

In Vitro Transcriptional Response of Eimeria tenella to Toltrazuril Reveals That Oxidative Stress and Autophagy Contribute to Its Anticoccidial Effect


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Overview of Sequencing Data and Gene Expression Patterns
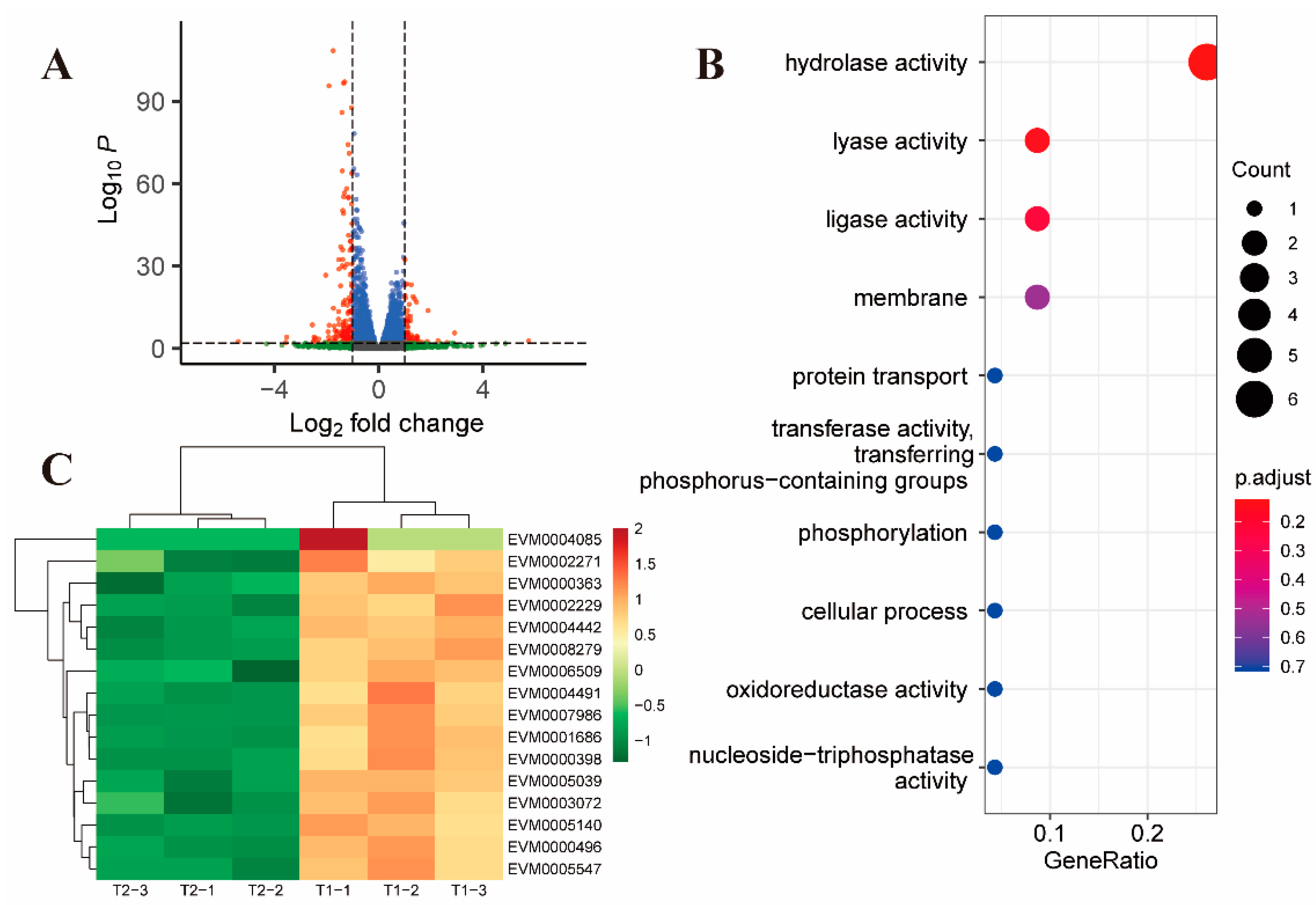
2.2. Gene Alterations in E. tenella in Response to Toltrazuril Treatment
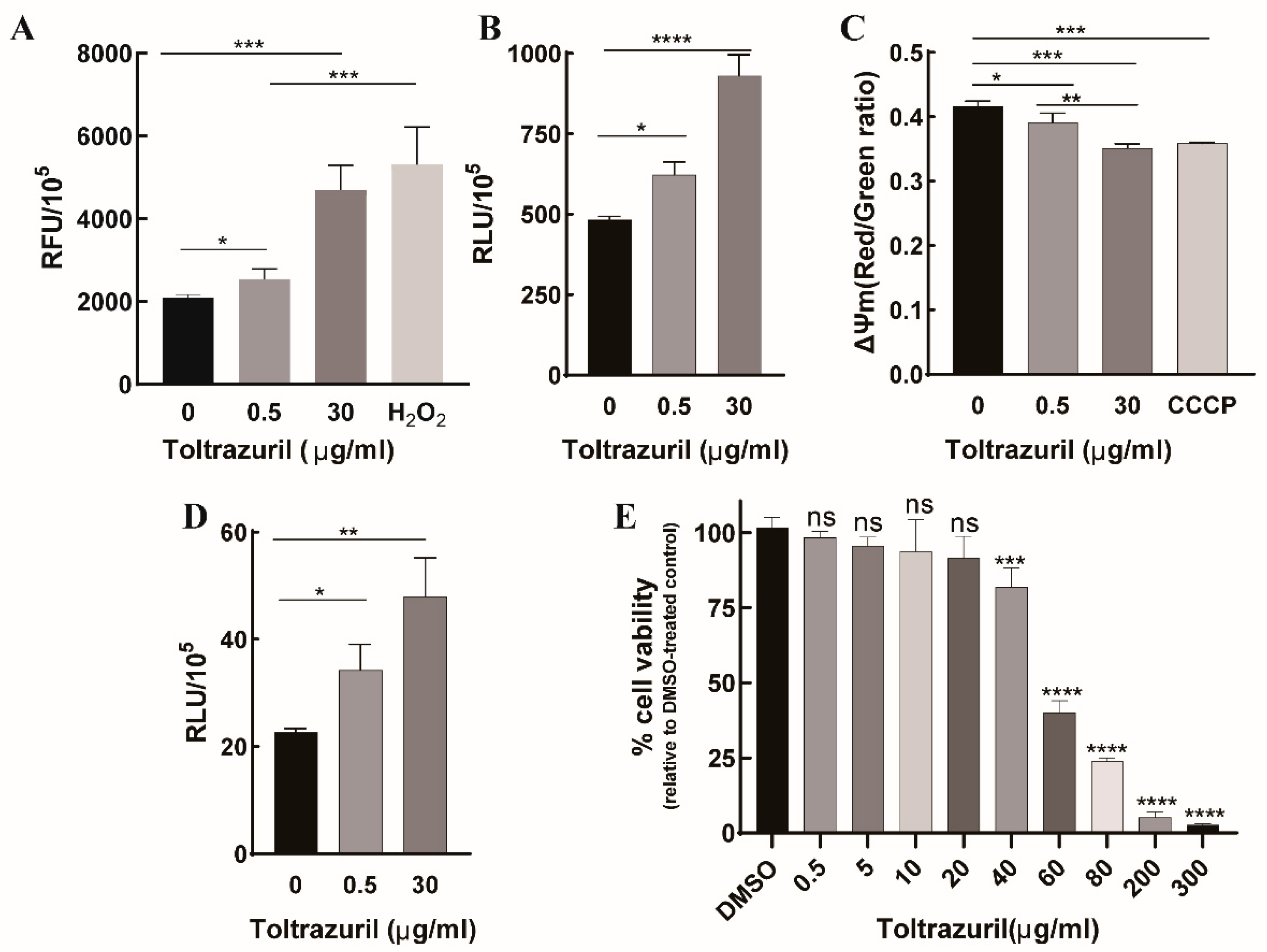
2.3. Toltrazuril Induces Intracellular Oxidative Stress and Autophagy in Parasites
3. Discussion
4. Materials and Methods
4.1. Animals and Parasites
4.2. Compounds
4.3. Merozoites Purification and Toltrazuril Treatment
4.4. RNA Extraction, Library Preparation and RNA-Seq
4.5. Bioinformatics
4.6. Real-Time PCR
4.7. Measurement of Intracellular ROS Levels
4.8. Mitochondrial ROS (mROS) Measurement
4.9. Measurement of Mitochondrial Membrane Potential
4.10. Autophagy Detection
4.11. Cytotoxicity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayer, J.; Donnelly, T.M. (Eds.) Coccidiosis. In Clinical Veterinary Advisor; W.B. Saunders: Saint Louis, MO, USA, 2013; pp. 346–348. [Google Scholar]
- Chaves Hernández, A.J. Poultry and Avian Diseases. In Encyclopedia of Agriculture and Food Systems; Van Alfen, N.K., Ed.; Academic Press: Oxford, UK, 2014; pp. 504–520. [Google Scholar]
- Cunha, A.F.d.; Santin, E.; Kogut, M. Editorial: Poultry Coccidiosis: Strategies to Understand and Control. Front. Vet. Sci. 2020, 7, 599322. [Google Scholar] [CrossRef]
- Chapman, H.D.; Barta, J.R.; Blake, D.; Gruber, A.; Jenkins, M.; Smith, N.C.; Suo, X.; Tomley, F.M. A selective review of advances in coccidiosis research. Adv. Parasitol. 2013, 83, 93–171. [Google Scholar] [CrossRef]
- Sander, V.A.; Sánchez López, E.F.; Mendoza Morales, L.; Ramos Duarte, V.A.; Corigliano, M.G.; Clemente, M. Use of Veterinary Vaccines for Livestock as a Strategy to Control Foodborne Parasitic Diseases. Front. Cell Infect. Microbiol. 2020, 10, 288. [Google Scholar] [CrossRef]
- Quiroz-Castañeda, R.E.; Dantán-González, E. Control of avian coccidiosis: Future and present natural alternatives. Biomed Res. Int. 2015, 2015, 430610. [Google Scholar] [CrossRef]
- Fatoba, A.J.; Adeleke, M.A. Transgenic Eimeria parasite: A potential control strategy for chicken coccidiosis. Acta Trop. 2020, 205, 105417. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Iqbal, Z.; Blake, D.; Khan, M.N.; Saleemi, M.K. Anticoccidial drug resistance in fowl coccidia: The state of play revisited. Worlds Poult. Sci. J. 2011, 67, 337–350. [Google Scholar] [CrossRef]
