The Variable Influences of Sea Level, Sedimentation and Exposure on Holocene Reef Development over a Cross-Shelf Transect, Central Great Barrier Reef

 , ,
, ,
Abstract
1. Introduction
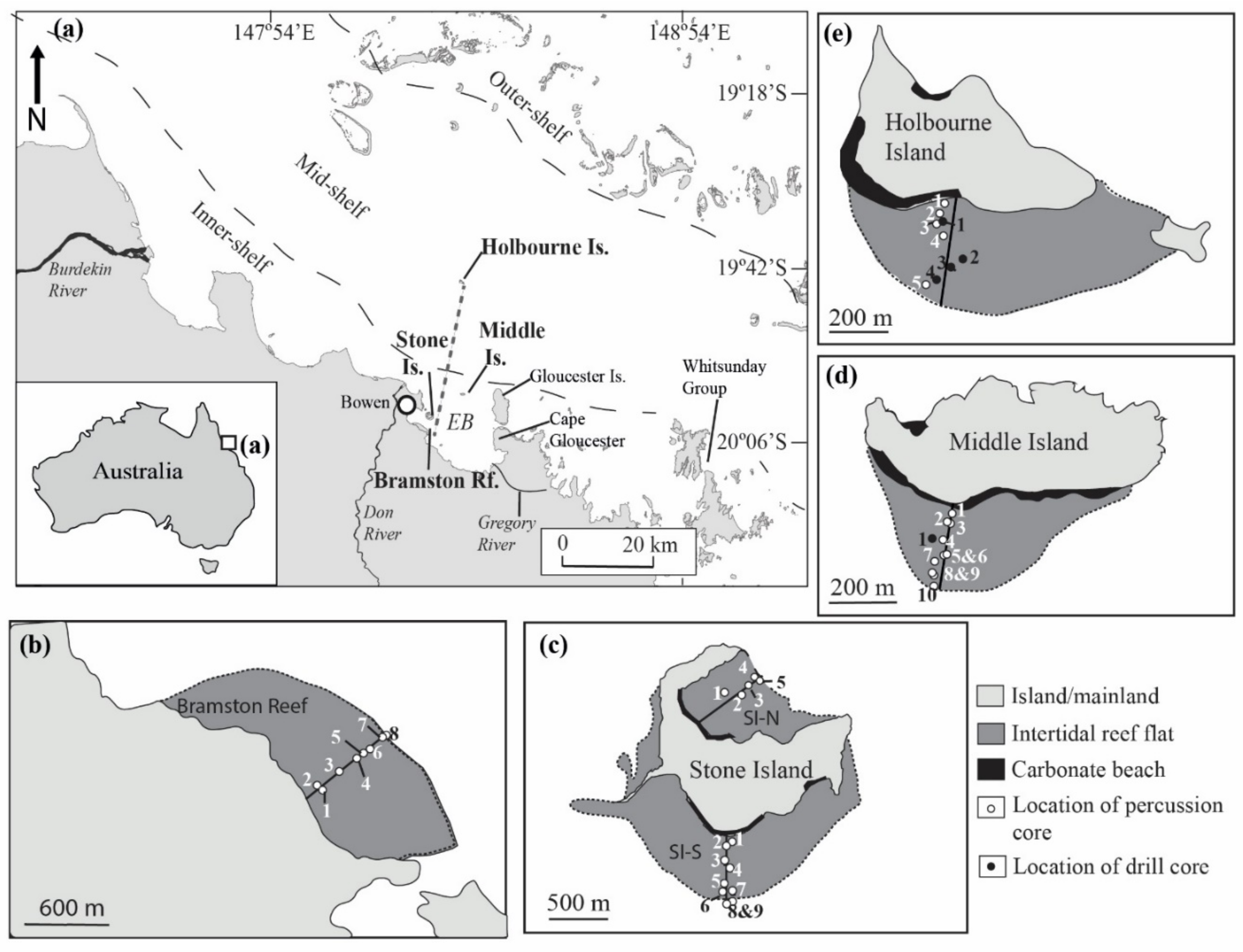
2. Materials and Methods
3. Results
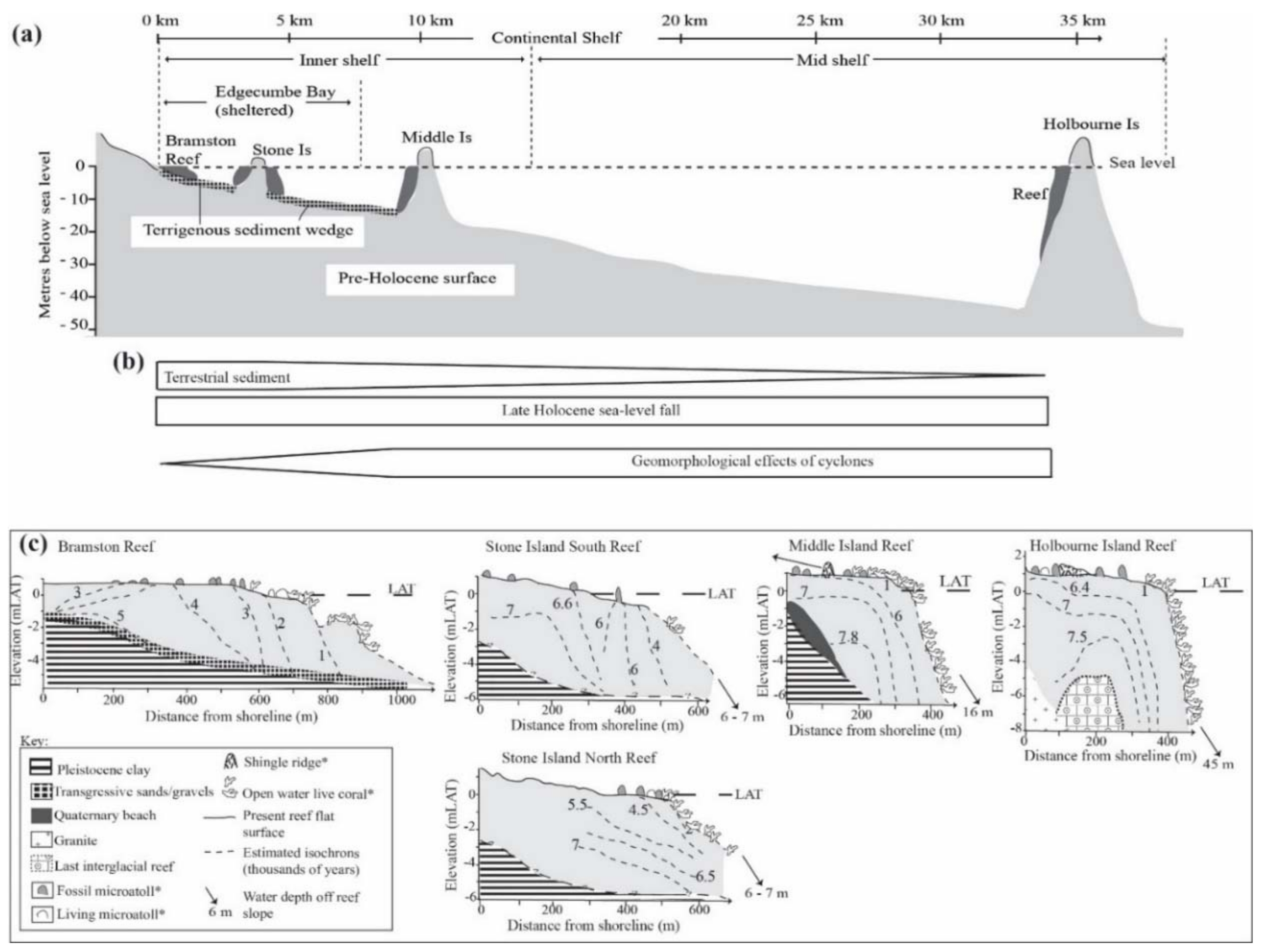
3.1. Nature and Depth of the Antecedent Surface
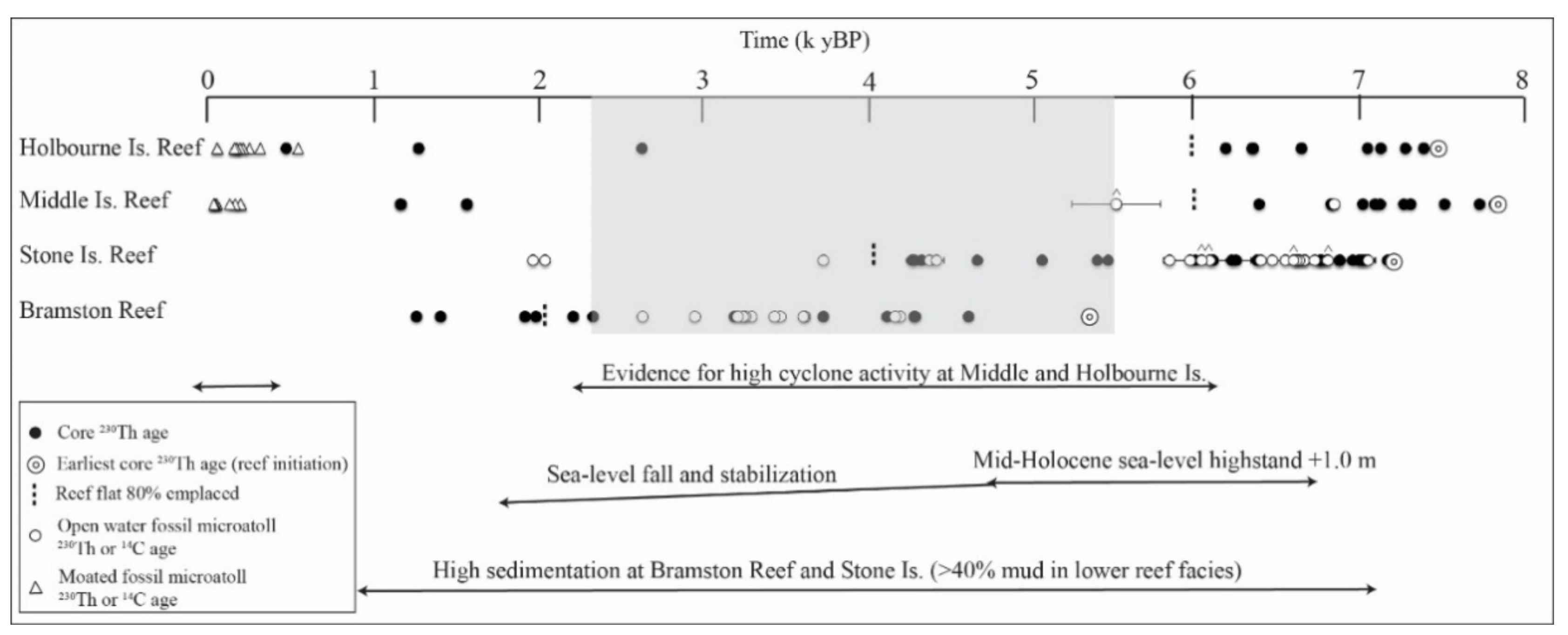
3.2. Timing, Formation, and Style of Reef Growth
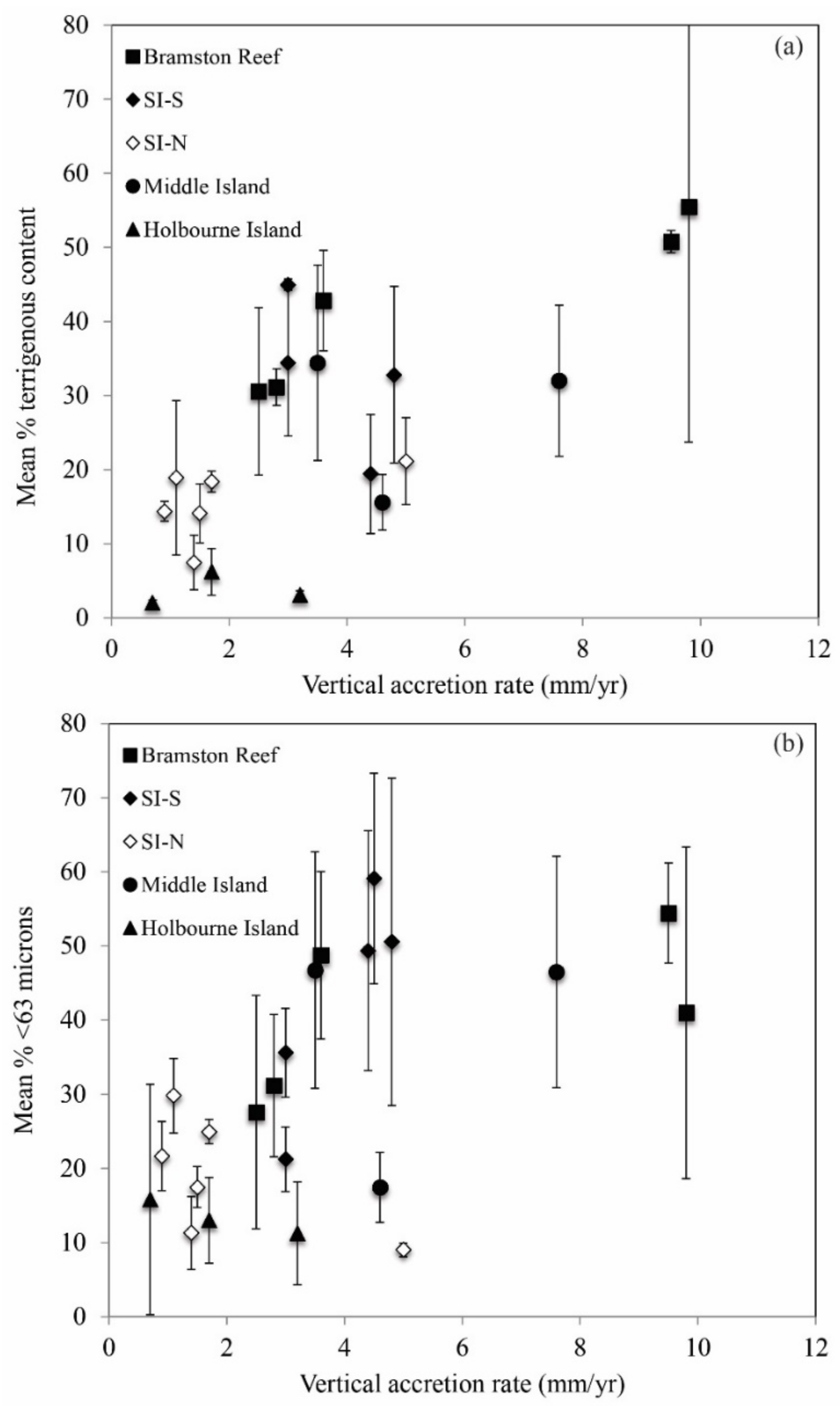
3.3. Reef Accretion Rates and Sedimentology
4. Discussion
4.1. Reef Foundations
4.2. Controls on Reef Initiation across the Shelf
4.3. Role of Exposure Conditions and Terrigenous Sediments on Reef Development and Preservation
4.4. Evidence for Changes in Disturbance Regimes since European Settlement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Done, T.J. Effects of tropical cyclone waves on ecological and geomorphological structures on the Great Barrier Reef. Cont. Shelf. Res. 1992, 12, 859–872. [Google Scholar] [CrossRef]
- Risk, M.J. Assessing the effects of sediments and nutrients on coral reefs. Curr. Opin. Environ. Sustain. 2014, 7, 108–117. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; et al. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629. [Google Scholar] [CrossRef] [PubMed]
- Pastorok, R.; Bilyard, G. Effects of sewage pollution on coral-reef communities. Mar. Ecol. Prog. Ser. 1985, 21, 175–189. [Google Scholar] [CrossRef]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 1–5. [Google Scholar] [CrossRef] [PubMed]
- DeVantier, L.M.; De’ath, G.; Turak, E.; Done, T.J.; Fabricius, K.E. Species richness and community structure of reef-building corals on the nearshore Great Barrier Reef. Coral Reefs 2006, 25, 329–340. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018. [Google Scholar] [CrossRef] [PubMed]
- Pandolfi, J.M.; Jackson, J.B.C. Ecological persistence interrupted in Caribbean coral reefs. Ecol. Lett. 2006, 9, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Smithers, S.G.; Hopley, D.; Parnell, K.E. Fringing and nearshore coral reefs of the Great Barrier Reef: Episodic Holocene development and future prospects. J. Coast. Res. 2006, 221, 175–187. [Google Scholar] [CrossRef]
- Bruno, J.F.; Precht, W.F.; Vroom, P.S.; Aronson, R.B. Coral reef baselines: How much macroalgae is natural? Mar. Pollut. Bull. 2014, 80, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Hongo, C.; Lin, K.; Shen, C.C. Late Holocene reef ecosystem baseline: Field evidence from the raised reef terraces of Kodakara and Kikai Islands, Ryukyu Islands, Japan. Quat. Int. 2017, 455, 8–17. [Google Scholar] [CrossRef]
- Kennedy, D.M.; Woodroffe, C.D. Fringing reef growth and morphology: A review. Earth-Sci. Rev. 2002, 57, 255–277. [Google Scholar] [CrossRef]
