
Mechanistic Insights into the Biological Effects of Engineered Nanomaterials: A Focus on Gold Nanoparticles


Abstract
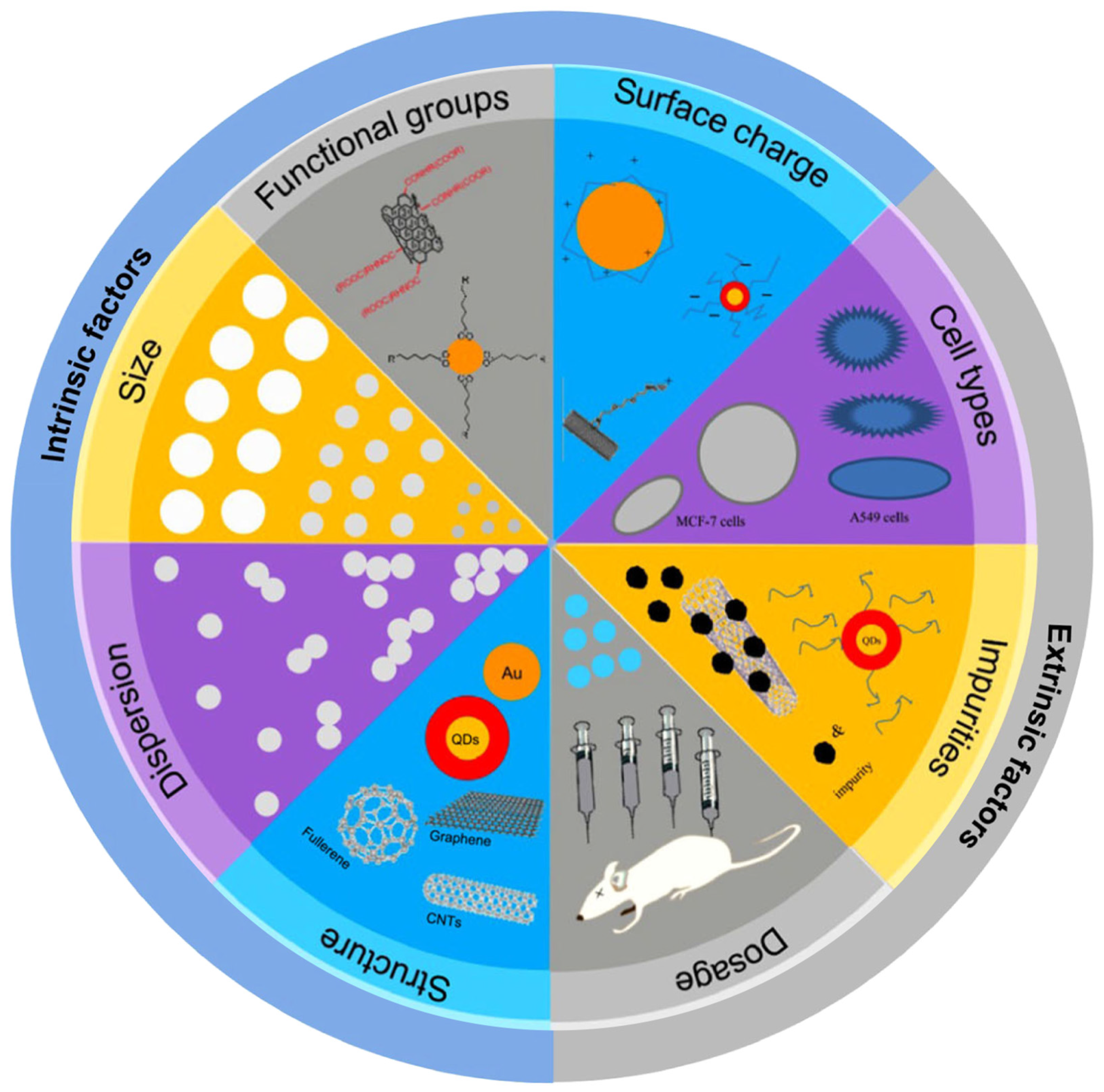
1. Introduction
Mechanistic Insights into the Biological Effects of ENMs—The Omics Era
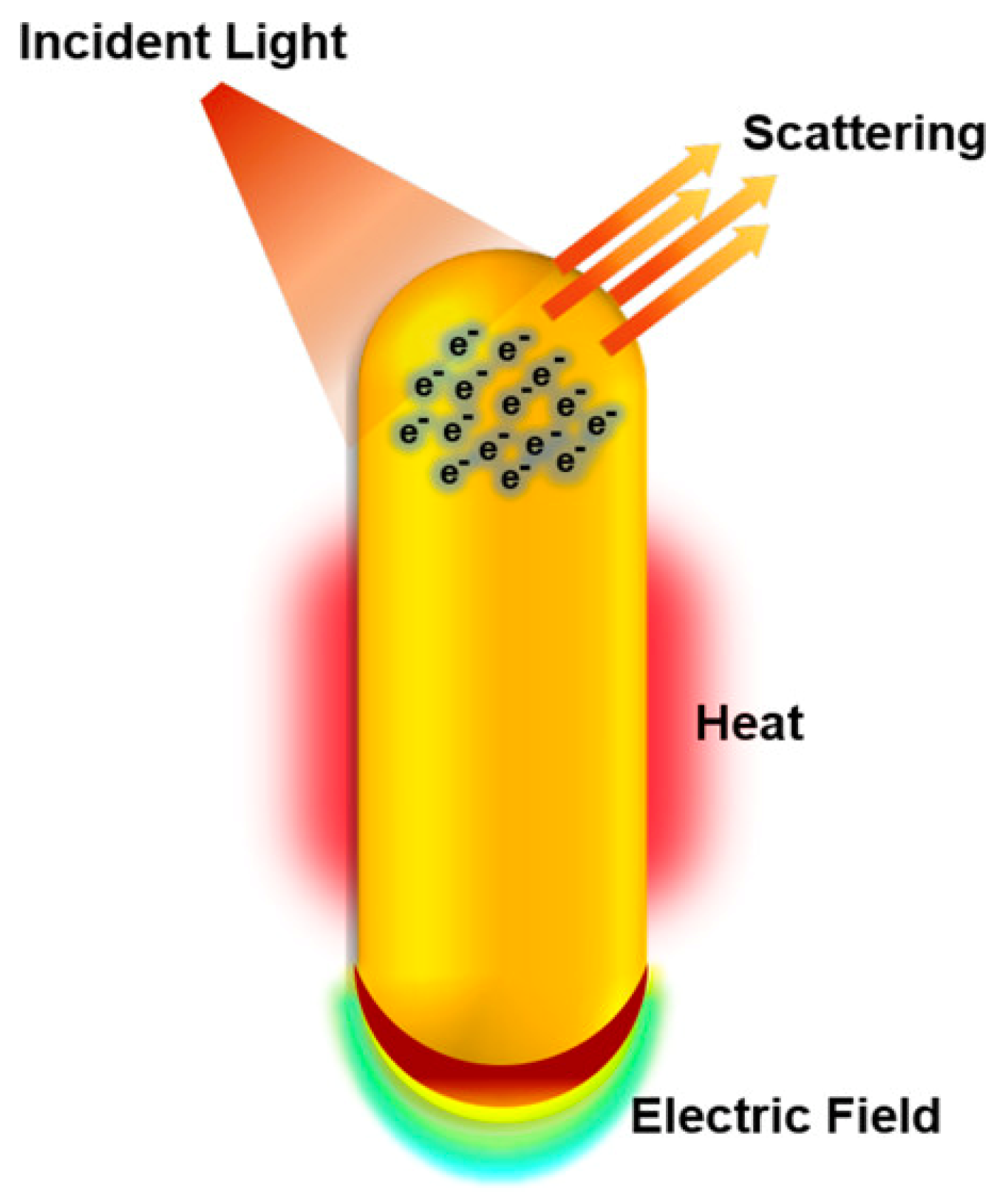
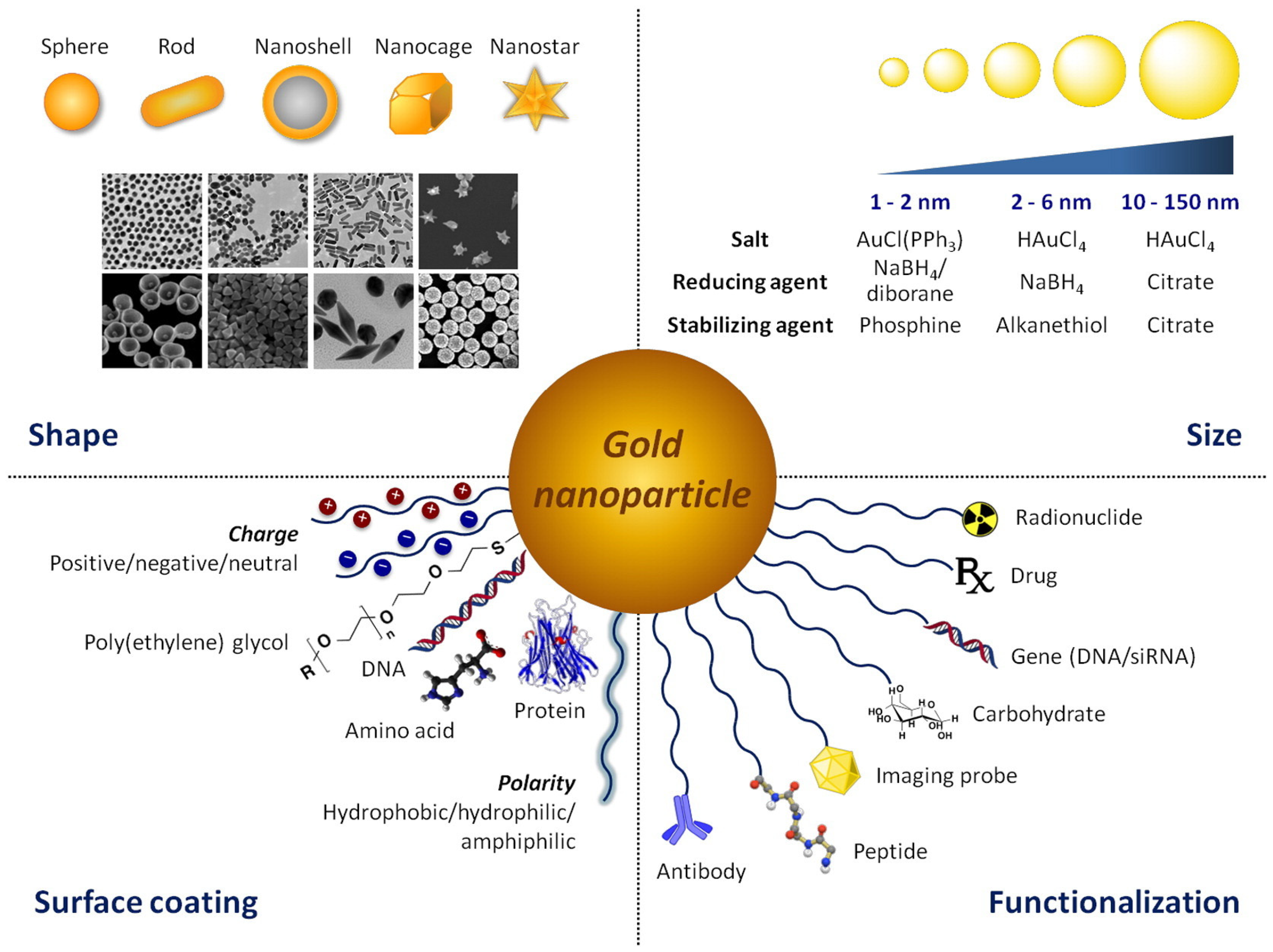
2. The Golden Age of (Nano) Medicine
Main Challenges in Gold-Based Nanomedicines Clinical Translation
3. Insights into the Molecular Effects of AuNPs in Biological Systems
3.1. Transcriptomics
3.2. Proteomics
3.3. Metabolomics
3.4. Epigenomics
3.4.1. DNA Methylation and Histone Modifications
3.4.2. Noncoding RNAs—miRNAs
3.5. Epitranscriptomics

{kind=link}
{kind=link}
{kind=link}
| Shape | Size | Surface Chemistry | Cell Type | Exposure | Method | Ref | |
|---|---|---|---|---|---|---|---|
| Epigenetics | |||||||
| DNA methylation | |||||||
| Spheres | 4 nm | thiol | hESCs (human embryonic stem cells) | 10 µg/mL; 24 h | Immunoprecipitation-based colorimetric assay | [104] | |
| Spheres | 4–5 nm | L-Glutathione (L-GSH) | HEK293FT (human embryonic kidney 293) and MGC-803 (human gastric carcinoma) | 100 µg/mL; 48 h | Dot blot assay | [105] | |
| Spheres | 5, 60, and 250 nm | citrate | Lung tissue (BALB/c mouse) | 5 and 50 µg; 48 h | LC-MS 1; bisulfite pyrosequencing | [106] | |
| Histone modifications | |||||||
| Spheres | 20 nm | citrate | MRC-5 (human normal lung fibroblasts) | 1 nM; 72 h | Immunofluorescence | [110] | |
