
Cognitive and Emotional Symptoms Induced by Chronic Stress Are Regulated by EGR1 in a Subpopulation of Hippocampal Pyramidal Neurons

 , , and
, , and
Abstract
1. Introduction
2. Results
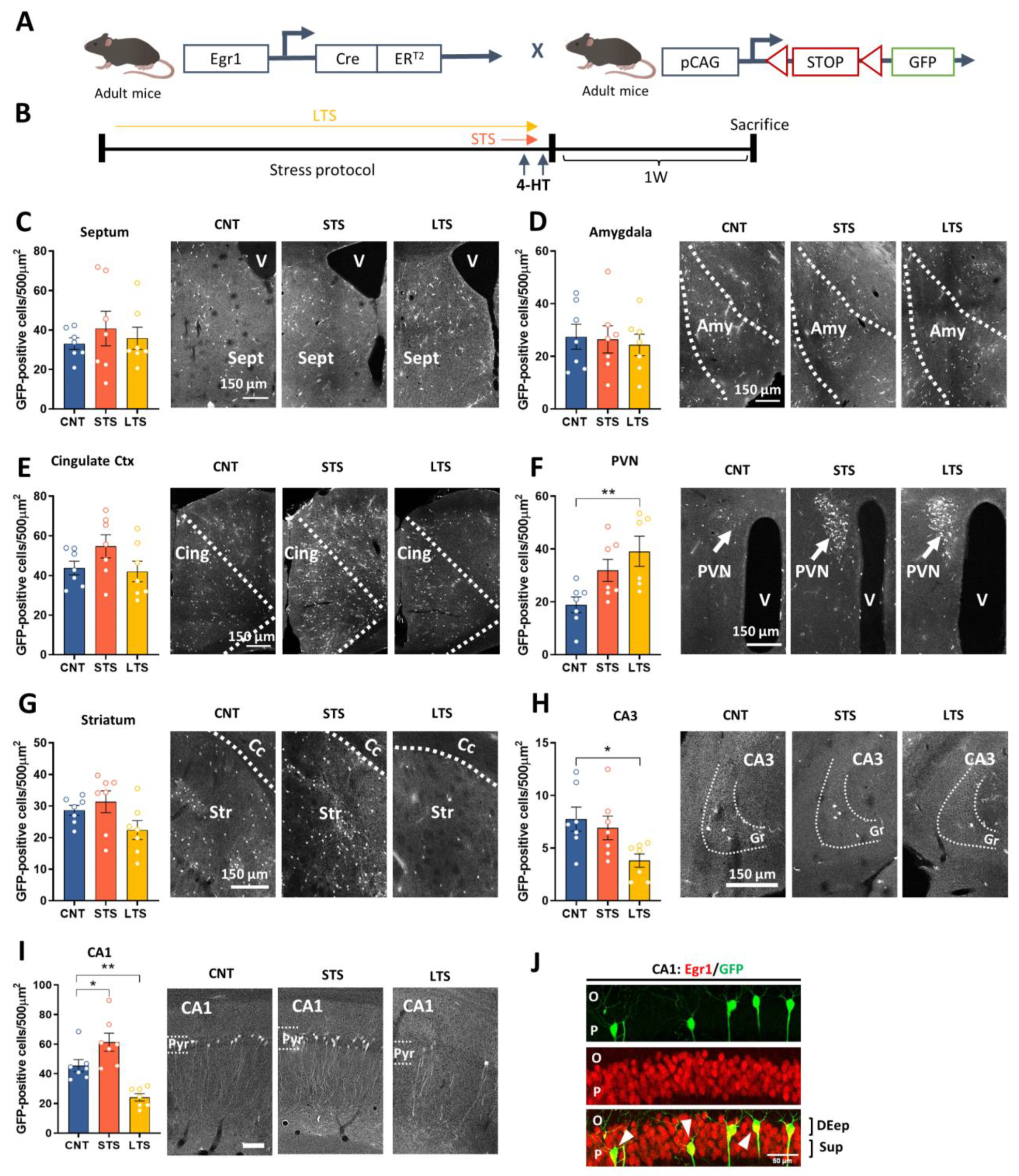
2.1. Egr1-Dependent Neural Activation in Hippocampal CA1 Depends on the Chronicity of the Stress
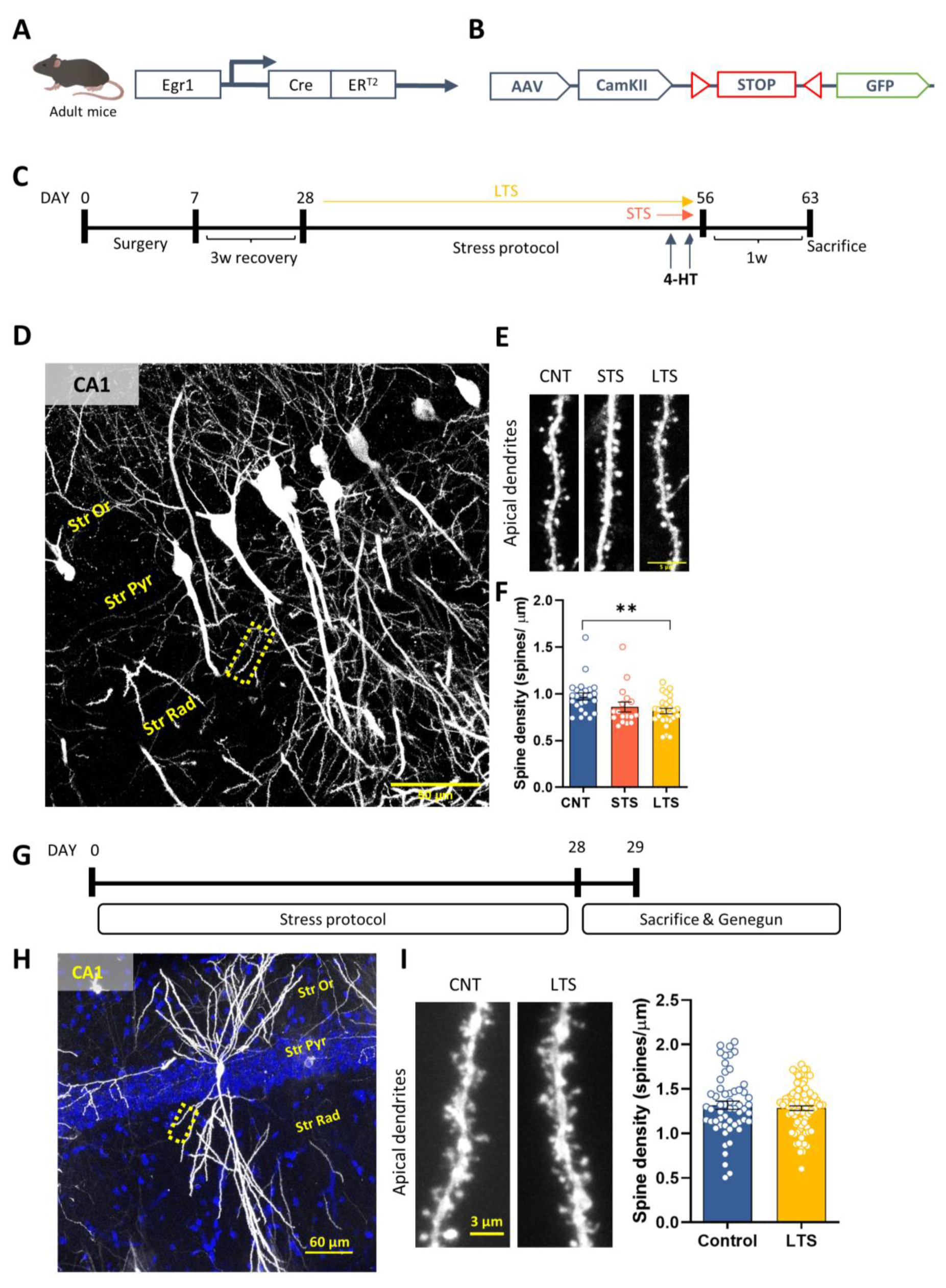
2.2. Specific Spine Density Alterations in Egr1-Positive Activated Cells
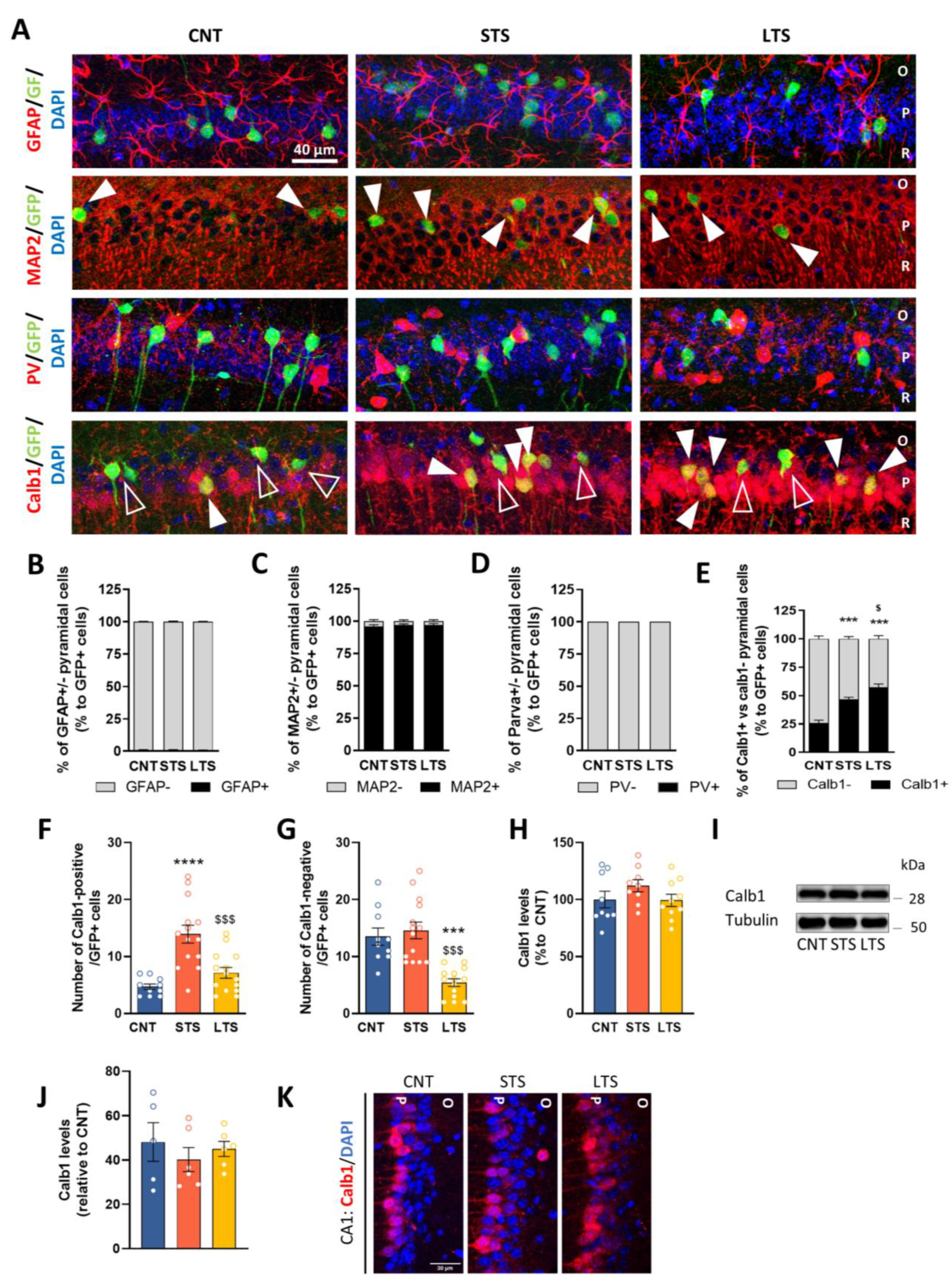
2.3. Different Hippocampal Subpopulations Are Activated Depending on Stress Duration
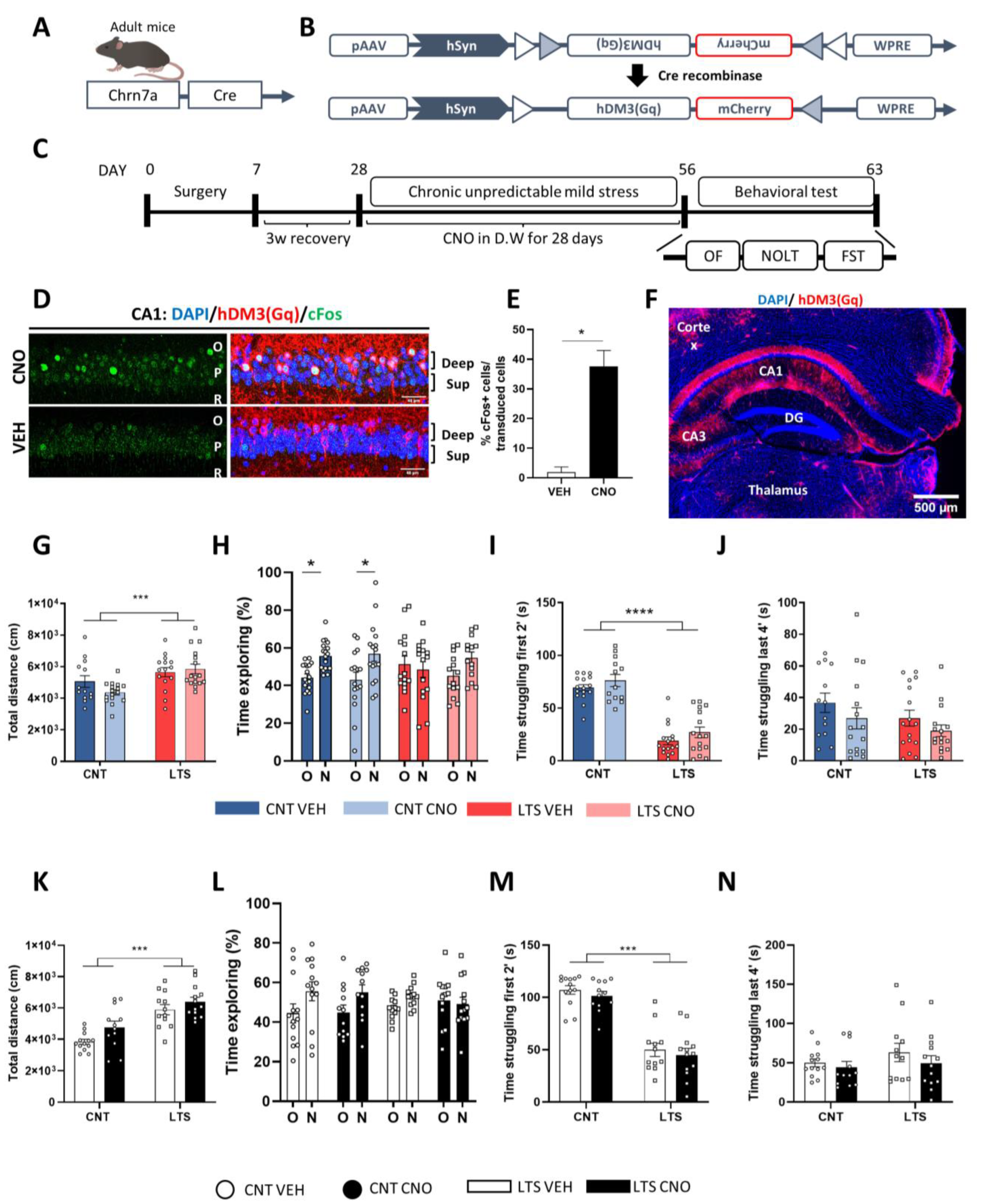
2.4. Chemogenetic Activation of Dorsal CA1 Deep Pyramidal Cells Does Not Prevent Sequelae Induced by CUMS
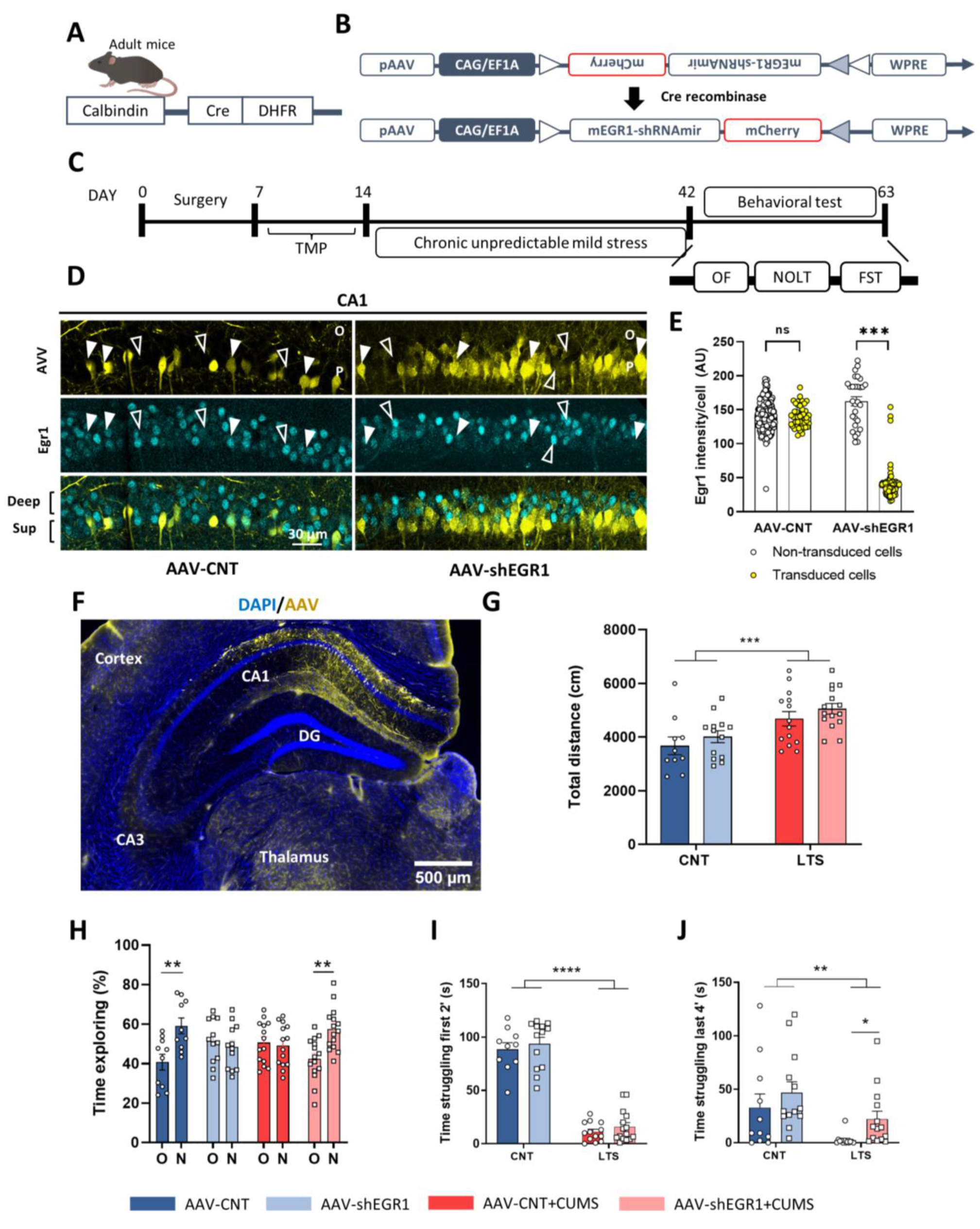
2.5. Specific Downregulation of Egr1 in CA1 Superficial Pyramidal Cells during CUMS Improves Depressive Sequelae
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Stereotaxic Surgery and Viral Transduction In Vivo
4.4. Chronic Unpredictable Mild Stress (CUMS)
4.5. Behavioral Tests
4.6. Tissue Fixation, Immunofluorescence and Confocal Imaging
4.7. Immunobloting
4.8. Gene Gun
4.9. Imaging Analysis
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, S.B.; Graham-Engeland, J.E.; Engeland, C.G.; Smyth, J.M.; Almeida, D.M.; Katz, M.J.; Lipton, R.B.; Mogle, J.A.; Munoz, E.; Ram, N.; et al. The Effects of Stress on Cognitive Aging, Physiology and Emotion (ESCAPE) Project. BMC Psychiatry 2015, 15, 146. [Google Scholar] [CrossRef]
- Vargas, T.; Conley, R.E.; Mittal, V.A. Chronic stress, structural exposures and neurobiological mechanisms: A stimulation, discrepancy and deprivation model of psychosis. Int. Rev. Neurobiol. 2020, 152, 41–69. [Google Scholar] [CrossRef]
- Nabeshima, T.; Kim, H.-C. Involvement of Genetic and Environmental Factors in the Onset of Depression. Exp. Neurobiol. 2013, 22, 235. [Google Scholar] [CrossRef]
- Richter-Levin, G.; Xu, L. How could stress lead to major depressive disorder? IBRO Rep. 2018, 4, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Van Oort, J.; Tendolkar, I.; Hermans, E.J.; Mulders, P.C.; Beckmann, C.F.; Schene, A.H.; Fernández, G.; van Eijndhoven, P.F. How the brain connects in response to acute stress: A review at the human brain systems level. Neurosci. Biobehav. Rev. 2017, 83, 281–297. [Google Scholar] [CrossRef]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353. [Google Scholar] [CrossRef]