- Montaggioni, L.F. History of Indo-Pacific coral reef systems since the last glaciation: Development patterns and controlling factors. Earth-Sci. Rev. 2005, 71, 1–75. [Google Scholar] [CrossRef]
- Davies, P.J. Subsurface solution unconformities at Heron Island, Great Barrier Reef. In Proceedings of the 2nd International Coral Reef Symposium, Brisbane, Australia, 22 June–2 July 1974; pp. 573–578. [Google Scholar]
- Hopley, D.; Mclean, R.F.; Marshall, J.; Smith, A.S. Holocene-Pleistocene boundary in a fringing reef: Hayman Island, North Queensland. Search 1978, 9, 323–325. [Google Scholar]
- Davies, P.J.; Marshall, J.F.; Hopley, D. Relationships between reef growth and sea level in the Great Barrier Reef. In Proceedings of the 5th International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; pp. 95–103. [Google Scholar]
- Marshall, J.F. Cross-shelf and facies related variations in submarine cementation in the central Great Barrier Reef. In Proceedings of the 5th International Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; pp. 221–226. [Google Scholar]
- Johnson, D.; Cuff, C.; Rhodes, E. Holocene reef sequences and geochemistry, Britomart reef, central Great Barrier Reef, Australia. Sedimentology 1984, 31, 515–529. [Google Scholar] [CrossRef]
- Partain, B.R.; Hopley, D. Morphology and development of the Cape Tribulation fringing reefs, Great Barrier Reef, Australia; Great Barrier Reef Marine Park Authority Technical Memorandum: Townsville, Australia, 1989. [Google Scholar]
- Johnson, D.P.; Risk, M.J. Fringing reef growth on a terrigenous mud formation, Fantome Island, central Great Barrier Reef, Australia. Sedimentology 1987, 34, 275–287. [Google Scholar] [CrossRef]
- Lewis, S.E.; Sloss, C.R.; Murray-Wallace, C.V.; Woodroffe, C.D.; Smithers, S.G. Post-glacial sea-level changes around the Australian margin: A review. Quat. Sci. Rev. 2013, 74, 115–138. [Google Scholar] [CrossRef]
- Larcombe, P.; Carter, R.M.; Dye, J.; Gagan, M.K.; Johnson, D.P. New evidence for episodic post-glacial sea-level rise, central Great Barrier Reef, Australia. Mar. Geol. 1995, 127, 1–44. [Google Scholar] [CrossRef]
- Chappell, J. Evidence for smoothly falling sea level relative to north Queensland, Australia, during the past 6,000 yr. Nature 1983, 302, 406–408. [Google Scholar] [CrossRef]
- Lewis, S.E.; Wüst, R.A.J.; Webster, J.M.; Collins, J.; Wright, S.A.; Jacobsen, G. Rapid relative sea-level fall along north-eastern Australia between 1200 and 800cal.yrBP: An appraisal of the oyster evidence. Mar. Geol. 2015, 370, 20–30. [Google Scholar] [CrossRef]
