DNA Barcoding and Taxonomic Challenges in Describing New Putative Species: Examples from Sootywing and Cloudywing Butterflies (Lepidoptera: Hesperiidae)

Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.1.1. Sootywings
2.1.2. Cloudywings
2.2. DNA Extraction and Sequence Analysis
3. Results and Discussion
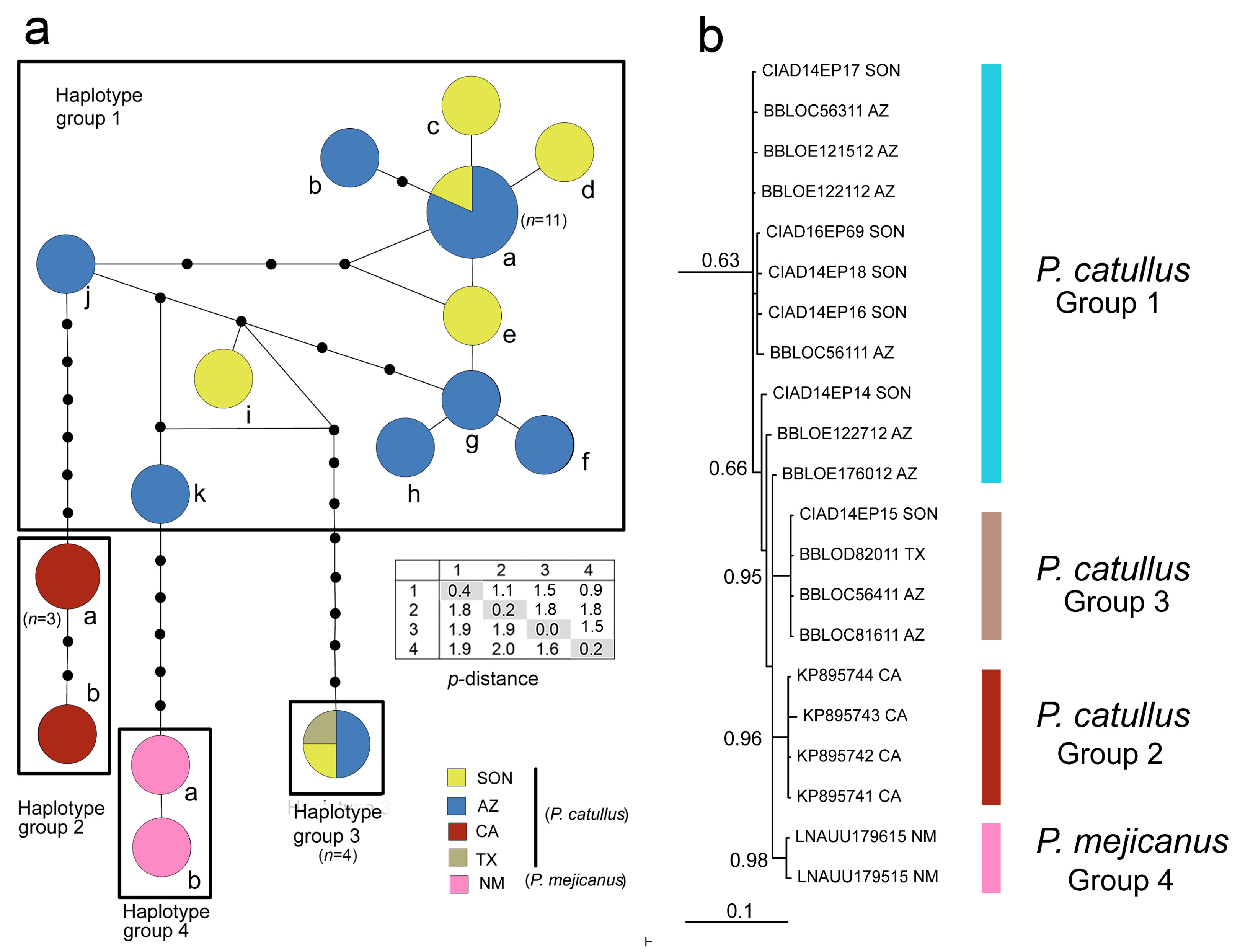
3.1. Sootywings
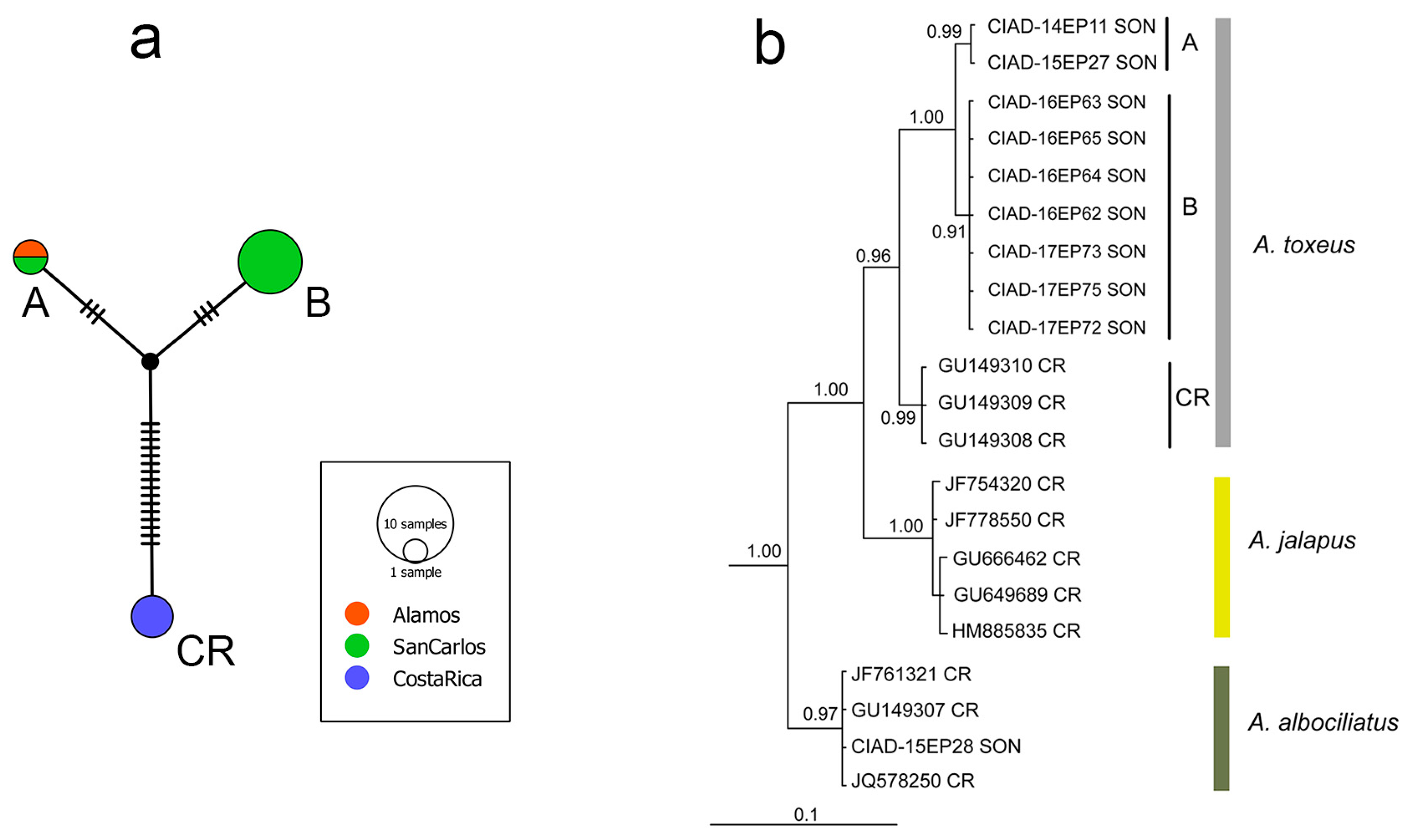
3.2. Cloudywings
4. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Jörger, K.M.; Schrödl, M. How to describe cryptic species? Practical challenges of molecular taxonomy. Front. Zool. 2013, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Sluys, R. The unappreciated, fundamentally analytical nature of taxonomy and the implications for the inventory of biodiversity. Biodivers. Conserv. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Pante, E.; Schoelinck, C.; Puillandre, N. From integrative taxonomy to species description: One step beyond. Syst. Biol. 2015, 64, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Pfeiler, E. Resurrection of the name Albula pacifica (Beebe, 1942) for the shafted bonefish (Albuliformes: Albulidae) from the eastern Pacific. Rev. Biol. Trop. 2008, 56, 839–844. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pfeiler, E.; Vergara-Quintanar, J.E.; Castrezana, S.; Caterino, M.S.; Markow, T.A. Phylogenetic relationships of Sonoran Desert cactus beetles in the tribe Hololeptini (Coleoptera: Histeridae: Histerinae), with comments on the taxonomic status of Iliotona beyeri. Mol. Phylogenet. Evol. 2010, 56, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, A.; Jones, R.W.; Pedraza-Lara, C.; Alvarado Villanueva, O.; Pfeiler, E. Reassessment of the phylogeography and intraspecific relationships of western and eastern populations of the boll weevil, Anthonomus grandis Boheman (Coleoptera: Curculionidae) in North America. Biol. J. Linn. Soc. 2017, 122, 29–45. [Google Scholar] [CrossRef]
