1. Introduction
Angiogenesis is the process in which new blood vessels are formed from pre-existing vessels. It is crucial in many physiological contexts, such as in wound healing, and as an adaptive response to hypoxia and ischemia [
1]. However, imbalance in angiogenic regulation can cause deleterious effects, as it accelerates inflammation-driven pathologies, as seen in atherosclerosis and cancer [
2,
3]. New vessels provide additional conduits for the delivery of inflammatory cells and cytokines that promote atherosclerotic plaque development and rupture. They also deliver the oxygen and nutrients necessary to sustain tumor growth and serve as potential routes for metastatic spread [
4].
Angiogenesis-associated conditions are highly prevalent globally, with cardiovascular disease (CVD) and cancer among the leading causes of morbidity and mortality worldwide. Current anti-angiogenic agents are limited, as they can interfere with physiological angiogenic processes, while pro-angiogenic therapies can potentially exacerbate chronic inflammation and inadvertently precipitate tumorigenesis [
5,
6]. Given the critical role of angiogenesis across such diverse pathologies, any agent capable of differentially modulating angiogenesis in a context-specific manner would be of great therapeutic value.
We previously identified a novel angiogenic role for tripartite motif-containing protein 2 (TRIM2) [
7]. Lentiviral short hairpin (sh)RNA knockdown of TRIM2 impaired endothelial cell tubule formation in both hypoxia and inflammatory conditions in vitro [
7]. We have also shown that TRIM2 knockdown attenuates the ability of human coronary artery endothelial cells (HCAECs) to migrate and proliferate in response to hypoxic and inflammatory stimuli. However, whether these effects are translated in vivo, and what the molecular mechanisms are underlying these, remains unknown.
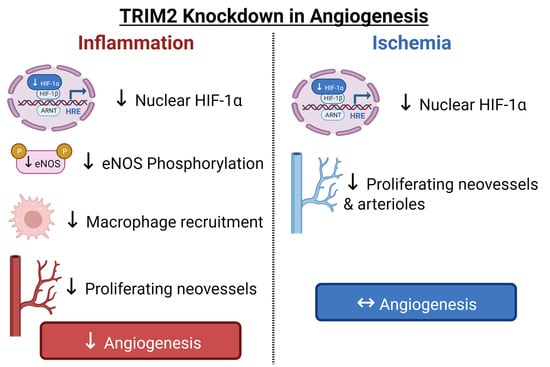
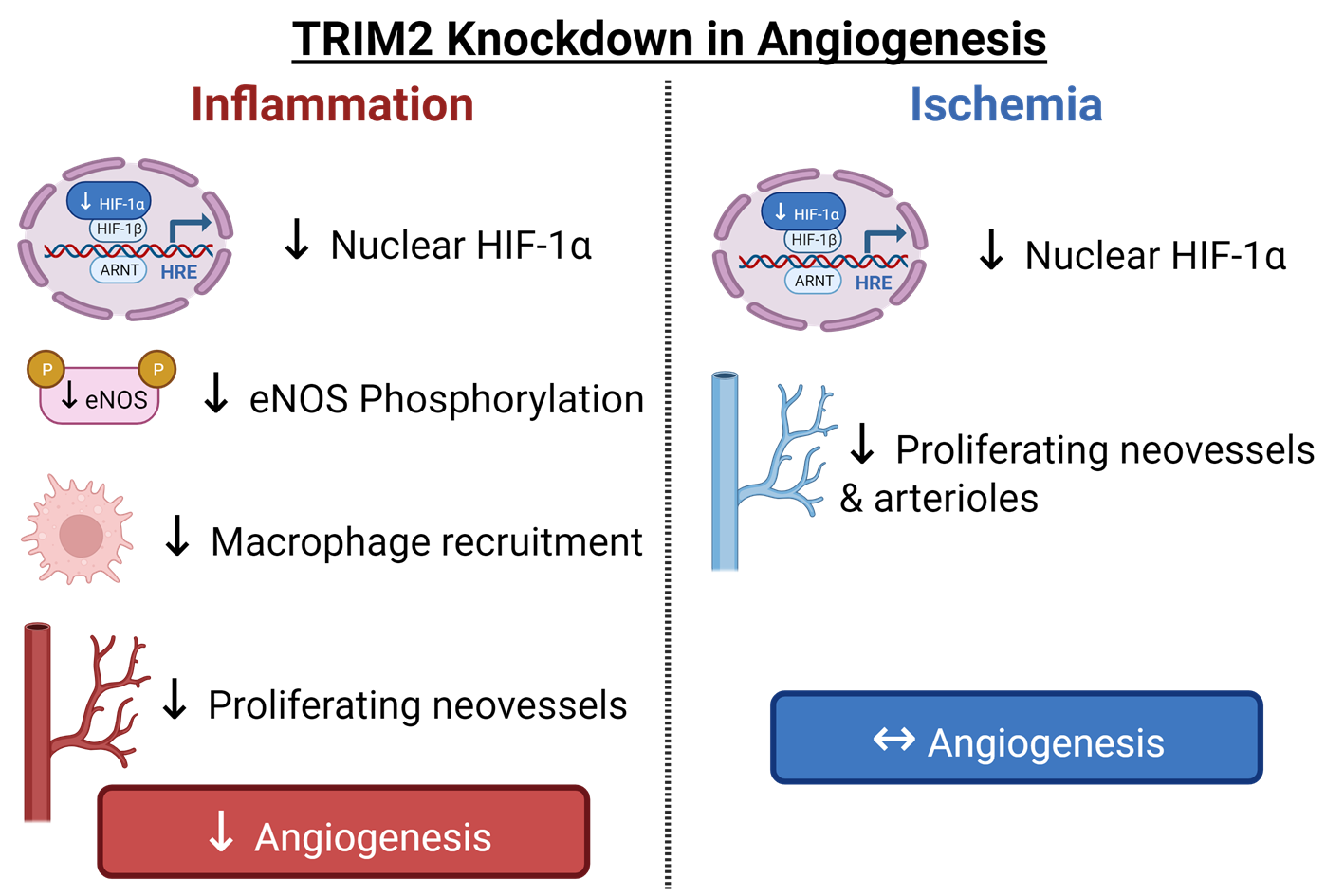
In this study, we used CRISPR/Cas9-generated homozygous Trim2 null (Trim2−/−) mice to evaluate the functional importance of TRIM2 in two well-validated models of pathological inflammation-driven angiogenesis and physiological hypoxia-mediated angiogenesis, namely the periarterial cuff and hindlimb ischemia models, respectively. In Trim2−/− mice, we report markedly attenuated infiltration of adventitial macrophages in response to femoral artery cuff placement, when compared to wildtype (WT) littermates concomitant with a reduction in mRNA levels of the macrophage marker cluster of differentiation 68 (Cd68). Mechanistically, we show that TRIM2 knockdown in human coronary artery endothelial cells (HCAECs) attenuates the induction of key mediators involved in the classical inflammation-driven angiogenic signaling pathway, including nuclear translocation of hypoxia-inducible factor (HIF)-1α and phosphorylation of downstream mediator endothelial nitric oxide synthase (eNOS).
In contrast, we find no significant differences in blood flow reperfusion despite a reduction in proliferating neovessels and arterioles in the ischemic hindlimbs of Trim2−/− and WT mice. In vitro, while the hypoxia-mediated induction of HIF-1α was tempered by TRIM2 knockdown, further downstream activation of angiogenic signaling proteins were unaffected. These findings collectively highlight a novel role for TRIM2 in the regulation of inflammation-driven angiogenesis and delineate the mechanistic basis for these effects. We propose TRIM2 to be a potential therapeutic target for diseases driven by pathological angiogenesis, unlimited by the usual adverse effects associated with inhibiting physiological angiogenesis.
3. Discussion
Dysregulated angiogenesis crucially underpins a wide range of chronic and debilitating diseases including atherosclerotic CVD and cancer. TRIM2 has emerged as a promising novel target that may differentially modulate both inflammation-driven pathological angiogenesis and hypoxia-stimulated physiological angiogenesis, as our previous studies have demonstrated impaired endothelial tubule formation in HCAECs with TRIM2 knockdown in vitro [
7]. Here, we report markedly reduced adventitial macrophage infiltration following
Trim2 deletion in a murine periarterial cuff model of inflammation-driven angiogenesis, concomitant with reduced proliferating adventitial neovessels and attenuated the induction of the inflammatory response. Correspondingly, we find that TRIM2 knockdown in HCAECs suppresses the TNFα-driven induction of several classical angiogenic mediators, particularly nuclear HIF-1α and reduced activation of the eNOS angiogenic signaling pathway.
Trim2 deletion, however, did not alter the capacity for blood flow reperfusion nor the extent of neovascularization in the murine hindlimb ischemia model despite a reduction in proliferating neovessels and arterioles. While TRIM2 knockdown in vitro suppressed the hypoxia-driven stimulation of nuclear HIF-1α, it did not affect downstream expression and activation of pro-angiogenic signaling pathways.
