Distribution of Cranberry Blue Butterflies (Agriades optilete) and Their Responses to Forest Disturbance from In Situ Oil Sands and Wildfires



Abstract

1. Introduction
- (1)
- Which forest types do cranberry blues inhabit?
- (2)
- What are the optimal conditions for observing cranberry blues?
- (3)
- How do cranberry blues respond to anthropogenic (i.e., clearing associated with in situ oil sands operations) and natural (i.e., wildfires) disturbance?
2. Materials and Methods
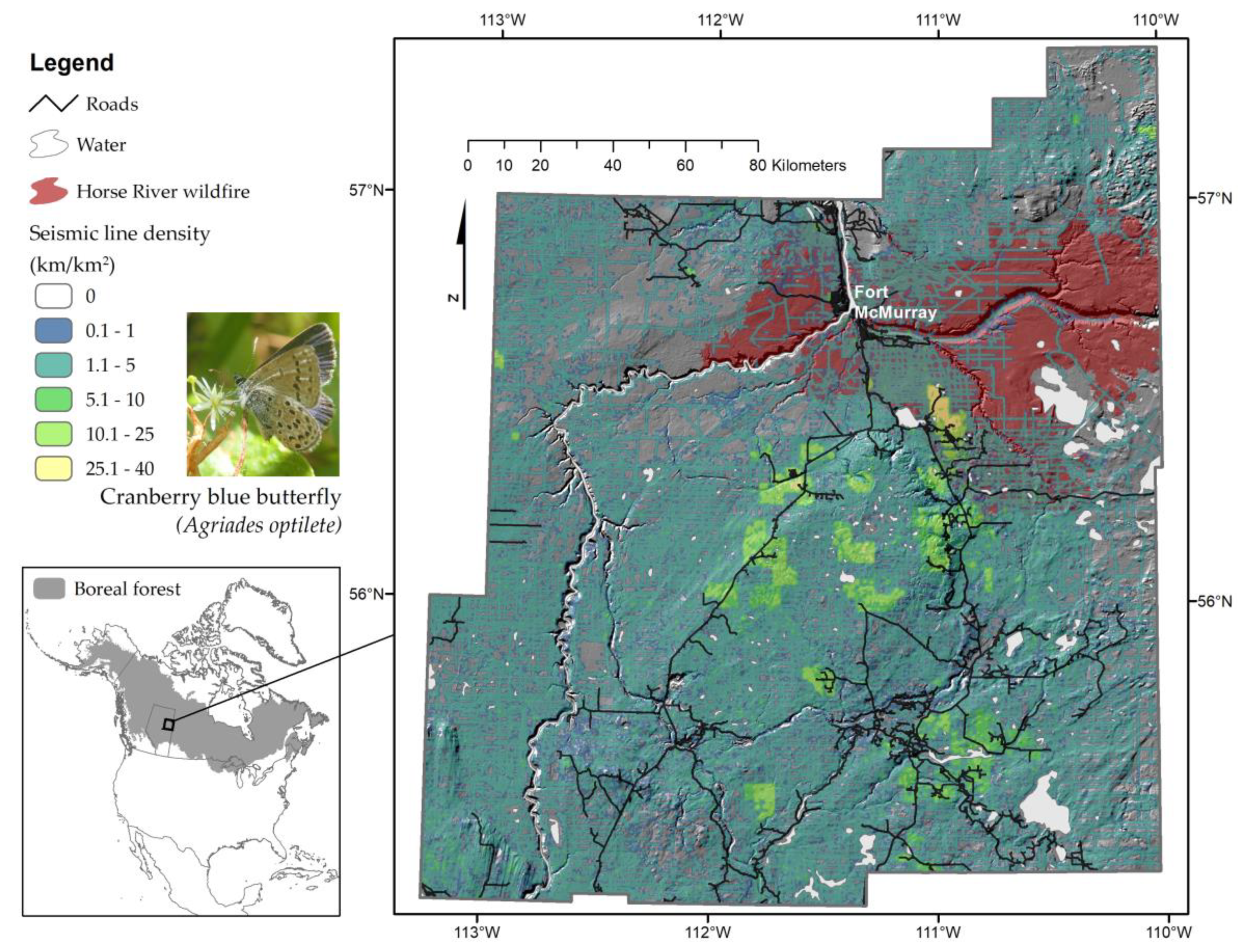
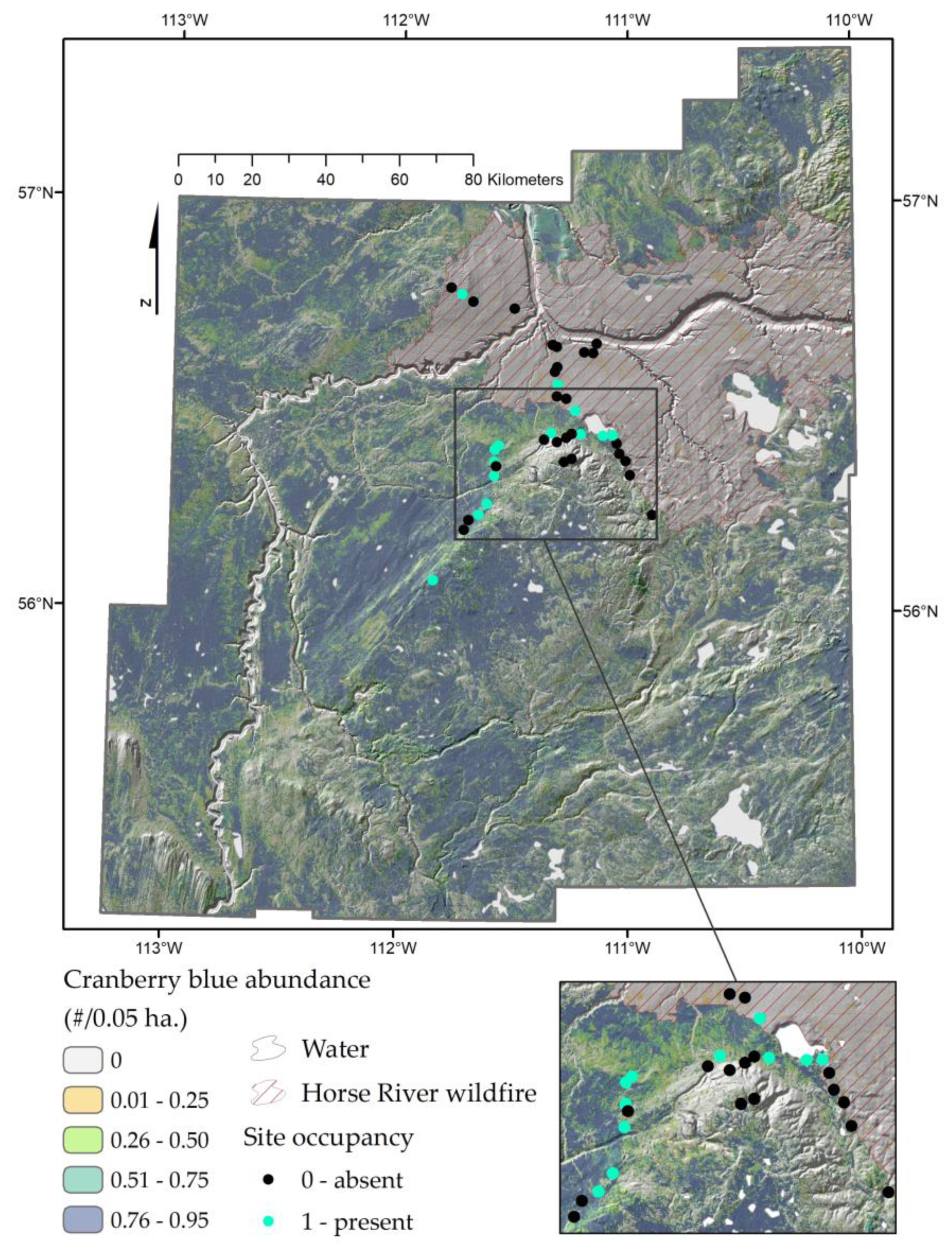
2.1. Study Area and Experimental Design
2.2. Statistical Design and Sampling Conditions
2.3. Responses to In Situ Oil Sands Disturbances
2.4. Responses to Wildfire and Distribution
2.5. Statistical Analysis
3. Results
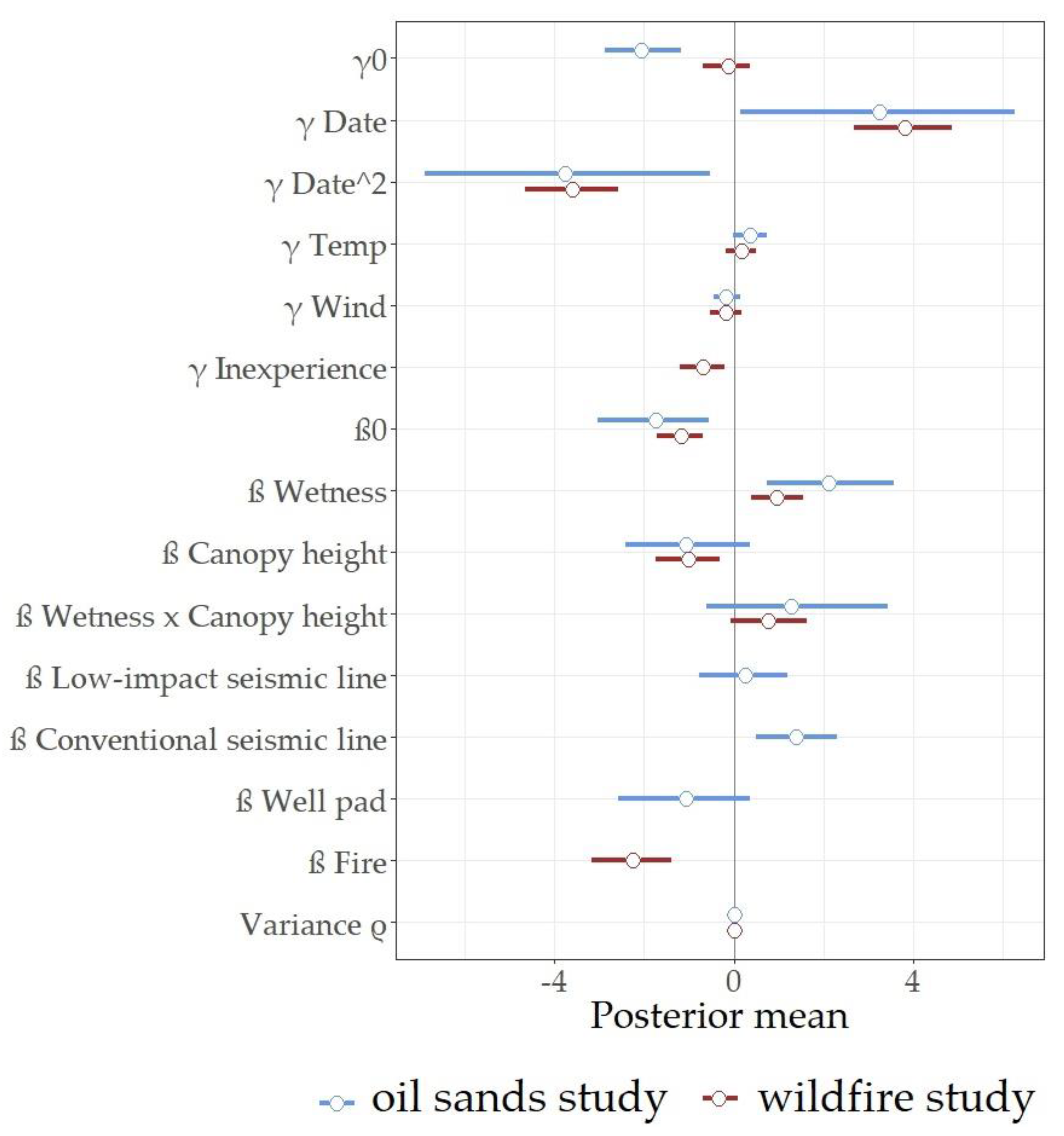
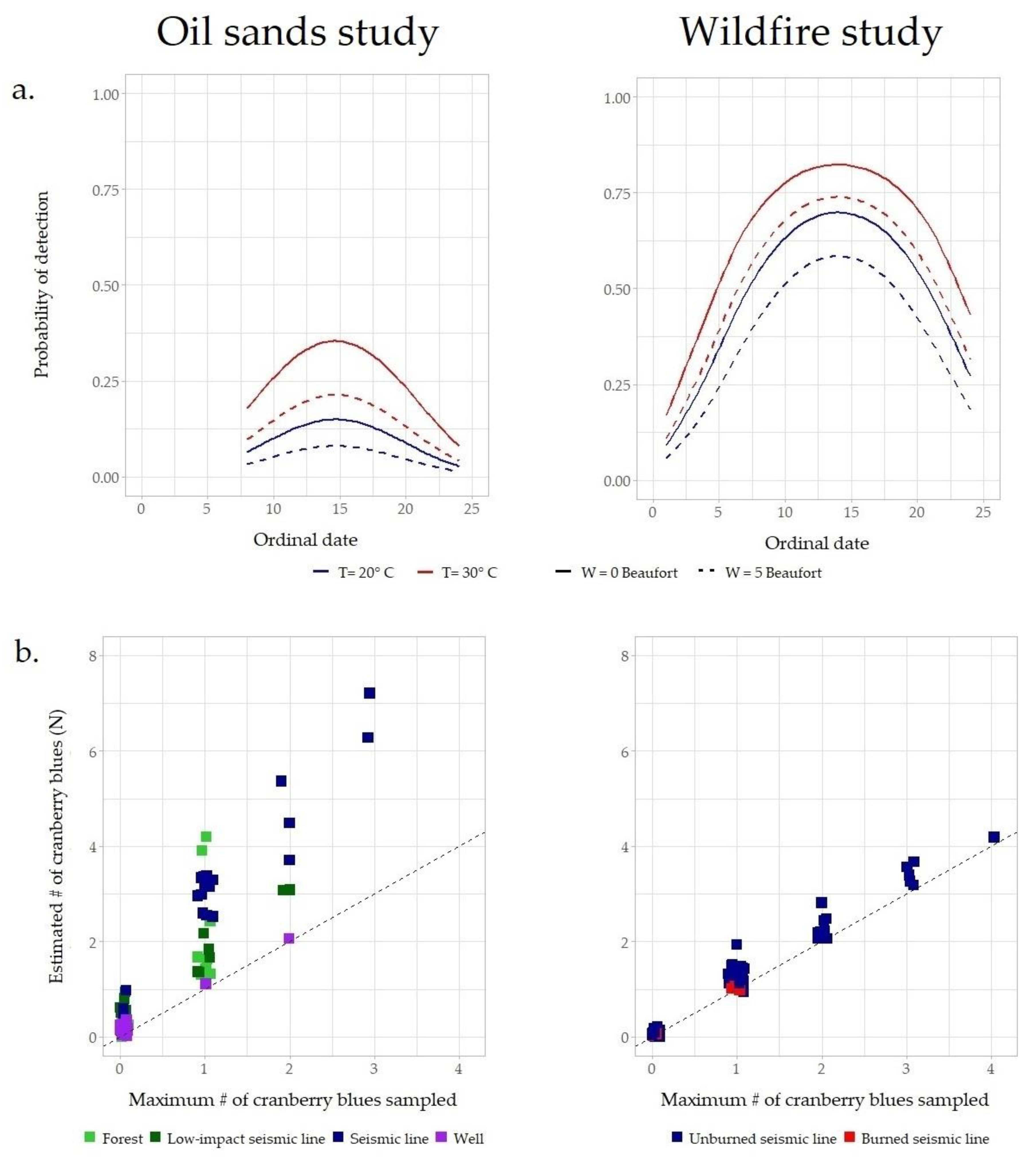
3.1. Probability of Detection of Cranberry Blues
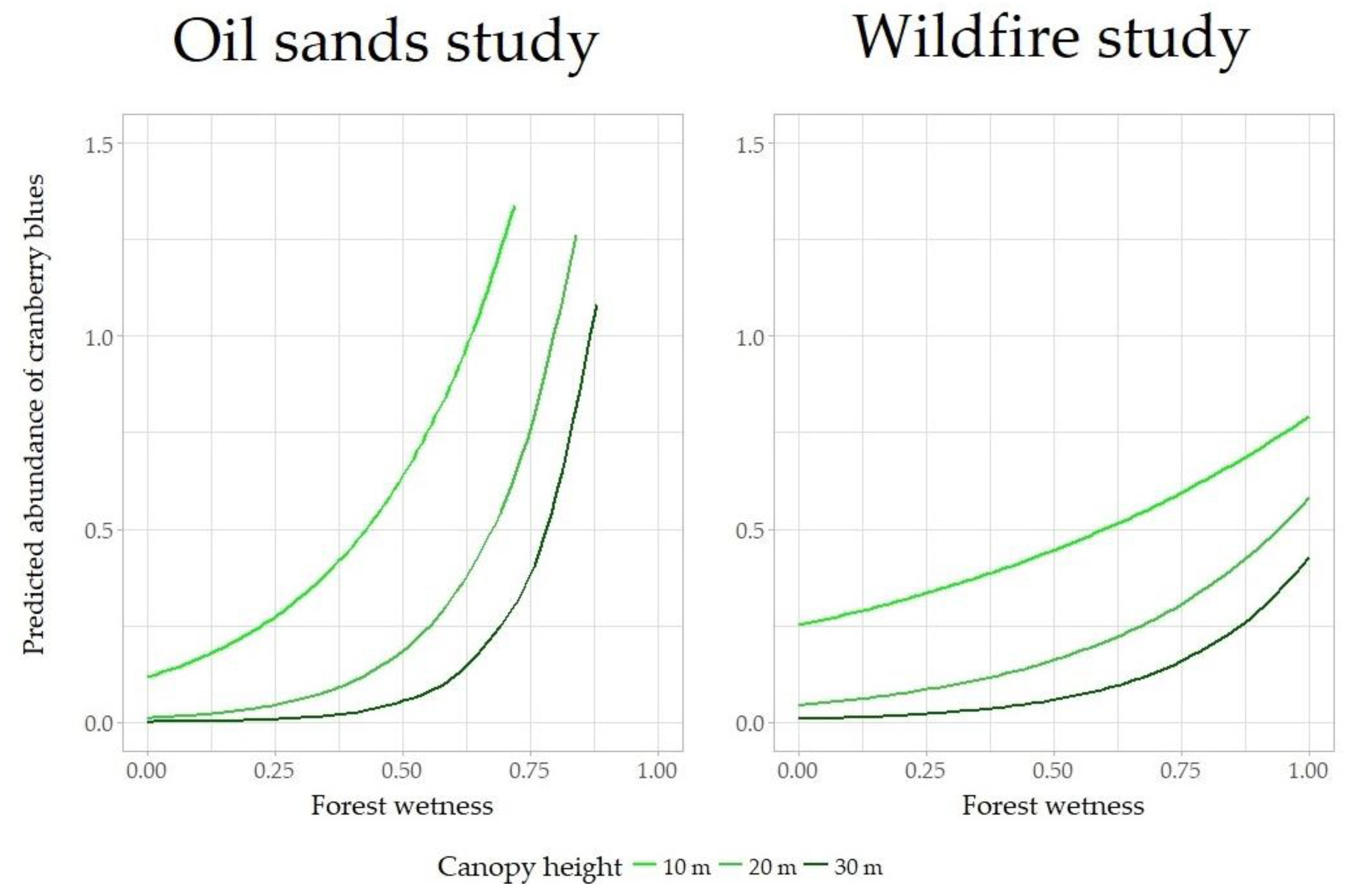
3.2. Abundance of Cranberry Blues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case-Study | Coefficient | Mean | Standard Deviation | 2.5% Quantile | 97.5% Quantile | p |