- Drew, L.W. Are we losing the science of taxonomy? BioScience 2011, 61, 942–946. [Google Scholar] [CrossRef]
- Cao, X.; Liu, J.; Chen, J.; Zheng, G.; Kuntner, M.; Agnarsson, I. Rapid dissemination of taxonomic discoveries based on DNA barcoding and morphology. Sci. Rep. 2016, 6, 37066. [Google Scholar] [CrossRef] [PubMed]
- Pelham, J.P. A catalogue of the butterflies of the United States and Canada. J. Res. Lepid. 2008, 40, 651–652. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R.; Harris, W.E. An emergent science on the brink of irrelevance: A review of the past 8 years of DNA barcoding. Mol. Ecol. Resour. 2012, 12, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Kvist, S. Barcoding in the dark: A critical view of the sufficiency of zoological DNA barcoding databases and a plea for broader integration of taxonomic knowledge. Mol. Phylogenet. Evol. 2013, 69, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Good, J.M.; Hird, S.; Reid, N.; Demboski, J.R.; Steppan, S.J.; Martin-Nims, T.R.; Sullivan, J. Ancient hybridization and mitochondrial capture between two species of chipmunks. Mol. Ecol. 2008, 17, 1313–1327. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [PubMed]
- Forister, M.L.; Nice, C.C.; Fordyce, J.A.; Gompert, Z.; Shapiro, A.M. Considering evolutionary processes in the use of single-locus genetic data for conservation, with examples from the Lepidoptera. J. Insect Conserv. 2008, 12, 37–51. [Google Scholar] [CrossRef]