- Ojimelukwe, A.E.; Emedhem, D.E.; Agu, G.O.; Nduka, F.O.; Abah, A.E. Populations of Eimeria tenella express resistance to commonly used anticoccidial drugs in southern Nigeria. Int. J. Vet. Sci. Med. 2018, 6, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.A.; Nguyen, B.T.; Cammayo, P.L.T.; Võ, T.C.; Naw, H.; Kim, S.; Kim, W.H.; Na, B.K.; Min, W. Epidemiological investigation and drug resistance of Eimeria species in Korean chicken farms. BMC Vet. Res. 2022, 18, 277. [Google Scholar] [CrossRef] [PubMed]
- Blake, D.P.; Tomley, F.M. Securing poultry production from the ever-present Eimeria challenge. Trends Parasitol. 2014, 30, 12–19. [Google Scholar] [CrossRef]
- Stock, M.L.; Elazab, S.T.; Hsu, W.H. Review of triazine antiprotozoal drugs used in veterinary medicine. J. Vet. Pharmacol. Ther. 2018, 41, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, X.; Zhao, Q.; She, R.; Xia, S.; Zhang, K.; Zhang, L.; Wang, X.; Wang, M.; Liu, Y.; et al. Anticoccidial activity of novel triazine compounds in broiler chickens. Vet. Parasitol. 2019, 267, 4–8. [Google Scholar] [CrossRef]
- Noack, S.; Chapman, H.D.; Selzer, P.M. Anticoccidial drugs of the livestock industry. Parasitol. Res. 2019, 118, 2009–2026. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Ortmann-Falkenstein, G.; Haberkorn, A. The effects of sym. Triazinones on developmental stages of Eimeria tenella, E. maxima and E. acervulina: A light and electron microscopical study. Z. Parasitenkd. 1984, 70, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Wang, H.; Xue, F.; Wang, X.; Fei, C.; Wang, M.; Zhang, T.; Yao, X.; He, P. Effects of diclazuril on apoptosis and mitochondrial transmembrane potential in second-generation merozoites of Eimeria tenella. Vet. Parasitol. 2010, 168, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Darius, A.K.; Mehlhorn, H.; Heydorn, A.O. Effects of toltrazuril and ponazuril on the fine structure and multiplication of tachyzoites of the NC-1 strain of Neospora caninum (a synonym of Hammondia heydorni) in cell cultures. Parasitol. Res. 2004, 92, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Bierbaum, S. Das Cyclophilin EtCyp20.5 aus Eimeria tenella als Rezeptor für das Antikokzidium Toltrazuril. Ph.D. Thesis, Düsseldorf University, Düsseldorf, Germany, 2010. [Google Scholar]