- Lewis, S.E.; Wüst, R.A.; Webster, J.M.; Shields, G.A. Mid-late Holocene sea-level variability in eastern Australia. Terra Nova 2008, 20, 74–81. [Google Scholar] [CrossRef]
- Leonard, N.D.; Zhao, J.X.; Welsh, K.J.; Feng, Y.X.; Smithers, S.G.; Pandolfi, J.M.; Clark, T.R. Holocene sea level instability in the southern Great Barrier Reef, Australia: High-precision U-Th dating of fossil microatolls. Coral Reefs 2016, 35, 625–639. [Google Scholar] [CrossRef]
- Perry, C.T.; Smithers, S.G. Cycles of coral reef “turn-on”, rapid growth and “turn-off” over the past 8500 years: A context for understanding modern ecological states and trajectories. Glob. Chang. Biol. 2011, 17, 76–86. [Google Scholar] [CrossRef]
- Perry, C.T.; Smithers, S.G.; Gulliver, P.; Browne, N.K. Evidence of very rapid reef accretion and reef growth under high turbidity and terrigenous sedimentation. Geology 2012, 40, 719–722. [Google Scholar] [CrossRef]
- Roff, G.; Clark, T.R.; Reymond, C.E.; Zhao, J.; Feng, Y.; McCook, L.J.; Done, T.J.; Pandolfi, J.M. Palaeoecological evidence of a historical collapse of corals at Pelorus Island, inshore Great Barrier Reef, following European settlement. Proc. R. Soc. B Biol. Sci. 2012, 280, 20122100. [Google Scholar] [CrossRef] [PubMed]
- Browne, N.K.; Smithers, S.G.; Perry, C.T. Coral reefs of the turbid inner-shelf of the Great Barrier Reef, Australia: An environmental and geomorphic perspective on their occurrence, composition and growth. Earth-Sci. Rev. 2012, 115, 1–20. [Google Scholar] [CrossRef]
- Kroon, F.J.; Kuhnert, P.M.; Henderson, B.L.; Wilkinson, S.N.; Kinsey-Henderson, A.; Abbott, B.; Brodie, J.E.; Turner, R.D.R. River loads of suspended solids, nitrogen, phosphorus and herbicides delivered to the Great Barrier Reef lagoon. Mar. Pollut. Bull. 2012, 65, 167–181. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, M.; Fallon, S.; Wyndham, T.; Hendy, E.; Lough, J.M.; Barnes, D. Coral record of increased sediment flux to the inner Great Barrier Reef since European settlement. Nature 2003, 421, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.; Carroll, C.; Ellis, R.; Hateley, L.; McCloskey, G.L.; Packett, R.; Dougall, C. Fentie Modelling Reductions of Pollutant Loads Due to Improved Management Practices in the Great Barrier Reef Catchments—Whole of GBR; Technical Report; Queensland Government: Brisbane, Australia, 2014.
- Fabricius, K.; De’ath, G.; McCook, L.; Turak, E.; Williams, D.M. Changes in algal, coral and fish assemblages along water quality gradients on the inshore Great Barrier Reef. Mar. Pollut. Bull. 2005, 51, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Larcombe, P.; Woolfe, K.J. Terrigenous sediments as influence upon Holocene nearshore coral reefs, central Great Barrier Reef, Australia. Aust. J. Earth Sci. 1999, 46, 141–154. [Google Scholar] [CrossRef]