- Zakharov, E.V.; Lobo, N.F.; Nowak, C.; Hellmann, J.J. Introgression as a likely cause of mtDNA paraphyly in two allopatric skippers (Lepidoptera: Hesperiidae). Heredity 2009, 102, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Kodandaramaiah, U.; Simonsen, T.J.; Bromilow, S.; Wahlberg, N.; Sperling, F. Deceptive single-locus taxonomy and phylogeography: Wolbachia-associated divergence in mitochondrial DNA is not reflected in morphology and nuclear markers in a butterfly species. Ecol. Evol. 2013, 3, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Cong, Q.; Shen, J.; Borek, D.; Robbins, R.K.; Opler, P.A.; Otwinowski, Z.; Grishin, N.V. When COI barcodes deceive: Complete genomes reveal introgression in hairstreaks. Proc. R. Soc. B 2017, 284, 20161735. [Google Scholar] [CrossRef] [PubMed]
- Hawlitschek, O.; Morinière, J.; Lehmann, G.U.C.; Lehmann, A.W.; Kropf, M.; Dunz, A.; Glaw, F.; Detcharoen, M.; Schmidt, S.; Hausmann, A.; et al. DNA barcoding of crickets, katydids and grasshoppers (Orthoptera) from Central Europe with focus on Austria, Germany and Switzerland. Mol. Ecol. Resour. 2017, 17, 1037–1053. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; deWaard, J.R.; Landry, J.F. DNA barcodes for 1/1000 of the animal kingdom. Biol. Lett. 2010, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.; Akhtar, S.; Rafi, M.A.; Mansoor, S.; Hebert, P.D.N. Mapping global biodiversity connections with DNA barcodes: Lepidoptera of Pakistan. PLoS ONE 2017, 12, e0174749. [Google Scholar] [CrossRef] [PubMed]
- Lavinia, P.D.; Nuñez Bustos, E.O.; Kopuchian, C.; Lijtmaer, D.A.; García, N.C.; Hebert, P.D.N.; Tubaro, P.L. Barcoding the butterflies of southern South America: Species delimitation efficacy, cryptic diversity and geographic patterns of divergence. PLoS ONE 2017, 12, e0186845. [Google Scholar] [CrossRef] [PubMed]
- Brower, A.V.Z. Alleviating the taxonomic impediment of DNA barcoding and setting a bad precedent: names for ten species of ‘Astraptes fulgerator’ (Lepidoptera: Hesperiidae: Eudaminae) with DNA–based diagnoses. Syst. Biodivers. 2010, 8, 485–491. [Google Scholar] [CrossRef]