- Vacanti, N.M.; Divakaruni, A.S.; Green, C.R.; Parker, S.J.; Henry, R.R.; Ciaraldi, T.P.; Murphy, A.N.; Metallo, C.M. Regulation of Substrate Utilization by the Mitochondrial Pyruvate Carrier. Mol. Cell 2014, 56, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Jian, C.; Xu, F.; Hou, T.; Sun, T.; Li, J.; Cheng, H.; Wang, X. Deficiency of PHB complex impairs respiratory supercomplex formation and activates mitochondrial flashes. J. Cell Sci. 2017, 130, 2620–2630. [Google Scholar] [CrossRef]
- Matz, J.M.; Goosmann, C.; Matuschewski, K.; Kooij, T.W.A. An Unusual Prohibitin Regulates Malaria Parasite Mitochondrial Membrane Potential. Cell Rep. 2018, 23, 756–767. [Google Scholar] [CrossRef]
- Tavares, A.; Gonçalves, J.; Florindo, C.; Tavares, A.A.; Soares, H. Mob1: Defining cell polarity for proper cell division. J. Cell Sci. 2012, 125, 516–527. [Google Scholar] [CrossRef]
- Delgado, I.L.S.; Tavares, A.; Francisco, S.; Santos, D.; Coelho, J.; Basto, A.P.; Zúquete, S.; Müller, J.; Hemphill, A.; Meissner, M.; et al. Characterization of a MOB1 Homolog in the Apicomplexan Parasite Toxoplasma gondii. Biology 2021, 10, 1233. [Google Scholar] [CrossRef]
- Melo-Hanchuk, T.D.; Slepicka, P.F.; Meirelles, G.V.; Basei, F.L.; Lovato, D.V.; Granato, D.C.; Pauletti, B.A.; Domingues, R.R.; Leme, A.F.P.; Pelegrini, A.L.; et al. NEK1 kinase domain structure and its dynamic protein interactome after exposure to Cisplatin. Sci. Rep. 2017, 7, 5445. [Google Scholar] [CrossRef] [PubMed]
- Vitulo, N.; Vezzi, A.; Galla, G.; Citterio, S.; Marino, G.; Ruperti, B.; Zermiani, M.; Albertini, E.; Valle, G.; Barcaccia, G. Characterization and evolution of the cell cycle-associated mob domain-containing proteins in eukaryotes. Evol. Bioinform. Online 2007, 3, 121–158. [Google Scholar] [CrossRef] [PubMed]
- Hammarton, T.C.; Lillico, S.G.; Welburn, S.C.; Mottram, J.C. Trypanosoma brucei MOB1 is required for accurate and efficient cytokinesis but not for exit from mitosis. Mol. Microbiol. 2005, 56, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, M.J.; Lehmann, M.; Muthalagi, M.; Jerome, M.E.; Brooks, C.F.; Szatanek, T.; Flynn, J.; Parrot, B.; Radke, J.; Striepen, B.; et al. Forward genetic analysis of the apicomplexan cell division cycle in Toxoplasma gondii. PLoS Pathog. 2008, 4, e36. [Google Scholar] [CrossRef]
- Alan, P.; Vandevoorde, K.R.; Joshi, B.; Cardoen, B.; Gao, G.; Mohammadzadeh, Y.; Hamarneh, G.; Nabi, I.R. Basal Gp78-dependent mitophagy promotes mitochondrial health and limits mitochondrial ROS. Cell Mol. Life Sci. 2022, 79, 565. [Google Scholar] [CrossRef]