- Browne, N.K.; Smithers, S.G.; Perry, C.T. Geomorphology and community structure of Middle Reef, central Great Barrier Reef, Australia: An inner-shelf turbid zone reef subject to episodic mortality events. Coral Reefs 2010, 29, 683–689. [Google Scholar] [CrossRef]
- Morgan, K.M.; Perry, C.T.; Johnson, J.A.; Smithers, S.G. Nearshore turbid-zone corals exhibit high bleaching tolerance on the Great Barrier Reef following the 2016 ocean warming event. Front. Mar. Sci. 2017, 4, 224. [Google Scholar] [CrossRef]
- Ryan, E.J.; Smithers, S.G.; Lewis, S.E.; Clark, T.R.; Zhao, J.X. Chronostratigraphy of Bramston Reef reveals a long-term record of fringing reef growth under muddy conditions in the central Great Barrier Reef. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 734–747. [Google Scholar] [CrossRef]
- Palmer, S.E.; Perry, C.T.; Smithers, S.G.; Gulliver, P. Internal structure and accretionary history of a nearshore, turbid-zone coral reef: Paluma Shoals, central Great Barrier Reef, Australia. Mar. Geol. 2010, 276, 14–29. [Google Scholar] [CrossRef]
- Roff, G.; Zhao, J.X.; Pandolfi, J.M. Rapid accretion of inshore reef slopes from the central Great Barrier Reef during the late Holocene. Geology 2015, 43, 343–346. [Google Scholar] [CrossRef]
- Kleypas, J.A.; Hopley, D. Reef development across a broad continental shelf, Southern Great Barrier Reef, Australia. In Proceedings of the 7th International Coral Reef Symposium, Guam, Micronesia, 22–27 June 1992; pp. 1129–1141. [Google Scholar]
- Ryan, E.J. Fringing Reef Growth on the Central Great Barrier Reef: Signatures of Sea-Level Change, Storms and Sedimentation. Unpublished Ph.D. Thesis, James Cook University, Townsville, Australia, 2016. [Google Scholar]
- Ryan, E.J.; Smithers, S.G.; Lewis, S.E.; Clark, T.R.; Zhao, J.X. The influence of sea level and cyclones on Holocene reef flat development: Middle Island, central Great Barrier Reef. Coral Reefs 2016, 35, 805–818. [Google Scholar] [CrossRef]
- Ryan, E.J.; Lewis, S.E.; Smithers, S.G.; Clark, T.R.; Zhao, J.X. Multi-scale records of reef development and condition provide context for contemporary changes on inshore reefs. Glob. Planet. Chang. 2016, 146, 162–178. [Google Scholar] [CrossRef]
- Ryan, E.J.; Smithers, S.G.; Lewis, S.E.; Clark, T.R.; Zhao, J.X.; Hua, Q. Fringing reef growth over a shallow last interglacial reef foundation at a mid-shelf high island: Holbourne Island, central Great Barrier Reef. Mar. Geol. 2018, 398, 137–150. [Google Scholar] [CrossRef]
- Larcombe, P.; Costen, A.; Woolfe, K.J. The hydrodynamic and sedimentary setting of nearshore coral reefs, Central Great Barrier Reef shelf, Australia: Paluma Shoals, a case study. Sedimentology 2001, 48, 811–835. [Google Scholar] [CrossRef]