- Grishin, N.V.; Janzen, D.H.; Hallwachs, W. Hiding behind gaudy looks, a new Central American species of Phareas (Hesperiidae: Eudaminae). J. Lepid. Soc. 2013, 67, 161–174. [Google Scholar]
- Garzón-Orduña, I.J.; Brower, A.V.Z.; Kamilari, M.; Iribar, A.; Murienne, J. Cracking the code: Examination of species delimitations among Hamadryas butterflies with DNA barcodes suggests Caribbean Cracker is Hamadryas februa Hübner (Nymphalidae: Biblidinae). J. Lepid. Soc. 2018, 72, 53–73. [Google Scholar] [CrossRef]
- Pfeiler, E.; Nazario-Yepiz, N.O.; Vargas, C.C.; Laclette, M.R.L. DNA barcodes suggest possible new cryptic species in the Codatractus melon species group (Hesperiidae: Eudaminae) in North America. J. Lepid. Soc. 2018, 72, 203–211. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Chen, X.; Murphy, R.W. Assessing DNA barcoding as a tool for species identification and data quality control. PLoS ONE 2013, 8, e57125. [Google Scholar] [CrossRef] [PubMed]
- Pfeiler, E.; Nazario-Yepiz, N.O.; Pérez-Gálvez, F.; ChávezMora, C.A.; Laclette, M.R.L.; Rendón-Salinas, E.; Markow, T.A. Population genetics of overwintering monarch butterflies, Danaus plexippus (Linnaeus), from central Mexico inferred from mitochondrial DNA and microsatellite markers. J. Hered. 2017, 108, 163–175. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pratt, G.F.; Rugman-Jones, P.F.; Stouthamer, R. Genetic characterization of North American sootywings (Lepidoptera: Hesperidae [sic]) using nuclear and mitochondrial loci validates specific status of MacNeill’s sootywing, Hesperopsis gracielae (MacNeill). Proc. Entomol. Soc. Wash. 2015, 117, 194–202. [Google Scholar] [CrossRef]
- Warren, A.D.; Davis, K.J.; Stangeland, E.M.; Pelham, J.P.; Grishin, N.V. [21-XI-2017]. 2013. Available online: http://www.butterfliesofamerica.com/ (accessed on 7 July 2018).