- Schmaal, L.; Veltman, D.J.; Van Erp, T.G.M.; Smann, P.G.; Frodl, T.; Jahanshad, N.; Loehrer, E.; Tiemeier, H.; Hofman, A.; Niessen, W.J.; et al. Subcortical brain alterations in major depressive disorder: Findings from the ENIGMA Major Depressive Disorder working group. Mol. Psychiatry 2016, 21, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ge, T.; Leng, Y.; Pan, Z.; Fan, J.; Yang, W.; Cui, R. The Role of Neural Plasticity in Depression: From Hippocampus to Prefrontal Cortex. Neural Plast. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Jarome, T.; Li, S.-J.; Kim, J.J.; Helmstetter, F.J. Chronic stress selectively reduces hippocampal volume in rats: A longitudinal MRI study. Neuroreport 2009, 20, 1554. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.M.; Yang, C.; Sawyer-Glover, A.M.; Moseley, M.E.; Schatzberg, A.F. Early life stress and inherited variation in monkey hippocampal Volumes. Arch. Gen. Psychiatry 2001, 58, 1145–1151. [Google Scholar] [CrossRef]
- Xu, Y.; Pan, J.; Sun, J.; Ding, L.; Ruan, L.; Reed, M.; Yu, X.; Klabnik, J.; Lin, D.; Li, J.; et al. Inhibition of phosphodiesterase 2 reverses impaired cognition and neuronal remodeling caused by chronic stress. Neurobiol. Aging 2015, 36, 955–970. [Google Scholar] [CrossRef] [PubMed]
- Alfarez, D.N.; Joëls, M.; Krugers, H.J. Chronic unpredictable stress impairs long-term potentiation in rat hippocampal CA1 area and dentate gyrus in vitro. Eur. J. Neurosci. 2003, 17, 1928–1934. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Li, M.-X.; Xu, C.; Chen, H.-B.; An, S.-C.; Ma, X.-M. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016, 2016, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Magariños, A.M.; Li, C.J.; Gal Toth, J.; Bath, K.G.; Jing, D.; Lee, F.S.; McEwen, B.S. Effect of brain-derived neurotrophic factor haploinsufficiency on stress-induced remodeling of hippocampal neurons. Hippocampus 2011, 21, 253–264. [Google Scholar] [CrossRef]
- Pawlak, R.; Rao, B.S.S.; Melchor, J.P.; Chattarji, S.; McEwen, B.; Strickland, S. Tissue plasminogen activator and plasminogen mediate stress-induced decline of neuronal and cognitive functions in the mouse hippocampus. Proc. Natl. Acad. Sci. USA 2005, 102, 18201. [Google Scholar] [CrossRef] [PubMed]
