Characterization of Genetic Diversity Conserved in the Gene Bank for Dutch Cattle Breeds


Abstract
1. Introduction
2. Materials and Methods
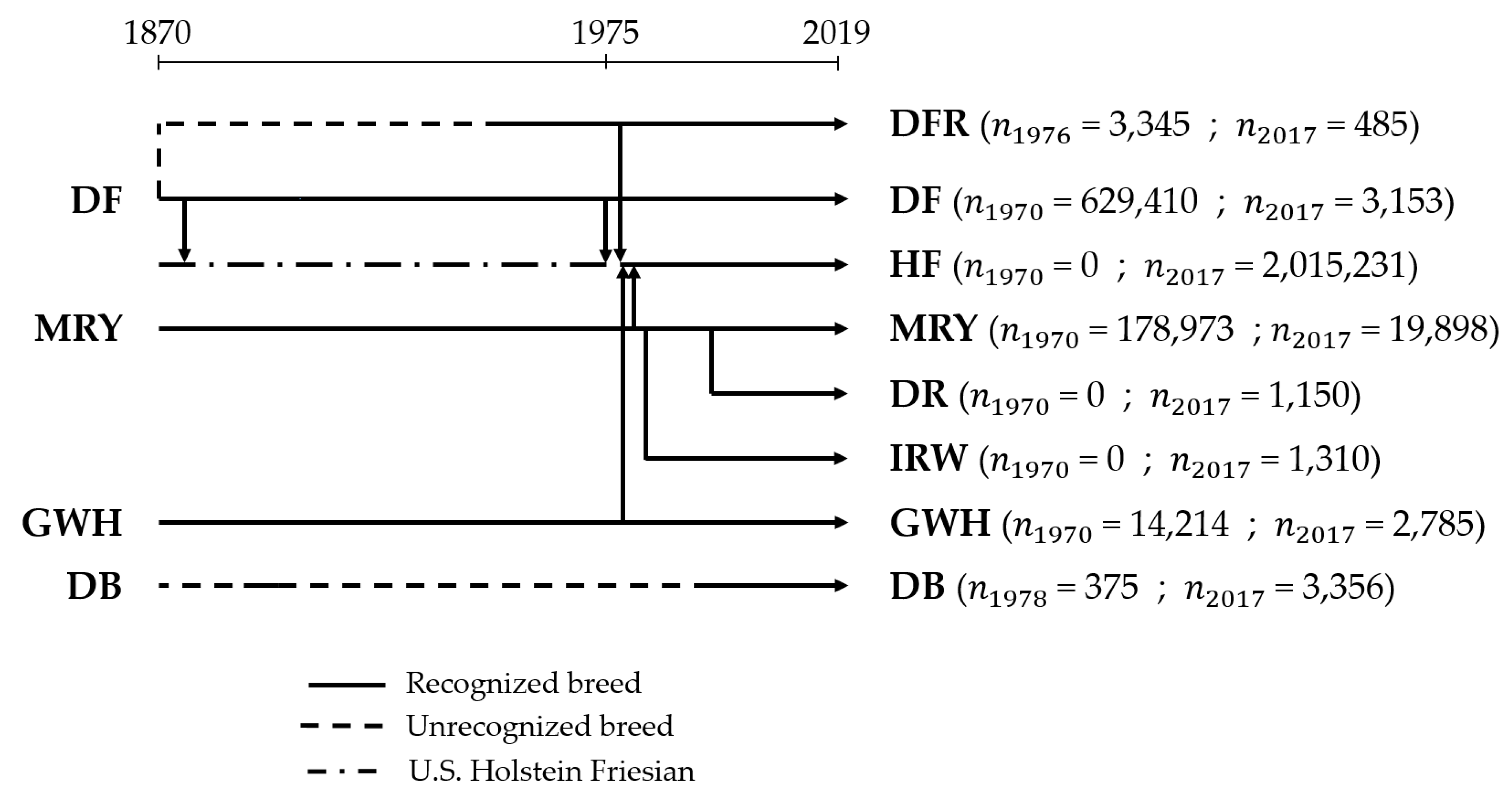
2.1. Data
2.2. Characterization of Genetic Diversity within and between Breeds
2.3. Optimizing Diversity in Collections
3. Results
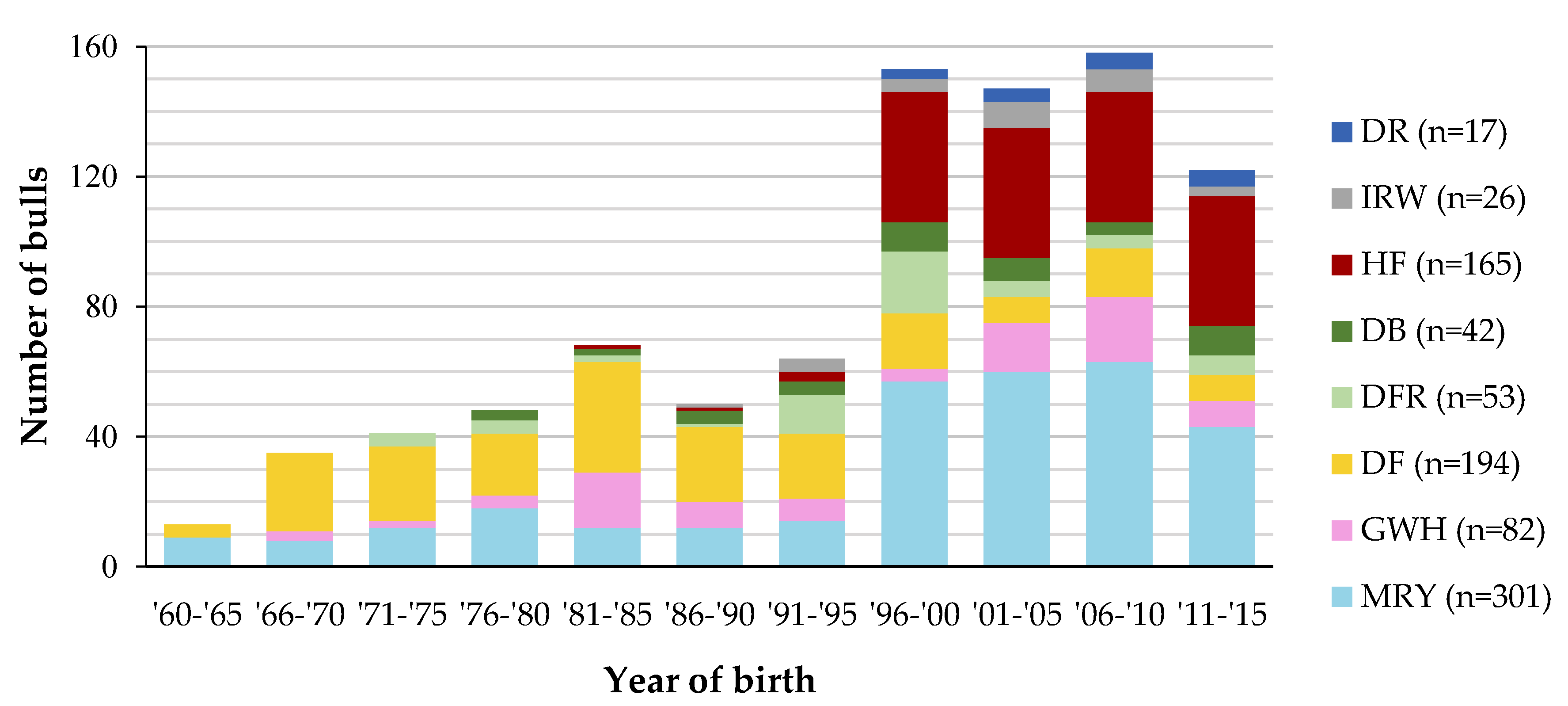
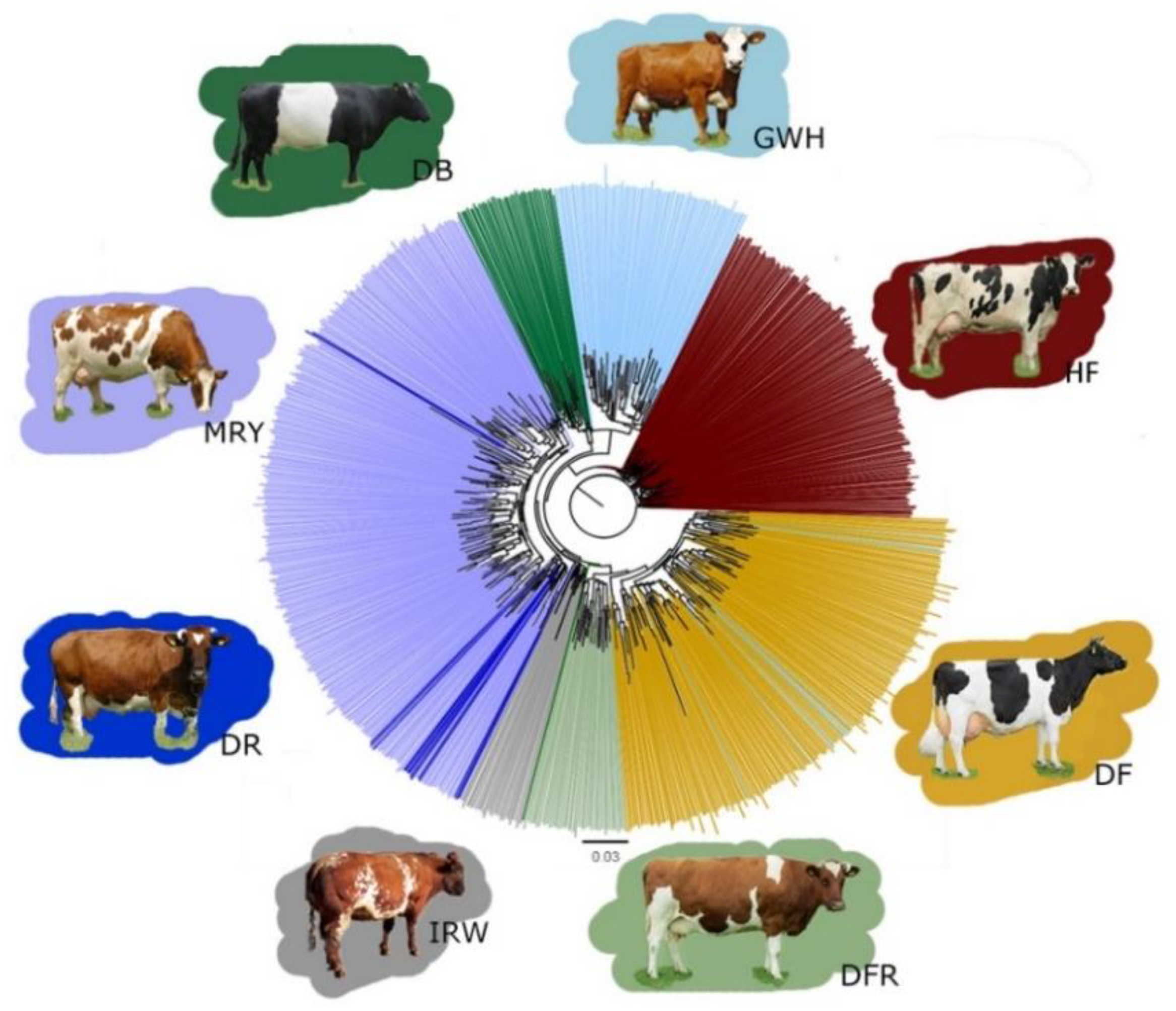
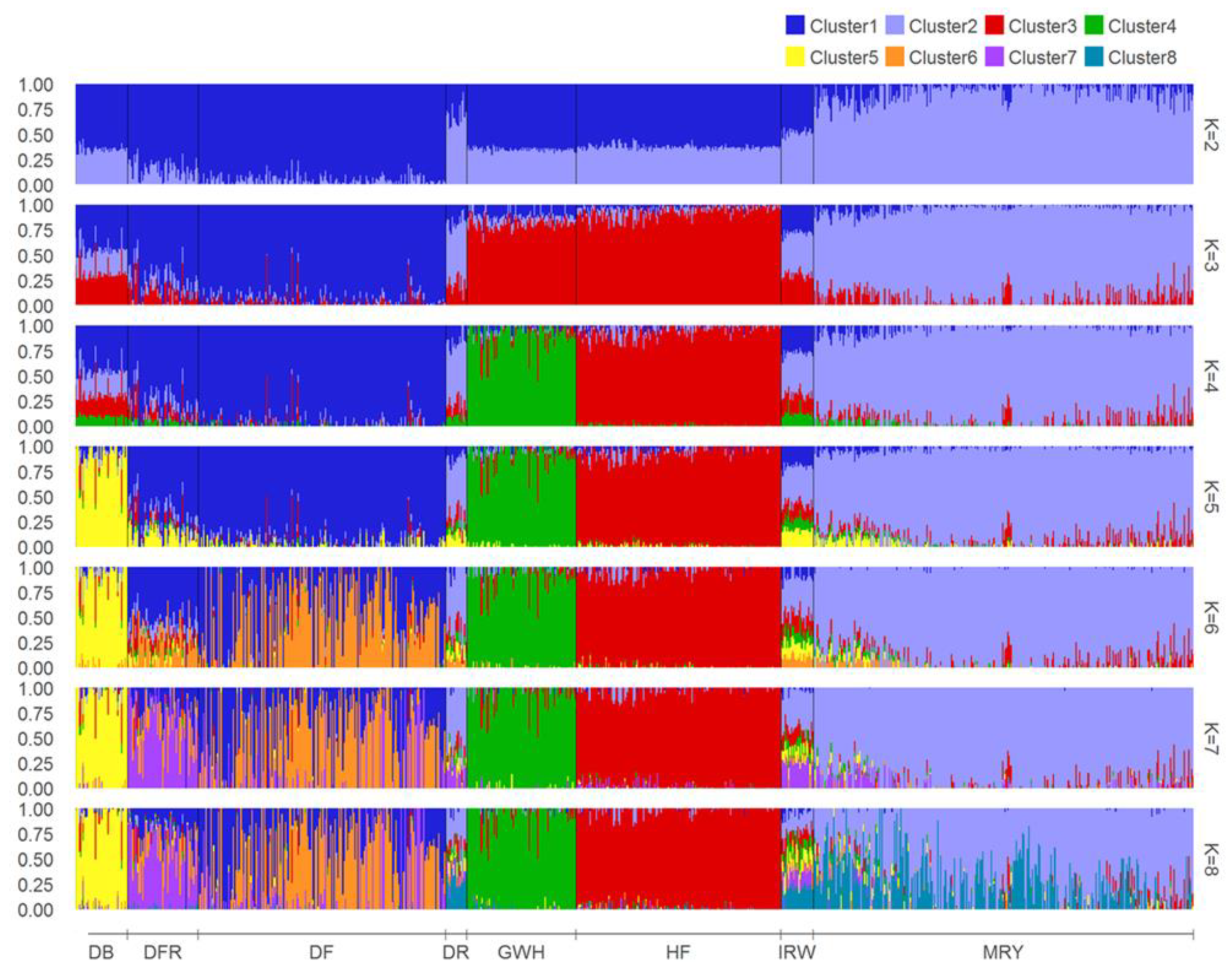
3.1. Characterization of Genetic Diversity within and between Breeds
3.2. Optimization of Diversity across Breeds
3.3. Optimization of Diversity within Breeds
4. Discussion
4.1. Characterization of Cattle Breeds in the Dutch Gene Bank
4.2. Management and Optimization of Genetic Diversity within Breeds
4.3. Value of Old and Recent Germplasm Material
4.4. Management of Genetic Diversity across Breeds
4.5. From Genotype to Sequence Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oldenbroek, K. Utilisation and Conservation of Farm Animal Genetic Resources; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 55–71. [Google Scholar]