| Spheres, Stars | ~20 nm, ~45 nm | Bis (p-sulfonatophenyl)phenylphosphine dehydrate dipotassium salt (BSPP) | dTHP-1 (differentiated human leukemia monocytic cell) and A549 (human lung adenocarcinoma) | 1 × 1011 partilces/mL; 24 h | LC-MS 1 followed by chromatin immunoprecipitation sequencing (ChIP-seq) | [111] | |
| miRNAs | |||||||
| Spheres | 20 nm | citrate | Blood cells (rats) | 1 wk; 2 mo | Microarray | [133] | |
| Spheres | 40 nm; 100 nm | citrate | Swiss mice fetal liver and lungs | 3.3 mg/kg (4 doses); 18 d of Swiss female mice gestation | Microarray | [123] | |
| Spheres | 20 nm | citrate | HDF (human normal dermal fibroblast) | 200 µM;.1, 4 and 8 h | RNA-Seq | [124] | |
| Spheres, Rods | 20 nm; 16 nm width, 46 nm length | citrate, poly (acrylic acid) (PAA); poly (ethylene glycol) (PEG) | HDF (human normal dermal fibroblast) | 0.1 nM; 24 h and 20 wk | RNA-Seq | [63] | |
| Epitranscriptomics | |||||||
| Rods | 21 nm width, 130 nm length | chitosan and 12-mer peptide | AML (Acute myeloid leukemia cells | 0.25 nM; 6 h | m6A-Seq followed by gene-specific m6A-qPCR and LC-MS/MS 2 | [131] | |
| Spheres | 3 nm | bovine serum albumin (BSA) | HEK293T (human embryonic kidney 293) | 200 μg/mL; 24 h | m6A-Seq followed by gene-specific m6A-qPCR | [132] | |
3.6. Multi-Omics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- ISO/TS 80004-1:2015; Nanotechnologies—Vocabulary—Part 1: Core Terms. International Organization for Standardization (ISO): Geneva, Switzerland, 2015; pp. 1–4.