- Shors, T.J.; Falduto, J.; Leuner, B. The opposite effects of stress on dendritic spines in male vs. female rats are NMDA receptor-dependent. Eur. J. Neurosci. 2004, 19, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Shors, T.J.; Chua, C.; Falduto, J. Sex Differences and Opposite Effects of Stress on Dendritic Spine Density in the Male Versus Female Hippocampus. J. Neurosci. 2001, 21, 6292–6297. [Google Scholar] [CrossRef]
- Cembrowski, M.S.; Bachman, J.L.; Wang, L.; Sugino, K.; Shields, B.C.; Spruston, N. Spatial Gene-Expression Gradients Underlie Prominent Heterogeneity of CA1 Pyramidal Neurons. Neuron 2016, 89, 351–368. [Google Scholar] [CrossRef]
- Valero, M.; de la Prida, L.M. The hippocampus in depth: A sublayer-specific perspective of entorhinal–hippocampal function. Curr. Opin. Neurobiol. 2018, 52, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Valero, M.; Cid, E.; Averkin, R.G.; Aguilar, J.; Sanchez-Aguilera, A.; Viney, T.J.; Gomez-Dominguez, D.; Bellistri, E.; De La Prida, L.M. Determinants of different deep and superficial CA1 pyramidal cell dynamics during sharp-wave ripples. Nat. Neurosci. 2015, 18, 1281–1290. [Google Scholar] [CrossRef]
- Gallo, F.T.; Katche, C.; Morici, J.F.; Medina, J.H.; Weisstaub, N.V. Immediate Early Genes, Memory and Psychiatric Disorders: Focus on c-Fos, Egr1 and Arc. Front. Behav. Neurosci. 2018, 12, 79. [Google Scholar] [CrossRef]
- Duclot, F.; Kabbaj, M. The Role of Early Growth Response 1 (EGR1) in Brain Plasticity and Neuropsychiatric Disorders. Front. Behav. Neurosci. 2017, 11, 35. [Google Scholar] [CrossRef]
- Girotti, M.; Pace, T.W.W.; Gaylord, R.I.; Rubin, B.A.; Herman, J.P.; Spencer, R.L. Habituation to repeated restraint stress is associated with lack of stress-induced c-fos expression in primary sensory processing areas of the rat brain. Neuroscience 2006, 138, 1067–1081. [Google Scholar] [CrossRef] [PubMed]
- Kozlovsky, N.; Matar, M.A.; Kaplan, Z.; Zohar, J.; Cohen, H. A distinct pattern of intracellular glucocorticoid-related responses is associated with extreme behavioral response to stress in an animal model of post-traumatic stress disorder. Eur. Neuropsychopharmacol. 2009, 19, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Musazzi, L.; Tornese, P.; Sala, N.; Popoli, M. Acute or Chronic? A Stressful Question. Trends Neurosci. 2017, 40, 525–535. [Google Scholar] [CrossRef]
