
Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases


Abstract
1. Introduction
2. Characteristics and Diseases Related to the Adenylate Kinase Isozymes
2.1. AK1
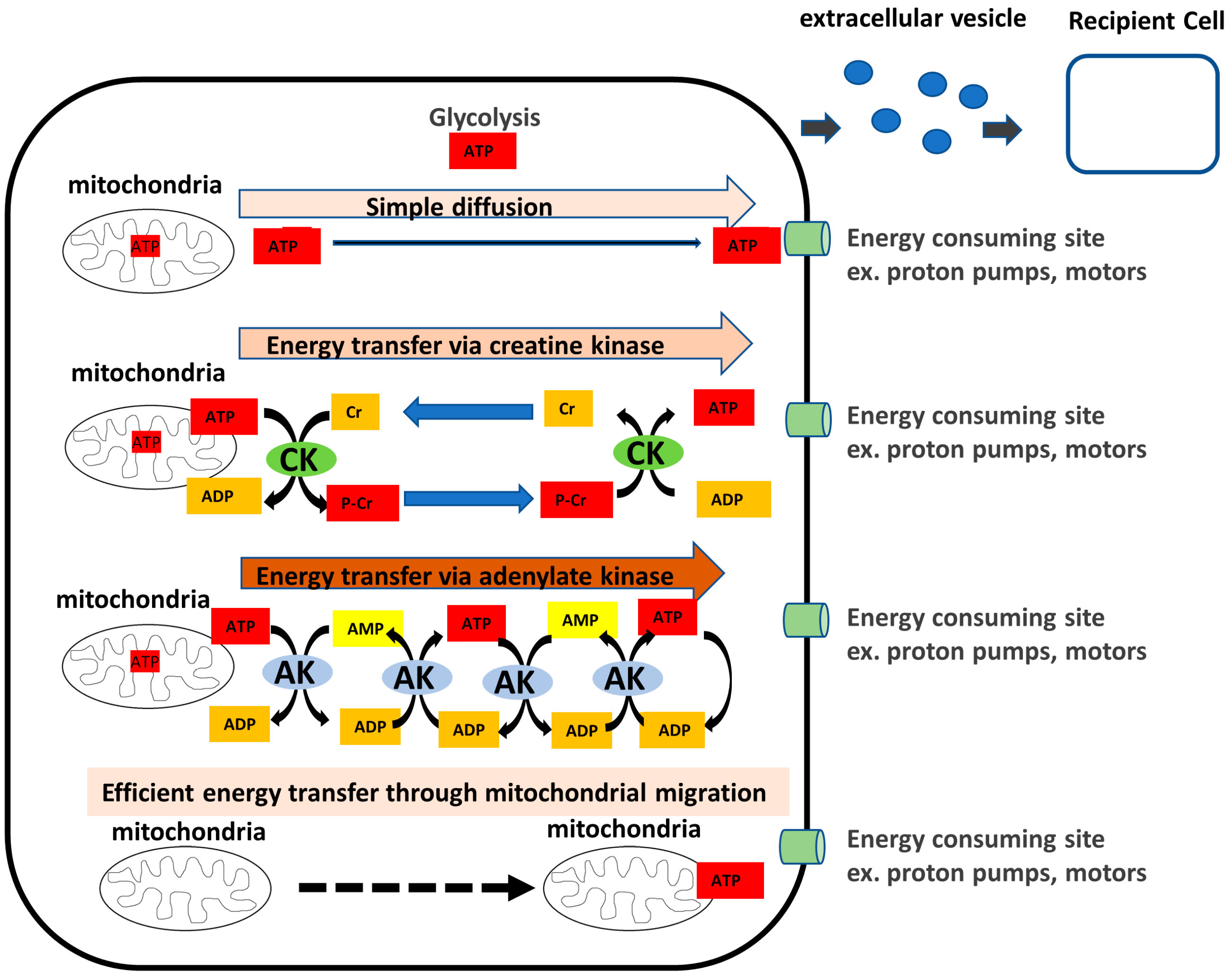
2.1.1. AK1 Exists in the Cytoplasm and in the Extracellular Space
2.1.2. AK1 Deficiency Causes Hematological Abnormalities
2.1.3. AK1 Expression Decreases in Non-Obstructive Azoospermia Patients
2.1.4. AK1 Expression Levels Are Associated with Behavioral Activity in an Animal Model
2.1.5. AK1 Knockout Affects Muscles, Heart Muscle, and Sperms
2.1.6. AK Isozymes and Other Enzymes Complement Each Other under AK1 Deficiency
2.2. AK2
2.2.1. AK2 Regulates Energy Metabolism in the Mitochondrial Intermembrane Space
2.2.2. AK2 Deficiency Causes Reticular Dysgenesis
2.2.3. AK2 Knockout in Animals Is Lethal
2.2.4. AK2 Expression Shows Circadian Variation
2.2.5. AK2 Suppresses Cancer
2.2.6. AK2 Promotes Cancer
2.3. AK3
2.3.1. AK3 Regulates Energy Metabolism in the Mitochondrial Matrix
2.3.2. AK3 Expression Level Is Associated with Survival in Cancer Patients
2.4. AK4
2.4.1. AK4 Is Involved in Oxidative Stress
2.4.2. AK4 Regulates Mitochondrial Activity under Hypoxia
2.4.3. AK4 Is Involved in Drug Resistance of Cancer Cells
2.4.4. AK4 Is a Useful Prognostic Marker for Cancer
2.4.5. AK4 Pseudogene Is Abundant in Circulating Exosomes in Patients with Pancreatic Adenocarcinoma
2.5. AK5
2.5.1. Anti-AK5 Antibodies Are Associated with Non-Paraneoplastic Limbic Encephalitis
2.5.2. AK5 Suppresses Cancer and Cell Growth through the AMPK/mTOR Pathway
2.6. AK6
2.6.1. AK6 Is Involved in Energy Metabolism in the Nucleus
2.6.2. AK6 Is Associated with Poor Cancer Prognosis
2.7. AK7
2.7.1. AK7 Deficiency Causes Primary Ciliary Dyskinesia and Infertility
2.7.2. AK7 Knockout Mice Exhibit Primary Ciliary Dyskinesia
2.7.3. AK7 Is A Useful Prognostic Marker for Ovarian Cancer
2.8. AK8
2.8.1. AK8 Is Associated with the Axoneme
2.8.2. AK8 Knockout Causes Hydrocephalus
2.9. AK9

{kind=link}
| Isozyme | Intracellular Localization/ Highly Expressed Tissues | Physiological Characteristics | Related Human Diseases | Animal Model/Cultured Cell |
|---|---|---|---|---|
| AK1 | Cytosol/nuscle, brain, heart, testis | Highly expressed in skeletal muscles, brain, and erythrocytes [1] | Non-obstructive azoospermia [30] Hematological abnormality [20,21,22] | KO mouse, inhibited sperm motility under specific conditions [30,33,99] Tg medaka, increased spontaneous locomotor activity [32] and reduced tolerance to ischemic stress [35] |
| AK2 | Mitochondrial intermembrane space/liver, kidney, heart | Supply of ADP to the adenine nucleotide transporter [100] Regulation of mitochondrial apoptosis [39] Transfer of energy from the mitochondria [36] | Reticular dysgenesis [40,41,42,47] | KO drosophila, lethal at the larval stage [1] KO zebrafish, reticular dysgenesis, and affected sensory hair cells [48] KO mouse, lethal in the early embryo [46] |
| AK3 | Mitochondrial matrix/all tissues except for red blood cell | Mitochondrial energy metabolism [1] Low expression in pancreatic beta cells [60] | Not reported | HeLa cell, slow growth [61] |
| AK4 | Mitochondrial matrix/kidney, liver, brain, heart, stomach, intestine, gonads | Mitochondrial energy metabolism [6,67] Hypoxia-related mitochondrial regulation [67,69,70] | Not reported | Smooth muscle cells, AK4 KD enhanced mitochondrial respiration and decreased glycolytic metabolism [68] Hela cell, AK4 KD decreased mitochondrial activity [70] |
| AK5 | Cytosol nucleus/highly expressed in brain | Energy regulation in brain [76] | Non-paraneoplastic limbic encephalitis [77] | AZ521 and MKN74 cells, AK5 KD inhibited growth and autophagy and increased apoptosis [80] |
| AK6 | Nucleus/all tissues | Nuclear energy metabolism [81] | Not reported | C. elegans, growth inhibition [82,83] |
| AK7 | Cytosol/lung, trachea, testis, mammary gland tissue | Expressed in ciliated tissues and testes [3] | Respiratory primary ciliary dyskinesia [91] Morphological anomalies of the flagella [92] Multiple morphological anomalies of the flagella, Oligoasthenoteratozoospermia [93] | KO mouse, primary ciliary dyskinesia hydrocephalus, abnormal spermatogenesis, mucus accumulation in the sinuses, and impaired spermatogenesis [3] |
| AK8 | Cytosol/liver, paancreas, lung, testis | Associated with the axoneme [95] | Not reported | KO mouse, hydrocephalus [96] |