|---|---|---|---|---|---|---|
| Oil sands | β (Intercept) | −1.73 | 0.64 | −3.03 | −0.55 | <0.01 |
| Oil sands | β well pad | −1.05 | 0.76 | −2.58 | 0.37 | 0.08 |
| Oil sands | β low-impact line | 0.25 | 0.50 | −0.76 | 1.19 | 0.31 |
| Oil sands | β conventional line | 1.37 | 0.46 | 0.51 | 2.29 | <0.01 |
| Oil sands | β Wetness | 2.12 | 0.72 | 0.74 | 3.58 | <0.01 |
| Oil sands | β Canopy height | −1.06 | 0.70 | −2.40 | 0.35 | 0.06 |
| Oil sands | β Wetness × Canopy height | 1.28 | 1.06 | −0.61 | 3.44 | 0.11 |
| Oil sands | γ (Intercept) | −2.05 | 0.43 | −2.88 | −1.17 | <0.01 |
| Oil sands | γ Date | 3.23 | 1.56 | 0.16 | 6.26 | 0.02 |
| Oil sands | γ Date^2 | −3.77 | 1.62 | −6.89 | −0.52 | 0.01 |
| Oil sands | γ Temperature | 0.35 | 0.19 | −0.01 | 0.73 | 0.04 |
| Oil sands | γ Wind | −0.16 | 0.15 | −0.44 | 0.16 | 0.14 |
| Oil sands | Variance ρ | <0.01 | <0.01 | <0.01 | <0.01 | 0.04 |
| Wildfire | β (Intercept) | −1.17 | 0.26 | −1.70 | −0.70 | <0.01 |
| Wildfire | β Wetness | 0.95 | 0.30 | 0.39 | 1.54 | <0.01 |