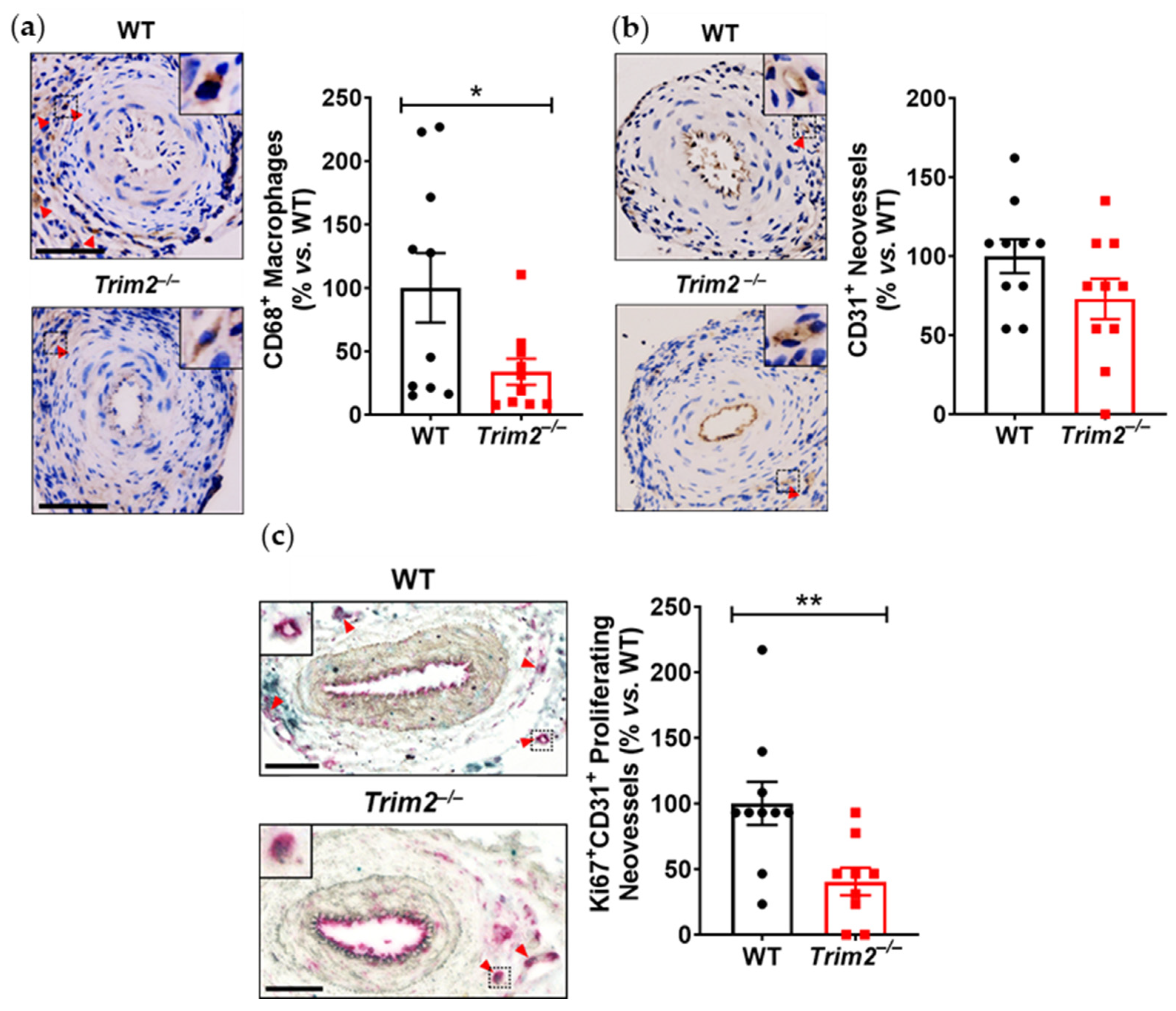
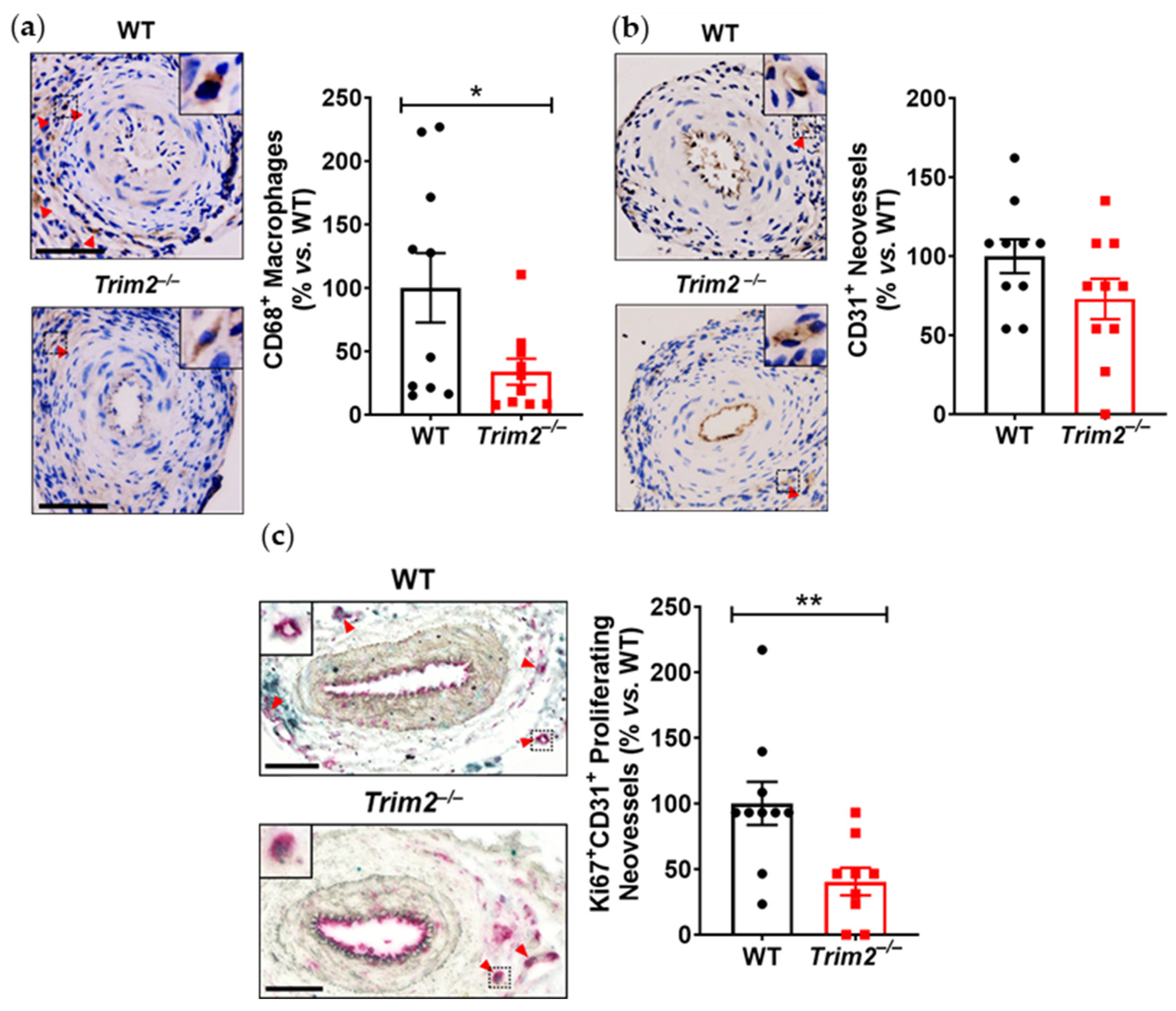
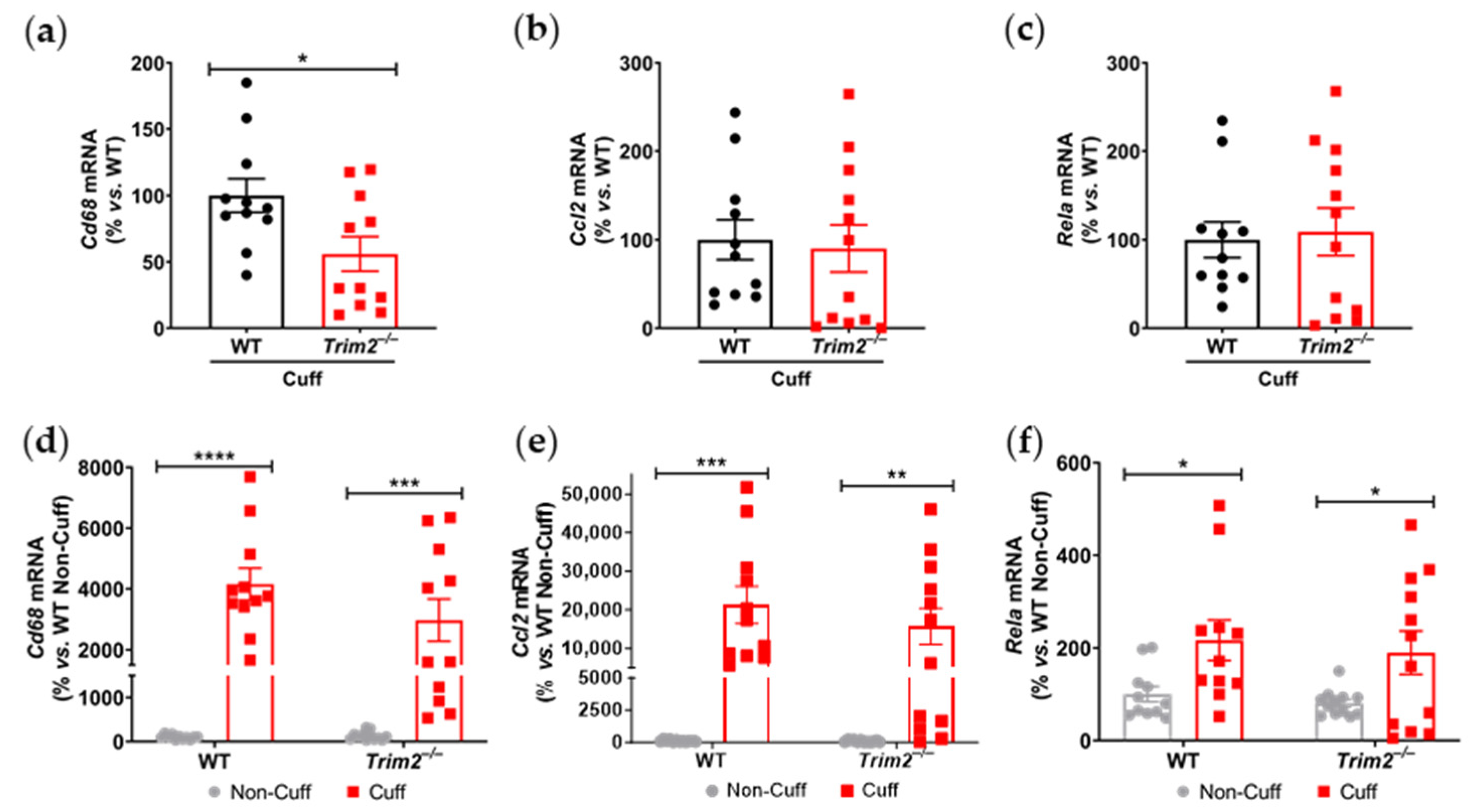
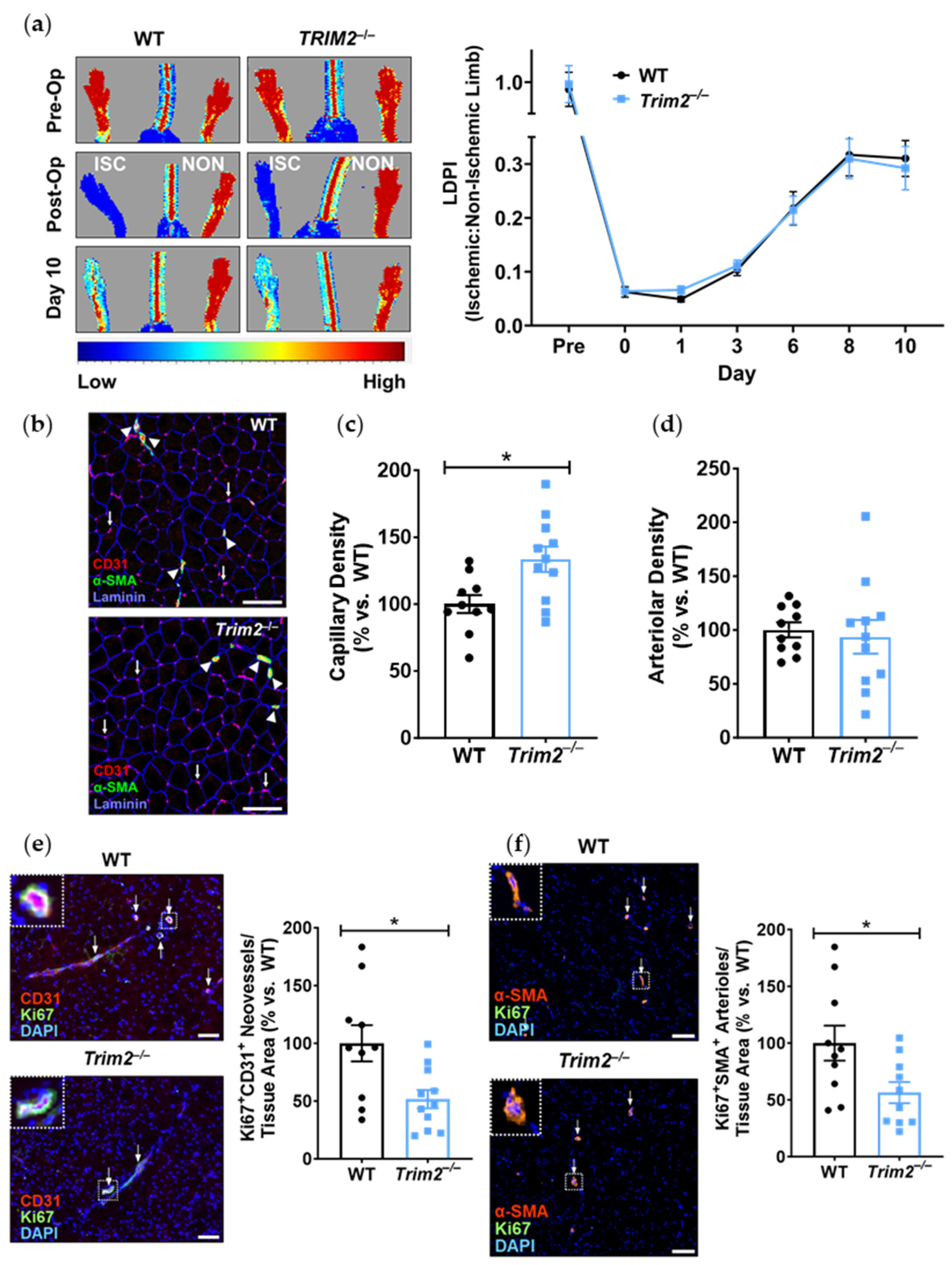
Our findings, particularly from the periarterial cuff model and the mechanistic studies, are consistent with our previous work showing the inhibition of inflammation-induced endothelial tubule formation after TRIM2 knockdown [
7]. The reduction in CD68
+ macrophage infiltration into the inflamed arteries of
Trim2−/− mice suggests that TRIM2 may be involved in broader mechanisms of inflammatory activation, which enhance vessel growth by stimulating a wealth of pro-angiogenic growth factors and mediators [
8]. The concomitant reduction in
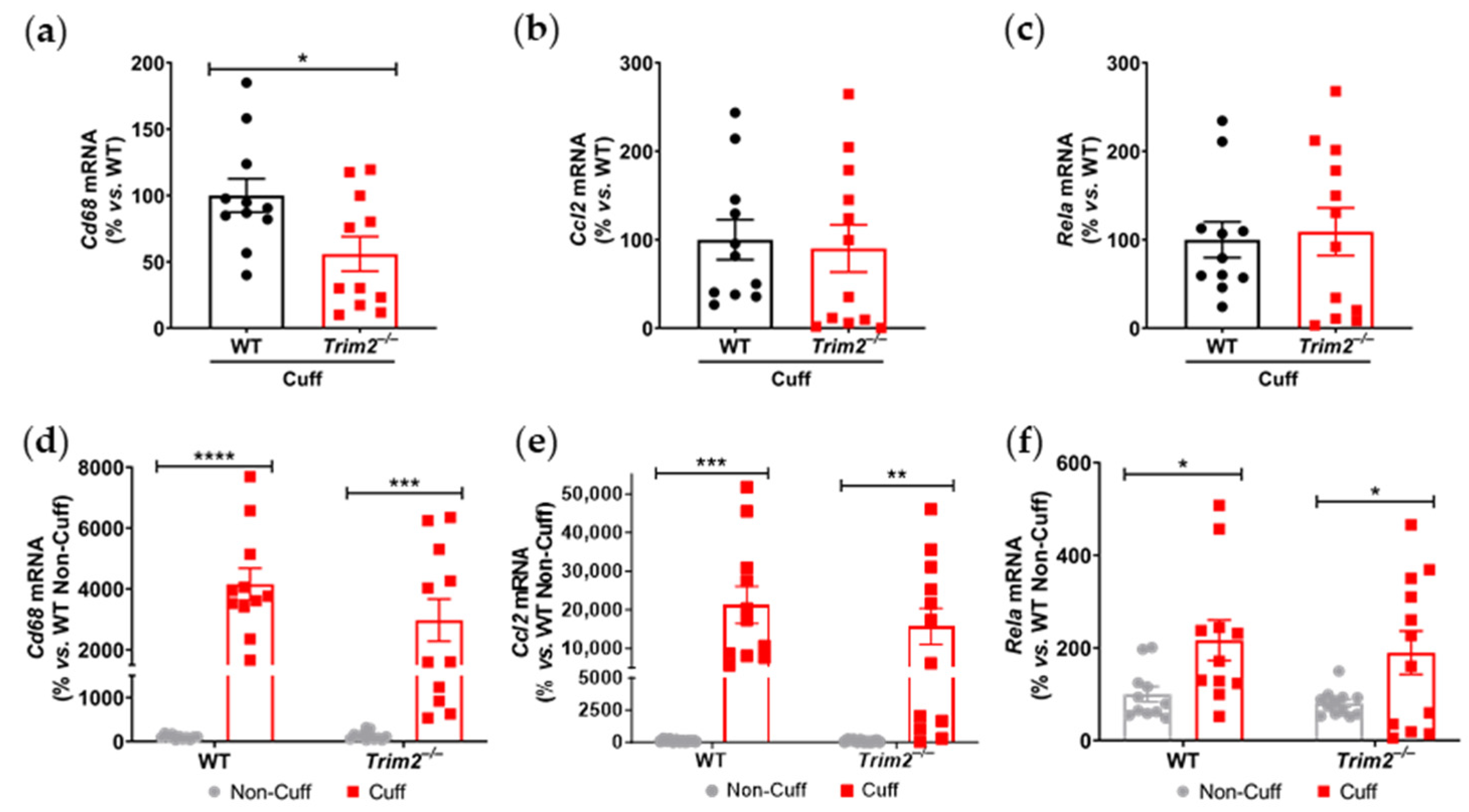
Cd68 mRNA levels in the cuffed arteries of
Trim2−/− mice further support the idea that TRIM2 plays a key role in regulating inflammation-driven pathological angiogenesis, particularly in the early stages of macrophage recruitment to the site of injury. The reduction in proliferating adventitial neovessels in the cuffed arteries of
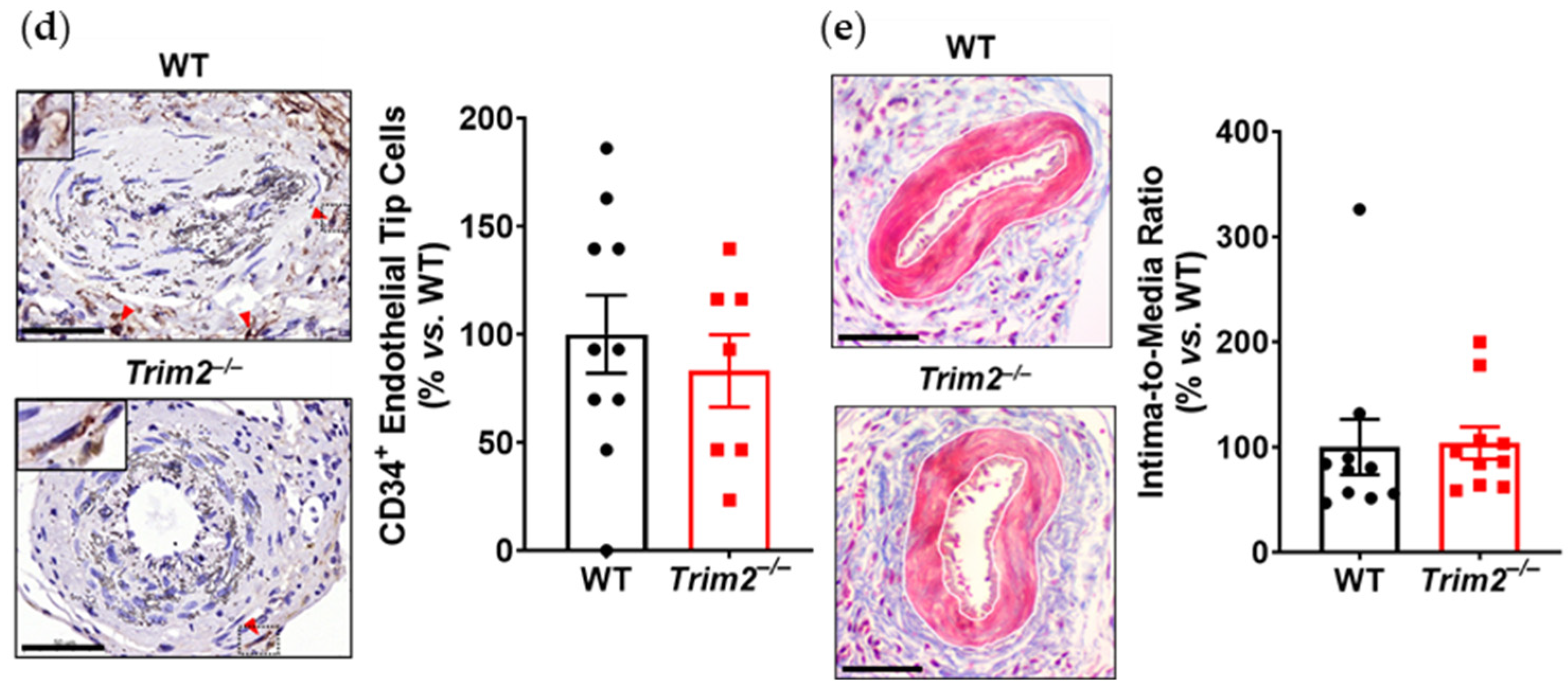
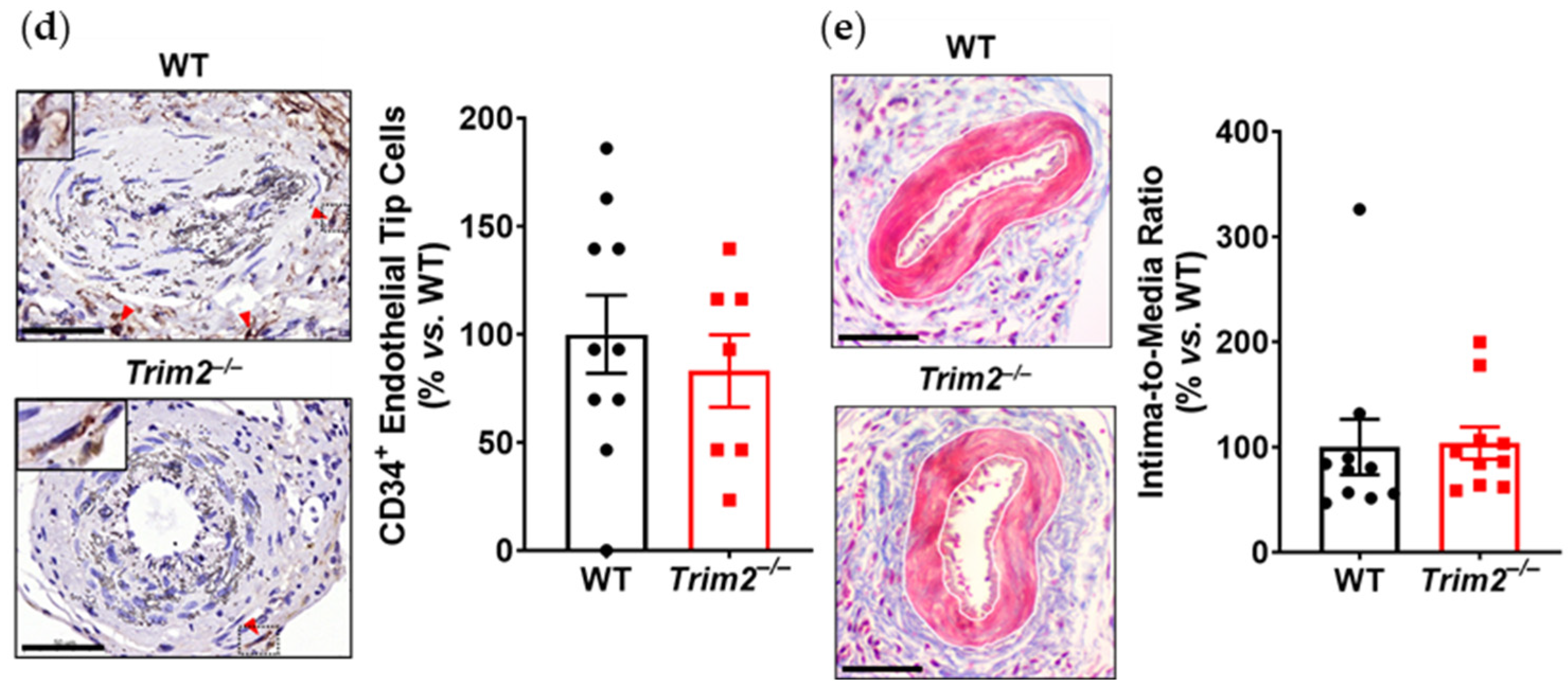
Trim2−/− mice is reflective of this. This was associated with a trend towards reduced total neovessels. Had the study duration been extended beyond 3 weeks, it could be hypothesized that a larger reduction in adventitial neovessels would be observed, as the blunted macrophage response would lead to fewer pro-angiogenic factors being released. No differences were observed in the presence of CD34