| AK9 | Cytosol, nucleus/ highly expressed in pituitary gland, trachea, thymus, testis, mammary gland | Highly expressed in the pituitary gland, trachea, thymus, and testis [97] | Congenital myasthenic syndrome [98] | Not reported |
| Isozyme/Entrez Gene ID | Tissue/Cell type | Reference |
|---|---|---|
| AK1/203 | Ovarian cancer cells | [101] |
| AK1/203 | Platelets | [102] |
| AK1/203 | Squamous carcinoma cells | [103] |
| AK1/203 | Colorectal cancer cells | [104] |
| AK1/203 | Colorectal cancer cells | [105] |
| AK1/203 | Urine | [106] |
| AK1/203 | Urine | [107] |
| AK2/204 | B-cells | [108] |
| AK2/204 | Colorectal cancer cells | [104] |
| AK2/204 | Colorectal cancer cells | [105] |
| AK2/204 | Dendritic cells | [109] |
| AK2/204 | Endothelial cells | [110] |
| AK2/204 | Ovarian cancer cells | [101] |
| AK2/204 | Prostate cancer cells | [111] |
| AK2/204 | Squamous carcinoma cells | [103] |
| AK3/50808 | Hepatocytes | [112] |
| AK4/205 | Squamous carcinoma cells | [103] |
| AK4/205 | Urine | [113] |
| AK4 pseudogene 3/645619 | Glioblastoma cells | [114] |
| AK5/26289 | Glioblastoma cells | [114] |
| AK5/26289 | Mesenchymal stem cells | [115] |
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noma, T. Dynamics of nucleotide metabolism as a supporter of life phenomena. J. Med. Investig. 2005, 52, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, C.; Solaroli, N.; Karlsson, A. The many isoforms of human adenylate kinases. Int. J. Biochem. Cell Biol. 2014, 49, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gonzalez, A.; Kourembanas, S.; Wyatt, T.A.; Mitsialis, S.A. Mutation of murine adenylate kinase 7 underlies a primary ciliary dyskinesia phenotype. Am. J. Respir. Cell Mol. Biol. 2009, 40, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Panayiotou, C.; Solaroli, N.; Xu, Y.; Johansson, M.; Karlsson, A. The characterization of human adenylate kinases 7 and 8 demonstrates differences in kinetic parameters and structural organization among the family of adenylate kinase isoenzymes. Biochem. J. 2011, 433, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Solaroli, N.; Panayiotou, C.; Johansson, M.; Karlsson, A. Identification of two active functional domains of human adenylate kinase 5. FEBS Lett. 2009, 583, 2872–2876. [Google Scholar] [CrossRef]
- Miyoshi, K.; Akazawa, Y.; Horiguchi, T.; Noma, T. Localization of adenylate kinase 4 in mouse tissues. Acta Histochem. Cytochem. 2009, 42, 55–64. [Google Scholar] [CrossRef]
- Ren, H.; Wang, L.; Bennett, M.; Liang, Y.; Zheng, X.; Lu, F.; Li, L.; Nan, J.; Luo, M.; Eriksson, S.; et al. The crystal structure of human adenylate kinase 6: An adenylate kinase localized to the cell nucleus. Proc. Natl. Acad. Sci. USA 2005, 102, 303–308. [Google Scholar] [CrossRef]
- Dzeja, P.P.; Terzic, A. Phosphotransfer networks and cellular energetics. J. Exp. Biol. 2003, 206 Pt 12, 2039–2047. [Google Scholar] [CrossRef]
- Aversa, R.; Petrescu, R.; Apicella, A.; Petrescu, F. Mitochondria are Naturally Micro Robots–A review. Am. J. Eng. Appl. Sci. 2016, 9, 991–1002. [Google Scholar] [CrossRef]
- Kinsey, S.T.; Locke, B.R.; Dillaman, R.M. Molecules in motion: Influences of diffusion on metabolic structure and function in skeletal muscle. J. Exp. Biol. 2011, 214 Pt 2, 263–274. [Google Scholar] [CrossRef]
- Record, M.; Subra, C.; Silvente-Poirot, S.; Poirot, M. Exosomes as intercellular signalosomes and pharmacological effectors. Biochem. Pharmacol. 2011, 81, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Zomer, A.; Maynard, C.; Verweij, F.J.; Kamermans, A.; Schafer, R.; Beerling, E.; Schiffelers, R.M.; de Wit, E.; Berenguer, J.; Ellenbroek, S.I.J.; et al. In Vivo imaging reveals extracellular vesicle-mediated phenocopying of metastatic behavior. Cell 2015, 161, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Park, H.K.; Auh, Q.S.; Nah, H.; Lee, J.S.; Moon, H.J.; Heo, D.N.; Kim, I.S.; Kwon, I.K. Emerging Potential of Exosomes in Regenerative Medicine for Temporomandibular Joint Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 1541. [Google Scholar] [CrossRef]
