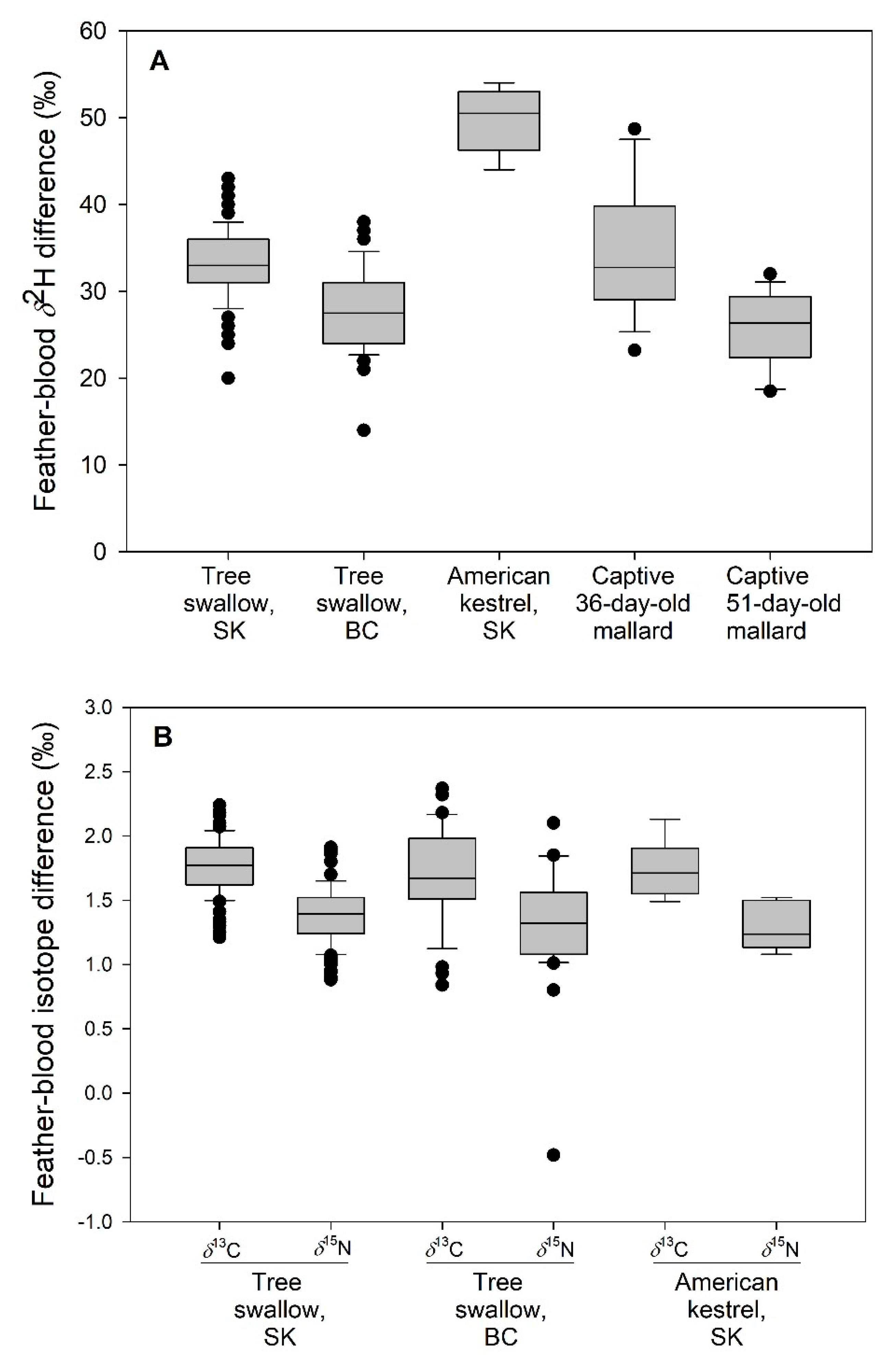
3.2. δ2H in Tree Swallow and American Kestrel Dietary Samples
3.3. Captive Mallard Ducklings
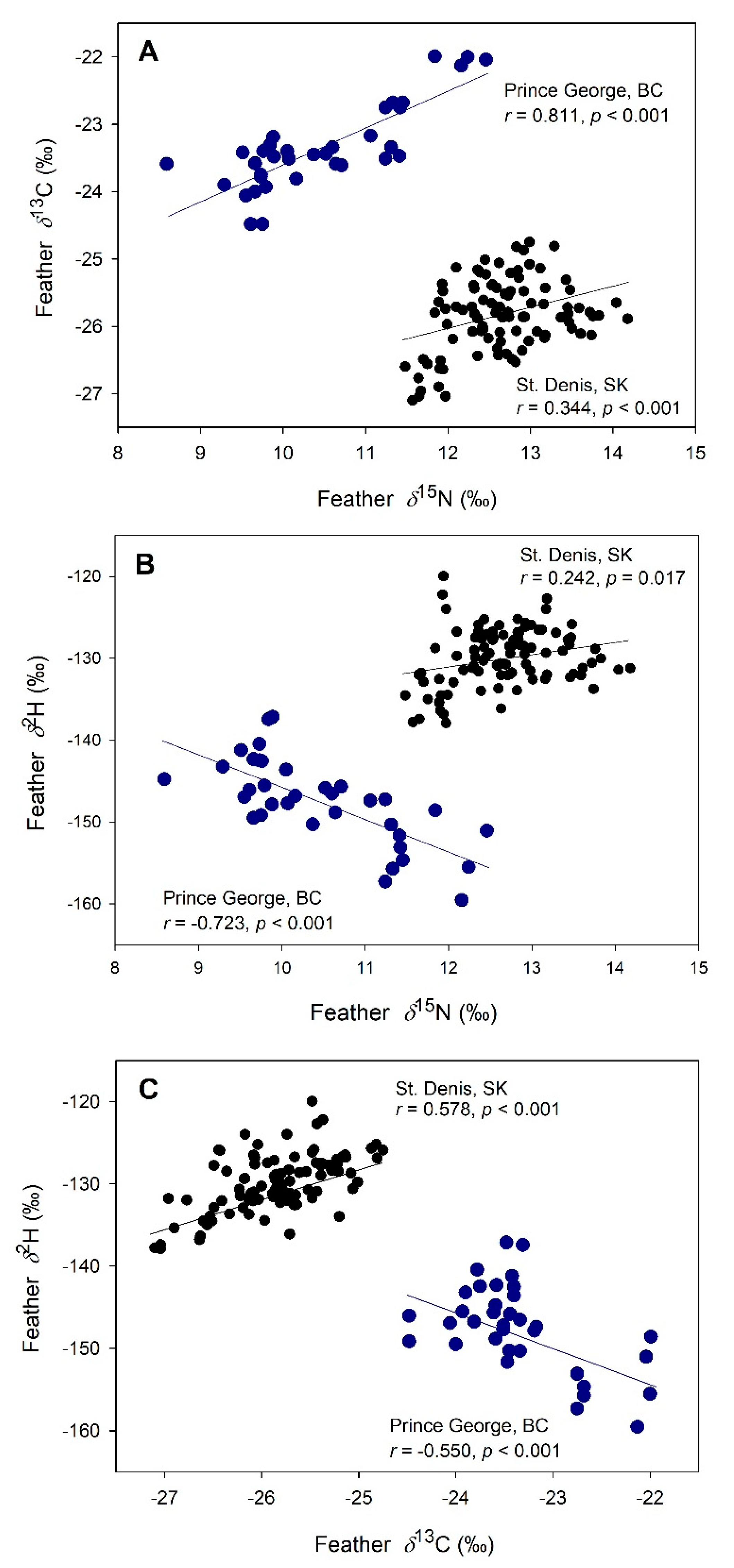
3.4. Exploring Blood-Feather Isotope Differences among Species and Sites
4. Discussion
Exploratory Blood-Feather Relationships
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hobson, K.A. Using stable isotopes to trace long-distance dispersal in birds and other taxa. Divers. Distrib. 2005, 11, 157–164. [Google Scholar] [CrossRef]
- Inger, R.; Bearhop, S. Applications of stable isotopes analyses to avian ecology. Ibis 2008, 150, 447–461. [Google Scholar] [CrossRef]
- Wolf, N.; Carleton, S.A.; Martínez del Rio, C.M. Ten years of experimental animal isotopic ecology. Funct. Ecol. 2009, 23, 17–26. [Google Scholar] [CrossRef]
- Boecklen, W.J.; Yarnes, C.T.; Cook, B.A.; James, A.C. On the use of stable isotopes in trophic ecology. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 411–440. [Google Scholar] [CrossRef] [Green Version]
- Vander Zanden, H.B.; Soto, D.X.; Bowen, G.J.; Hobson, K.A. Expanding the isotopic toolbox: Application of hydrogen and oxygen stable isotope ratios to food web studies. Front. Ecol. Evol. 2016, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Van Wijk, R.E.; Barshep, Y.; Hobson, K.A. On the use of stable hydrogen isotope measurements (δ2H) to discern trophic level in avian terrestrial food webs. Diversity 2021, 13, 202. [Google Scholar] [CrossRef]
- Hobson, K.A.; Van Wilgenburg, S.L.; Wassenaar, L.I.; Larson, K. Linking hydrogen (δ2H) isotopes in feathers and precipitation: Sources of variance and consequences for assignment to isoscapes. PLoS ONE 2012, 7, e35137. [Google Scholar] [CrossRef]
- Vander Zanden, H.B.; Wunder, M.B.; Hobson, K.A.; Van Wilgenburg, S.L.; Wassenaar, L.I.; Welker, J.M.; Bowen, G.J. Contrasting assignment of migratory organisms to geographic origins using long-term versus year-specific precipitation isotope maps. Methods Ecol. Evol. 2014, 5, 891–900. [Google Scholar] [CrossRef] [Green Version]
- Nordell, C.J.; Haché, S.; Bayne, E.M.; Sólymos, P.; Foster, K.R.; Godwin, C.M.; Krikun, R.; Pyle, P.; Hobson, K.A. Within-site variation in feather stable hydrogen isotope (δ2Hf) values of boreal songbirds: Implications for assignment to molt origin. PLoS ONE 2016, 11, e0163957. [Google Scholar] [CrossRef]