| Wildfire | β Canopy height | −1.02 | 0.37 | −1.74 | −0.32 | <0.01 |
| Wildfire | β Wetness × Canopy height | 0.77 | 0.44 | −0.07 | 1.64 | 0.04 |
| Wildfire | β Fire | −2.24 | 0.45 | −3.16 | −1.38 | <0.01 |
| Wildfire | γ (Intercept) | −0.13 | 0.27 | −0.68 | 0.37 | 0.31 |
| Wildfire | γ Date | 3.80 | 0.57 | 2.67 | 4.86 | <0.01 |
| Wildfire | γ Date^2 | −3.59 | 0.54 | −4.65 | −2.56 | <0.01 |
| Wildfire | γ Temperature | 0.17 | 0.17 | −0.17 | 0.50 | 0.16 |
| Wildfire | γ Wind | −0.17 | 0.18 | −0.52 | 0.17 | 0.16 |
| Wildfire | γ Observer | −0.70 | 0.26 | −1.20 | −0.20 | <0.01 |
| Wildfire | Variance ρ | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
References
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Lefebvre, V.; Peres, C.A.; Banks-Leite, C.; Wearn, O.R.; Marsh, C.J.; Butchart, S.H.M.; Arroyo-Rodríguez, V.; Barlow, J.; Cerezo, A.; et al. Creation of forest edges has a global impact on forest vertebrates. Nature 2017, 551, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Fahrig, L. Ecological Responses to Habitat Fragmentation Per Se. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Didham, R.K.; Bascompte, J.; Wardle, D.A. Global change and species interactions in terrestrial ecosystems. Ecol. Lett. 2008, 11, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Devictor, V.; Van Swaay, C.; Brereton, T.; Brotons, L.; Chamberlain, D.; Heliölö, J.; Herrando, S.; Julliard, R.; Kuussaari, M.; Lindström, Å.; et al. Differences in the climatic debts of birds and butterflies at a continental scale. Nat. Clim. Chang. 2012, 2, 121–124. [Google Scholar] [CrossRef]
- Hanski, I. Habitat fragmentation and species richness. J. Biogeogr. 2015, 42, 989–993. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.J.; Didham, R.K.; Banks-leite, C.; Barlow, J.; Ewers, R.M.; Rosindell, J.; Holt, R.D.; Gonzalez, A.; Pardini, R.; Damschen, E.I.; et al. Is habitat fragmentation good for biodiversity? Biol. Conserv. 2018, 226, 9–15. [Google Scholar] [CrossRef]
- Martinson, H.M.; Fagan, W.F. Trophic disruption: A meta-analysis of how habitat fragmentation affects resource consumption in terrestrial arthropod systems. Ecol. Lett. 2014, 17, 1178–1189. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.M.; Hudgens, B.; Damschen, E.I.; Levey, D.J.; Orrock, J.L.; Tewksbury, J.J.; Weldon, A.J. Assessing positive and negative ecological effects of corridors. In Sources, Sinks and Sustainability; Liu, J., Hull, V., Morzillo, A.T., Wiens, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 475–503. ISBN 9780511842399. [Google Scholar]
- Fisher, J.T.; Burton, A.C. Wildlife winners and losers in an oil sands landscape. Front. Ecol. Environ. 2018, 16, 323–328. [Google Scholar] [CrossRef]
- Thomas, J.A. Butterfly communities under threat. Science 2016, 353, 216–218. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017. [Google Scholar] [CrossRef] [PubMed]
- Menéndez, R.; González-Megías, A.; Lewis, O.T.; Shaw, M.R.; Thomas, C.D. Escape from natural enemies during climate-driven range expansion: A case study. Ecol. Entomol. 2008, 33, 413–421. [Google Scholar] [CrossRef]