- Arslan, F.; Lai, R.C.; Smeets, M.B.; Akeroyd, L.; Choo, A.; Aguor, E.N.; Timmers, L.; van Rijen, H.V.; Doevendans, P.A.; Pasterkamp, G.; et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013, 10, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Yegutkin, G.G.; Samburski, S.S.; Jalkanen, S. Soluble purine-converting enzymes circulate in human blood and regulate extracellular ATP level via counteracting pyrophosphatase and phosphotransfer reactions. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 1328–1330. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Samburski, S.S.; Mortensen, S.P.; Jalkanen, S.; Gonzalez-Alonso, J. Intravascular ADP and soluble nucleotidases contribute to acute prothrombotic state during vigorous exercise in humans. J. Physiol. 2007, 579 Pt 2, 553–564. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Wieringa, B.; Robson, S.C.; Jalkanen, S. Metabolism of circulating ADP in the bloodstream is mediated via integrated actions of soluble adenylate kinase-1 and NTPDase1/CD39 activities. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2012, 26, 3875–3883. [Google Scholar] [CrossRef]
- Kesimer, M.; Kirkham, S.; Pickles, R.J.; Henderson, A.G.; Alexis, N.E.; Demaria, G.; Knight, D.; Thornton, D.J.; Sheehan, J.K. Tracheobronchial air-liquid interface cell culture: A model for innate mucosal defense of the upper airways? Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L92–L100. [Google Scholar] [CrossRef]
- Szeinberg, A.; Kahana, D.; Gavendo, S.; Zaidman, J.; Ben-Ezzer, J. Hereditary deficiency of adenylate kinase in red blood cell. Acta Haematol. 1969, 42, 111–126. [Google Scholar] [CrossRef]
- Boivin, P.; Galand, C.; Hakim, J.; Simony, D.; Seligman, M. A new erythroenzymopathy: Congenital non-spherocytic hemolytic anemia and hereditary erythrocytic adenylate kinase deficiency. La Presse Med. 1971, 79, 215–218. [Google Scholar]
- Miwa, S.; Fujii, H.; Tani, K.; Takahashi, K.; Takizawa, T.; Igarashi, T. Red cell adenylate kinase deficiency associated with hereditary nonspherocytic hemolytic anemia: Clinical and biochemical studies. Am. J. Hematol. 1983, 14, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Carson, D.; Dannawi, H.; Forman, L.; Kuhl, W.; West, C.; Westwood, B. Metabolic compensation for profound erythrocyte adenylate kinase deficiency. A hereditary enzyme defect without hemolytic anemia. J. Clin. Investig. 1983, 72, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Lachant, N.A.; Zerez, C.R.; Barredo, J.; Lee, D.W.; Savely, S.M.; Tanaka, K.R. Hereditary erythrocyte adenylate kinase deficiency: A defect of multiple phosphotransferases? Blood 1991, 77, 2774–2784. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, S.; Igarashi, M.; Tanizawa, Y.; Yamada, M.; Kishi, F.; Kajii, T.; Fujii, H.; Miwa, S.; Sakurai, M.; Nakazawa, A. Human adenylate kinase deficiency associated with hemolytic anemia. A single base substitution affecting solubility and catalytic activity of the cytosolic adenylate kinase. J. Biol. Chem. 1989, 264, 10148–10155. [Google Scholar] [CrossRef]
- Mascarello, F.; De Luca, G.; Rizzotti, M. Histochemical demonstration of uridine diphospho-glucoso-4′-epimerase activity. Riv. Istochim. Norm. E Patol. 1976, 20, 231–236. [Google Scholar]
- Bianchi, P.; Zappa, M.; Bredi, E.; Vercellati, C.; Pelissero, G.; Barraco, F.; Zanella, A. A case of complete adenylate kinase deficiency due to a nonsense mutation in AK-1 gene (Arg 107 --> Stop, CGA --> TGA) associated with chronic haemolytic anaemia. Br. J. Haematol. 1999, 105, 75–79. [Google Scholar] [CrossRef]
- Toren, A.; Brok-Simoni, F.; Ben-Bassat, I.; Holtzman, F.; Mandel, M.; Neumann, Y.; Ramot, B.; Rechavi, G.; Kende, G. Congenital haemolytic anaemia associated with adenylate kinase deficiency. Br. J. Haematol. 1994, 87, 376–380. [Google Scholar] [CrossRef]
- He, S.; Chen, H.; Guo, X.; Gao, J. Red cell adenylate kinase deficiency in China: Molecular study of 2 new mutations (413G > A, 223dupA). BMC Med. Genom. 2022, 15, 102. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, G.; Zhang, H.; Chen, F.; Chen, Y.; Zhuang, Y.; Huang, Z.; Zou, F.; Liu, M.; An, G.; et al. Adenylate kinase 1 deficiency disrupts mouse sperm motility under conditions of energy stressdagger. Biol. Reprod. 2020, 103, 1121–1131. [Google Scholar] [CrossRef]
- Horiguchi, T.; Miyatake, Y.; Miyoshi, K.; Tanimura, A.; Hagita, H.; Sakaue, H.; Noma, T. Gene-expression profile reveals the genetic and acquired phenotypes of hyperactive mutant SPORTS rat. J. Med. Investig. 2020, 67, 51–61. [Google Scholar] [CrossRef]