- McKechnie, A.E.; Wolf, B.O.; Martínez del Rio, C. Deuterium stable isotope ratios as tracers of water resource use: An experimental test with rock doves. Oecologia 2004, 140, 191–200. [Google Scholar] [CrossRef]
- Betini, G.S.; Hobson, K.A.; Wassenaar, L.I.; Norris, D.R. Stable hydrogen isotope values (δD) in songbird nestlings: Effects of diet, temperature and body size. Can. J. Zool. 2009, 87, 767–772. [Google Scholar] [CrossRef]
- Criste, P.; Møller, A.P.; Saino, N.; De Lope, F. Genetic and environmental components of phenotypic variation in immune response and body size of a colonial bird, Delichon urbica (the house martin). Heredity 2000, 85, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Tella, J.L.; Bortolotti, G.R.; Forero, M.G.; Dawson, R.D. Environmental and genetic variation in T-cell mediated immune response of fledgling American kestrels. Oecologia 2000, 123, 453–459. [Google Scholar] [CrossRef]
- Evans, M.R.; Roberts, M.L.; Buchanan, K.L.; Goldsmith, A.R. Heritability of corticosterone response and changes in life history traits during selection in the zebra finch. J. Evol. Biol. 2006, 19, 343–352. [Google Scholar] [CrossRef]
- Griebel, I.A.; Fairhurst, G.D.; Marchant, T.A.; Clark, R.G. Effects of parental and nest site characteristics on nestling quality in the tree swallow (Tachycineta bicolor). Can. J. Zool. 2018, 97, 63–71. [Google Scholar] [CrossRef]
- Pym, R.A.E. Nutritional Genetics. In Poultry Breeding and Genetics; Crawford, R.D., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 847–876. [Google Scholar]
- Bortolotti, L.E.; Clark, R.G.; Wassenaar, L.I. Hydrogen isotope variability in prairie wetland systems: Implications for studies of migratory connectivity. Ecol. Appl. 2013, 23, 110–121. [Google Scholar] [CrossRef]
- Fairhurst, G.D.; Treen, G.; Clark, R.G.; Bortolotti, G.R. Nestling corticosterone response to microclimate in an altricial bird. Can. J. Zool. 2012, 90, 1422–1430. [Google Scholar] [CrossRef]
- Johns, D.W.; Marchant, T.A.; Fairhurst, G.D.; Speakman, J.; Clark, R.G. Biomarker of burden: Feather corticosterone reflects energetic expenditure and allostatic overload in captive waterfowl. Funct. Ecol. 2017, 32, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Magozzi, C.; Vander Zanden, H.B.; Wunder, M.B.; Bowen, G.J. Mechanistic model predicts tissue–environment relationships and trophic shifts in animal hydrogen and oxygen isotope ratios. Oecologia 2019, 191, 777–789. [Google Scholar] [CrossRef]