- Lambeck, K.; Nakada, M. Late Pleistocene and Holocene sea-level change along the Australian coast. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1990, 89, 143–176. [Google Scholar] [CrossRef]
- Scoffin, T.P.; Stoddart, D.R. The nature and significance of microatolls. Philos. Trans. R. Soc. Lond. B 1978, 284, 99–122. [Google Scholar] [CrossRef]
- Clark, T.R.; Roff, G.; Zhao, J.X.; Feng, Y.X.; Done, T.J.; Pandolfi, J.M. Testing the precision and accuracy of the U-Th chronometer for dating coral mortality events in the last 100 years. Quat. Geochronol. 2014, 23, 35–45. [Google Scholar] [CrossRef]
- Clark, T.R.; Zhao, J.X.; Roff, G.; Feng, Y.X.; Done, T.J.; Nothdurft, L.D.; Pandolfi, J.M. Discerning the timing and cause of historical mortality events in modern Porites from the Great Barrier Reef. Geochim. Cosmochim. Acta 2014, 138, 57–80. [Google Scholar] [CrossRef]
- Zhou, H.; Zhao, J.; Qing, W.; Feng, Y.; Tang, J. Speleothem-derived Asian Summer Monsoon variations in Central China, 54-46ka. J. Quat. Sci. 2011, 26, 781–790. [Google Scholar] [CrossRef]
- Chappell, J.; Chivas, A.; Wallensky, E.; Polach, H.A.; Aharon, P. Holocene palaeo-environmental changes, central to north Great Barrier Reef inner zone. J. Aust. Geol. Geophys. 1983, 8, 223–235. [Google Scholar]
- Hopley, D. Contrasting evidence for Holocene sea levels with special reference to the Bowen-Whitsunday area of Queensland. In Geographical Essays in Honour of Gilbert J. Butland; Douglas, I., Hobbs, J.E., Pigram, J.J., Eds.; University of New England: Armidale, Australia, 1975; pp. 51–84. [Google Scholar]
- Stuiver, M.; Reimer, P.J. Extended 14C data base and revised Calib 3.0 14C age calibration program. Radiocarbon 1993, 35, 215–230. [Google Scholar] [CrossRef]
- Neumann, A.C.; Macintyre, I.G. Reef response to sea level rise: Keep-up, catch-up or give-up. In Proceedings of the 5th Int Coral Reef Congress, Tahiti, French Polynesia, 27 May–1 June 1985; pp. 105–110. [Google Scholar]
- Kleypas, J.A. Coral reef development under naturally turbid conditions: Fringing reefs near Broad Sound, Australia. Coral Reefs 1996, 15, 153–167. [Google Scholar] [CrossRef]
- Dechnik, B.; Webster, J.M.; Webb, G.E.; Nothdurft, L.; Dutton, A.; Braga, J.C.; Zhao, J.X.; Duce, S.; Sadler, J. The evolution of the Great Barrier Reef during the Last Interglacial Period. Glob. Planet. Chang. 2017, 149, 53–71. [Google Scholar] [CrossRef]
- Purdy, E.G. Reef Configuration: Cause and Effect. Soc. Econ. Paleontol. Mineral. 1974, 18, 9–76. [Google Scholar]
- Webster, J.M.; Davies, P.J. Coral variation in two deep drill cores: Significance for the Pleistocene development of the Great Barrier Reef. Sediment. Geol. 2003, 159, 61–80. [Google Scholar] [CrossRef]