- Maruyama, M.; Furukawa, Y.; Kinoshita, M.; Mukaiyama, A.; Akiyama, S.; Yoshimura, T. Adenylate kinase 1 overexpression increases locomotor activity in medaka fish. PLoS ONE 2022, 17, e0257967. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.; Dzeja, P.P.; Oerlemans, F.; Simonetti, A.W.; Heerschap, A.; de Haan, A.; Rush, P.S.; Terjung, R.R.; Wieringa, B.; Terzic, A. Adenylate kinase 1 gene deletion disrupts muscle energetic economy despite metabolic rearrangement. EMBO J. 2000, 19, 6371–6381. [Google Scholar] [CrossRef] [PubMed]
- Janssen, E.; Terzic, A.; Wieringa, B.; Dzeja, P.P. Impaired intracellular energetic communication in muscles from creatine kinase and adenylate kinase (M-CK/AK1) double knock-out mice. J. Biol. Chem. 2003, 278, 30441–30449. [Google Scholar] [CrossRef]
- Pucar, D.; Bast, P.; Gumina, R.J.; Lim, L.; Drahl, C.; Juranic, N.; Macura, S.; Janssen, E.; Wieringa, B.; Terzic, A.; et al. Adenylate kinase AK1 knockout heart: Energetics and functional performance under ischemia-reperfusion. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H776–H782. [Google Scholar] [CrossRef] [PubMed]
- Khoo, J.C.; Russell, P.J. Isoenzymes of adenylate kinase in human tissue. Biochim. Biophys. Acta 1972, 268, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Nobumoto, M.; Yamada, M.; Song, S.; Inouye, S.; Nakazawa, A. Mechanism of mitochondrial import of adenylate kinase isozymes. J. Biochem. 1998, 123, 128–135. [Google Scholar] [CrossRef]
- Brdiczka, D.; Beutner, G.; Ruck, A.; Dolder, M.; Wallimann, T. The molecular structure of mitochondrial contact sites. Their role in regulation of energy metabolism and permeability transition. BioFactors 1998, 8, 235–242. [Google Scholar] [CrossRef]
- Lee, H.J.; Pyo, J.O.; Oh, Y.; Kim, H.J.; Hong, S.H.; Jeon, Y.J.; Kim, H.; Cho, D.H.; Woo, H.N.; Song, S.; et al. AK2 activates a novel apoptotic pathway through formation of a complex with FADD and caspase-10. Nat. Cell Biol. 2007, 9, 1303–1310. [Google Scholar] [CrossRef]
- Lagresle-Peyrou, C.; Six, E.M.; Picard, C.; Rieux-Laucat, F.; Michel, V.; Ditadi, A.; Demerens-de Chappedelaine, C.; Morillon, E.; Valensi, F.; Simon-Stoos, K.L.; et al. Human adenylate kinase 2 deficiency causes a profound hematopoietic defect associated with sensorineural deafness. Nat. Genet. 2009, 41, 106–111. [Google Scholar] [CrossRef]
- Pannicke, U.; Honig, M.; Hess, I.; Friesen, C.; Holzmann, K.; Rump, E.M.; Barth, T.F.; Rojewski, M.T.; Schulz, A.; Boehm, T.; et al. Reticular dysgenesis (aleukocytosis) is caused by mutations in the gene encoding mitochondrial adenylate kinase 2. Nat. Genet. 2009, 41, 101–105. [Google Scholar] [CrossRef]
- Ichikawa, S.; Prockop, S.; Cunningham-Rundles, C.; Sifers, T.; Conner, B.R.; Wu, S.; Karam, R.; Walsh, M.F.; Fiala, E. Reticular dysgenesis caused by an intronic pathogenic variant in AK2. Mol. Case Stud. 2020, 6, a005017. [Google Scholar] [CrossRef] [PubMed]
- Burkart, A.; Shi, X.; Chouinard, M.; Corvera, S. Adenylate kinase 2 links mitochondrial energy metabolism to the induction of the unfolded protein response. J. Biol. Chem. 2011, 286, 4081–4089. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, A.; Miyoshi, K.; Horiguchi, T.; Hagita, H.; Fujisawa, K.; Noma, T. Mitochondrial Activity and Unfolded Protein Response are Required for Neutrophil Differentiation. Cell. Physiol. Biochem. 2018, 47, 1936–1950. [Google Scholar] [CrossRef] [PubMed]
- Glaser, M.; Nulty, W.; Vagelos, P.R. Role of adenylate kinase in the regulation of macromolecular biosynthesis in a putative mutant of Escherichia coli defective in membrane phospholipid biosynthesis. J. Bacteriol. 1975, 123, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yamada, S.; Park, S.; Klepinin, A.; Kaambre, T.; Terzic, A.; Dzeja, P. Adenylate kinase AK2 isoform integral in embryo and adult heart homeostasis. Biochem. Biophys. Res. Commun. 2021, 546, 59–64. [Google Scholar] [CrossRef]
- Rissone, A.; Weinacht, K.G.; la Marca, G.; Bishop, K.; Giocaliere, E.; Jagadeesh, J.; Felgentreff, K.; Dobbs, K.; Al-Herz, W.; Jones, M.; et al. Reticular dysgenesis-associated AK2 protects hematopoietic stem and progenitor cell development from oxidative stress. J. Exp. Med. 2015, 212, 1185–1202. [Google Scholar] [CrossRef]
- Rissone, A.; Jimenez, E.; Bishop, K.; Carrington, B.; Slevin, C.; Wincovitch, S.M.; Sood, R.; Candotti, F.; Burgess, S.M. A model for reticular dysgenesis shows impaired sensory organ development and hair cell regeneration linked to cellular stress. Dis. Model. Mech. 2019, 12, dmm040170. [Google Scholar] [CrossRef]