- Meehan, T.D.; Rosenfield, R.N.; Atudorei, V.N.; Bielfeldt, J.; Rosenfield, L.J.; Stewart, A.C.; Stoudt, W.E.; Bozek, M.A. Variation in stable-hydrogen isotope ratios between adult and nestling Cooper’s hawks. Condor 2003, 105, 567–572. [Google Scholar] [CrossRef]
- Smith, A.D.; Dufty, A.M. Variation in the stable-hydrogen isotope composition of northern goshawk feathers: Relevance to the study of migratory origins. Condor 2005, 107, 547–558. [Google Scholar] [CrossRef]
- Greenwood, J.L.; Dawson, R.D. Correlates of deuterium enrichment in the feathers of adult American kestrels of known origin. Condor 2011, 113, 555–564. [Google Scholar] [CrossRef]
- Shutler, D.; Clark, R.G. Causes and consequences of tree swallow (Tachycineta bicolor) dispersal in Saskatchewan. Auk 2003, 120, 619–631. [Google Scholar] [CrossRef]
- Griebel, I.A.; Dawson, R.D. Experimental reduction of nestling hemoglobin concentration in combination with ectoparasite load manipulation affects nestling morphology and begging behavior, but not adult behavior. J. Ornithol. 2020, 161, 35–45. [Google Scholar] [CrossRef]
- Griebel, I.A.; Dawson, R.D.; Clark, R.G. Cavity type influences abundance of nest-dwelling avian blow flies: An experiment with tree swallows. Ecol. Entomol. 2020, 45, 434–443. [Google Scholar] [CrossRef]
- Dawson, R.D. Timing of breeding and environmental factors as determinants of reproductive performance of tree swallows. Can. J. Zool. 2008, 86, 843–850. [Google Scholar] [CrossRef]
- Dawson, R.D.; Bortolotti, G.R. Fire in the boreal forest: Proximate effects on reproduction and long-term consequences for territory occupancy of American kestrels. Ecoscience 2006, 13, 75–81. [Google Scholar] [CrossRef]
- Bortolotti, G.R.; Tella, J.L.; Forero, M.G.; Dawson, R.D.; Negro, J.J. Genetics, local environment and health as factors influencing plasma carotenoids in wild American kestrels (Falco sparverius). Proc. R. Soc. Lond. B 2000, 267, 1433–1438. [Google Scholar] [CrossRef] [Green Version]