- Talebian, S.; Rodrigues, T.; Das Neves, J.; Sarmento, B.; Langer, R.; Conde, J. Facts and Figures on Materials Science and Nanotechnology Progress and Investment. ACS Nano 2021, 15, 15940–15952. [Google Scholar] [CrossRef]
- Chris, B. Introduction to Nanoscience and Nanotechnology; MEMS Nanoelectron; John Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 1–416. [Google Scholar]
- Ross, G. Risks and Benefits. Nat. Nanotechnol. 2006, 1, 151. [Google Scholar] [CrossRef]
- Baucher, M.-A.; Scott, R.; Cannizzaro, C.; Standridge, S.; Nesbitt, E.; Fadel, T. Symposium on Assessing the Economic Impact of Nanotechnology: Synthesis Report. Organ. Econ. Co-Oper. Dev. 2013, 4, 1–80. Available online: https://www.oecd.org/sti/emerging-tech/ (accessed on 5 November 2022).
- Kisby, T.; Yilmazer, A.; Kostarelos, K. Reasons for Success and Lessons Learnt from Nanoscale Vaccines against COVID-19. Nat. Nanotechnol. 2021, 16, 843–850. [Google Scholar] [CrossRef]
- Falagan-Lotsch, P.; Grzincic, E.M.; Murphy, C.J. New Advances in Nanotechnology-Based Diagnosis and Therapeutics for Breast Cancer: An Assessment of Active-Targeting Inorganic Nanoplatforms. Bioconjug. Chem. 2017, 28, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Barenholz, Y. Doxil®—The First FDA-Approved Nano-Drug: Lessons Learned. J. Control. Release 2012, 160, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-Based Drug Delivery in Cancer Therapy and Its Role in Overcoming Drug Resistance. Front. Mol. Biosci. 2020, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- The Nanodatabase. Consum. Prod. 2013. Available online: https://nanodb.dk/en/ (accessed on 26 October 2022).
- Global Nanomaterials Market Forecast 2020–2028. INKWOOD Res. 2022. Available online: https://inkwoodresearch.com/reports/nanomaterials-market/ (accessed on 15 October 2022).
- Zhao, H.; Wang, Y.; Bao, L.; Chen, C. Engineering Nano-Bio Interfaces from Nanomaterials to Nanomedicines. Acc. Mater. Res. 2022, 3, 812–829. [Google Scholar] [CrossRef]
- Đorđević, S.; Gonzalez, M.M.; Conejos-Sánchez, I.; Carreira, B.; Pozzi, S.; Acúrcio, R.C.; Satchi-Fainaro, R.; Florindo, H.F.; Vicent, M.J. Current Hurdles to the Translation of Nanomedicines from Bench to the Clinic. Drug Deliv. Transl. Res. 2022, 12, 500–525. [Google Scholar] [CrossRef]
- Wilhelm, S.; Tavares, A.J.; Dai, Q.; Ohta, S.; Audet, J.; Dvorak, H.F.; Chan, W.C.W. Analysis of Nanoparticle Delivery to Tumours. Nat. Rev. Mater. 2016, 1, 16014. [Google Scholar] [CrossRef]
- Bondarenko, O.; Mortimer, M.; Kahru, A.; Feliu, N.; Javed, I.; Kakinen, A.; Lin, S.; Xia, T.; Song, Y.; Davis, T.P.; et al. Nanotoxicology and Nanomedicine: The Yin and Yang of Nano-Bio Interactions for the New Decade. Nano Today 2021, 39, 101–184. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; Leite, P.E.; Falagan-Lotsch, P.; Benetti, F.; Micheletti, C.; Budtz, H.C.; Jacobsen, N.R.; Lisboa-Filho, P.N.; Rocha, L.A.; Kühnel, D.; et al. Challenges on the Toxicological Predictions of Engineered Nanoparticles. NanoImpact 2017, 8, 59–72. [Google Scholar] [CrossRef]
- Yang, W.; Wang, L.; Mettenbrink, E.M.; DeAngelis, P.L.; Wilhelm, S. Nanoparticle Toxicology. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Halappanavar, S.; Vogel, U.; Wallin, H.; Yauk, C.L. Promise and Peril in Nanomedicine: The Challenges and Needs for Integrated Systems Biology Approaches to Define Health Risk. WIREs Nanomed. Nanobiotechnology 2018, 10, 1–7. [Google Scholar] [CrossRef]
- Nymark, P.; Kohonen, P.; Hongisto, V.; Grafström, R.C. Toxic and Genomic Influences of Inhaled Nanomaterials as a Basis for Predicting Adverse Outcome. Ann. Am. Thorac. Soc. 2018, 15, S91–S97. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Meng, H.; Yan, L.; Wang, B.; Zhao, Y. Nanosurface Chemistry and Dose Govern the Bioaccumulation and Toxicity of Carbon Nanotubes, Metal Nanomaterials and Quantum Dots in Vivo. Sci. Bull. 2015, 60, 3–20. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Meng, H.; Wang, X.; Lin, S.; Ji, Z.; Zhang, H. Nanomaterial Toxicity Testing in the 21st Century: Use of a Predictive Toxicological Approach and High-Throughput Screening. Acc. Chem. Res. 2013, 46, 607–621. [Google Scholar] [CrossRef]
- Costa, P.M.; Fadeel, B. Emerging Systems Biology Approaches in Nanotoxicology: Towards a Mechanism-Based Understanding of Nanomaterial Hazard and Risk. Toxicol. Appl. Pharmacol. 2016, 299, 101–111. [Google Scholar] [CrossRef]
- National Research Council. Toxicity Testing in the 21st Century: A Vision and a Strategy; National Academies Press: Washington, DC, USA, 2007; ISBN 0309109922. [Google Scholar]
- Hartung, T. Toxicology for the Twenty-First Century. Nature 2009, 460, 208–212. [Google Scholar] [CrossRef]
- Feliu, N.; Kohonen, P.; Ji, J.; Zhang, Y.; Karlsson, H.L.; Palmberg, L.; Nyström, A.; Fadeel, B. Next-Generation Sequencing Reveals Low-Dose Effects of Cationic Dendrimers in Primary Human Bronchial Epithelial Cells. ACS Nano 2015, 9, 146–163. [Google Scholar] [CrossRef]
- Kohonen, P.; Parkkinen, J.A.; Willighagen, E.L.; Ceder, R.; Wennerberg, K.; Kaski, S.; Grafström, R.C. A Transcriptomics Data-Driven Gene Space Accurately Predicts Liver Cytopathology and Drug-Induced Liver Injury. Nat. Commun. 2017, 8, 15932. [Google Scholar] [CrossRef] [PubMed]
- Halappanavar, S.; Rahman, L.; Nikota, J.; Poulsen, S.S.; Ding, Y.; Jackson, P.; Wallin, H.; Schmid, O.; Vogel, U.; Williams, A. Ranking of Nanomaterial Potency to Induce Pathway Perturbations Associated with Lung Responses. NanoImpact 2019, 14, 100158. [Google Scholar] [CrossRef]
- Fadeel, B.; Farcal, L.; Hardy, B.; Vázquez-Campos, S.; Hristozov, D.; Marcomini, A.; Lynch, I.; Valsami-Jones, E.; Alenius, H.; Savolainen, K. Advanced Tools for the Safety Assessment of Nanomaterials. Nat. Nanotechnol. 2018, 13, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Sturla, S.J.; Boobis, A.R.; FitzGerald, R.E.; Hoeng, J.; Kavlock, R.J.; Schirmer, K.; Whelan, M.; Wilks, M.F.; Peitsch, M.C. Systems Toxicology: From Basic Research to Risk Assessment. Chem. Res. Toxicol. 2014, 27, 314–329. [Google Scholar] [CrossRef]