- Laine, M.A.; Sokolowska, E.; Dudek, M.; Callan, S.A.; Hyytiä, P.; Hovatta, I. Brain activation induced by chronic psychosocial stress in mice. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Fee, C.; Prevot, T.; Misquitta, K.; Banasr, M.; Sibille, E. Chronic Stress-induced Behaviors Correlate with Exacerbated Acute Stress-induced Cingulate Cortex and Ventral Hippocampus Activation. Neuroscience 2020, 440, 113–129. [Google Scholar] [CrossRef]
- Flak, J.N.; Solomon, M.B.; Jankord, R.; Krause, E.G.; Herman, J.P. Identification of chronic stress-activated regions reveals a potential recruited circuit in rat brain. Eur. J. Neurosci. 2012, 36, 2547–2555. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Floriou-Servou, A.; von Ziegler, L.; Waag, R.; Schläppi, C.; Germain, P.L.; Bohacek, J. The Acute Stress Response in the Multiomic Era. Biol. Psychiatry 2021, 89, 1116–1126. [Google Scholar] [CrossRef]
- Longueville, S.; Nakamura, Y.; Brami-Cherrier, K.; Coura, R.; Hervé, D.; Girault, J.-A. Long-lasting tagging of neurons activated by seizures or cocaine administration in Egr1-CreERT2 transgenic mice. Eur. J. Neurosci. 2021, 53, 1450–1472. [Google Scholar] [CrossRef] [PubMed]
- Brito, V.; Montalban, E.; Sancho-Balsells, A.; Pupak, A.; Flotta, F.; Masana, M.; Ginés, S.; Alberch, J.; Martin, C.; Girault, J.-A.; et al. Hippocampal Egr1-dependent neuronal ensembles negatively regulate motor learning. J. Neurosci. 2022, 42, 5346–5360. [Google Scholar] [CrossRef] [PubMed]
- Román-Albasini, L.; Díaz-Véliz, G.; Olave, F.A.; Aguayo, F.I.; García-Rojo, G.; Corrales, W.A.; Silva, J.P.; Ávalos, A.M.; Rojas, P.S.; Aliaga, E.; et al. Antidepressant-relevant behavioral and synaptic molecular effects of long-term fasudil treatment in chronically stressed male rats. Neurobiol. Stress 2020, 13, 100234. [Google Scholar] [CrossRef]
- Roustazade, R.; Radahmadi, M.; Yazdani, Y. Therapeutic effects of saffron extract on different memory types, anxiety, and hippocampal BDNF and TNF-α gene expressions in sub-chronically stressed rats. Nutr. Neurosci. 2021, 25, 192–206. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.B.; Adamec, R.E.; Thompson, B.L. Expression of egr-1 (zif268) mRNA in select fear-related brain regions following exposure to a predator. Behav. Brain Res. 2005, 162, 279–288. [Google Scholar] [CrossRef]