- FAO. Global Plan of Action for Animal Genetic Resources and the Interlaken Declaration; FAO: Rome, Italy, 2007. [Google Scholar]
- Bennewitz, J.; Simianer, H.; Meuwissen, T.H.E. Investigations on Merging Breeds in Genetic Conservation Schemes. J. Dairy Sci. 2008, 91, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Smith, C. Genetic aspects of conservation in farm livestock. Livest. Prod. Sci. 1984, 11, 37–48. [Google Scholar] [CrossRef]
- Eding, H.; Meuwissen, T.H.E. Linear methods to estimate kinships from genetic marker data for the construction of core sets in genetic conservation schemes. J. Anim. Breed. Genet. 2003, 120, 289–302. [Google Scholar] [CrossRef]
- Theunissen, B. Breeding without Mendelism: Theory and practice of dairy cattle breeding in The Netherlands 1900–1950. J. Hist. Biol. 2008, 41, 637–676. [Google Scholar] [CrossRef]
- Fernández, J.; Meuwissen, T.H.E.; Toro, M.A.; Mäki-Tanila, A. Management of genetic diversity in small farm animal populations. Animal 2011, 5, 1684–1698. [Google Scholar] [CrossRef]
- Danchin-Burge, C.; Hiemstra, S.J.; Blackburn, H. Ex situ conservation of Holstein-Friesian cattle: Comparing the Dutch, French, and US germplasm collections. J. Dairy Sci. 2011, 94, 4100–4108. [Google Scholar] [CrossRef]
- Doekes, H.P.; Veerkamp, R.F.; Bijma, P.; Hiemstra, S.J.; Windig, J.J. Trends in genome-wide and region-specific genetic diversity in the Dutch-Flemish Holstein-Friesian breeding program from 1986 to 2015. Genet. Sel. Evol. 2018, 50, 1–16. [Google Scholar] [CrossRef]
- CMD. Jaarverslag Centrale Melkcontrole Dienst (Central Milkrecording Service); Stichting Centrale Melk Controle Dienst: Arnhem, The Netherlands, 1971. [Google Scholar]
- Maurice van Eijndhoven, M. Genetic Variation of Milk Fatty Acid Composition Between and Within Dairy Cattle Breeds. 2014. Available online: https://edepot.wur.nl/322983 (accessed on 28 November 2019).