- Pyle, R.M. The Butterflies of Cascadia; Seattle Audubon Society: Seattle, WA, USA, 2002; pp. 1–420. ISBN 0-914516-13-2. [Google Scholar]
- Pfeiler, E.; Laclette, M.R.L.; Markow, T.A. Butterfly biodiversity in a threatened coastal desert ecosystem of northwestern Mexico, with a focus on the life history and ecology of potentially endangered species. J. Lepid. Soc. 2016, 70, 47–60. [Google Scholar] [CrossRef]
- Bailowitz, R.; Brock, J. Danforth, D. Annotated checklist of the butterflies (Lepidoptera) of Sonora, Mexico. Dugesiana 2017, 24, 125–147. [Google Scholar]
- Felger, R.S.; Carnahan, S.D.; Sánchez-Escalante, J.J. Oasis at the Desert Edge: Flora of Cañón del Nacapule. Sonora, Mexico, 2017; pp. 1–220. Available online: http://www.tumamoc.arizona.edu (accessed on 31 July 2018).
- Brown, J.W.; Real, H.G.; Faulkner, D.K. Butterflies of Baja California; Lepidoptera Research Foundation, Inc.: Beverly Hills, CA, USA, 1992; pp. 1–129. ISBN 9611464-4-3. [Google Scholar]
- Lotts, K.; Naberhaus, T. Butterflies and Moths of North America. 2017. Available online: http://www.butterfliesandmoths.org/ (accessed on 23 April 2018).
- Lindsey, A.W.; Bell, E.L.; Williams, R.C., Jr. The Hesperioidea of North America. J. Sci. Lab. Denison. 1931, 26, 141–142. [Google Scholar]
- Glassberg, J. Butterflies through Binoculars: The West; Oxford University Press: New York, NY, USA, 2001; ISBN 0-19-510669-5. [Google Scholar]
- Janzen, D.H.; Hallwachs, W.; Burns, J.M.; Hajibabaei, M.; Bertrand, C.; Hebert, P.D.N. Reading the complex skipper butterfly fauna of one tropical place. PLoS ONE 2011, 6, e19874. [Google Scholar] [CrossRef] [PubMed]
- Pfeiler, E.; Laclette, M.R.L.; Markow, T.A. Polyphyly in Urbanus and Astraptes (Hesperiidae: Eudaminae) assessed using mitochondrial DNA barcodes, with a reinstated status proposed for Achalarus. J. Lepid. Soc. 2016, 70, 85–95. [Google Scholar]
- Williams, R.C., Jr.; Bell, E.L. Studies in the American Hesperioidea. (Lepidoptera). Paper IV (Lepidoptera). On the synonymy and the male genitalia of some species. Trans. Am. Entomol. Soc. 1934, 60, 265–280. [Google Scholar]
- Janzen, D.H.; Hallwachs, W. Dynamic Database for An Inventory of the Macrocaterpillar Fauna, and its Food Plants and Parasitoids, of Area de Conservación Guanacaste (ACG), Northwestern Costa Rica (SRNP voucher codes). 2009. Available online: http://janzen.sas.upenn.edu (accessed on 18 August 2018).
- SEINet. Southwest Environmental Information Network. Available online: http://swbiodiversity.org/seinet/index.php (accessed on 31 July 2018).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Servín-Garcidueñas, L.E.; Martínez-Romero, E. Complete mitochondrial genome recovered from the gut metagenome of overwintering monarch butterflies, Danaus plexippus (L.) (Lepidoptera: Nymphalidae, Danainae). Mitochondr. DNA 2014, 25, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Scott, J.A.; Ellis, S.L.; Eff, D. New records, range extensions, and field data for Colorado butterflies and skippers. J. Lepid. Soc. 1968, 22, 159–171. [Google Scholar]