- Hartung, T.; FitzGerald, R.E.; Jennings, P.; Mirams, G.R.; Peitsch, M.C.; Rostami-Hodjegan, A.; Shah, I.; Wilks, M.F.; Sturla, S.J. Systems Toxicology: Real World Applications and Opportunities. Chem. Res. Toxicol. 2017, 30, 870–882. [Google Scholar] [CrossRef]
- Sau, T.K.; Murphy, C.J. Seeded High Yield Synthesis of Short Au Nanorods in Aqueous Solution. Langmuir 2004, 20, 6414–6420. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Murphy, C.J. Mini Gold Nanorods with Tunable Plasmonic Peaks beyond 1000 Nm. Chem. Mater. 2018, 30, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.J.; Chang, H.; Falagan-Lotsch, P.; Gole, M.T.; Hofmann, D.M.; Hoang, K.N.L.; McClain, S.M.; Meyer, S.M.; Turner, J.G.; Unnikrishnan, M.; et al. Virus-Sized Gold Nanorods: Plasmonic Particles for Biology. Acc. Chem. Res. 2019, 52, 2124–2135. [Google Scholar] [CrossRef]
- Burrows, N.D.; Lin, W.; Hinman, J.G.; Dennison, J.M.; Vartanian, A.M.; Abadeer, N.S.; Grzincic, E.M.; Jacob, L.M.; Li, J.; Murphy, C.J. Surface Chemistry of Gold Nanorods. Langmuir 2016, 32, 9905–9921. [Google Scholar] [CrossRef]
- Wu, M.; Vartanian, A.M.; Chong, G.; Pandiakumar, A.K.; Hamers, R.J.; Hernandez, R.; Murphy, C.J. Solution NMR Analysis of Ligand Environment in Quaternary Ammonium-Terminated Self-Assembled Monolayers on Gold Nanoparticles: The Effect of Surface Curvature and Ligand Structure. J. Am. Chem. Soc. 2019, 141, 4316–4327. [Google Scholar] [CrossRef]
- Her, S.; Jaffray, D.A.; Allen, C. Gold Nanoparticles for Applications in Cancer Radiotherapy: Mechanisms and Recent Advancements. Adv. Drug Deliv. Rev. 2017, 109, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Dykman, L.; Khlebtsov, N. Gold Nanoparticles in Biomedical Applications: Recent Advances and Perspectives. Chem. Soc. Rev. 2012, 41, 2256–2282. [Google Scholar] [CrossRef] [PubMed]
- Crowe, H.W. Treatment of Rheumatoid Arthritis with Gold Salts. Lancet 1934, 224, 845–846. [Google Scholar] [CrossRef]
- Rastinehad, A.R.; Anastos, H.; Wajswol, E.; Winoker, J.S.; Sfakianos, J.P.; Doppalapudi, S.K.; Carrick, M.R.; Knauer, C.J.; Taouli, B.; Lewis, S.C.; et al. Gold Nanoshell-Localized Photothermal Ablation of Prostate Tumors in a Clinical Pilot Device Study. Proc. Natl. Acad. Sci. USA 2019, 116, 18590–18596. [Google Scholar] [CrossRef] [PubMed]
- Kumthekar, P.; Ko, C.H.; Paunesku, T.; Dixit, K.; Sonabend, A.M.; Bloch, O.; Tate, M.; Schwartz, M.; Zuckerman, L.; Lezon, R.; et al. A First-in-Human Phase 0 Clinical Study of RNA Interference–Based Spherical Nucleic Acids in Patients with Recurrent Glioblastoma. Sci. Transl. Med. 2021, 13, eabb3945. [Google Scholar] [CrossRef]
- Dreaden, E.C.; Alkilany, A.M.; Huang, X.; Murphy, C.J.; El-Sayed, M.A. The Golden Age: Gold Nanoparticles for Biomedicine. Chem. Soc. Rev. 2012, 41, 2740–2779. [Google Scholar] [CrossRef]
- van de Looij, S.M.; Hebels, E.R.; Viola, M.; Hembury, M.; Oliveira, S.; Vermonden, T. Gold Nanoclusters: Imaging, Therapy, and Theranostic Roles in Biomedical Applications. Bioconjug. Chem. 2022, 33, 4–23. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef]
- Libutti, S.K.; Paciotti, G.F.; Byrnes, A.A.; Alexander, H.R.; Gannon, W.E.; Walker, M.; Seidel, G.D.; Yuldasheva, N.; Tamarkin, L. Phase I and Pharmacokinetic Studies of CYT-6091, a Novel PEGylated Colloidal Gold-RhTNF Nanomedicine. Clin. Cancer Res. 2010, 16, 6139–6149. [Google Scholar] [CrossRef]
- Vucic, S.; Kiernan, M.C.; Menon, P.; Huynh, W.; Rynders, A.; Ho, K.S.; Glanzman, R.; Hotchkin, M.T. Study Protocol of RESCUE-ALS: A Phase 2, Randomised, Double-Blind, Placebo-Controlled Study in Early Symptomatic Amyotrophic Lateral Sclerosis Patients to Assess Bioenergetic Catalysis with CNM-Au8 as a Mechanism to Slow Disease Progression. BMJ Open 2021, 11, e041479. [Google Scholar] [CrossRef]
- Tatovic, D.; McAteer, M.A.; Barry, J.; Barrientos, A.; Rodríguez Terradillos, K.; Perera, I.; Kochba, E.; Levin, Y.; Dul, M.; Coulman, S.A.; et al. Safety of the Use of Gold Nanoparticles Conjugated with Proinsulin Peptide and Administered by Hollow Microneedles as an Immunotherapy in Type 1 Diabetes. Immunother. Adv. 2022, 2, ltac002. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Liu, L.; Morin, E.E.; Liu, M.; Schwendeman, A. Survey of Clinical Translation of Cancer Nanomedicines—Lessons Learned from Successes and Failures. Acc. Chem. Res. 2019, 52, 2673–2683. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Soliman, M.G.; Sun, X.; Pelaz, B.; Feliu, N.; Parak, W.J.; Liu, S. How Entanglement of Different Physicochemical Properties Complicates the Prediction of in Vitro and in Vivo Interactions of Gold Nanoparticles. ACS Nano 2018, 12, 10104–10113. [Google Scholar] [CrossRef]
- Chetwynd, A.J.; Lynch, I. The Rise of the Nanomaterial Metabolite Corona, and Emergence of the Complete Corona. Environ. Sci. Nano 2020, 7, 1041–1060. [Google Scholar] [CrossRef]
- Lin, S.; Mortimer, M.; Chen, R.; Kakinen, A.; Riviere, J.E.; Davis, T.P.; Ding, F.; Ke, P.C. NanoEHS beyond Toxicity—Focusing on Biocorona. Environ. Sci. Nano 2017, 4, 1433–1454. [Google Scholar] [CrossRef]
- Cantarutti, C.; Hunashal, Y.; La Rosa, C.; Condorelli, M.; Giorgetti, S.; Bellotti, V.; Fogolari, F.; Esposito, G. The Corona of Protein-Gold Nanoparticle Systems: The Role of Ionic Strength. Phys. Chem. Chem. Phys. 2022, 24, 1630–1637. [Google Scholar] [CrossRef]
- Ali, M.R.K.; Rahman, M.A.; Wu, Y.; Han, T.; Peng, X.; Mackey, M.A.; Wang, D.; Shin, H.J.; Chen, Z.G.; Xiao, H.; et al. Efficacy, Long-Term Toxicity, and Mechanistic Studies of Gold Nanorods Photothermal Therapy of Cancer in Xenograft Mice. Proc. Natl. Acad. Sci. USA 2017, 114, E3110–E3118. [Google Scholar] [CrossRef]
- Balfourier, A.; Luciani, N.; Wang, G.; Lelong, G.; Ersen, O.; Khelfa, A.; Alloyeau, D.; Gazeau, F.; Carn, F. Unexpected Intracellular Biodegradation and Recrystallization of Gold Nanoparticles. Proc. Natl. Acad. Sci. USA 2020, 117, 103–113. [Google Scholar] [CrossRef]
- Yang, J.A.; Lohse, S.E.; Murphy, C.J. Tuning Cellular Response to Nanoparticles via Surface Chemistry and Aggregation. Small 2014, 10, 1642–1651. [Google Scholar] [CrossRef]