- Riva, F.; Acorn, J.H.; Nielsen, S.E. Localized disturbances from oil sands developments increase butterfly diversity and abundance in Alberta’s boreal forests. Biol. Conserv. 2018, 217, 173–180. [Google Scholar] [CrossRef]
- Swengel, S.R.; Schlicht, D.; Olsen, F.; Swengel, A.B. Declines of prairie butterflies in the midwestern USA. J. Insect Conserv. 2011, 15, 327–339. [Google Scholar] [CrossRef]
- Van Swaay, C.; Van Strien, A.; Harpke, A. The European Grassland Butterfly Indicator: 1990–2011; Lund University: Lund, Sweden, 2013. [Google Scholar]
- Sands, D.P.A.; New, T.R. Conservation of the Richmond Birdwing Butterfly in Australia; Springer: Dordrecht, The Netherlands, 2013; ISBN 9789400771703. [Google Scholar]
- Van Swaay, C.; Cuttelod, A.; Collins, S.; Maes, D.; López Munguira, M.; Šašić, M.; Settele, J.; Verovnik, R.; Verstrael, T.; Warren, M.; et al. European Red List of Butterfies; International Union for Conservation of Nature: Gland, Switzerland, 2010; ISBN 9789279141515. [Google Scholar]
- Xerces Society Red Listed butterflies of North America. Available online: https://xerces.org/red-lists/ (accessed on 16 October 2018).