- Hall, J.; Thomas, K.L.; Everitt, B.J. Cellular Imaging of Zif268 Expression in the Hippocampus and Amygdala during Contextual and Cued Fear Memory Retrieval: Selective Activation of Hippocampal CA1 Neurons during the Recall of Contextual Memories. J. Neurosci. 2001, 21, 2186–2193. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.S.; Tocco, G.; Shors, T.J.; Thompson, R.F. Activation of immediate early genes after acute stress. Neuroreport 1991, 2, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Melia, K.R.; Ryabinin, A.E.; Schroeder, R.; Bloom, F.E.; Wilson, M.C.; Anokhin, P.K. Induction and Habituation of Immediate Early Gene Expression in Rat Brain by Acute and Repeated Restraint Stress. J. Neurosci. 1994, 14, 5929–5938. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, W.E.; Herman, J.P.; Battaglia, D.F.; Akil, H.; Watson, S.J. Pattern and time course of immediate early gene expression in rat brain following acute stress. Neuroscience 1995, 64, 477–505. [Google Scholar] [CrossRef]
- Olsson, T.; Seckl, J.R. Ketanserin selectively blocks acute stress-induced changes in ngf1-a and mineralocorticoid receptor gene expression in hippocampal neurons. Pergamon Neurosci. 1997, 76, 441–448. [Google Scholar] [CrossRef]
- Knapska, E.; Kaczmarek, L. A gene for neuronal plasticity in the mammalian brain: Zif268/Egr-1/NGFI-A/Krox-24/TIS8/ZENK? Prog. Neurobiol. 2004, 74, 183–211. [Google Scholar] [CrossRef]
- Covington, H.E.; Lobo, M.K.; Maze, I.; Vialou, V.; Hyman, J.M.; Zaman, S.; LaPlant, Q.; Mouzon, E.; Ghose, S.; Tamminga, C.A.; et al. Antidepressant Effect of Optogenetic Stimulation of the Medial Prefrontal Cortex. J. Neurosci. 2010, 30, 16082–16090. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ono, K.; Ouchi, H.; Tsushima, R.; Murakami, Y. Social isolation stress down-regulates cortical early growth response 1 (Egr-1) expression in mice. Neurosci. Res. 2012, 73, 257–262. [Google Scholar] [CrossRef]