+ endothelial tip cells. The intima-to-media ratio, though, which assesses the formation of a thickened neointima as an ‘outside-in’ response to adventitial inflammation, was not affected by
Trim2 deletion, indicating that the reduction in adventitial neovessels was a specific effect on angiogenesis and not a consequence of the development of a smaller neointimal or media. Furthermore, recent clinical studies have identified TRIM2 as a potential oncogene in human cancer cell lines including colorectal carcinoma, epithelial ovarian carcinoma and osteosarcoma [
9,
10,
11]. These observations fit with a postulated role for TRIM2 in inflammation-driven angiogenesis, a hallmark of cancer development and progression.
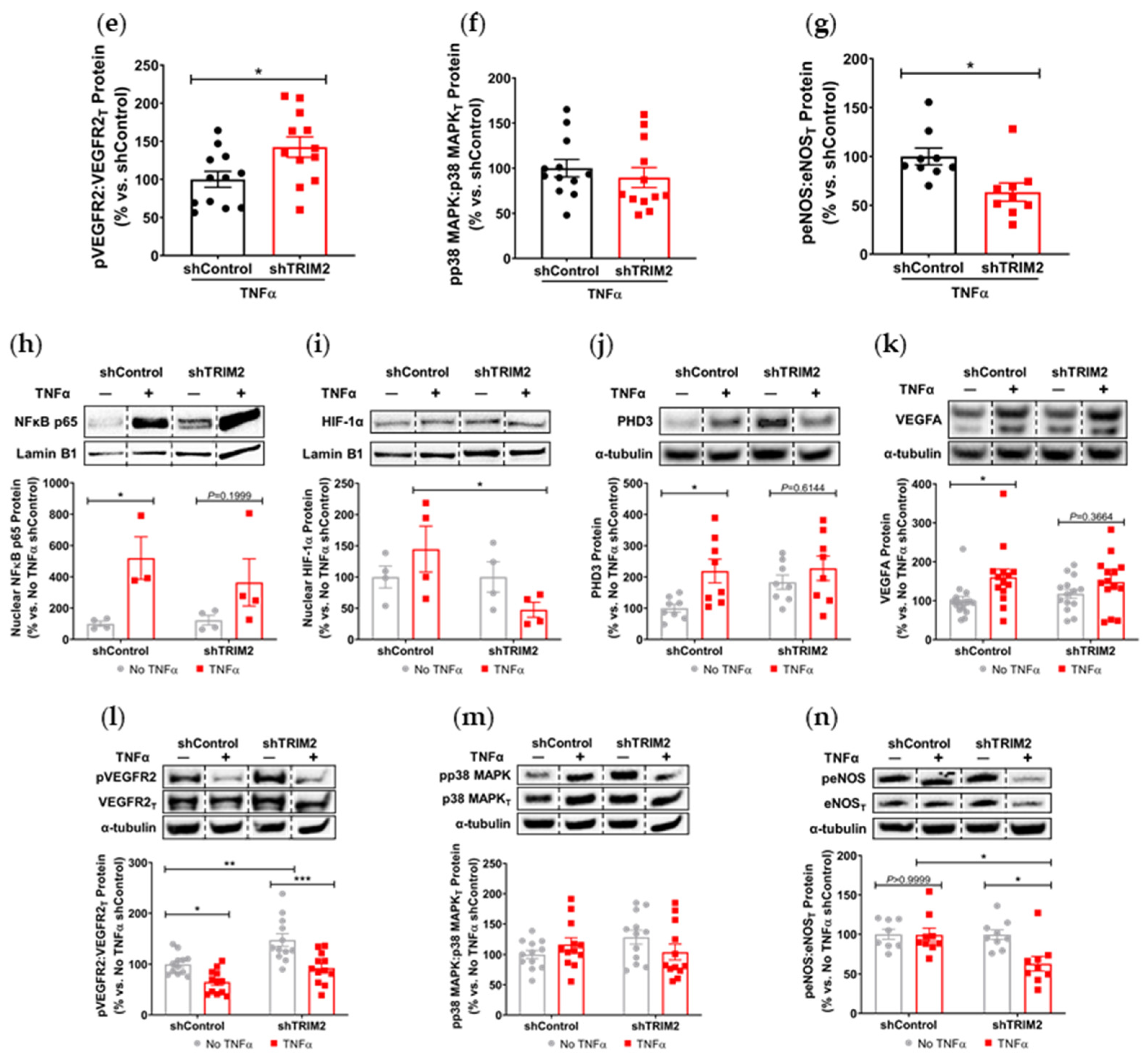
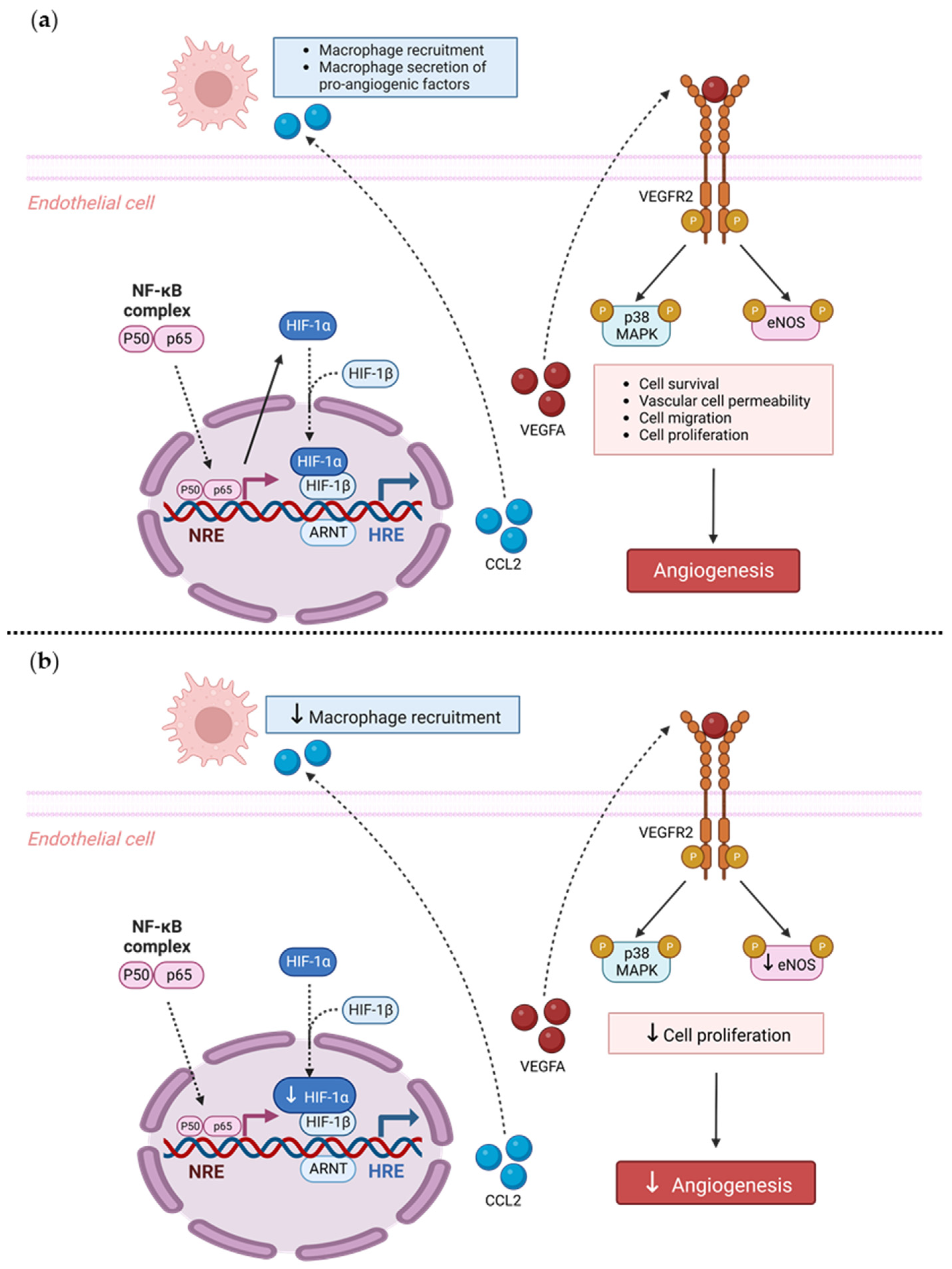
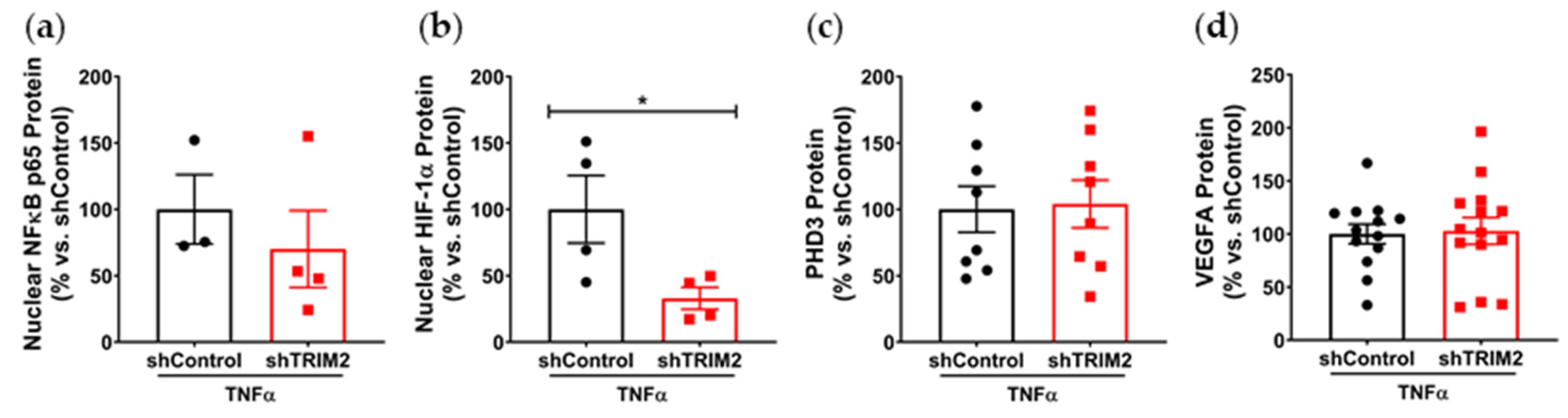
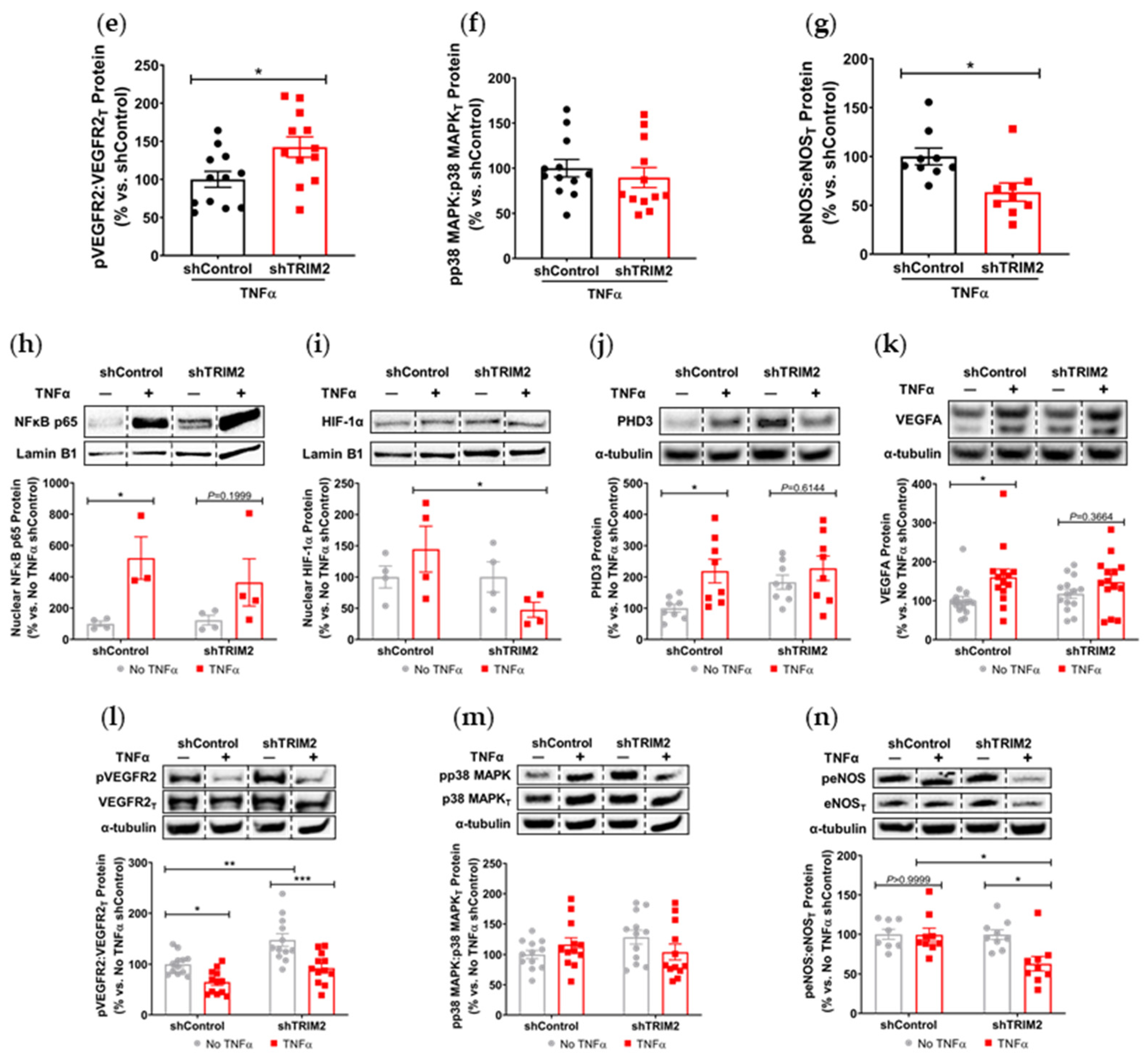
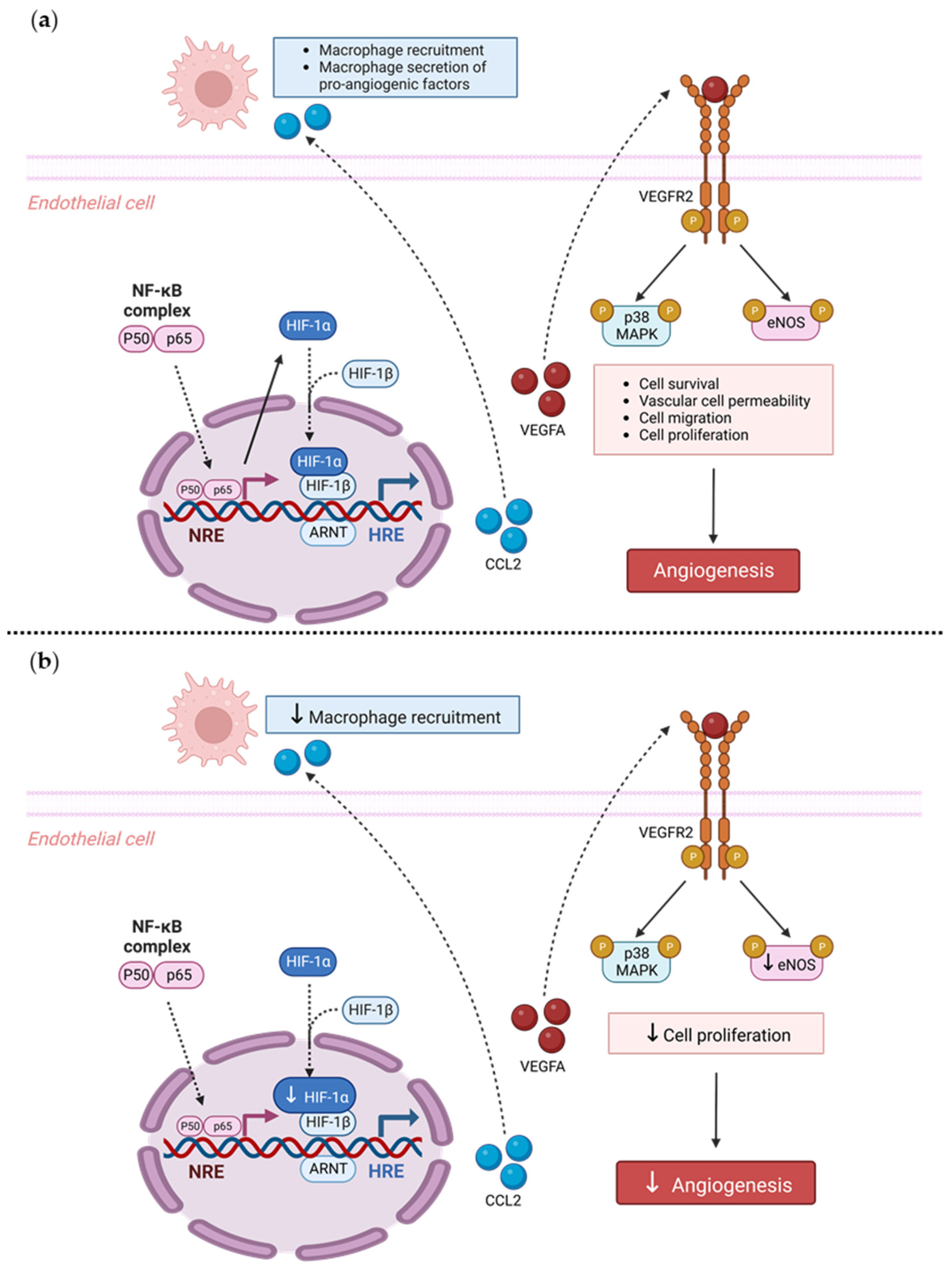
Our findings also suggest a potential mechanistic pathway by which TRIM2 may be directing angiogenic responses to inflammation. Direct comparisons of TNFα-stimulated cells in vitro showed a significant reduction in nuclear HIF-1α levels and eNOS activation. Intriguingly, when compared to their respective baseline unstimulated controls, we found that the extent of inflammatory induction of nuclear NF-κB, PHD3 and VEGFA were less pronounced in shTRIM2 cells. Under stimulation with cytokines like TNFα, it is possible that TRIM2, functioning as a ubiquitin ligase [