- Van Helden, W.; Minkema, D. Inventarisatie van Zeldzame Huisdierrassen; SZH: Groningen, The Netherlands, 1978. [Google Scholar]
- CGN. Rassenlijst Nederlandse Landbouwhuisdierrassen en hun Risicostatus; CGN: Wageningen, The Netherlands, 2017. [Google Scholar]
- Malécot, G. Mathématiques de l’Hérédité; Masson & Cie: Paris, France, 1948. [Google Scholar]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Figtree v1.4. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 21 June 2019).
- Raj, A.; Stephens, M.; Pritchard, J.K. FastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.M. POPHELPER: An R package and web app to analyse and visualize population structure. Mol. Ecol. Resour. 2017, 17, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Berg, P.; Windig, J.J. Genomic Management of Animal Genetic Diversity; Oldenbroek, K., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; pp. 162–166. [Google Scholar]
- Grundy, B.; Villanueva, B.; Woolliams, J.A. Dynamic selection procedures for constrained inbreeding and their consequences for pedigree development. Genet. Res. 1998, 72, 159–168. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Sonesson, A.K. Maximizing the Response of Selection with a Predefined Rate of Inbreeding: Overlapping Generations. J. Anim. Sci. 1997, 76, 2575–2583. [Google Scholar] [CrossRef] [PubMed]
- Eynard, S.E.; Windig, J.J.; Hulsegge, I.; Hiemstra, S.J.; Calus, M.P.L. The impact of using old germplasm on genetic merit and diversity—A cattle breed case study. J. Anim. Breed. Genet. 2018, 135, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E. Gencont: An operational tool for controlling inbreeding in selection and conservation schemes. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 18–23 August 2002. [Google Scholar]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.A.M.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 1–16. [Google Scholar] [CrossRef]
- Mil, R.; Van Nauta, W.J. Fundamentfokkerij Fries Hollands vee. Available online: https://edepot.wur.nl/183413 (accessed on 28 November 2019).