- Lei, Y.; Tang, D.; Liao, G.; Xu, L.; Liu, S.; Chen, Q.; Li, C.; Duan, J.; Wang, K.; Wang, J.; et al. The crystal structure of Atg18 reveals a new binding site for Atg2 in Saccharomyces cerevisiae. Cell. Mol. Life Sci. 2021, 78, 2131–2143. [Google Scholar] [CrossRef]
- Zheng, W.; Xie, W.; Yin, D.; Luo, R.; Liu, M.; Guo, F. ATG5 and ATG7 induced autophagy interplays with UPR via PERK signaling. Cell Commun. Signal. 2019, 17, 42. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.Y.; Liu, Y.J.; Feng, J.; Wu, Y.; Shen, H.M.; Lu, G.D. Full-coverage regulations of autophagy by ROS: From induction to maturation. Autophagy 2022, 18, 1240–1255. [Google Scholar] [CrossRef]
- Kma, L.; Baruah, T.J. The interplay of ROS and the PI3K/Akt pathway in autophagy regulation. Biotechnol. Appl. Biochem. 2022, 69, 248–264. [Google Scholar] [CrossRef]
- Blake, D.P.; Knox, J.; Dehaeck, B.; Huntington, B.; Rathinam, T.; Ravipati, V.; Ayoade, S.; Gilbert, W.; Adebambo, A.O.; Jatau, I.D.; et al. Re-calculating the cost of coccidiosis in chickens. Vet. Res. 2020, 51, 115. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Rippey, N.S.; Toivio-Kinnucan, M.A.; Blagburn, B.L. Ultrastructural effects of diclazuril against Toxoplasma gondii and investigation of a diclazuril-resistant mutant. J. Parasitol. 1995, 81, 459–466. [Google Scholar] [CrossRef]
- Maes, L.; Coussement, W.; Vanparijs, O.; Marsboom, R. In vivo action of the anticoccidial diclazuril (Clinacox) on the developmental stages of Eimeria tenella: A histological study. J. Parasitol. 1988, 74, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, A.; Maes, L.; Coussement, W.; Vanparijs, O.; Lauwers, F.; Vlaminckx, E.; Borgers, M.; Marsboom, R. In vivo action of the anticoccidial diclazuril (Clinacox) on the developmental stages of Eimeria tenella: An ultrastructural evaluation. J. Parasitol. 1988, 74, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Verheyen, A.; Maes, L.; Coussement, W.; Vanparijs, O.; Lauwers, F.; Vlaminckx, E.; Marsboom, R. Ultrastructural evaluation of the effects of diclazuril on the endogenous stages of Eimeria maxima and E. brunetti in experimentally inoculated chickens. Parasitol. Res. 1989, 75, 604–610. [Google Scholar] [CrossRef]
- Melin, J.; Schulz, C.; Wrobel, L.; Bernhard, O.; Chacinska, A.; Jahn, O.; Schmidt, B.; Rehling, P. Presequence recognition by the tom40 channel contributes to precursor translocation into the mitochondrial matrix. Mol. Cell Biol. 2014, 34, 3473–3485. [Google Scholar] [CrossRef] [PubMed]