12], may contribute to the stabilization and nuclear translocation of the transcription factors NF-κB and HIF-1α, likely by promoting proteasomal degradation of their cytosolic inhibitors, such as PHD3 in the latter case [
13]. These transcription factors, in turn, promote VEGFA expression, which activates endothelial cell migration and tubule formation through numerous intracellular pathways [
14,
15], including the phosphorylation of p38 MAPK and eNOS. NF-κB p65 and HIF-1α may also stimulate angiogenesis downstream of TRIM2 by upregulating inflammatory cytokines and chemokines like CCL2, leading to the recruitment and activation of macrophages that help to potentiate the inflammation-driven angiogenic response.
It is not clear whether TRIM2 mediates VEGFA-related effects via VEGFR2, as only a modest decrease in Tyr
1175 phosphorylation was seen following TRIM2 knockdown. TRIM2 could be targeting alternative VEGFR2 phosphorylation sites. While there was no change in VEGFR2 phosphorylation at Tyr
1175, it is possible that other key tyrosine sites like Tyr
801, Tyr
1054 or Tyr
1059 may be involved, each of which may activate a distinct set of signal transduction mechanisms and cellular responses [
14]. Specifically, VEGFR2 phosphorylation at Tyr
801 also contributes to Akt-dependent eNOS activation and nitric oxide release from endothelial cells [
16,
17]. Another potential target of TRIM2 is neuropilin-1 (NRP1), one of the key co-receptors for VEGFR2. NRP1 is highly expressed in endothelial cells and neurons and can bind to both VEGFA and the class 3 semaphorins, a family of axonal guidance proteins, thus forming a key link between angiogenesis and neurogenesis [
15,
18]. Given the association of TRIM2 with axonal outgrowth and development [
12,
19], and now angiogenesis, it is possible that NRP1 and/or its semaphorin ligands may be involved in regulating angiogenic function by TRIM2. Further studies examining a possible link between NRP1 and TRIM2 would be useful to clarify the mechanistic pathway.
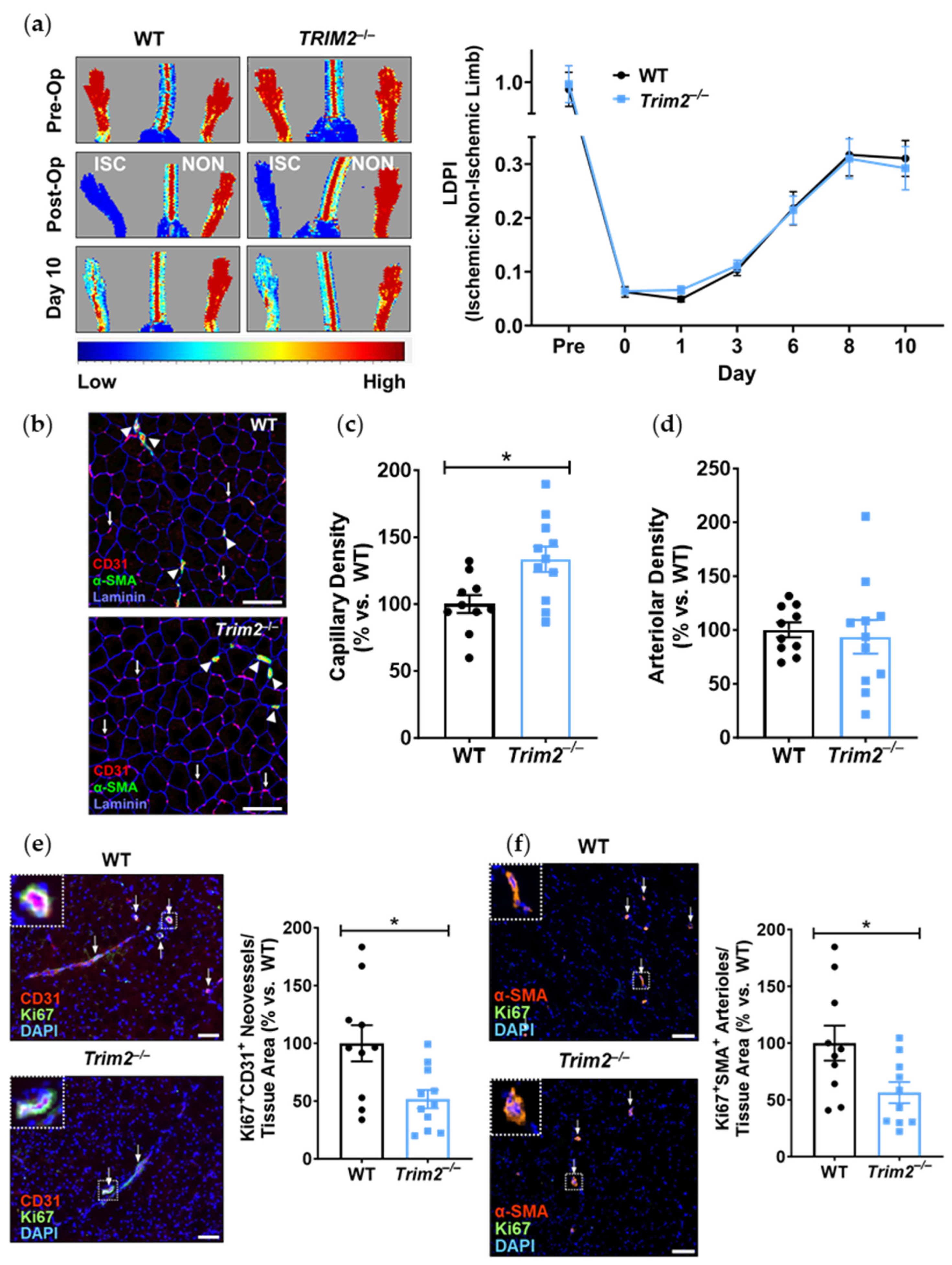
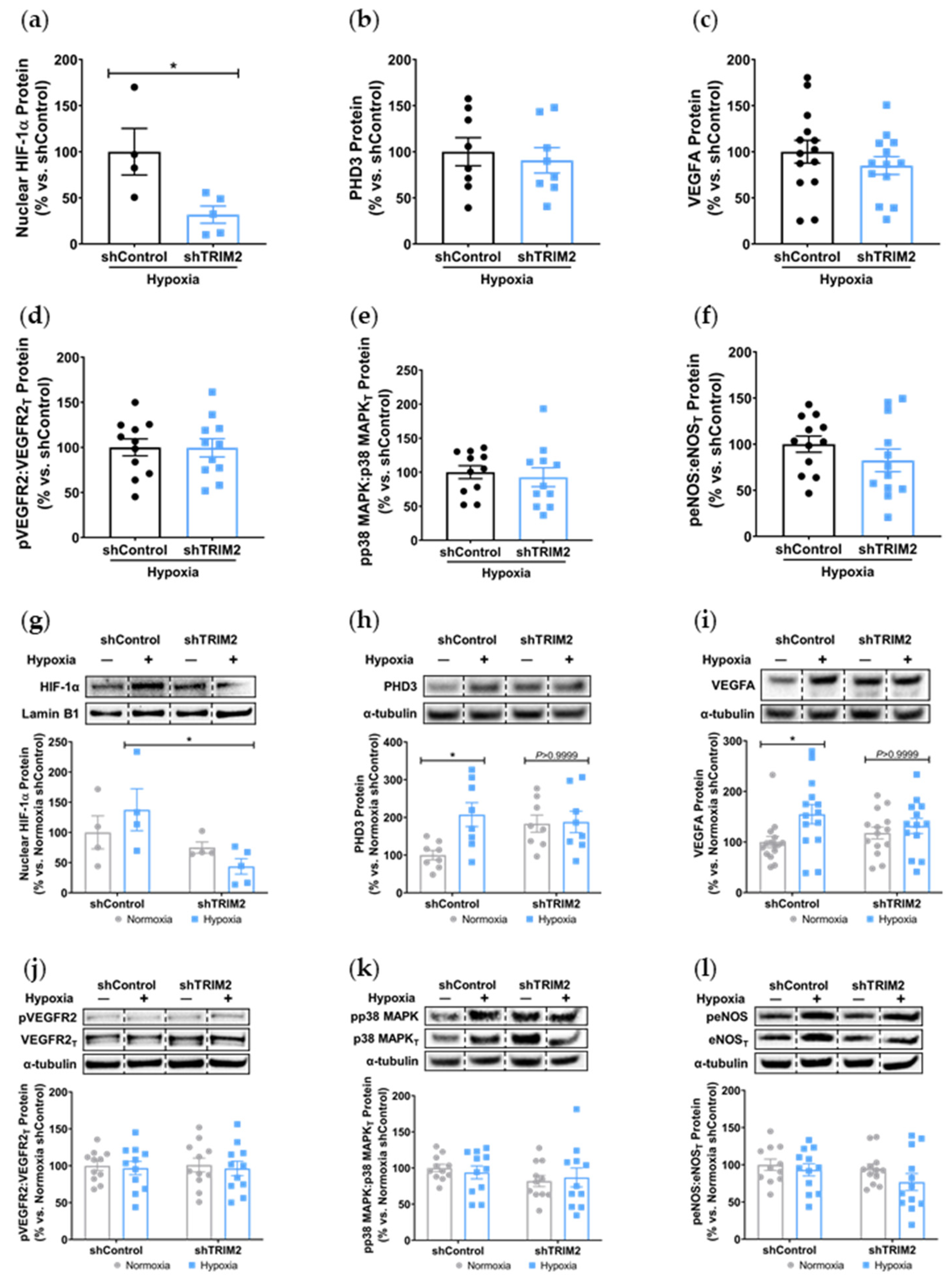
Consistent with our previous work which demonstrated impairment of hypoxia-stimulated tubule formation in vitro with TRIM2 knockdown [
7],
Trim2 deletion reduced the number of proliferating neovessels and arterioles in the ischemic tissue in vivo. Interestingly, there was an increase in total CD31
+ vessels in the ischemic hindlimbs of
Trim2−/− mice, suggestive of a potential negative feedback loop. In the late stages of angiogenesis, vessel pruning occurs, whereby capillaries disintegrate, to facilitate mature vessel formation during tissue remodeling [