- FAO. Cryoconservation of Animal Genetic Resources; FAO Animal: Rome, Italy, 2012. [Google Scholar]
- Doekes, H.P.; Veerkamp, R.F.; Bijma, P.; Hiemstra, S.J.; Windig, J. Value of the Dutch Holstein Friesian germplasm collection to increase genetic variability and improve genetic merit. J. Dairy Sci. 2018, 101, 10022–10033. [Google Scholar] [CrossRef]
- Silva, R.D.O.; Moran, D.; Vosough, B.; Sipke, A.; Hiemstra, J. Optimizing ex situ genetic resource collections for European livestock conservation. J. Anim. Breed. Genet. 2019, 136, 63–73. [Google Scholar]
- Wang, Y.; Bennewitz, J.; Wellmann, R. Novel optimum contribution selection methods accounting for conflicting objectives in breeding programs for livestock breeds with historical migration. Genet. Sel. Evol. 2017, 49, 1–12. [Google Scholar] [CrossRef]
- Eynard, S.E.; Windig, J.J.; Hiemstra, S.J.; Calus, M.P.L. Whole-genome sequence data uncover loss of genetic diversity due to selection. Genet. Sel. Evol. 2016, 48, 1–14. [Google Scholar] [CrossRef]
- Albrechtsen, A.; Nielsen, F.C.; Nielsen, R. Ascertainment Biases in SNP Chips Affect Measures of Population Divergence. Mol. Biol. Evol. 2010, 27, 2534–2547. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
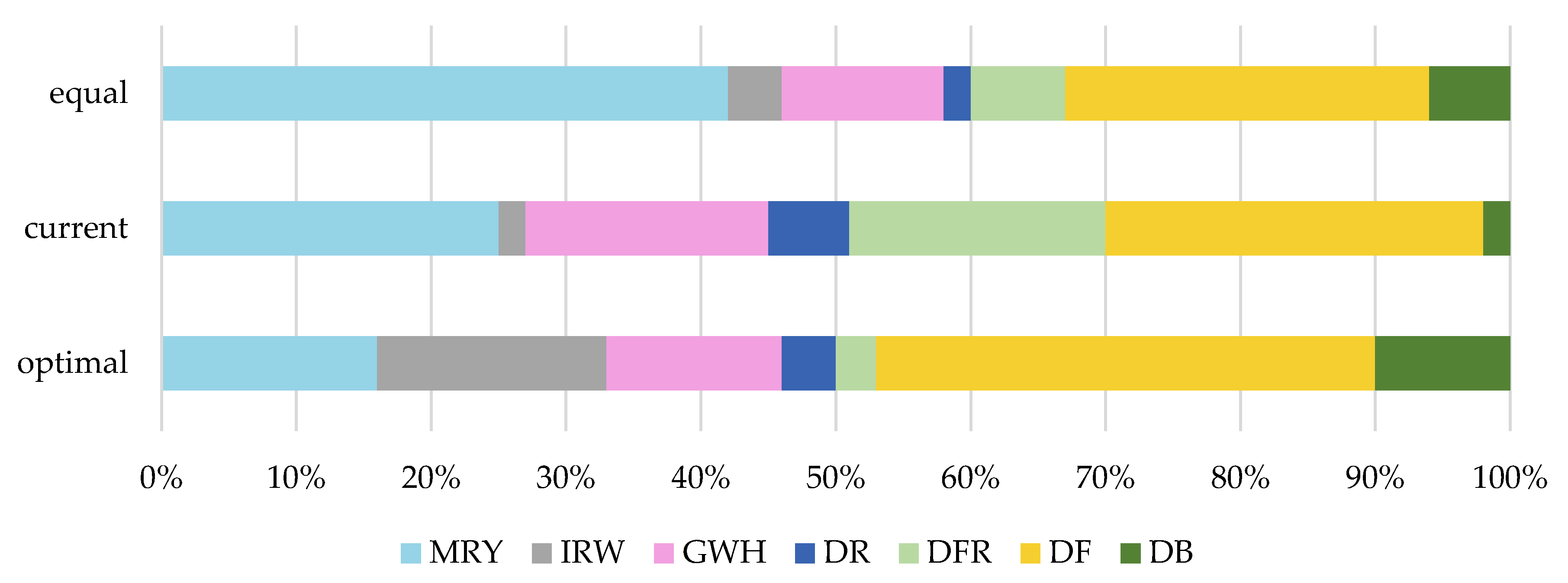{kind=link}
| Breed | DB | DF | DFR | DR | GWH | HF | IRW | MRY |
|---|---|---|---|---|---|---|---|---|
| DB | 66.87 | |||||||
| DF | 63.61 | 66.71 | ||||||
| DFR | 63.58 | 65.51 | 66.47 | |||||
| DR | 63.04 | 62.89 | 63.01 | 65.80 | ||||
| GWH | 62.89 | 62.97 | 63.02 | 62.97 | 69.05 | |||
| HF | 61.91 | 61.82 | 61.89 | 61.83 | 61.91 | 64.24 | ||
| IRW | 63.04 | 63.06 | 63.11 | 63.64 | 63.02 | 61.75 | 64.69 | |
| MRY | 63.11 | 62.78 | 62.92 | 64.76 | 62.93 | 62.02 | 63.67 | 66.44 |
| Breed | Equal | Current | Optimal | Current-Equal | Optimal-Current |
|---|---|---|---|---|---|
| DB | 66.87 | 65.57 | 64.33 | −1.3 | −1.24 |
| DF | 66.71 | 65.92 | 63.36 | −0.79 | −2.56 |
| DFR | 66.47 | 65.24 | 64.77 | −1.23 | −0.47 |