- Lamia, K.A.; Sachdeva, U.M.; DiTacchio, L.; Williams, E.C.; Alvarez, J.G.; Egan, D.F.; Vasquez, D.S.; Juguilon, H.; Panda, S.; Shaw, R.J.; et al. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science 2009, 326, 437–440. [Google Scholar] [CrossRef]
- Asher, G.; Schibler, U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell Metab. 2011, 13, 125–137. [Google Scholar] [CrossRef]
- Bass, J.; Takahashi, J.S. Circadian rhythms: Redox redux. Nature 2011, 469, 476–478. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, I.; Albrecht, U.; Ripperger, J.A. The role of clock genes and rhythmicity in the liver. Mol. Cell. Endocrinol. 2012, 349, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Fustin, J.M.; Doi, M.; Yamada, H.; Komatsu, R.; Shimba, S.; Okamura, H. Rhythmic nucleotide synthesis in the liver: Temporal segregation of metabolites. Cell Rep. 2012, 1, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, H.J.; Oh, Y.; Choi, S.G.; Hong, S.H.; Kim, H.J.; Lee, S.Y.; Choi, J.W.; Su Hwang, D.; Kim, K.S.; et al. The DUSP26 phosphatase activator adenylate kinase 2 regulates FADD phosphorylation and cell growth. Nat. Commun. 2014, 5, 3351. [Google Scholar] [CrossRef]
- Liu, H.; Pu, Y.; Amina, Q.; Wang, Q.; Zhang, M.; Song, J.; Guo, J.; Mardan, M. Prognostic and therapeutic potential of Adenylate kinase 2 in lung adenocarcinoma. Sci. Rep. 2019, 9, 17757. [Google Scholar] [CrossRef]
- Cai, F.; Xu, H.; Zha, D.; Wang, X.; Li, P.; Yu, S.; Yao, Y.; Chang, X.; Chen, J.; Lu, Y.; et al. AK2 Promotes the Migration and Invasion of Lung Adenocarcinoma by Activating TGF-beta/Smad Pathway In vitro and In vivo. Front. Pharmacol. 2021, 12, 714365. [Google Scholar] [CrossRef]
- Colletti, M.; Petretto, A.; Galardi, A.; Di Paolo, V.; Tomao, L.; Lavarello, C.; Inglese, E.; Bruschi, M.; Lopez, A.A.; Pascucci, L.; et al. Proteomic Analysis of Neuroblastoma-Derived Exosomes: New Insights into a Metastatic Signature. Proteomics 2017, 17, 1600430. [Google Scholar] [CrossRef]
- Klepinin, A.; Zhang, S.; Klepinina, L.; Rebane-Klemm, E.; Terzic, A.; Kaambre, T.; Dzeja, P. Adenylate Kinase and Metabolic Signaling in Cancer Cells. Front. Oncol. 2020, 10, 660. [Google Scholar] [CrossRef]
- Chang, X.; Ravi, R.; Pham, V.; Bedi, A.; Chatterjee, A.; Sidransky, D. Adenylate kinase 3 sensitizes cells to cigarette smoke condensate vapor induced cisplatin resistance. PLoS ONE 2011, 6, e20806. [Google Scholar] [CrossRef]
- Pullen, T.J.; Khan, A.M.; Barton, G.; Butcher, S.A.; Sun, G.; Rutter, G.A. Identification of genes selectively disallowed in the pancreatic islet. Islets 2010, 2, 89–95. [Google Scholar] [CrossRef]
- Fujisawa, K.; Wakazaki, M.; Matsuzaki, A.; Matsumoto, T.; Yamamoto, N.; Noma, T.; Takami, T. Adenylate Kinase Isozyme 3 Regulates Mitochondrial Energy Metabolism and Knockout Alters HeLa Cell Metabolism. Int. J. Mol. Sci. 2022, 23, 4316. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Wu, C.; Wang, M.; Wei, K.; Wang, J. Identification of novel cell glycolysis related gene signature predicting survival in patients with breast cancer. Sci. Rep. 2021, 11, 3986. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.Y.; Feng, S.C.; Sun, Y.Q.; Jiang, G.Q. MiR-96-5p promotes breast cancer migration by activating MEK/ERK signaling. J. Gene Med. 2020, 22, e3188. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kikkawa, R.; Yamada, H.; Horii, I. Investigation of proteomic biomarkers in in vivo hepatotoxicity study of rat liver: Toxicity differentiation in hepatotoxicants. J. Toxicol. Sci. 2006, 31, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Strom, A.L.; Zhai, J.; Gal, J.; Bao, S.; Gong, W.; Zhu, H. Enzymatically inactive adenylate kinase 4 interacts with mitochondrial ADP/ATP translocase. Int. J. Biochem. Cell Biol. 2009, 41, 1371–1380. [Google Scholar] [CrossRef]
- Vendrov, A.E.; Madamanchi, N.R.; Hakim, Z.S.; Rojas, M.; Runge, M.S. Thrombin and NAD(P)H oxidase-mediated regulation of CD44 and BMP4-Id pathway in VSMC, restenosis, and atherosclerosis. Circ. Res. 2006, 98, 1254–1263. [Google Scholar] [CrossRef]
- Lanning, N.J.; Looyenga, B.D.; Kauffman, A.L.; Niemi, N.M.; Sudderth, J.; DeBerardinis, R.J.; MacKeigan, J.P. A mitochondrial RNAi screen defines cellular bioenergetic determinants and identifies an adenylate kinase as a key regulator of ATP levels. Cell Rep. 2014, 7, 907–917. [Google Scholar] [CrossRef]