- Martin, J.M.; Braga, J.C.; Rivas, P. Coral successions in Upper Tortonian reefs in SE Spain. Lethaia 1989, 22, 271–286. [Google Scholar] [CrossRef]
- Insalaco, E. Facies and palaeoecology of Upper Jurassic (Middle Oxfordian) coral reefs in England. Facies 1999, 40, 81–100. [Google Scholar] [CrossRef]
- Santodomingo, N.; Novak, V.; Pretkovic, V.; Marshall, N.; Di Martino, E.; Giudice Cappelli, E.L.; Rösler, A.; Reich, S.; Braga, J.C.; Renema, W.; et al. A diverse patch reef from turbid waters in the middle Miocene (East Kalimantan, Indonesia). Palaios 2015, 30, 128–149. [Google Scholar] [CrossRef]
- Wilson, M.E.J.; Lokier, S.W. Siliciclastic and volcaniclastic influences on equatorial carbonates: Insights from the Neogene of Indonesia. Sedimentology 2002, 49, 583–601. [Google Scholar] [CrossRef]
- Smithers, S.; Larcombe, P. Late Holocene initiation and growth of a nearshore turbid-zone coral reef: Paluma Shoals, central Great Barrier Reef, Australia. Coral Reefs 2003, 22, 499–505. [Google Scholar] [CrossRef]
- Cabioch, G.; Montaggioni, L.F.; Faure, G. Holocene initiation and development of New Caledonian fringing reefs, SW Pacific. Coral Reefs 1995, 14, 131–140. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Fabricius, K.E. Shifting roles of heterotrophy and autotrophy in coral energetics under varying turbidity. J. Exp. Mar. Biol. Ecol. 2000, 252, 221–253. [Google Scholar] [CrossRef]
- Dechnik, B.; Webster, J.M.; Webb, G.E.; Nothdurft, L.; Zhao, J.X. Successive phases of Holocene reef flat development: Evidence from the mid- to outer Great Barrier Reef. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 466, 221–230. [Google Scholar] [CrossRef]
- Tudhope, A.W.; Scoffin, T.P. Growth and structure of fringing reefs in a muddy environment, South Thailand. J. Sediment. Res. Sect. A Sediment. Petrol. Process. 1994, 64, 752–764. [Google Scholar] [CrossRef]
- Perry, C.T.; Smithers, S.G.; Gulliver, P. Rapid vertical accretion on a “young” shore-detached turbid zone reef: Offshore Paluma Shoals, central Great Barrier Reef, Australia. Coral Reefs 2013, 32, 1143–1148. [Google Scholar] [CrossRef]
- Blanchon, P.; Jones, B.; Kalbfleisch, W. Anatomy of a fringing reef around Grand Cayman: Storm rubble, not coral framework. J. Sediment. Res. 1997, 67, 1–16. [Google Scholar]
- Pandolfi, J.M.; Minchin, P. A comparison of taxonomic composition and diversity between reef coral life and death assemblages in Madang Lagoon, Papua New Guinea. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 119, 321–341. [Google Scholar] [CrossRef]