- Bao, L.; Cui, X.; Chen, C. Toxicology for Nanotechnology. In Micro/Nano Technologies; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–22. ISBN 9789811393747. [Google Scholar]
- Truong, L.; Zaikova, T.; Baldock, B.L.; Balik-Meisner, M.; To, K.; Reif, D.M.; Kennedy, Z.C.; Hutchison, J.E.; Tanguay, R.L. Systematic Determination of the Relationship between Nanoparticle Core Diameter and Toxicity for a Series of Structurally Analogous Gold Nanoparticles in Zebrafish. Nanotoxicology 2019, 13, 879–893. [Google Scholar] [CrossRef]
- Isoda, K.; Tanaka, A.; Fuzimori, C.; Echigoya, M.; Taira, Y.; Taira, I.; Shimizu, Y.; Akimoto, Y.; Kawakami, H.; Ishida, I. Toxicity of Gold Nanoparticles in Mice Due to Nanoparticle/Drug Interaction Induces Acute Kidney Damage. Nanoscale Res. Lett. 2020, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Villiers, C.L.; Freitas, H.; Couderc, R.; Villiers, M.B.; Marche, P.N. Analysis of the Toxicity of Gold Nano Particles on the Immune System: Effect on Dendritic Cell Functions. J. Nanoparticle Res. 2010, 12, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Vetten, M.A.; Tlotleng, N.; Tanner Rascher, D.; Skepu, A.; Keter, F.K.; Boodhia, K.; Koekemoer, L.A.; Andraos, C.; Tshikhudo, R.; Gulumian, M. Label-Free in Vitro Toxicity and Uptake Assessment of Citrate Stabilised Gold Nanoparticles in Three Cell Lines. Part. Fibre Toxicol. 2013, 10, 1–15. [Google Scholar] [CrossRef]
- Grzincic, E.M.; Murphy, C.J. Gold Nanorods Indirectly Promote Migration of Metastatic Human Breast Cancer Cells in Three-Dimensional Cultures. ACS Nano 2015, 9, 6801–6816. [Google Scholar] [CrossRef] [PubMed]
- Carnovale, C.; Bryant, G.; Shukla, R.; Bansal, V. Identifying Trends in Gold Nanoparticle Toxicity and Uptake: Size, Shape, Capping Ligand, and Biological Corona. ACS Omega 2019, 4, 242–256. [Google Scholar] [CrossRef]
- Falagan-Lotsch, P.; Grzincic, E.M.; Murphy, C.J. One Low-Dose Exposure of Gold Nanoparticles Induces Long-Term Changes in Human Cells. Proc. Natl. Acad. Sci. USA 2016, 113, 13318–13323. [Google Scholar] [CrossRef] [PubMed]
- Falagan-Lotsch, P.; Murphy, C.J. Network-Based Analysis Implies Critical Roles of MicroRNAs in the Long-Term Cellular Responses to Gold Nanoparticles. Nanoscale 2020, 12, 21172–21187. [Google Scholar] [CrossRef]
- Kinaret, P.A.S.; Serra, A.; Federico, A.; Kohonen, P.; Nymark, P.; Liampa, I.; Ha, M.K.; Choi, J.-S.; Jagiello, K.; Sanabria, N.; et al. Transcriptomics in Toxicogenomics, Part I: Experimental Design, Technologies, Publicly Available Data, and Regulatory Aspects. Nanomaterials 2020, 10, 750. [Google Scholar] [CrossRef]
- Luijten, M.; Wackers, P.F.K.; Rorije, E.; Pennings, J.L.A.; Heusinkveld, H.J. Relevance of in Vitro Transcriptomics for in Vivo Mode of Action Assessment. Chem. Res. Toxicol. 2021, 34, 452–459. [Google Scholar] [CrossRef]
- Grzincic, E.M.; Yang, J.A.; Drnevich, J.; Falagan-Lotsch, P.; Murphy, C.J. Global Transcriptomic Analysis of Model Human Cell Lines Exposed to Surface-Modified Gold Nanoparticles: The Effect of Surface Chemistry. Nanoscale 2015, 7, 1349–1362. [Google Scholar] [CrossRef]
- Bajak, E.; Fabbri, M.; Ponti, J.; Gioria, S.; Ojea-Jiménez, I.; Collotta, A.; Mariani, V.; Gilliland, D.; Rossi, F.; Gribaldo, L. Changes in Caco-2 Cells Transcriptome Profiles upon Exposure to Gold Nanoparticles. Toxicol. Lett. 2015, 233, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Chueh, P.J.; Liang, R.Y.; Lee, Y.H.; Zeng, Z.M.; Chuang, S.M. Differential Cytotoxic Effects of Gold Nanoparticles in Different Mammalian Cell Lines. J. Hazard. Mater. 2014, 264, 303–312. [Google Scholar] [CrossRef]
- Serra, A.; Letunic, I.; Fortino, V.; Handy, R.D.; Fadeel, B.; Tagliaferri, R.; Greco, D. INSIdE NANO: A Systems Biology Framework to Contextualize the Mechanism-of-Action of Engineered Nanomaterials. Sci. Rep. 2019, 9, 179. [Google Scholar] [CrossRef]
- Schlumpf, U.; Meyer, M.; Ulrich, J.; Friede, R.L. Neurologic Complications Induced by Gold Treatment. Arthritis Rheum. 1983, 26, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Gambari, P.; Ostuni, P.; Lazzarin, P.; Tavolato, B.; Todesco, S. Neurotoxicity Following a Very High Dose of Oral Gold (Auranofin). Arthritis Rheum. 1984, 27, 1316–1317. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.Y.; Tu, H.F.; Tan, X.; Yeh, Y.S.; Chueh, P.J.; Chuang, S.M. A Gene Signature for Gold Nanoparticle-Exposed Human Cell Lines. Toxicol. Res. (Camb) 2015, 4, 365–375. [Google Scholar] [CrossRef]
- Silins, I.; Högberg, J. Combined Toxic Exposures and Human Health: Biomarkers of Exposure and Effect. Int. J. Environ. Res. Public Health 2011, 8, 629–647. [Google Scholar] [CrossRef]
- Samim, A.R.; Arshad, M.; Vaseem, H. An Insight into Various Biomarkers to Study Toxicological Impact of Nanoparticles in Fishes: Explored and Missing Information. Int. J. Environ. Sci. Technol. 2022, 1–20. [Google Scholar] [CrossRef]
- Hossen, M.N.; Elechalawar, C.K.; Sjoelund, V.; Moore, K.; Mannel, R.; Bhattacharya, R.; Mukherjee, P. Experimental Conditions Influence the Formation and Composition of the Corona around Gold Nanoparticles. Cancer Nanotechnol. 2021, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Zhang, J.; Li, M.; Yang, D.; Liu, D.; Song, S.; Fu, J.; Zhang, H.; Dai, W.; Wang, X.; et al. Proteomic Analysis of Intracellular Protein Corona of Nanoparticles Elucidates Nano-Trafficking Network and Nano-Bio Interactions. Theranostics 2020, 10, 1213–1229. [Google Scholar] [CrossRef]
- del Pilar Chantada-Vázquez, M.; López, A.C.; Vence, M.G.; Vázquez-Estévez, S.; Acea-Nebril, B.; Calatayud, D.G.; Jardiel, T.; Bravo, S.B.; Núñez, C. Proteomic Investigation on Bio-Corona of Au, Ag and Fe Nanoparticles for the Discovery of Triple Negative Breast Cancer Serum Protein Biomarkers. J. Proteomics 2020, 212, 103581. [Google Scholar] [CrossRef]