- Wujak, M.; Veith, C.; Wu, C.Y.; Wilke, T.; Kanbagli, Z.I.; Novoyatleva, T.; Guenther, A.; Seeger, W.; Grimminger, F.; Sommer, N.; et al. Adenylate Kinase 4-A Key Regulator of Proliferation and Metabolic Shift in Human Pulmonary Arterial Smooth Muscle Cells via Akt and HIF-1alpha Signaling Pathways. Int. J. Mol. Sci. 2021, 22, 10371. [Google Scholar] [CrossRef]
- Kong, F.; Binas, B.; Moon, J.H.; Kang, S.S.; Kim, H.J. Differential expression of adenylate kinase 4 in the context of disparate stress response strategies of HEK293 and HepG2 cells. Arch. Biochem. Biophys. 2013, 533, 11–17. [Google Scholar] [CrossRef]
- Fujisawa, K.; Terai, S.; Takami, T.; Yamamoto, N.; Yamasaki, T.; Matsumoto, T.; Yamaguchi, K.; Owada, Y.; Nishina, H.; Noma, T.; et al. Modulation of anti-cancer drug sensitivity through the regulation of mitochondrial activity by adenylate kinase 4. J. Exp. Clin. Cancer Res. 2016, 35, 48. [Google Scholar] [CrossRef]
- Liu, X.; Gonzalez, G.; Dai, X.; Miao, W.; Yuan, J.; Huang, M.; Bade, D.; Li, L.; Sun, Y.; Wang, Y. Adenylate Kinase 4 Modulates the Resistance of Breast Cancer Cells to Tamoxifen through an m(6)A-Based Epitranscriptomic Mechanism. Mol. Ther. 2020, 28, 2593–2604. [Google Scholar] [CrossRef] [PubMed]
- Xin, F.; Yao, D.W.; Fan, L.; Liu, J.H.; Liu, X.D. Adenylate kinase 4 promotes bladder cancer cell proliferation and invasion. Clin. Exp. Med. 2019, 19, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Jan, Y.H.; Tsai, H.Y.; Yang, C.J.; Huang, M.S.; Yang, Y.F.; Lai, T.C.; Lee, C.H.; Jeng, Y.M.; Huang, C.Y.; Su, J.L.; et al. Adenylate kinase-4 is a marker of poor clinical outcomes that promotes metastasis of lung cancer by downregulating the transcription factor ATF3. Cancer Res. 2012, 72, 5119–5129. [Google Scholar] [CrossRef] [PubMed]
- Jan, Y.H.; Lai, T.C.; Yang, C.J.; Lin, Y.F.; Huang, M.S.; Hsiao, M. Adenylate kinase 4 modulates oxidative stress and stabilizes HIF-1alpha to drive lung adenocarcinoma metastasis. J. Hematol. Oncol. 2019, 12, 12. [Google Scholar] [CrossRef]
- Li, L.; Deng, T.; Zhang, Q.; Yang, Y.; Liu, Y.; Yuan, L.; Xie, M. AK4P1 is a cancer-promoting pseudogene in pancreatic adenocarcinoma cells whose transcripts can be transmitted by exosomes. Oncol. Lett. 2022, 23, 163. [Google Scholar] [CrossRef]
- Van Rompay, A.R.; Johansson, M.; Karlsson, A. Identification of a novel human adenylate kinase. cDNA cloning, expression analysis, chromosome localization and characterization of the recombinant protein. Eur. J. Biochem. 1999, 261, 509–517. [Google Scholar] [CrossRef]
- Muniz-Castrillo, S.; Hedou, J.J.; Ambati, A.; Jones, D.; Vogrig, A.; Pinto, A.L.; Benaiteau, M.; de Broucker, T.; Fechtenbaum, L.; Labauge, P.; et al. Distinctive clinical presentation and pathogenic specificities of anti-AK5 encephalitis. Brain 2021, 144, 2709–2721. [Google Scholar] [CrossRef]
- Miyamoto, K.; Fukutomi, T.; Akashi-Tanaka, S.; Hasegawa, T.; Asahara, T.; Sugimura, T.; Ushijima, T. Identification of 20 genes aberrantly methylated in human breast cancers. Int. J. Cancer 2005, 116, 407–414. [Google Scholar] [CrossRef]
- Ahn, B.; Chae, Y.S.; Lee, S.K.; Kim, M.; Kim, H.S.; Moon, J.W.; Park, S.H. Identification of novel DNA hypermethylation of the adenylate kinase 5 promoter in colorectal adenocarcinoma. Sci. Rep. 2021, 11, 12626. [Google Scholar] [CrossRef]
- Zhang, L.H.; Wang, Z.; Fan, Q.Q.; Yin, Z.J.; Jin, L.F.; Mao, Y.; Hua, D. AK5, a novel prognosis marker, inhibits apoptosis and promotes autophagy as well as proliferation in human gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9900–9906. [Google Scholar]
- Santama, N.; Ogg, S.C.; Malekkou, A.; Zographos, S.E.; Weis, K.; Lamond, A.I. Characterization of hCINAP, a novel coilin-interacting protein encoded by a transcript from the transcription factor TAFIID32 locus. J. Biol. Chem. 2005, 280, 36429–36441. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Zhang, J.; Li, T.; Hang, R.; Liu, Y.; Tian, Y.; Huang, D.; Qu, L.; Cao, X.; Ji, J.; et al. The ATPase hCINAP regulates 18S rRNA processing and is essential for embryogenesis and tumour growth. Nat. Commun. 2016, 7, 12310. [Google Scholar] [CrossRef] [PubMed]
- Zhai, R.; Meng, G.; Zhao, Y.; Liu, B.; Zhang, G.; Zheng, X. A novel nuclear-localized protein with special adenylate kinase properties from Caenorhabditis elegans. FEBS Lett. 2006, 580, 3811–3817. [Google Scholar] [CrossRef] [PubMed]