- Clark, T.R.; Leonard, N.D.; Zhao, J.-X.; Brodie, J.; McCook, L.J.; Wachenfeld, D.R.; Duc Nguyen, A.; Markham, H.L.; Pandolfi, J.M. Historical photographs revisited: A case study for dating and characterizing recent loss of coral cover on the inshore Great Barrier Reef. Sci. Rep. 2016, 6, 19285. [Google Scholar] [CrossRef] [PubMed]
- Bartley, R.; Croke, J.; Bainbridge, Z.T.; Austin, J.M.; Kuhnert, P.M. Combining contemporary and long-term erosion rates to target erosion hot-spots in the Great Barrier Reef, Australia. Anthropocene 2015, 10, 1–12. [Google Scholar] [CrossRef]
- Bainbridge, Z.; Lewis, S.; Bartley, R.; Fabricius, K.; Collier, C.; Waterhouse, J.; Garzon-Garcia, A.; Robson, B.; Burton, J.; Wenger, A.; et al. Fine sediment and particulate organic matter: A review and case study on ridge-to-reef transport, transformations, fates, and impacts on marine ecosystems. Mar. Pollut. Bull. 2018, 135, 1205–1220. [Google Scholar] [CrossRef]
- Lewis, S.E.; Olley, J.; Furuichi, T.; Sharma, A.; Burton, J. Complex sediment deposition history on a wide continental shelf: Implications for the calculation of accumulation rates on the Great Barrier Reef. Earth Planet. Sci. Lett. 2014, 393, 146–158. [Google Scholar] [CrossRef]
- Lough, J.M.; Cooper, T.F. New insights from coral growth band studies in an era of rapid environmental change. Earth-Sci. Rev. 2011, 108, 170–184. [Google Scholar] [CrossRef]
- Inoue, M.; Ishikawa, D.; Miyaji, T.; Yamazaki, A.; Suzuki, A.; Yamano, H.; Kawahata, H.; Watanabe, T. Evaluation of Mn and Fe in coral skeletons (Porites spp.) as proxies for sediment loading and reconstruction of 50 yrs of land use on Ishigaki Island, Japan. Coral Reefs 2014, 33, 363–373. [Google Scholar] [CrossRef]
- Lewis, S.E.; Brodie, J.E.; McCulloch, M.T.; Mallela, J.; Jupiter, S.D.; Stuart Williams, H.; Lough, J.M.; Matson, E.G. An assessment of an environmental gradient using coral geochemical records, Whitsunday Islands, Great Barrier Reef, Australia. Mar. Pollut. Bull. 2012, 65, 306–319. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Bramston Reef | Stone Island South | Stone Island North | Middle Island | Holbourne Island |
|---|---|---|---|---|---|
| Distance offshore from mainland coast near Bowen | 0 km | 3 km | 3 km | 10 km | 40 km |
| Reef flat width (m) | 900 | 450 | 400 | 330 | 440 |
| Number of cores collected | 8 | 9 | 5 | 11 | 9 |
| Maximum core depth (m) | 4.6 | 5.1 | 5.0 | 7.2 | 8.3 |
| Number of 230Th ages in cores | 13 | 14 | 11 | 14 | 16 |
| Earliest recorded age of initiation (yBP) | 5396 ± 51 | 7247 ± 23 | 7064 ± 17 | 7873 ± 17 | 7520 ± 20 |
| Antecedent substrate | Transgressive sands and lag gravels overlying Pleistocene clay | Not recovered | Not recovered | Regolith clay or unconsolidated carbonate sediments | Last interglacial reef or granite |