- Scott, J.A.; Davenport, K.E. Weeds can also be stars: A new subspecies of Pholisora catullus (Hesperiidae). Papilio (New Series) 2017, 26, 2–5. [Google Scholar]
- Puillandre, N.; Macpherson, E.; Lambourdière, J.; Cruaud, C.; Boisselier-Dubayle, M.C.; Samadi, S. Barcoding type specimens helps to identify synonyms and an unnamed new species in Eumunida Smith, 1883 (Decapoda: Eumunididae). Invertebr. Syst. 2011, 25, 322–333. [Google Scholar] [CrossRef]
- Hausmann, A.; Miller, S.E.; Holloway, J.D.; deWaard, J.R.; Pollock, D.; Prosser, S.W.J.; Hebert, P.D.N. Calibrating the taxonomy of a megadiverse insect family: 3000 DNA barcodes from geometrid type specimens (Lepidoptera, Geometridae). Genome 2016, 59, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Warren, A.D. Butterflies of Oregon Their Taxonomy, Distribution, and Biology; C.P. Gillette Museum of Arthropod Diversity: Fort Collins, CO, USA, 2005; ISBN 1084-8819. [Google Scholar]
- Plötz, C. Einige Hesperiinen-Gattungen und deren Arten. Berliner Entomologische Zeitschrift 1882, 26, 253–266. [Google Scholar]
- Mabille, P.; Boullet, E. Essai de Révision de la Famille des Hespérides. Ann. Sci. Nat. Zool. 1912, 16, 158–159. [Google Scholar]
- Skinner, H. A new species of Eudamus. Can. Entomol. 1892, 24, 164. [Google Scholar] [CrossRef]
- Skinner, H.; Williams, R.C., Jr. On the male genitalia of the larger Hesperiidae of North America. Trans. Am. Entomol Soc. 1922, 48, 109–127. [Google Scholar]
- Warren, A.D. Florida Museum of Natural History, University of Florida, Gainesville, FL, USA. Personal communication. 2017. [Google Scholar]
- Pfenninger, M.; Schwenk, K. Cryptic animal species are homogeneously distributed among taxa and biogeographical regions. BMC Evol. Biol. 2007, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Karanovic, T.; Djurakic, M.; Eberhard, S.M. Cryptic species or inadequate taxonomy? Implementation of 2D geometric morphometrics based on integumental organs as landmarks for delimitation and description of copepod taxa. Syst. Biol. 2016, 65, 304–327. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Locality | GenBank | BOLD | Specimen ID | Date Collected | Hap | bp |
|---|---|---|---|---|---|---|
| Pholisora catullus | ||||||
| MEX; Sonora; San Carlos | MH310828 | CIAD 14–EP12 | 02 Oct. 2014 | 1a | 658 | |
| MEX; Sonora; San Carlos | MH310832 | CIAD 14–EP14 | 02 Oct. 2014 | 1i | 658 | |
| MEX; Sonora; San Carlos | MH310833 | CIAD 14–EP15 | 24 Sep. 2014 | 3 | 658 | |
| MEX; Sonora; San Carlos | MH310830 | CIAD 14–EP16 | 24 Sep. 2014 | 1c | 658 | |
| MEX; Sonora; San Carlos | MH310831 | CIAD 14–EP17 | 06 Jul. 2014 | 1e | 658 | |
| MEX; Sonora; San Carlos | MH310829 | CIAD 14–EP18 | 10 Jul. 2014 | 1d | 658 | |
| MEX; Sonora; La Aduana | MH310827 | CIAD 16–EP69 | 02 Aug. 2016 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOC352-11 | BIOUG01455-F12 | 11 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOC561-11 | BIOUG01457-H07 | 11 Aug. 2011 | 1b | 658 | |
| USA; AZ; Pima Co. | BBLOC563-11 | BIOUG01457-H09 | 11 Aug. 2011 | 1f | 658 | |