- Zhang, T.; Gaffrey, M.J.; Thrall, B.D.; Qian, W.-J. Mass Spectrometry-Based Proteomics for System-Level Characterization of Biological Responses to Engineered Nanomaterials. Anal. Bioanal. Chem. 2018, 410, 6067–6077. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Gaffrey, M.J.; Thomas, D.G.; Weber, T.J.; Hess, B.M.; Weitz, K.K.; Piehowski, P.D.; Petyuk, V.A.; Moore, R.J.; Qian, W.-J.; et al. A Proteome-Wide Assessment of the Oxidative Stress Paradigm for Metal and Metal-Oxide Nanomaterials in Human Macrophages. NanoImpact 2020, 17, 100194. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Huang, Y.; Lü, X. Proteomic Analysis of Molecular Biocompatibility of Gold Nanoparticles to Human Dermal Fibroblasts-Fetal. J. Biomed. Nanotechnol. 2013, 9, 40–52. [Google Scholar] [CrossRef]
- Tsai, Y.Y.; Huang, Y.H.; Chao, Y.L.; Hu, K.Y.; Chin, L.-T.; Chou, S.H.; Hour, A.L.; Yao, Y.-D.; Tu, C.S.; Liang, Y.J.; et al. Identification of the Nanogold Particle-Induced Endoplasmic Reticulum Stress by Omic Techniques and Systems Biology Analysis. ACS Nano 2011, 5, 9354–9369. [Google Scholar] [CrossRef]
- Ng, C.-T.; Yung, L.-Y.L.; Swa, H.L.-F.; Poh, R.W.-Y.; Gunaratne, J.; Bay, B.-H. Altered Protein Expression Profile Associated with Phenotypic Changes in Lung Fibroblasts Co-Cultured with Gold Nanoparticle-Treated Small Airway Epithelial Cells. Biomaterials 2015, 39, 31–38. [Google Scholar] [CrossRef]
- Tarasova, N.K.; Gallud, A.; Ytterberg, A.J.; Chernobrovkin, A.; Aranzaes, J.R.; Astruc, D.; Antipov, A.; Fedutik, Y.; Fadeel, B.; Zubarev, R.A. Cytotoxic and Proinflammatory Effects of Metal-Based Nanoparticles on THP-1 Monocytes Characterized by Combined Proteomics Approaches. J. Proteome Res. 2017, 16, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Tzur, D.; Knox, C.; Eisner, R.; Guo, A.C.; Young, N.; Cheng, D.; Jewell, K.; Arndt, D.; Sawhney, S.; et al. HMDB: The Human Metabolome Database. Nucleic Acids Res. 2007, 35, 521–526. [Google Scholar] [CrossRef]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond Biomarkers and towards Mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Shin, T.H.; Lee, D.Y.; Lee, H.S.; Park, H.J.; Jin, M.S.; Paik, M.J.; Manavalan, B.; Mo, J.S.; Lee, G. Integration of Metabolomics and Transcriptomics in Nanotoxicity Studies. BMB Rep. 2018, 51, 14–20. [Google Scholar] [CrossRef]
- Viant, M.R.; Ebbels, T.M.D.; Beger, R.D.; Ekman, D.R.; Epps, D.J.T.; Kamp, H.; Leonards, P.E.G.; Loizou, G.D.; MacRae, J.I.; van Ravenzwaay, B.; et al. Use Cases, Best Practice and Reporting Standards for Metabolomics in Regulatory Toxicology. Nat. Commun. 2019, 10, 3041. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Toward Merging Untargeted and Targeted Methods in Mass Spectrometry-Based Metabolomics and Lipidomics. Anal. Chem. 2016, 88, 524–545. [Google Scholar] [CrossRef]
- Lindeque, J.Z.; Matthyser, A.; Mason, S.; Louw, R.; Taute, C.J.F. Metabolomics Reveals the Depletion of Intracellular Metabolites in HepG2 Cells after Treatment with Gold Nanoparticles. Nanotoxicology 2018, 12, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lü, X.; Chen, R.; Chen, Y. Comparative Study of the Effects of Gold and Silver Nanoparticles on the Metabolism of Human Dermal Fibroblasts. Regen. Biomater. 2020, 7, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Enea, M.; Araújo, A.M.; Peixoto de Almeida, M.; Soares, M.E.; Gonçalves-Monteiro, S.; Guedes de Pinho, P.; Pereira, E.; Bastos, M.d.L.; Carmo, H. A Metabolomic Approach for the in Vivo Study of Gold Nanospheres and Nanostars after a Single-Dose Intravenous Administration to Wistar Rats. Nanomaterials 2019, 9, 1606. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chen, M.; Ji, X.; Mao, Z.; Zhang, X.; Wang, X.; Xia, Y. Metabolomic Profiles Delineate the Potential Role of Glycine in Gold Nanorod-Induced Disruption of Mitochondria and Blood-Testis Barrier Factors in TM-4 Cells. Nanoscale 2014, 6, 8265–8273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, J.; Zhuo, Y.; Zou, Z.; Li, Y.; Yang, H.; Xie, W.; Zeng, J.; Deng, Y.; Cai, S.; et al. Untargeted Metabolomics Reveals Alterations in the Metabolic Reprogramming of Prostate Cancer Cells by Double-Stranded DNA-Modified Gold Nanoparticles. Biomater. Adv. 2022, 135, 212745. [Google Scholar] [CrossRef] [PubMed]
- Cote, I.L.; McCullough, S.D.; Hines, R.N.; Vandenberg, J.J. Application of Epigenetic Data in Human Health Risk Assessment. Curr. Opin. Toxicol. 2017, 6, 71–78. [Google Scholar] [CrossRef]
- Cote, I.; Vandenberg, J.J.; Druwe, I.L.; Angrish, M.M. Incorporating Epigenetics into a Risk Assessment Framework; Elsevier Inc.: Amsterdam, The Netherlands, 2018; ISBN 9780128124338. [Google Scholar]
- Chorley, B.N. Epigenetics in Risk Assessment: Clarity or Confusion? In Proceedings of the Environmental and Molecular Mutagenesis & Genomics Society 2021 Virtual Annual Meeting, Virtual Meeting, 22–25 September 2021; pp. 1–20. [Google Scholar] [CrossRef]
- Lu, X.; Miousse, I.R.; Pirela, S.V.; Moore, J.K.; Melnyk, S.; Koturbash, I.; Demokritou, P. In Vivo Epigenetic Effects Induced by Engineered Nanomaterials: A Case Study of Copper Oxide and Laser Printer-Emitted Engineered Nanoparticles. Nanotoxicology 2016, 10, 629–639. [Google Scholar] [CrossRef]
- González-Palomo, A.K.; Saldaña-Villanueva, K.; Cortés-García, J.D.; Fernández-Macias, J.C.; Méndez-Rodríguez, K.B.; Pérez Maldonado, I.N. Effect of Silver Nanoparticles (AgNPs) Exposure on MicroRNA Expression and Global DNA Methylation in Endothelial Cells EA.Hy926. Environ. Toxicol. Pharmacol. 2021, 81, 103543. [Google Scholar] [CrossRef]
- Pogribna, M.; Hammons, G. Epigenetic Effects of Nanomaterials and Nanoparticles. J. Nanobiotechnol. 2021, 19, 1–18. [Google Scholar] [CrossRef]
- Cedar, H.; Bergman, Y. Linking DNA Methylation and Histone Modification: Patterns and Paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef]
- Ehrlich, M. DNA Hypomethylation in Cancer Cells. Epigenomics 2009, 1, 239–259. [Google Scholar] [CrossRef]
- Jin, Z.; Liu, Y. DNA Methylation in Human Diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, A.; Nakanishi, M. Navigating the DNA Methylation Landscape of Cancer. Trends Genet. 2021, 37, 1012–1027. [Google Scholar] [CrossRef]
- Senut, M.-C.; Zhang, Y.; Liu, F.; Sen, A.; Ruden, D.M.; Mao, G. Size-Dependent Toxicity of Gold Nanoparticles on Human Embryonic Stem Cells and Their Neural Derivatives. Small 2016, 12, 631–646. [Google Scholar] [CrossRef]