- Granneman, S.; Nandineni, M.R.; Baserga, S.J. The putative NTPase Fap7 mediates cytoplasmic 20S pre-rRNA processing through a direct interaction with Rps14. Mol. Cell. Biol. 2005, 25, 10352–10364. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.; Zhai, R.; Liu, B.; Zheng, X. Identification of a novel nuclear-localized adenylate kinase from Drosophila melanogaster. Biochem. (Moscow) 2008, 73, 38–43. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, L.; Qin, N.; Cao, M.; Liang, X.; Wang, R. hCINAP is potentially a direct target gene of HIF-1 and is required for hypoxia-induced EMT and apoptosis in cervical cancer cells. Biochem. Cell Biol. 2021, 99, 203–213. [Google Scholar] [CrossRef]
- Ji, Y.; Yang, C.; Tang, Z.; Yang, Y.; Tian, Y.; Yao, H.; Zhu, X.; Zhang, Z.; Ji, J.; Zheng, X. Adenylate kinase hCINAP determines self-renewal of colorectal cancer stem cells by facilitating LDHA phosphorylation. Nat. Commun. 2017, 8, 15308. [Google Scholar] [CrossRef]
- Xu, R.; Yang, Y.; Zheng, X. Unique structural features of the adenylate kinase hCINAP/AK6 and its multifaceted functions in carcinogenesis and tumor progression. FEBS Lett. 2021, 595, 2071–2084. [Google Scholar] [CrossRef]
- Lonergan, K.M.; Chari, R.; Deleeuw, R.J.; Shadeo, A.; Chi, B.; Tsao, M.S.; Jones, S.; Marra, M.; Ling, V.; Ng, R.; et al. Identification of novel lung genes in bronchial epithelium by serial analysis of gene expression. Am. J. Respir. Cell Mol. Biol. 2006, 35, 651–661. [Google Scholar] [CrossRef]
- Milara, J.; Armengot, M.; Mata, M.; Morcillo, E.J.; Cortijo, J. Role of adenylate kinase type 7 expression on cilia motility: Possible link in primary ciliary dyskinesia. Am. J. Rhinol. Allergy 2010, 24, 181–185. [Google Scholar] [CrossRef]
- Mata, M.; Lluch-Estelles, J.; Armengot, M.; Sarrion, I.; Carda, C.; Cortijo, J. New adenylate kinase 7 (AK7) mutation in primary ciliary dyskinesia. Am. J. Rhinol. Allergy 2012, 26, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Lores, P.; Coutton, C.; El Khouri, E.; Stouvenel, L.; Givelet, M.; Thomas, L.; Rode, B.; Schmitt, A.; Louis, B.; Sakheli, Z.; et al. Homozygous missense mutation L673P in adenylate kinase 7 (AK7) leads to primary male infertility and multiple morphological anomalies of the flagella but not to primary ciliary dyskinesia. Hum. Mol. Genet. 2018, 27, 1196–1211. [Google Scholar] [CrossRef] [PubMed]
- Xiang, M.; Wang, Y.; Xu, W.; Zheng, N.; Deng, H.; Zhang, J.; Duan, Z.; Zha, X.; Zhang, W.; Song, G.; et al. A novel homozygous missense mutation in AK7 causes multiple morphological anomalies of the flagella and oligoasthenoteratozoospermia. J. Assist. Reprod. Genet. 2022, 39, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Lewis, H.; Sloan, S.H.; Foos, R.Y. Massive intraocular hemorrhage associated with anticoagulation and age-related macular degeneration. Graefe’s Arch. Clin. Exp. Ophthalmol. 1988, 226, 59–64. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Cao, W.; Aghajanian, H.K.; Haig-Ladewig, L.; Lin, A.M.; Al-Alao, O.; Gerton, G.L. Adenine nucleotide metabolism and a role for AMP in modulating flagellar waveforms in mouse sperm. Biol. Reprod. 2014, 90, 128. [Google Scholar] [CrossRef] [PubMed]
- Vogel, P.; Read, R.W.; Hansen, G.M.; Payne, B.J.; Small, D.; Sands, A.T.; Zambrowicz, B.P. Congenital hydrocephalus in genetically engineered mice. Vet. Pathol. 2012, 49, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Amiri, M.; Conserva, F.; Panayiotou, C.; Karlsson, A.; Solaroli, N. The human adenylate kinase 9 is a nucleoside mono- and diphosphate kinase. Int. J. Biochem. Cell Biol. 2013, 45, 925–931. [Google Scholar] [CrossRef]
- Lam, C.W.; Wong, K.S.; Leung, H.W.; Law, C.Y. Limb girdle myasthenia with digenic RAPSN and a novel disease gene AK9 mutations. Eur. J. Hum. Genet. 2017, 25, 192–199. [Google Scholar] [CrossRef]
- Janssen, E.; de Groof, A.; Wijers, M.; Fransen, J.; Dzeja, P.P.; Terzic, A.; Wieringa, B. Adenylate kinase 1 deficiency induces molecular and structural adaptations to support muscle energy metabolism. J. Biol. Chem. 2003, 278, 12937–12945. [Google Scholar] [CrossRef]
- Chen, R.P.; Liu, C.Y.; Shao, H.L.; Zheng, W.W.; Wang, J.X.; Zhao, X.F. Adenylate kinase 2 (AK2) promotes cell proliferation in insect development. BMC Mol. Biol. 2012, 13, 31. [Google Scholar] [CrossRef]