- Chacinska, A.; Koehler, C.M.; Milenkovic, D.; Lithgow, T.; Pfanner, N. Importing mitochondrial proteins: Machineries and mechanisms. Cell 2009, 138, 628–644. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, J.B.; Brunstein, M.E.; Bozkurt, S.; Alves, L.; Wegner, M.; Kaulich, M.; Pohl, C.; Münch, C. Protein import motor complex reacts to mitochondrial misfolding by reducing protein import and activating mitophagy. Nat. Commun. 2022, 13, 5164. [Google Scholar] [CrossRef]
- Radheshyam, M.; Madhulika, N. Superoxide Dismutase: A Key Enzyme for the Survival of Intracellular Pathogens in Host. In Reactive Oxygen Species; Rizwan, A., Ed.; IntechOpen: Rijeka, Croatia, 2021; Chapter 3. [Google Scholar]
- Couto, N.; Wood, J.; Barber, J. The role of glutathione reductase and related enzymes on cellular redox homoeostasis network. Free Radic. Biol. Med. 2016, 95, 27–42. [Google Scholar] [CrossRef]
- Bosch, S.S.; Kronenberger, T.; Meissner, K.A.; Zimbres, F.M.; Stegehake, D.; Izui, N.M.; Schettert, I.; Liebau, E.; Wrenger, C. Oxidative Stress Control by Apicomplexan Parasites. Biomed. Res. Int. 2015, 2015, 351289. [Google Scholar] [CrossRef]
- Labbé, M.; Péroval, M.; Bourdieu, C.; Girard-Misguich, F.; Péry, P. Eimeria tenella enolase and pyruvate kinase: A likely role in glycolysis and in others functions. Int. J. Parasitol. 2006, 36, 1443–1452. [Google Scholar] [CrossRef]
- Liu, L.-l.; Chen, Z.-g.; Mi, R.-s.; Zhang, K.-y.; Liu, Y.-c.; Jiang, W.; Fei, C.-z.; Xue, F.-q.; Li, T. Effect of Acetamizuril on enolase in second-generation merozoites of Eimeria tenella. Vet. Parasitol. 2016, 215, 88–91. [Google Scholar] [CrossRef]
- Zhou, B.-h.; Ding, H.-y.; Yang, J.-y.; Chai, J.; Guo, H.-w.; Wang, H.-w. Effects of diclazuril on the expression of enolase in second-generation merozoites of Eimeria tenella. Poult. Sci. 2020, 99, 6402–6409. [Google Scholar] [CrossRef]
- Szatmári, Z.; Sass, M. The autophagic roles of Rab small GTPases and their upstream regulators: A review. Autophagy 2014, 10, 1154–1166. [Google Scholar] [CrossRef]
- Hyttinen, J.M.; Niittykoski, M.; Salminen, A.; Kaarniranta, K. Maturation of autophagosomes and endosomes: A key role for Rab7. Biochim. Biophys. Acta 2013, 1833, 503–510. [Google Scholar] [CrossRef]