- Hodges, T.E.; Green, M.R.; Simone, J.J.; McCormick, C.M. Effects of social context on endocrine function and Zif268 expression in response to an acute stressor in adolescent and adult rats. Int. J. Dev. Neurosci. 2014, 35, 25–34. [Google Scholar] [CrossRef]
- Northcutt, K.V.; Lonstein, J.S. Social contact elicits immediate-early gene expression in dopaminergic cells of the male prairie vole extended olfactory amygdala. Neuroscience 2009, 163, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, H.; Lv, Y.; Liu, C.; Sun, W.; Tian, L. Downregulation of Egr-1 Expression Level via GluN2B Underlies the Antidepressant Effects of Ketamine in a Chronic Unpredictable Stress Animal Model of Depression. Neuroscience 2018, 372, 38–45. [Google Scholar] [CrossRef]
- Ieraci, A.; Mallei, A.; Popoli, M. Social Isolation Stress Induces Anxious-Depressive-Like Behavior and Alterations of Neuroplasticity-Related Genes in Adult Male Mice. Neural Plast. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Clements, K.M.; Wainwright, P.E. Swim stress increases hippocampal Zif268 expression in the spontaneously hypertensive rat. Brain Res. Bull. 2010, 82, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Isingrini, E.; Camus, V.; Le Guisquet, A.M.; Pingaud, M.; Devers, S.; Belzung, C. Association between repeated unpredictable chronic mild stress (UCMS) procedures with a high fat diet: A model of fluoxetine resistance in mice. PLoS ONE 2010, 5, e10404. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Rex, C.S.; Rice, C.J.; Dubé, C.M.; Gall, C.M.; Lynch, G.; Barama, T.Z. Correlated memory defects and hippocampal dendritic spine loss after acute stress involve corticotropin-releasing hormone signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 13123–13128. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.S.; Zhong, G.; Cao, H.X.; Hu, Y.; Hong, X.Y.; Li, T.; Li, X.; Liu, Q.; Wang, Q.; Ke, D.; et al. Repeated Restraint Stress Led to Cognitive Dysfunction by NMDA Receptor-Mediated Hippocampal CA3 Dendritic Spine Impairments in Juvenile Sprague-Dawley Rats. Front. Mol. Neurosci. 2020, 13, 552787. [Google Scholar] [CrossRef]
- Krzystyniak, A.; Baczynska, E.; Magnowska, M.; Antoniuk, S.; Roszkowska, M.; Zareba-Koziol, M.; Das, N.; Basu, S.; Pikula, M.; Wlodarczyk, J. Prophylactic Ketamine Treatment Promotes Resilience to Chronic Stress and Accelerates Recovery: Correlation with Changes in Synaptic Plasticity in the CA3 Subregion of the Hippocampus. Int. J. Mol. Sci. 2019, 20, 1726. [Google Scholar] [CrossRef]