- New, T.R. Conservation Biology of Lycaenidae (Butterflies); IUCN Species Survival Commission: Gland, Switzerland, 1993. [Google Scholar]
- Burke, R.J.; Fitzsimmons, J.M.; Kerr, J.T. A mobility index for Canadian butterfly species based on naturalists’ knowledge. Biodivers. Conserv. 2011, 20, 2273–2295. [Google Scholar] [CrossRef]
- Riva, F.; Barbero, F.; Bonelli, S.; Balletto, E.; Casacci, L.P. The acoustic repertoire of lycaenid butterfly larvae. Bioacoustics 2017, 26, 77–90. [Google Scholar] [CrossRef]
- Bird, C.D.; Hilchie, G.J.; Kondla, N.G.; Pike, E.M.; Sperling, F.A.H. Alberta Butterflies; Alberta Public Affairs Bureau/Queens Printer, The Provincial Museum of Alberta: Edmonton, AL, USA, 1995; ISBN 0-7732-1672-3. [Google Scholar]
- Viljur, M.L.; Teder, T. Butterflies take advantage of contemporary forestry: Clear-cuts as temporary grasslands. For. Ecol. Manag. 2016, 376, 118–125. [Google Scholar] [CrossRef]
- NatureServe Explorer: An Online Encyclopedia of Life [Web Application]. Version 7.1. 2017. Available online: http://www.natureserve.org/explorer (accessed on 16 October 2018).
- Van Rensen, C.K.; Nielsen, S.E.; White, B.; Vinge, T.; Lieffers, V.J. Natural regeneration of forest vegetation on legacy seismic lines in boreal habitats in Alberta’s oil sands region. Biol. Conserv. 2015, 184, 127–135. [Google Scholar] [CrossRef]
- Flannigan, M.D.; Krawchuk, M.A.; de Groot, W.J.; Wotton, M.B.; Gowman, L.M. Implications of changing climate for global wildland fire. Int. J. Wildl. Fire 2009, 18, 483–507. [Google Scholar] [CrossRef]
- Jaeger, J.A.G. Landscape division, splitting index, and effective mesh size: New measures of landscape fragmentation. Landsc. Ecol. 2000, 15, 115–130. [Google Scholar] [CrossRef]
- Dabros, A.; Pyper, M.; Castilla, G. Seismic lines in the boreal and arctic ecosystems of North America: Environmental impacts, challenges, and opportunities. Environ. Rev. 2018, 16, 1–16. [Google Scholar] [CrossRef]
- Riva, F.; Acorn, J.H.; Nielsen, S.E. Narrow anthropogenic corridors direct the movement of a generalist boreal butterfly. Biol. Lett. 2018. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.G.; Stocks, B.J. Forest fires and sustainability in the boreal forests of Canada. Ambio 1998, 27, 545–550. [Google Scholar]
- Burton, P.J.; Parisien, M.A.; Hicke, J.A.; Hall, R.J.; Freeburn, J.T. Large fires as agents of ecological diversity in the North American boreal forest. Int. J. Wildl. Fire 2008, 17, 754–767. [Google Scholar] [CrossRef]
- Simms, C.D. Canada’s Fort McMurray fire: Mitigating global risks. Lancet Glob. Health 2016, 4, e520. [Google Scholar] [CrossRef]
- New, T.R. Insects, Fire and Conservation; Springer: Basel, Switzerland, 2014; ISBN 9783319080963. [Google Scholar]
- Swengel, A.B. A literature review of insect responses to fire, compared to other conservation managements of open habitat. Biodivers. Conserv. 2001, 10, 1141–1169. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guo, X.; Coops, N.C.; Tompalski, P.; Nielsen, S.E.; Bater, C.W.; John Stadt, J. Regional mapping of vegetation structure for biodiversity monitoring using airborne lidar data. Ecol. Inform. 2017, 38, 50–61. [Google Scholar] [CrossRef]
- Oltean, G.S.; Comeau, P.G.; White, B. Linking the depth-to-water topographic index to soil moisture on boreal forest sites in Alberta. For. Sci. 2016, 62, 154–165. [Google Scholar] [CrossRef]
- Murphy, P.N.C.; Ogilvie, J.; Connor, K.; Arp, P.A. Mapping wetlands: A comparison of two different approaches for New Brunswick, Canada. Wetlands 2007, 27, 846–854. [Google Scholar] [CrossRef]
- Mao, L.; Bater, C.W.; Stadt, J.J.; White, B.; Tompalski, P.; Coops, N.C.; Nielsen, S.E. Environmental landscape determinants of maximum forest canopy height of boreal forests. J. Plant Ecol. 2017. [Google Scholar] [CrossRef]
- Pollard, E.; Yates, T.J. Monitoring Butterflies for Ecology and Conservation: The British Butterfly Monitoring Scheme; Chapman and Hall: London, UK, 1993; ISBN 0412402203. [Google Scholar]
- Van Swaay, C.A.M.; Nowicki, P.; Settele, J.; Van Strien, A.J. Butterfly monitoring in Europe: Methods, applications and perspectives. Biodivers. Conserv. 2008, 17, 3455–3469. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 17 October 2018).