20]. We therefore postulate that the changes seen with increased capillary density yet reduced number of proliferating neovessels may be indicative of vessel pruning at this late stage post-ischemia. Furthermore, revascularization in the hindlimb ischemia model is primarily facilitated through arteriogenesis [
21]. No differences were observed in overall arteriolar density, which is consistent with the lack of change seen in blood flow reperfusion. However, while there were significant changes at a cellular/tissue level, this did not seem to impact the recovery of blood flow reperfusion to the ischemic hindlimb. These incongruous findings may reflect inherent angiogenic compensatory mechanisms in vivo that may be activated in response to
Trim2 deletion, perhaps starting early in embryonic development and thereby rendering
Trim2 redundant. Mechanistically, while the hypoxia-driven increase in nuclear HIF-1α was attenuated in TRIM2-deficient HCAECs, downstream angiogenic signaling was not altered. The paucity of effects on these intracellular mediators may explain the lack of differences between WT and
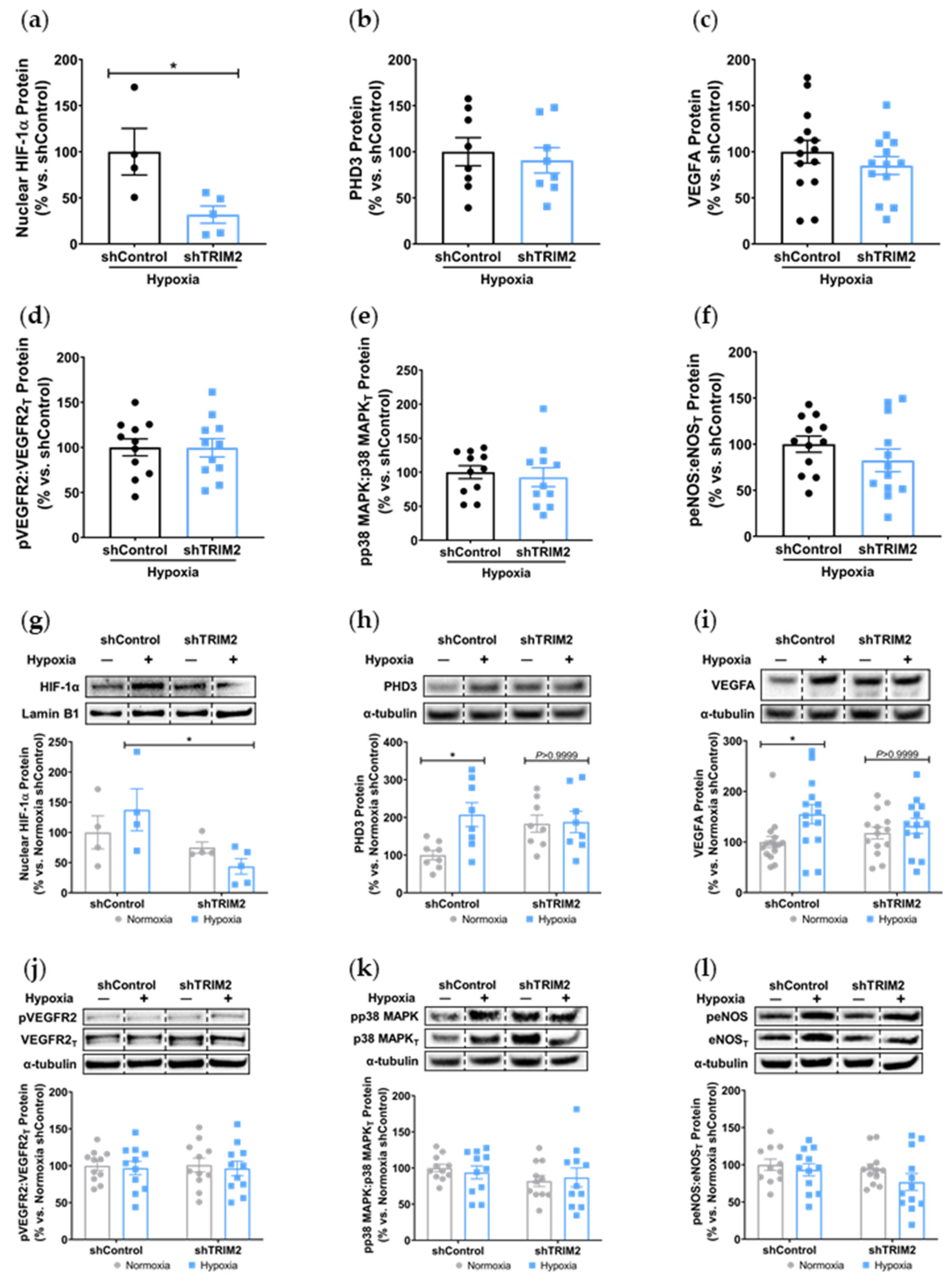
Trim2−/− mice in their angiogenic responses to hypoxia. This could prove clinically useful, as anti-TRIM2 therapies may be developed to suppress pathological inflammatory angiogenesis, without the adverse effects of impairing hypoxia-driven physiological angiogenesis.
The discordant in vitro and in vivo findings may also reflect the activation of compensatory angiogenic mechanisms in response to
Trim2 deletion in vivo. Future studies could explore inducible and endothelial cell-specific
Trim2 silencing to exclude such effects as angiogenic responses may be countered by
Trim2 deletion in other cell types, like vascular smooth muscle cells, pericytes and immune cells [
22]. It is also plausible that TRIM2 may target non-classical pathways downstream of VEGFA to confer its angiogenic effects in hypoxia, or it could be modulating other angiogenic factors like the fibroblast growth factors or angiopoietins. Future studies could evaluate a broader range of signaling targets to better elucidate the mechanistic basis by which TRIM2 may be modulating endothelial responses to hypoxia.
Overall, we have shown, for the first time, that TRIM2 is functionally important in regulating pathological angiogenic responses to inflammation in vivo, via modulation of classical angiogenic mediators HIF-1α, NF-κB p65, and VEGFA and downstream targets of VEGFA. Given that TRIM2 has no effect on physiological ischemia-driven angiogenesis, targeted TRIM2 inhibition could prove therapeutically useful for diseases driven predominantly by pathological angiogenesis including atherosclerosis and cancer, without the adverse effects of inhibiting physiological angiogenesis.