- Ma, Y.; Fu, H.; Zhang, C.; Cheng, S.; Gao, J.; Wang, Z.; Jin, W.; Conde, J.; Cui, D. Chiral Antioxidant-Based Gold Nanoclusters Reprogram DNA Epigenetic Patterns. Sci. Rep. 2016, 6, 33436. [Google Scholar] [CrossRef]
- Tabish, A.M.; Poels, K.; Byun, H.M.; Luyts, K.; Baccarelli, A.A.; Martens, J.; Kerkhofs, S.; Seys, S.; Hoet, P.; Godderis, L. Changes in DNA Methylation in Mouse Lungs after a Single Intra-Tracheal Administration of Nanomaterials. PLoS ONE 2017, 12, e0169886. [Google Scholar] [CrossRef] [PubMed]
- Grant, P.A. A Tale of Histone Modifications. Genome Biol. 2001, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Torres, I.O.; Fujimori, D.G. Functional Coupling between Writers, Erasers and Readers of Histone and DNA Methylation. Curr. Opin. Struct. Biol. 2015, 35, 68–75. [Google Scholar] [CrossRef]
- Shyamasundar, S.; Ng, C.T.; Lanry Yung, L.Y.; Dheen, S.T.; Bay, B.H. Epigenetic Mechanisms in Nanomaterial-Induced Toxicity. Epigenomics 2015, 7, 395–411. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Silveira, C.P.; Cai, Q.; Dawson, K.A.; Cagney, G.; Yan, Y. Nanoscale Shape-Dependent Histone Modifications. PNAS Nexus 2022, 1, pgac172. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.K.S.; Malakar, A.K.; Chakraborty, S. Interplay between MiRNAs and Human Diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef] [PubMed]
- Tribolet, L.; Kerr, E.; Cowled, C.; Bean, A.G.D.; Stewart, C.R.; Dearnley, M.; Farr, R.J. MicroRNA Biomarkers for Infectious Diseases: From Basic Research to Biosensing. Front. Microbiol. 2020, 11, 1197. [Google Scholar] [CrossRef]
- Mendell, J.T.; Olson, E.N. MicroRNAs in Stress Signaling and Human Disease. Cell 2012, 148, 1172–1187. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Song, Y. Preferential Regulation of MiRNA Targets by Environmental Chemicals in the Human Genome. BMC Genom. 2011, 12, 1–13. [Google Scholar] [CrossRef]
- Ballesteros, S.; Vales, G.; Velázquez, A.; Pastor, S.; Alaraby, M.; Marcos, R.; Hernández, A. Micrornas as a Suitable Biomarker to Detect the Effects of Long-Term Exposures to Nanomaterials. Studies on TiO2 NP and MWCNT. Nanomaterials 2021, 11, 3458. [Google Scholar] [CrossRef]
- Ventura, C.; Vieira, L.; Silva, C.; Sousa-Uva, A.; Silva, M.J. Functional Effects of Differentially Expressed MicroRNAs in A549 Cells Exposed to MWCNT-7 or Crocidolite. Toxicol. Lett. 2020, 328, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Buckley, A.; Marczylo, T.; Seiffert, J.; Römer, I.; Warren, J.; Hodgson, A.; Chung, K.F.; Gant, T.W.; Smith, R.; et al. The Small Airway Epithelium as a Target for the Adverse Pulmonary Effects of Silver Nanoparticle Inhalation. Nanotoxicology 2018, 12, 539–553. [Google Scholar] [CrossRef]
- Huang, Y.; Lü, X.; Lü, X. Study of Key Biological Pathways and Important MicroRNAs Involved in Silver Nanoparticles Induced Cytotoxicity Based on MicroRNA Sequencing Technology. J. Biomed. Nanotechnol. 2018, 14, 2042–2055. [Google Scholar] [CrossRef]
- Sun, B.; Liu, R.; Ye, N.; Xiao, Z.D. Comprehensive Evaluation of Microrna Expression Profiling Reveals the Neural Signaling Specific Cytotoxicity of Superparamagnetic Iron Oxide Nanoparticles (SPIONs) through N-Methyl-D-Aspartate Receptor. PLoS ONE 2015, 10, e0121671. [Google Scholar] [CrossRef]
- Balansky, R.; Longobardi, M.; Ganchev, G.; Iltcheva, M.; Nedyalkov, N.; Atanasov, P.; Toshkova, R.; De Flora, S.; Izzotti, A. Transplacental Clastogenic and Epigenetic Effects of Gold Nanoparticles in Mice. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2013, 751–752, 42–48. [Google Scholar] [CrossRef]
- Huang, Y.; Lü, X.; Qu, Y.; Yang, Y.; Wu, S. MicroRNA Sequencing and Molecular Mechanisms Analysis of the Effects of Gold Nanoparticles on Human Dermal Fibroblasts. Biomaterials 2015, 37, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling. Curr. Biol. 2014, 24, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Rechavi, G. Epitranscriptome Regulation. Nat. Struct. Mol. Biol. 2018. [Google Scholar] [CrossRef]
- Kumar, S.; Mohapatra, T. Deciphering Epitranscriptome: Modification of MRNA Bases Provides a New Perspective for Post-Transcriptional Regulation of Gene Expression. Front. Cell Dev. Biol. 2021, 9, 628415. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the Human and Mouse M6A RNA Methylomes Revealed by M6A-Seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, B.; Nie, Z.; Duan, L.; Xiong, Q.; Jin, Z.; Yang, C.; Chen, Y. The Role of M6A Modification in the Biological Functions and Diseases. Signal Transduct. Target. Ther. 2021, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Subbarayalu, P.; Medina, D.; Nirzhor, S.; Timilsina, S.; Rajamanickam, S.; Eedunuri, V.K.; Gupta, Y.; Zheng, S.; Abdelfattah, N.; et al. M6A RNA Methylation Regulates Histone Ubiquitination to Support Cancer Growth and Progression. Cancer Res. 2022, 82, 1872–1889. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Han, M.; Cao, K.; Li, Q.; Pang, J.; Dou, L.; Liu, S.; Shi, Z.; Yan, F.; Feng, S. Gold Nanorods Exhibit Intrinsic Therapeutic Activity via Controlling N 6-Methyladenosine-Based Epitranscriptomics in Acute Myeloid Leukemia. ACS Nano 2021, 15, 17689–17704. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Wang, J.; Fang, K.; Hou, W.; Li, B.; Zhao, J.; Ma, X. RNA M6A Alterations Induced by Biomineralization Nanoparticles: A Proof-of-Concept Study of Epitranscriptomics for Nanotoxicity Evaluation. Nanoscale Res. Lett. 2022, 17, 23. [Google Scholar] [CrossRef]
- Chew, W.S.; Poh, K.W.; Siddiqi, N.J.; Alhomida, A.S.; Yu, L.E.; Ong, W.Y. Short- and Long-Term Changes in Blood MiRNA Levels after Nanogold Injection in Rats-Potential Biomarkers of Nanoparticle Exposure. Biomarkers 2012, 17, 750–757. [Google Scholar] [CrossRef]
- Scala, G.; Kinaret, P.; Marwah, V.; Sund, J.; Fortino, V.; Greco, D. Multi-Omics Analysis of Ten Carbon Nanomaterials Effects Highlights Cell Type Specific Patterns of Molecular Regulation and Adaptation. NanoImpact 2018, 11, 99–108. [Google Scholar] [CrossRef]
- Aragoneses-Cazorla, G.; Buendia-Nacarino, M.P.; Mena, M.L.; Luque-Garcia, J.L. A Multi-Omics Approach to Evaluate the Toxicity Mechanisms Associated with Silver Nanoparticles Exposure. Nanomaterials 2022, 12, 1762. [Google Scholar] [CrossRef]