- Wassenaar, L.I.; Hobson, K.A. Comparative equilibration and online technique for determination of non-exchangeable hydrogen of keratins for use in animal migration studies. Isot. Environ. Health Stud. 2003, 39, 211–217. [Google Scholar] [CrossRef]
- Soto, D.X.; Koehler, G.; Wassenaar, L.I.; Hobson, K.A. Re-evaluation of the hydrogen stable isotopic composition of keratin calibration standards for wildlife and forensic science applications. Rapid Commun. Mass Specrom. 2017, 31, 1193–1203. [Google Scholar] [CrossRef]
- SAS Institute. Statistical Analysis System, version 9.4; SAS Institute: Cary, NC, USA, 2016. [Google Scholar]
- Elgin, A.S.; Clark, R.G.; Morrissey, C.A. Tree swallow selection of wetlands in agricultural landscapes predicted by central place foraging theory. Condor Ornithol. Appl. 2020, 122, duaa039. [Google Scholar] [CrossRef]
- Michelson, C.I.; Clark, R.G.; Morrissey, C.A. Diets of adult and nestling tree swallows in contrasting agricultural environments: Evidence from stable isotope analyses. Condor Ornithol. Appl. 2018, 120, 751–764. [Google Scholar]
- Wolf, N.; Newsome, S.D.; Fogel, M.L.; Martínez del Rio, C.M. An experimental exploration of the incorporation of hydrogen isotopes from dietary sources into avian tissues. J. Exp. Biol. 2012, 215, 1915–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wunder, M.; Hobson, K.A.; Kelly, J.; Marra, P.; Wassenaar, L.I.; Stricker, C.; Doucette, R. Does a lack of design and repeatability compromise scientific criticism? A response to Smith et al. Auk 2009, 126, 922–926. [Google Scholar] [CrossRef]
- Grénier, C.S.V.; Guglielmo, C.G.; Mitchell, G.W.; Falconer, M.; Hobson, K.A. Nutritional consequences of breeding away from riparian habitats in bank swallows: New evidence from multiple endogenous markers. Conserv. Physiol. 2021, 9, coaa140. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fixed Effects | Random Effects | ||||||
|---|---|---|---|---|---|---|---|
| Species/Isotope | Year | Sampling Date | Cross-Foster | Box Type | Natal Nest | Rearing Nest | n |
| Tree swallows (SK) | |||||||
| δ2H | 14.08 *** | 0.54 | 0.14 | 0.22 | 0.16 a | 3.81 *** | 228 |
| δ13C | na | 8.88 ** | 0.33 | 0.66 | 0.22 | 3.07 *** | 96 |
| δ15N | na | 0.4 | 0.09 | 0.65 | inest | 3.15 ** | 96 |
| Tree swallows (BC) | |||||||
| δ2H | na | 1.54 | 7.10 * | na | 0.64 | 1.61 * | 35 |
| δ13C | na | 12.00 ** | 5.99 * | na | 0.78 | 1.76 * | 35 |