4. Materials and Methods
4.1. Animal Studies
All experimental procedures were conducted with approval from the SAHMRI Animal Ethics Committee (#SAM335) and conformed to the Australian Code for the Care and Use of Animals for Scientific Purposes (National Health and Medical Research Council, Australia). A Trim2−/− mouse line was generated by the South Australian Genome Editing facility using a CRISPR/Cas9 approach. In brief, Cas9 protein was injected into C57BL/6J murine embryos along with two guide RNA sequences. These guide RNAs were designed such that non-homologous end joining of the DNA following CRISPR/Cas9 activity would result in excision of a DNA fragment containing exon 2 of Trim2, leading to a frameshift in the coding sequence and an early stop codon in exon 3. The founder male carrying this mutant Trim2 allele was back-crossed to wildtype (WT) female C57BL/6J mice, generating identical heterozygous offspring which were subsequently crossed to generate homozygous Trim2 knockout (Trim2−/−) mice. Male wildtype (WT) and Trim2−/− mouse littermates were housed in a temperature and humidity-controlled environment under a 12 h light/dark cycle with ad libitum access to water and standard mouse chow. They underwent surgery at 8 weeks of age.
4.2. Plasma Glucose and Lipid Analyses
Plasma glucose concentrations were determined using a glucometer (Accu-Chek® Performa, Roche, Basel, Switzerland), while total plasma and HDL cholesterol concentrations were measured enzymatically (439-17501, Wako Diagnostics, Richmond, VA, USA). HDL cholesterol concentrations were determined following polyethylene glycol precipitation of apoB-containing lipoproteins, while LDL cholesterol concentrations were calculated by subtracting HDL from total cholesterol concentrations. Triglyceride concentrations were determined using a colorimetric assay (290-63701, Wako Diagnostics).
4.3. Periarterial Cuff Model
The femoral periarterial cuff model is an established model of inflammation-driven neointima formation and adventitial angiogenesis [
23,
24], processes which are known to contribute to atherosclerotic plaque development. A non-occlusive 2 mm length of polyethylene cuff was placed around the left femoral artery to trigger a localized inflammatory response, while a sham operation was performed on the right femoral artery as a parallel control. The animals were sacrificed 21 days post-surgery by overdose of isoflurane and intracardiac puncture, followed by perfusion with phosphate-buffered saline (PBS) via the left ventricle. The femoral arteries (complete with cuff) were excised for histochemical analyses.
Excised femoral arteries were fixed in 10% (v/v) formalin for 24 h then embedded in 3% (w/v) agarose prior to tissue processing and paraffin embedding. Angiogenic responses to cuff placement were assessed via immunohistochemistry on 5 μm sections, probing for CD68 (Bio-Rad, Hercules, CA, USA, Cat# MCA1957GA, RRID:AB_324217) to assess macrophage infiltration and CD31 (Abcam, Cambridge, UK, Cat# ab28365, RRID:AB_726365) to detect adventitial vessels. Proliferating neovessels were determined by co-staining tissue sections with proliferation marker Ki-67 (Thermo Fisher Scientific, Waltham, MA, USA, Cat# 14-5698-82, RRID:AB_10854564) and CD31. Endothelial tip cells were determined by staining sections with CD34 (Abcam, Cat# ab8158, RRID:AB_306316). Masson’s trichrome staining was performed with a Trichrome Stain Kit (ab150686, Abcam) to assess intima-to-media ratio as a measure of neointimal responses to inflammatory stimulation. All histological sections were photographed with a Zeiss Axio Scan.Z1 Digital Slide Scanner (Carl Zeiss Microscopy, Oberkochen, Baden-Württemberg, Germany), and image analysis was performed using Image-Pro Premier software (v9.0.4, Media Cybernetics, Rockville, MD, USA).
An additional cohort of mice underwent the same procedure and were sacrificed 24 h post-surgery for gene expression analysis. Total RNA was isolated from the femoral arteries with TRI® reagent (Sigma-Aldrich, St. Louis, MO, USA) and quantitated spectrophotometrically. Then, 200 ng of total RNA was reverse transcribed using the iScript cDNA synthesis kit (Bio-Rad). Quantitative real-time PCR was performed for Cd68 (F: 5′-GGACAGCTTACCTTTGGATTCAA-3′; R: 5′-CTGTGGGAAGGACACATTGTATTC-3′), Ccl2 (F: 5′-GCTGGAGCATCCACGTGTT-3′; R: 5′-ATCTTGCTGGTGAATGAGTAGCA-3′), NF-κB p65 (Rela, forward [F]: 5′-AGTATCCATAGCTTCCAGAACC-3′; reverse [R]: 5′-ACTGC-ATTCAAGTCATAGTCC-3′) and 36B4 (F: 5′-CAACGGCAGCA-TTTATAACCC-3′; R: 5′-CCCATTGATGATGGAGTGTGG-3′). Relative gene expression was calculated using the ΔΔCt method, normalized to 36B4 and WT non-cuffed arteries.
4.4. Hindlimb Ischemia Model
The hindlimb ischemia model is a well-validated model of physiological angiogenesis in response to tissue ischemia [
25]. Hindlimb ischemia was induced by ligation and excision of the left superficial and deep femoral arteries, along with the left femoral vein down to the saphenous artery. A sham procedure was performed on the contralateral hindlimb as a parallel control. Hindlimb blood reperfusion was determined by laser Doppler imaging (moorLDI2-IR, Moor Instruments, Devon, UK), performed prior to and immediately following surgery, then at days 1, 3, 6, 8 and 10 post-surgery. Animals were sacrificed 10 days post-surgery by isoflurane overdose and intracardiac puncture, and the gastrocnemius muscles of both hindlimbs were collected for histological analyses.
Gastrocnemius muscles from both ischemic and non-ischemic hindlimbs were OCT-embedded and frozen on dry ice. Sections were taken across the medial plane of the gastrocnemius muscle (anterior distal hindlimb). This region is known to provide the most consistent and uniform responses to ischemic induction [
25,
26,
27]. To histologically assess angiogenic responses to ischemia, immunofluorescence was performed on 5 µm tissue sections, staining with CD31 (Abcam, Cat# ab28364, RRID:AB_726362) to detect neovessels, α-smooth muscle actin (α-SMA, Sigma-Aldrich, Cat# F3777, RRID:AB_476977) to detect arterioles and laminin (Millipore, Burlington, MA, USA, Cat# MAB1905, RRID:AB_94392) to detect myocytes. Proliferating neovessels and arterioles were determined by co-staining tissue sections with Ki-67 (Thermo Fisher Scientific, Cat# 11-5698-82, RRID:AB_11151330) and either CD31 or α-SMA, respectively. Images were taken using an Eclipse Ni-E fluorescent microscope (Nikon Instruments, Tokyo, Japan). CD31
+ neovessels and α-SMA
+ arterioles were quantified using CellProfiler software (
www.cellprofiler.org, accessed on 9 January 2023, Broad Institute of MIT and Harvard, Boston, MA, USA), while the myocytes were manually quantified using ImageJ (
https://imagej.net/ij/, accessed on 9 January 2023, National Institutes of Health, Bethesda, MD, USA).
4.5. Lentiviral shRNA Knockdown of TRIM2 In Vitro
Human coronary artery endothelial cells (HCAECs, Cell Applications, San Diego, CA, USA) were cultured in MesoEndo Cell Growth Medium (212-500, Cell Applications) and used at passages 3–4. HCAECs were seeded at 5 × 10
4 cells/well in 6-well plates and cultured at 37 °C and 5% CO
2 overnight. The cells were exposed to 1 × 10
4 infectious units (IFU)/mL of lentiviral particles containing shRNA against TRIM2 (shTRIM2) or a random control sequence (shControl) for 24 h in the presence of polybrene. Transduced HCAECs were trypsinized, counted and seeded at a density of 1.5 × 10
5 cells/well and 8 × 10
4 cells/well for the inflammation and hypoxia experiments, respectively. HCAECs were then either incubated for 4.5 h with 0.6 ng/mL TNFα (to mimic inflammation) or for 6 h at 5% CO
2 and 1.2% O
2 balanced with N
2 (to mimic hypoxia). To measure phosphorylated proteins, HCAECs were stimulated with 10 ng/mL recombinant human VEGF
165 protein (R&D Systems) 15 min prior to harvest. Nuclear proteins were isolated from cell lysates using the NE-PER
® Nuclear and Cytoplasmic Extraction kit (Thermo Fisher Scientific). Whole-cell protein lysates were extracted using RIPA buffer [
7,
28]. Each experiment was performed at least four times independently with triplicates for each condition.
4.6. Protein Expression
Nuclear and whole-cell protein extracts were subjected to Western blot analysis and probed with primary antibodies for NF-κB p65 (Abcam, Cat# ab16502, RRID:AB_443394), HIF-1α (Novus Biologicals, Centennial, CO, USA, Cat# NB100-105, RRID:AB_10001154), PHD3 (Novus, Cat# NB100-303, RRID:AB_10003302), VEGFA (Abcam, Cat# ab46154, RRID:AB_2212642), phosphorylated (Tyr1175) VEGFR2 (Cell Signaling Technology, Danvers, MA, USA, Cat# 2478, RRID:AB_331377), total VEGFR2 (Cell Signaling Technology, Cat# 2479, RRID:AB_2212507), phosphorylated (Thr180/Tyr182) p38 MAPK (Cell Signaling Technology, Cat# 4511, RRID:AB_2139682), total p38 MAPK (Cell Signaling Technology, Cat# 8690, RRID:AB_10999090), phosphorylated (Ser1177) eNOS (BD Biosciences, Franklin Lakes, NJ, USA, Cat# 612393, RRID:AB_399751) and total eNOS (BD Biosciences, Cat# 610297, RRID:AB_397691). Even protein loading was confirmed with lamin B1 (Abcam, Cat# ab16048, RRID:AB_443298) for nuclear fractions or α-tubulin (Abcam, Cat# ab40742, RRID:AB_880625) for whole-cell lysates.
4.7. Statistics
Data are expressed as mean ± SEM. Comparisons were made using unpaired Student’s t-tests or two-way ANOVA followed by post hoc analysis using Bonferroni’s multiple comparison tests. Significance was set at a two-sided p < 0.05.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}