- Qu, Y.; Yang, C.; Ren, Q.; Ma, M.; Dong, C.; Hashimoto, K.; Regional, H.K. Regional differences in dendritic spine density confer resilience to chronic social defeat stress. Acta Neuropsychiatr. 2017, 30, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Mclaughlin, K.J.; Baran, S.E.; Wright, R.L.; Conrad, C.D. Chronic stress enhances spatial memory in ovariectomized female rats despite CA3 dendritic retraction: Possible involvement of CA1 neurons. Neuroscience 2005, 135, 1045–1054. [Google Scholar] [CrossRef]
- Qiao, H.; An, S.C.; Ren, W.; Ma, X.M. Progressive alterations of hippocampal CA3-CA1 synapses in an animal model of depression. Behav. Brain Res. 2014, 275, 191–200. [Google Scholar] [CrossRef]
- Patel, D.; Anilkumar, S.; Chattarji, S.; Buwalda, B. Repeated social stress leads to contrasting patterns of structural plasticity in the amygdala and hippocampus. Behav. Brain Res. 2018, 347, 314–324. [Google Scholar] [CrossRef] [PubMed]
- Leem, Y.-H.; Park, J.-S.; Chang, H.; Park, J.; Kim, H.-S. Exercise Prevents Memory Consolidation Defects Via Enhancing Prolactin Responsiveness of CA1 Neurons in Mice Under Chronic Stress. Mol. Neurobiol. 2019, 56, 6609–6625. [Google Scholar] [CrossRef] [PubMed]
- Donohue, H.S.; Gabbott, P.L.A.; Davies, H.A.; Rodríguez, J.J.; Cordero, M.I.; Sandi, C.; Medvedev, N.I.; Popov, V.I.; Colyer, F.M.; Peddie, C.J.; et al. Chronic restraint stress induces changes in synapse morphology in stratum lacunosum-moleculare CA1 rat hippocampus: A stereological and three-dimensional ultrastructural study. Neuroscience 2006, 140, 597–606. [Google Scholar] [CrossRef]
- Conrad, C.D.; Mclaughlin, K.J.; Huynh, T.N.; El-Ashmawy, M.; Sparks, M. Chronic Stress and a Cyclic Regimen of Estradiol Administration Separately Facilitate Spatial Memory: Relationship with CA1 Spine Density and Dendritic Complexity. Behav. Neurosci. 2012, 126, 142. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.J.; Wilson, J.O.; Harman, J.; Wright, R.L.; Wieczorek, L.; Gomez, J.; Korol, D.L.; Conrad, C.D. Chronic 17β-estradiol or cholesterol prevents stress-induced hippocampal CA3 dendritic retraction in ovariectomized female rats: Possible correspondence between CA1 spine properties and spatial acquisition. Hippocampus 2010, 20, 768–786. [Google Scholar] [CrossRef]