| Shallowest depth 1 to antecedent surface (m) | 1.9 | n/a | n/a | 2.6 | 6.0 |
| Maximum depth 1 to antecedent surface (m) | 3.7 | >5.1 | >5.0 | 7.0 | 8.1 |
| Time when ~80% of reef flat had been developed (yBP) 2 | 2000 | 5000 | 4000 | 6000 | 6000 |
| Range of average net vertical accretion rates (mm/yr) 3 | 2.5–9.8 | 0.3–4.8 | 0.9–5.0 | 3.5–7.6 | 0.7–3.2 |
| Number of coral genera recorded in cores | 25 | 20 | 23 | 15 | At least 10 |
| Reference | [39] | [45] | [45] | [44] | [46] |
| Facies Name | Facies Description | Bramston Reef | Stone Island South | Stone Island North | Middle Island | Holbourne Island | |
|---|---|---|---|---|---|---|---|
| Facies A | Intertidal sands | % mud | 14.2 ± 5.2 | 4.2 ± 2.0 | 9.6 ± 5.2 | 3.3 ± 1.0 | 2.1 ± 0.8 |
| % CaCO3 | 56.0 ± 24.1 | 96.9 ± 2.3 | 92.7 ± 2.8 | 96.0 ± 0.5 | 97.3 ± 0.7 | ||
| Facies B | Reef framework, sandy matrix | % mud | 17.1 ± 4.6 | 8.6 ± 4.4 | 13.5 ± 7.0 | 9.8 ± 4.4 | 6.5 ± 2.0 |
| % CaCO3 | 73.1 ± 12.0 | 91.7 ± 8.8 | 87.4 ± 6.0 | 92.9 ± 1.9 | 97.4 ± 1.3 | ||
| Facies C | Reef framework, muddy-sand matrix | % mud | 32.4 ± 12.5 | 35.9 ± 12.7 | 20.7 ± 5.3 | 22.0 ± 10.2 | 18.0 ± 8.3 |
| % CaCO3 | 63.6 ± 13.7 | 75.4 ± 7.9 | 81.7 ± 9.7 | 84.2 ± 5.4 | 92.8 ± 3.9 | ||
| Facies D | Reef framework, muddy matrix | % mud | 53.8 ± 17.4 | 47.8 ± 13.9 | n/a | 46.5 ± 15.6 | n/a |
| % CaCO3 | 55.6 ± 7.1 | 70.5 ± 9.5 | n/a | 67.9 ± 10.2 | n/a | ||
| Facies E | Mud-silt unit | % mud | 41.1 ± 19.1 | n/a | n/a | n/a | n/a |
| % CaCO3 | 47.0 ± 2.6 | n/a | n/a | n/a | n/a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryan, E.J.; Smithers, S.G.; Lewis, S.E.; Clark, T.R.; Zhao, J.-x. The Variable Influences of Sea Level, Sedimentation and Exposure on Holocene Reef Development over a Cross-Shelf Transect, Central Great Barrier Reef. Diversity 2018, 10, 110. https://doi.org/10.3390/d10040110
Ryan EJ, Smithers SG, Lewis SE, Clark TR, Zhao J-x. The Variable Influences of Sea Level, Sedimentation and Exposure on Holocene Reef Development over a Cross-Shelf Transect, Central Great Barrier Reef. Diversity. 2018; 10(4):110. https://doi.org/10.3390/d10040110
Chicago/Turabian StyleRyan, Emma J., Scott G. Smithers, Stephen E. Lewis, Tara R. Clark, and Jian-xin Zhao. 2018. "The Variable Influences of Sea Level, Sedimentation and Exposure on Holocene Reef Development over a Cross-Shelf Transect, Central Great Barrier Reef" Diversity 10, no. 4: 110. https://doi.org/10.3390/d10040110
APA StyleRyan, E. J., Smithers, S. G., Lewis, S. E., Clark, T. R., & Zhao, J.-x. (2018). The Variable Influences of Sea Level, Sedimentation and Exposure on Holocene Reef Development over a Cross-Shelf Transect, Central Great Barrier Reef. Diversity, 10(4), 110. https://doi.org/10.3390/d10040110