- Nath, S.; Patra, D.; Nag, A.; Kundu, R. Sesquiterpenoid-rich Java Ginger rhizome extract prompts autophagic cell death in cervical cancer cell SiHa mainly by modulating cellular redox homeostasis. 3 Biotech 2023, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Uribe, P.; Meriño, J.; Matus, C.E.; Schulz, M.; Zambrano, F.; Villegas, J.V.; Conejeros, I.; Taubert, A.; Hermosilla, C.; Sánchez, R. Autophagy is activated in human spermatozoa subjected to oxidative stress and its inhibition impairs sperm quality and promotes cell death. Hum. Reprod. 2022, 37, 680–695. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhao, Y.; Ding, Y.; Ruan, Y.; Li, X.; Zhou, Q.; Zhou, Y.; Zhang, C.; Hu, L.; Zhao, X.; et al. Autophagy Ameliorates Reactive Oxygen Species-Induced Platelet Storage Lesions. Oxid. Med. Cell Longev. 2022, 2022, 1898844. [Google Scholar] [CrossRef] [PubMed]
- Eckert, J.; Braun, R.; Shirley, M.; Coudert, P. Biotechnology: Guidelines on Techniques in Coccidiosis Research COST European Cooperation in the Field of Scientific and Technical Research; The European Cooperation in Science and Technology: Brussels, Belgium, 1995. [Google Scholar]
- Schwarz, R.S.; Fetterer, R.H.; Rosenberg, G.H.; Miska, K.B. Coccidian merozoite transcriptome analysis from Eimeria maxima in comparison to Eimeria tenella and Eimeria acervulina. J. Parasitol. 2010, 96, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, L.; Si, H.; Liu, X.; Suo, X.; Hu, D. Early Transcriptional Response to Monensin in Sensitive and Resistant Strains of Eimeria tenella. Front. Microbiol. 2022, 13, 934153. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, H.; Du, S.; Song, X.; Hu, D. In Vitro Transcriptional Response of Eimeria tenella to Toltrazuril Reveals That Oxidative Stress and Autophagy Contribute to Its Anticoccidial Effect. Int. J. Mol. Sci. 2023, 24, 8370. https://doi.org/10.3390/ijms24098370
Zhang L, Zhang H, Du S, Song X, Hu D. In Vitro Transcriptional Response of Eimeria tenella to Toltrazuril Reveals That Oxidative Stress and Autophagy Contribute to Its Anticoccidial Effect. International Journal of Molecular Sciences. 2023; 24(9):8370. https://doi.org/10.3390/ijms24098370
Chicago/Turabian StyleZhang, Lei, Hongtao Zhang, Shiqi Du, Xingju Song, and Dandan Hu. 2023. "In Vitro Transcriptional Response of Eimeria tenella to Toltrazuril Reveals That Oxidative Stress and Autophagy Contribute to Its Anticoccidial Effect" International Journal of Molecular Sciences 24, no. 9: 8370. https://doi.org/10.3390/ijms24098370
APA StyleZhang, L., Zhang, H., Du, S., Song, X., & Hu, D. (2023). In Vitro Transcriptional Response of Eimeria tenella to Toltrazuril Reveals That Oxidative Stress and Autophagy Contribute to Its Anticoccidial Effect. International Journal of Molecular Sciences, 24(9), 8370. https://doi.org/10.3390/ijms24098370