- Liang, B.; Peng, P.; Chen, S.; Li, L.; Zhang, M.; Cao, D.; Yang, J.; Li, H.; Gui, T.; Li, X.; et al. Characterization and proteomic analysis of ovarian cancer-derived exosomes. J. Proteom. 2013, 80, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Pienimaeki-Roemer, A.; Kuhlmann, K.; Bottcher, A.; Konovalova, T.; Black, A.; Orso, E.; Liebisch, G.; Ahrens, M.; Eisenacher, M.; Meyer, H.E.; et al. Lipidomic and proteomic characterization of platelet extracellular vesicle subfractions from senescent platelets. Transfusion 2015, 55, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Tan, H.S.; Datta, A.; Lai, R.C.; Zhang, H.; Meng, W.; Lim, S.K.; Sze, S.K. Hypoxic tumor cell modulates its microenvironment to enhance angiogenic and metastatic potential by secretion of proteins and exosomes. Mol. Cell. Proteom. 2010, 9, 1085–1099. [Google Scholar] [CrossRef] [PubMed]
- Tauro, B.J.; Greening, D.W.; Mathias, R.A.; Mathivanan, S.; Ji, H.; Simpson, R.J. Two distinct populations of exosomes are released from LIM1863 colon carcinoma cell-derived organoids. Mol. Cell. Proteom. 2013, 12, 587–598. [Google Scholar] [CrossRef]
- Choi, D.S.; Choi, D.Y.; Hong, B.S.; Jang, S.C.; Kim, D.K.; Lee, J.; Kim, Y.K.; Kim, K.P.; Gho, Y.S. Quantitative proteomics of extracellular vesicles derived from human primary and metastatic colorectal cancer cells. J. Extracell. Vesicles 2012, 1, 18704. [Google Scholar] [CrossRef]
- Gonzales, P.A.; Pisitkun, T.; Hoffert, J.D.; Tchapyjnikov, D.; Star, R.A.; Kleta, R.; Wang, N.S.; Knepper, M.A. Large-scale proteomics and phosphoproteomics of urinary exosomes. J. Am. Soc. Nephrol. 2009, 20, 363–379. [Google Scholar] [CrossRef]
- Raj, D.A.; Fiume, I.; Capasso, G.; Pocsfalvi, G. A multiplex quantitative proteomics strategy for protein biomarker studies in urinary exosomes. Kidney Int. 2012, 81, 1263–1272. [Google Scholar] [CrossRef]
- Buschow, S.I.; van Balkom, B.W.; Aalberts, M.; Heck, A.J.; Wauben, M.; Stoorvogel, W. MHC class II-associated proteins in B-cell exosomes and potential functional implications for exosome biogenesis. Immunol. Cell Biol. 2010, 88, 851–856. [Google Scholar] [CrossRef]
- Kowal, J.; Arras, G.; Colombo, M.; Jouve, M.; Morath, J.P.; Primdal-Bengtson, B.; Dingli, F.; Loew, D.; Tkach, M.; Thery, C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc. Natl. Acad. Sci. USA 2016, 113, E968–E977. [Google Scholar] [CrossRef]
- de Jong, O.G.; Verhaar, M.C.; Chen, Y.; Vader, P.; Gremmels, H.; Posthuma, G.; Schiffelers, R.M.; Gucek, M.; van Balkom, B.W. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J. Extracell. Vesicles 2012, 1, 18396. [Google Scholar] [CrossRef]
- Inder, K.L.; Zheng, Y.Z.; Davis, M.J.; Moon, H.; Loo, D.; Nguyen, H.; Clements, J.A.; Parton, R.G.; Foster, L.J.; Hill, M.M. Expression of PTRF in PC-3 Cells modulates cholesterol dynamics and the actin cytoskeleton impacting secretion pathways. Mol. Cell. Proteom. 2012, 11, M111.012245. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qin, H.; Poon, T.C.; Sze, S.C.; Ding, X.; Co, N.N.; Ngai, S.M.; Chan, T.F.; Wong, N. Hepatocellular carcinoma-derived exosomes promote motility of immortalized hepatocyte through transfer of oncogenic proteins and RNAs. Carcinogenesis 2015, 36, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Prunotto, M.; Farina, A.; Lane, L.; Pernin, A.; Schifferli, J.; Hochstrasser, D.F.; Lescuyer, P.; Moll, S. Proteomic analysis of podocyte exosome-enriched fraction from normal human urine. J. Proteom. 2013, 82, 193–229. [Google Scholar] [CrossRef] [PubMed]
- Skog, J.; Wurdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujisawa, K. Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases. Int. J. Mol. Sci. 2023, 24, 5561. https://doi.org/10.3390/ijms24065561
Fujisawa K. Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases. International Journal of Molecular Sciences. 2023; 24(6):5561. https://doi.org/10.3390/ijms24065561
Chicago/Turabian StyleFujisawa, Koichi. 2023. "Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases" International Journal of Molecular Sciences 24, no. 6: 5561. https://doi.org/10.3390/ijms24065561
APA StyleFujisawa, K. (2023). Regulation of Adenine Nucleotide Metabolism by Adenylate Kinase Isozymes: Physiological Roles and Diseases. International Journal of Molecular Sciences, 24(6), 5561. https://doi.org/10.3390/ijms24065561