- Royle, J.A. N-Mixture Models for Estimating Population Size from Spatially Replicated Counts. Biometrics 2004, 60, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Latimer, A.M.; Wu, S.; Gelfand, A.E.; Silander, J.A. Building statistical models to analyze species distributions. Ecol. Appl. 2006, 16, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Lichstein, J.W.; Simons, T.R.; Shriner, S.A.; Franzreb, K.E. Spatial autocorrelation and autoregressive models in ecology. Ecol. Monogr. 2002, 72, 445–463. [Google Scholar] [CrossRef]
- Lahoz-Monfort, J.J.; Guillera-Arroita, G.; Wintle, B.A. Imperfect detection impacts the performance of species distribution models. Glob. Ecol. Biogeogr. 2014, 23, 504–515. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, A.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Nowicki, P.; Settele, J.; Henry, P.-Y.; Woyciechowski, M. Butterfly Monitoring Methods: The ideal and the Real World. Isr. J. Ecol. Evol. 2008, 54, 69–88. [Google Scholar] [CrossRef]
- Rabinowitz, D.; Cairns, S.; Dillon, T. Seven forms of rarity and their frequency in the flora of the British Isles. In Conservation Biology: The Science of Scarcity and Diversity; Elsevier: Amsterdam, The Netherlands, 1986; ISBN 0-87893-794-3. [Google Scholar]
- Hart, S.A.; Chen, H.Y.H. Fire, Logging, and Overstory Affect Understory Abundance, Diversity, and Composition in Boreal Forest. Ecol. Monogr. 2008, 78, 123–140. [Google Scholar] [CrossRef]
- Stern, E.; Riva, F.; Nielsen, S. Effects of Narrow Linear Disturbances on Light and Wind Patterns in Fragmented Boreal Forests in Northeastern Alberta. Forests 2018, 9, 486. [Google Scholar] [CrossRef]
- Stralberg, D.; Wang, X.; Parisien, M.-A.; Robinne, F.-N.; Sólymos, P.; Mahon, C.L.; Nielsen, S.E.; Bayne, E.M. Wildfire-mediated vegetation change in boreal forests of Alberta, Canada. Ecosphere 2018, 9, e02156. [Google Scholar] [CrossRef]





| Oil sands Study (2016) | Wildfire Study (2017) | |||||
|---|---|---|---|---|---|---|
| Point-count area | 20 m2 (5 m × 4 m) | 500 m2 (50 m × 10 m) | ||||
| Treatment | Forest | Low-impact seismic line | Conventional seismic line | Exploratory well pads | Unburned seismic line | Burned seismic line |
| Number of point-count locations | 30 | 30 | 30 | 30 | 5 per site, in 22 sites (N = 110) | 5 per site, in 18 sites (N = 90) |
| Number of visits per point-count | 4, with one observer | 4, with one observer | 4, with one observer | 4, with one observer | 2, with two observers | 2, with two observers |
| Samples per treatment | 120 | 120 | 120 | 120 | 440 | 360 |
| Number of cranberry blues observed | 16 | 12 | 33 | 3 | 115 | 9 |
| Point-counts with cranberry blues | 10/30 (33%) | 7/30 (23%) | 16/30 (53%) | 2/30 (7%) | 34/110 (31%) | 6/90 (7%) |
| Sites with cranberry blues | n.a. | 11/22 (50%) | 3/18 (17%) | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riva, F.; Acorn, J.H.; Nielsen, S.E. Distribution of Cranberry Blue Butterflies (Agriades optilete) and Their Responses to Forest Disturbance from In Situ Oil Sands and Wildfires. Diversity 2018, 10, 112. https://doi.org/10.3390/d10040112
Riva F, Acorn JH, Nielsen SE. Distribution of Cranberry Blue Butterflies (Agriades optilete) and Their Responses to Forest Disturbance from In Situ Oil Sands and Wildfires. Diversity. 2018; 10(4):112. https://doi.org/10.3390/d10040112
Chicago/Turabian StyleRiva, Federico, John H. Acorn, and Scott E. Nielsen. 2018. "Distribution of Cranberry Blue Butterflies (Agriades optilete) and Their Responses to Forest Disturbance from In Situ Oil Sands and Wildfires" Diversity 10, no. 4: 112. https://doi.org/10.3390/d10040112
APA StyleRiva, F., Acorn, J. H., & Nielsen, S. E. (2018). Distribution of Cranberry Blue Butterflies (Agriades optilete) and Their Responses to Forest Disturbance from In Situ Oil Sands and Wildfires. Diversity, 10(4), 112. https://doi.org/10.3390/d10040112