| USA; AZ; Pima Co. | BBLOC562-11 | BIOUG01457-H08 | 11 Aug. 2011 | ‘1’ | 621a | |
| USA; AZ; Pima Co. | BBLOC564-11 | BIOUG01457-H10 | 11 Aug. 2011 | 3 | 658 | |
| USA; AZ; Pima Co. | BBLOC816-11 | BIOUG01466-D02 | 11 Aug. 2011 | 3 | 658 | |
| USA; AZ; Pima Co. | BBLOE1210-12 | BIOUG01985-A02 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1211-12 | BIOUG01985-A03 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1212-12 | BIOUG01985-A04 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1215-12 | BIOUG01985-A07 | 15 Aug. 2011 | 1h | 658 | |
| USA; AZ; Pima Co. | BBLOE1217-12 | BIOUG01985-A09 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1221-12 | BIOUG01985-B01 | 15 Aug. 2011 | 1g | 658 | |
| USA; AZ; Pima Co. | BBLOE1223-12 | BIOUG01985-B03 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1225-12 | BIOUG01985-B05 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1226-12 | BIOUG01985-B06 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1227-12 | BIOUG01985-B07 | 15 Aug. 2011 | 1j | 658 | |
| USA; AZ; Pima Co. | BBLOE1228-12 | BIOUG01985-B08 | 15 Aug. 2011 | 1a | 658 | |
| USA; AZ; Pima Co. | BBLOE1760-12 | BIOUG01990-G05 | 15 Aug. 2011 | 1k | 658 | |
| USA; CA; San Diego Co.b | KP895741 | 11 Apr. 2008 | 2a | 657 | ||
| USA; CA; San Diego Co.b | KP895742 | 11 Apr. 2008 | 2a | 657 | ||
| USA; CA; San Diego Co.b | KP895743 | GBMIN81625-17 | 11 Apr. 2008 | 2b | 657 | |
| USA; CA; San Diego Co.b | KP895744 | GBMIN81626-17 | 11 Apr. 2008 | 2a | 657 | |
| USA; CA; San Diego Co. | LOCBF4342-15 | BIOUG23748-F06 | 07 Apr. 2013 | ‘2’ | 588 a | |
| USA; CA; San Diego Co. | LOCBF1343-13 | BIOUG06805-H01 | 07 Apr. 2013 | ‘2’ | 624 a | |
| USA; CA; San Diego Co. | LOCBF1344-13 | BIOUG06805-H02 | 07 Apr. 2013 | ‘2’ | 625 a | |
| USA; CA; San Diego Co. | LOCBF1345-13 | BIOUG06805-H03 | 07 Apr. 2013 | ‘2’ | 625 a | |
| USA; TX; Randall Co. | BBLOD820-11 | BIOUG01568-D06 | 09 July 2011 | 3 | 658 | |
| Pholisora mejicanus | ||||||
| USA; NM; Colfax Co. | LNAUU1796-15 | USNM_ENT 01201719 | 07 Aug. 1989 | 4a | 658 | |
| USA; NM; Colfax Co. | LNAUU1795-15 | USNM_ENT 01201718 | 07 Aug. 1989 | 4b | 658 | |
| USA; NM; Torrance Co. | NGSFT3796-16 | USNM_ENT 01201720 | 10 Aug. 1946 | ‘4’ | 658 a | |
| USA; CO: Chaffee Co. | NGSFT3797-16 | USNM_ENT 01201721 | No date | ‘4’ | 208 a | |
| Achalarus toxeus | ||||||
| MEX; Sonora; San Carlos | KT290038 | CIAD 14–EP11 | 08 Sep. 2014 | A | 658 | |
| MEX; Sonora; Alamos | KU041804 | CIAD 15–EP27 | 03 Aug. 2015 | A | 658 | |
| MEX; Sonora; San Carlos | MH807206 | CIAD 16–EP62 | 28 Mar. 2016 | B | 658 | |
| MEX; Sonora; San Carlos | MH807207 | CIAD 16–EP63 | 04 Apr. 2016 | B | 658 | |
| MEX; Sonora; San Carlos | MH807208 | CIAD 16–EP64 | 10 Apr. 2016 | B | 658 | |
| MEX; Sonora; San Carlos | MH807209 | CIAD 16–EP65 | 17 Apr. 2016 | B | 658 | |
| MEX; Sonora; San Carlos | MH807210 | CIAD 17–EP72 | 06 Apr. 2017 | B | 658 | |
| MEX; Sonora; San Carlos | MH807211 | CIAD 17–EP73 | 08 Apr. 2017 | B | 658 | |
| MEX; Sonora; San Carlos | MH807212 | CIAD 17–EP75 | 14 Apr. 2017 | B | 658 | |