| δ15N | na | 3.80 a | 0.45 | na | inest | 1.81 * | 35 |
| American kestrels | |||||||
| δ2H | na | na | 0.28 | na | 0.57 | inest | 16 |
| δ13C | na | na | 0.84 | na | 0.76 | 1.14 | 16 |
| δ15N | na | na | 0.27 | na | inest | 0.11 | 16 |
| Stable Isotope | Species | |||
|---|---|---|---|---|
| Tissue | Tree Swallow (BC) | Tree Swallow (SK) | Americanm Kestrel (SK) | |
| δ2H | Blood | −175 ± 5 (36) | −163 ± 3 (96) | −154 ± 2 (8) |
| Feather | −147 ± 5 (36) | −130 ± 3 (96) | −105 ± 3 (16) | |
| Difference | 28 ± 5 (36) | 33 ± 4 (96) | 50 ± 4 (8) | |
| δ15N | Blood | 9.1 ± 0.9 (36) | 11.3 ± 0.6 (96) | 6.0 ± 0.1 (8) |
| Feather | 10.5 ± 0.9 (35) | 12.7 ± 0.6 (96) | 7.5 ± 0.3 (16) | |
| Difference | 1.3 ± 0.4 (35) | 1.4 ± 0.2 (96) | 1.3 ± 0.2 (8) | |
| δ13C | Blood | −25.0 ± 0.5 (36) | −27.6 ± 0.4 (96) | −25.2 ± 0.5 (8) |
| Feather | −23.3 ± 0.6 (35) | −25.8 ± 0.5 (96) | −23.7 ± 0.5 (16) | |
| Difference | 1.7 ± 0.4 (35) | 1.8 ± 0.2 (96) | 1.7 ± 0.2 (8) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clark, R.G.; Dawson, R.D.; Greenwood, J.L.; Johns, D.W.; Wassenaar, L.I.; Hobson, K.A. Experimental Evaluation of δ2H, δ13C and δ15N Variability in Blood and Feathers of Wild and Captive Birds: Implications for Interspecific Food Web Studies. Diversity 2021, 13, 495. https://doi.org/10.3390/d13100495
Clark RG, Dawson RD, Greenwood JL, Johns DW, Wassenaar LI, Hobson KA. Experimental Evaluation of δ2H, δ13C and δ15N Variability in Blood and Feathers of Wild and Captive Birds: Implications for Interspecific Food Web Studies. Diversity. 2021; 13(10):495. https://doi.org/10.3390/d13100495
Chicago/Turabian StyleClark, Robert G., Russell D. Dawson, Jennifer L. Greenwood, David W. Johns, Leonard I. Wassenaar, and Keith A. Hobson. 2021. "Experimental Evaluation of δ2H, δ13C and δ15N Variability in Blood and Feathers of Wild and Captive Birds: Implications for Interspecific Food Web Studies" Diversity 13, no. 10: 495. https://doi.org/10.3390/d13100495
APA StyleClark, R. G., Dawson, R. D., Greenwood, J. L., Johns, D. W., Wassenaar, L. I., & Hobson, K. A. (2021). Experimental Evaluation of δ2H, δ13C and δ15N Variability in Blood and Feathers of Wild and Captive Birds: Implications for Interspecific Food Web Studies. Diversity, 13(10), 495. https://doi.org/10.3390/d13100495