| DR | 65.80 | 64.31 | 63.97 | −1.49 | −0.34 |
| GWH | 69.05 | 67.91 | 66.23 | −1.14 | −1.68 |
| IRW | 64.69 | 64.36 | 61.57 | −0.33 | −2.79 |
| MRY | 66.44 | 66.12 | 64.56 | −0.32 | −1.56 |
| Birth Year | DB | DF | DFR | DR | GWH | IRW | MRY | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | sel | c | n | sel | c | n | sel | c | n | sel | c | n | sel | c | n | sel | c | n | sel | c | |
| 1960–1969 | 24 | 4 | 11.9 | 2 | 1 | 0.6 | 16 | 5 | 13.9 | ||||||||||||
| 1970–1979 | 3 | 1 | 0.8 | 44 | 7 | 10.8 | 6 | 4 | 9.9 | 5 | 1 | 2.6 | 29 | 14 | 28.4 | ||||||
| 1980–1989 | 6 | 6 | 36.9 | 47 | 9 | 29.5 | 2 | 1 | 0.6 | 23 | 6 | 22.8 | 1 | 1 | 1.1 | 20 | 6 | 3.5 | |||
| 1990–1999 | 10 | 9 | 26.1 | 45 | 4 | 16.1 | 28 | 22 | 63.6 | 10 | 2 | 9.6 | 6 | 5 | 57.8 | 64 | 7 | 11.2 | |||
| 2000–2009 | 14 | 11 | 16.2 | 20 | 4 | 31.6 | 8 | 7 | 11.1 | 11 | 11 | 62.7 | 32 | 14 | 41.6 | 13 | 13 | 27.0 | 123 | 14 | 22.5 |
| 2010–2015 | 9 | 6 | 20.0 | 10 | 0 | 0 | 9 | 8 | 14.8 | 6 | 6 | 37.3 | 10 | 7 | 22.7 | 6 | 6 | 14.0 | 51 | 14 | 20.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Breukelen, A.E.; Doekes, H.P.; Windig, J.J.; Oldenbroek, K. Characterization of Genetic Diversity Conserved in the Gene Bank for Dutch Cattle Breeds. Diversity 2019, 11, 229. https://doi.org/10.3390/d11120229
van Breukelen AE, Doekes HP, Windig JJ, Oldenbroek K. Characterization of Genetic Diversity Conserved in the Gene Bank for Dutch Cattle Breeds. Diversity. 2019; 11(12):229. https://doi.org/10.3390/d11120229
Chicago/Turabian Stylevan Breukelen, Anouk E., Harmen P. Doekes, Jack J. Windig, and Kor Oldenbroek. 2019. "Characterization of Genetic Diversity Conserved in the Gene Bank for Dutch Cattle Breeds" Diversity 11, no. 12: 229. https://doi.org/10.3390/d11120229
APA Stylevan Breukelen, A. E., Doekes, H. P., Windig, J. J., & Oldenbroek, K. (2019). Characterization of Genetic Diversity Conserved in the Gene Bank for Dutch Cattle Breeds. Diversity, 11(12), 229. https://doi.org/10.3390/d11120229