- Gardner, L.; Kostarelos, K.; Mallick, P.; Dive, C.; Hadjidemetriou, M. Nano-Omics: Nanotechnology-Based Multidimensional Harvesting of the Blood-Circulating Cancerome. Nat. Rev. Clin. Oncol. 2022, 19, 551–561. [Google Scholar] [CrossRef]
- Gioria, S.; Vicente, J.L.; Barboro, P.; La Spina, R.; Tomasi, G.; Urbán, P.; Kinsner-Ovaskainen, A.; François, R.; Chassaigne, H. A Combined Proteomics and Metabolomics Approach to Assess the Effects of Gold Nanoparticles in Vitro. Nanotoxicology 2016, 10, 736–748. [Google Scholar] [CrossRef]
- Gallud, A.; Klöditz, K.; Ytterberg, J.; Östberg, N.; Katayama, S.; Skoog, T.; Gogvadze, V.; Chen, Y.Z.; Xue, D.; Moya, S.; et al. Cationic Gold Nanoparticles Elicit Mitochondrial Dysfunction: A Multi-Omics Study. Sci. Rep. 2019, 9, 4366. [Google Scholar] [CrossRef]
- Salamanca-Buentello, F.; Daar, A.S. Nanotechnology, Equity and Global Health. Nat. Nanotechnol. 2021, 16, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Fadeel, B. Nanomaterial Characterization: Understanding Nano-Bio Interactions. Biochem. Biophys. Res. Commun. 2022, 633, 45–51. [Google Scholar] [CrossRef] [PubMed]



| Name | Composition | Investigated Application | Clinicaltrials.Gov ID (Phase) |
|---|---|---|---|
| Aurimune | 7 nm PEGylated colloidal gold particles loaded with TNF-α | treatment of advanced and metastatic solid tumors | NCT00356980; NCT00436410 (both Phase I) |
| AuroLase | ~150 nm diameter gold-silica nanoshells coated with PEG | photothermal therapy against lung, head and neck and prostate tumors | NCT01679470 (not applicable) NCT00848042 (not applicable) NCT02680535 (not applicable) |
| NU-0129 | gold nanoparticles loaded with siRNA specific for the oncogene Bcl2L12, and OEG | treatment of glioblastoma | NCT03020017 (early Phase I) |
| CNM-Au8 | 13 nm gold nanocrystals | Parkinson’s disease, multiple sclerosis, amyotrophic lateral sclerosis | NCT03815916 (Phase II) NCT03993171 (Phase II) NCT04098406 (Phase II) |
| C19A3-GNP | ultrasmall-AuNPs (less than 5 nm) loaded with a proinsulin-derived peptide | treatment of type 1 diabetes | NCT02837094 (Phase I) |
| Gold Factor | 8–28 nm AuNPs with varying shapes | treatment of arthritis | NCT05347602 (not applicable) |
| Shape | Size | Surface Chemistry | Cell Type | Exposure | Method | Ref |
|---|---|---|---|---|---|---|
| Spheres | 20 nm | citrate, poly(allylamine hydrochloride) (PAH), and lipid coatings combined with alkanethiols or PAH | HDF (human dermal fibroblasts); PC3 (human prostate cancer) | 0.1 nM (HDF); 1 nM (PC3); 24–48 h | Microarray | [66] |
| Spheres | 5 nm | citrate | Caco2 (human epithelial colorectal adenocarcinoma) | 300 μM; 72 h | Microarray | [67] |
| Rods | 10–40 nm length | no data | MRC-5 (human normal lung fibroblasts) | 360 ng/mL; 24 h | Microarray | [68] |
| Spheres | 4 nm | citrate | HDF (primary human dermal fibroblasts) | no data; 1 d, 2 wk, 2 mo | Microarray | [53] |
| Spheres | 39, 41, and 45 nm | no data | A549 (human lung adenocarcinoma); HEK293 (human embryonic kidney 293), HepG2 (human hepatocellular carcinoma), and AGS (human gastric adenocarcinoma) | 360 ng/mL; 24 h | Microarray | [72] |
| Shape | Size | Surface Chemistry | Cell Type | Exposure | Method | Ref |
|---|---|---|---|---|---|---|
| Spheres | 20 nm | citrate | HDF-f (human dermal fibroblasts-fetal) | 200 μM; 1, 4 and 8 h | 2D-DIGE/MS 1 | [80] |
| Spheres | 5.9 nm | no surface coating (“naked”) | K562 (human chronic myelogenous leukemia) | 2.95 nM; 48 h | 2D-DIGE/MS 1; protein microarray | [81] |
| Spheres | 20 nm | citrate | MRC-5 (human normal lung fibroblast) in co-culture with SAECs (small airway epithelial cells) | 1 nM, 72 h (SAECs) | LC-MS/MS 2 | [82] |
| Spheres | 10–20 nm and 32–54 nm (bimodal) | amine groups (NH2) | THP-1 (human leukemia monocytic cell) | 15 μg/mL; 3 h | LC-MS/MS 2 | [83] |
| Shape | Size | Surface Chemistry | Cell Type | Exposure | Method | Ref |
|---|---|---|---|---|---|---|
| Spheres | 18 nm | citrate, poly-(sodium styrene sulfonate (PSSNa), or poly-vinylpyrrolidone (PVP) | HepG2 (human hepatocellular carcinoma) | 0.25 nM (PSSNa and PVP) and 0.5 nM (citrate); 3 h | GC-MS 1 and LC-MS/MS 2 | [89] |
| Spheres | 20 nm | citrate | HDF (human normal dermal fibroblasts) | 200 μM; 4, 8 and 24 h | LC-QTOF-MS 3 | [90] |
| Rods | 10 nm width, 40 nm length | Cetyltrimethylammonium Bromide (CTAB) | TM-4 (mouse Sertoli cells) | 10 nm; 24 h | GC-MS 1 | [92] |
| Spheres | 50 nm | double-stranded DNA | PC3 and DU145 (human prostate cancer cell lines) | 20 μg/mL; 24 h | LC-QTOF-MS 3 | [93] |
| Shape | Size | Surface Chemistry | Cell Type | Exposure | Method | Ref |
|---|---|---|---|---|---|---|
| Spheres | 5 nm; 30 nm | citrate | Caco 2 (human epithelial colorectal adenocarcinoma) | 300 μM; 72 h | 2D-DIGE/MS 1; LC-HRMS/MS 2 | [137] |
| Spheres | 5 nm; 20 nm | alkyl ammonium bromide, alkyl sodium carboxylate, or poly(ethylene glycol) (PEG) | THP-1 (human leukemia monocytic cell) | trancriptomics: 27 μg/mL (5 nm); 4 μg/mL (20 nm); 6 h. proteomics: 35 μg/mL (5 nm); 15 μg/mL (20 nm); 24 h | RNA-Seq; HPLC-MS 3 | [138] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.H.A.; Falagan-Lotsch, P. Mechanistic Insights into the Biological Effects of Engineered Nanomaterials: A Focus on Gold Nanoparticles. Int. J. Mol. Sci. 2023, 24, 4109. https://doi.org/10.3390/ijms24044109
Nguyen NHA, Falagan-Lotsch P. Mechanistic Insights into the Biological Effects of Engineered Nanomaterials: A Focus on Gold Nanoparticles. International Journal of Molecular Sciences. 2023; 24(4):4109. https://doi.org/10.3390/ijms24044109
Chicago/Turabian StyleNguyen, Nhung H. A., and Priscila Falagan-Lotsch. 2023. "Mechanistic Insights into the Biological Effects of Engineered Nanomaterials: A Focus on Gold Nanoparticles" International Journal of Molecular Sciences 24, no. 4: 4109. https://doi.org/10.3390/ijms24044109
APA StyleNguyen, N. H. A., & Falagan-Lotsch, P. (2023). Mechanistic Insights into the Biological Effects of Engineered Nanomaterials: A Focus on Gold Nanoparticles. International Journal of Molecular Sciences, 24(4), 4109. https://doi.org/10.3390/ijms24044109