| Costa Rica; ACG | GU149310 | 03-SRNP-237 | 20 Jan. 2003 c | CR | 658 | |
| Costa Rica; ACG | GU149309 | 02-SRNP-10179 | 31 Mar. 2002 | CR | 658 | |
| Costa Rica; ACG | GU149308 | 02-SRNP-10180 | 19 Apr. 2002 | CR | 658 | |
| Costa Rica; ACG | DQ291763 | 96-SRNP-8690 | 09 Oct. 1996 | CR | 568 | |
| Costa Rica; ACG | DQ291762 | 93-SRNP-7539 | 14 Dec. 1993 | CR | 393 | |
| Achalarus albociliatus | ||||||
| MEX; Sonora; Alamos | KU041805 | CIAD 15–EP28 | 04 Aug. 2015 | 658 | ||
| Costa Rica; ACG | JF761321 | 07-SRNP-12984 | 08 Jun. 2007 | 658 | ||
| Costa Rica; ACG | GU149307 | 02-SRNP-4786 | 15 Apr. 2002 | 658 | ||
| Costa Rica; ACG | JQ578250 | 06-SRNP-101470 | 07 Apr. 2006 | 658 | ||
| Achalarus jalapus | ||||||
| Costa Rica; ACG | GU666462 | 08-SRNP-22900 | 26 Sep. 2008 | 658 | ||
| Costa Rica; ACG | GU649689 | 08-SRNP-23350 | 05 Nov. 2008 | 658 | ||
| Costa Rica; ACG | HM885835 | 09-SRNP-20140 | 02 Mar. 2009 | 658 | ||
| Costa Rica; ACG | JF754320 | 08-SRNP-30979 | 20 May 2008 | 658 | ||
| Costa Rica; ACG | JF778550 | 08-SRNP-22673 | 22 Sep. 2008 | 658 |
| Haplotype Group/Species | Nucleotide Site No. | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 43 | 58 | 271 | 355 | 367 | 418 | 448 | 542 | 634 | |
| 1 P. catullus (n = 22) | A | T | C | T | C | T | T | C | C |
| 2 P. catullus (n = 8) | A | C | C | C | C | C | T | C | T |
| 3 P. catullus (n = 4) | A | C | T | T | T | C | C | T | C |
| 4 P. mejicanus (n = 3) | G | C | T | T | T | T | T | C | C |
| Haplotype | Nucleotide Site No. | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 46 | 79 | 205 | 212 | 217 | 277 | 283 | 313 | 407 | 415 | 421 | 472 | 490 | 508 | 574 | 628 | |
| Son A (n = 2) | C | A | T | C | A | C | T | C | C | G | C | C | C | T | C | G |
| Son B (n = 7) | C | A | T | C | A | C | T | C | C | G | C | C | C | T | C | G |
| CR (n = 3) | T | T | C | T | G | T | C | T | T | A | T | T | T | C | T | A |
| CR/DQ291763 * | T | T | C | T | G | T | C | T | T | A | T | T | T | C | – | – |
| CR/DQ291762 * | – | – | – | – | – | T | C | T | T | A | T | T | T | C | T | A |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeiler, E. DNA Barcoding and Taxonomic Challenges in Describing New Putative Species: Examples from Sootywing and Cloudywing Butterflies (Lepidoptera: Hesperiidae). Diversity 2018, 10, 111. https://doi.org/10.3390/d10040111
Pfeiler E. DNA Barcoding and Taxonomic Challenges in Describing New Putative Species: Examples from Sootywing and Cloudywing Butterflies (Lepidoptera: Hesperiidae). Diversity. 2018; 10(4):111. https://doi.org/10.3390/d10040111
Chicago/Turabian StylePfeiler, Edward. 2018. "DNA Barcoding and Taxonomic Challenges in Describing New Putative Species: Examples from Sootywing and Cloudywing Butterflies (Lepidoptera: Hesperiidae)" Diversity 10, no. 4: 111. https://doi.org/10.3390/d10040111
APA StylePfeiler, E. (2018). DNA Barcoding and Taxonomic Challenges in Describing New Putative Species: Examples from Sootywing and Cloudywing Butterflies (Lepidoptera: Hesperiidae). Diversity, 10(4), 111. https://doi.org/10.3390/d10040111