- Qin, X.; Jiang, Y.; Tse, Y.C.; Wang, Y.; Wong, T.P.; Paudel, H.K. Early Growth Response 1 (Egr-1) Regulates N-Methyl-d-aspartate Receptor (NMDAR)-dependent Transcription of PSD-95 and α-Amino-3-hydroxy-5-methyl-4-isoxazole Propionic Acid Receptor (AMPAR) Trafficking in Hippocampal Primary Neurons. J. Biol. Chem. 2015, 290, 29603–29616. [Google Scholar] [CrossRef] [PubMed]
- Krugers, H.J.; Koolhaas, J.M.; Medema, R.M.; Korf, J. Prolonged subordination stress increases Calbindin-D28k immunoreactivity in the rat hippocampal CA1 area. Brain Res. 1996, 729, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Xie, X.M.; Yu, J.Y.; Sun, Y.X.; Liao, X.M.; Wang, X.X.; Su, Y.A.; Liu, Y.J.; Schmidt, M.V.; Wang, X.D.; et al. Suppressed Calbindin Levels in Hippocampal Excitatory Neurons Mediate Stress-Induced Memory Loss. Cell Rep. 2017, 21, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Revest, J.M.; Di Blasi, F.; Kitchener, P.; Rougé-Pont, F.; Desmedt, A.; Turiault, M.; Tronche, F.; Piazza, P.V. The MAPK pathway and Egr-1 mediate stress-related behavioral effects of glucocorticoids. Nat. Neurosci. 2005 85 2005, 8, 664–672. [Google Scholar] [CrossRef]
- Revest, J.M.; Kaouane, N.; Mondin, M.; Le Roux, A.; Rougé-Pont, F.; Vallée, M.; Barik, J.; Tronche, F.; Desmedt, A.; Piazza, P.V. The enhancement of stress-related memory by glucocorticoids depends on synapsin-Ia/Ib. Mol. Psychiatry 2010, 15, 1140–1151. [Google Scholar] [CrossRef]
- Saunderson, E.A.; Spiers, H.; Mifsud, K.R.; Gutierrez-Mecinas, M.; Trollope, A.F.; Shaikh, A.; Mill, J.; Reul, J.M.H.M. Stress-induced gene expression and behavior are controlled by DNA methylation and methyl donor availability in the dentate gyrus. Proc. Natl. Acad. Sci. USA 2016, 113, 4830–4835. [Google Scholar] [CrossRef]
- Davis, S.; Renaudineau, S.; Poirier, R.; Poucet, B.; Save, E.; Laroche, S. The formation and stability of recognition memory: What happens upon recall? Front. Behav. Neurosci. 2010, 4, 177. [Google Scholar] [CrossRef]
- Besnard, A.; Caboche, J.; Laroche, S. Recall and Reconsolidation of Contextual Fear Memory: Differential Control by ERK and Zif268 Expression Dosage. PLoS ONE 2013, 8, 72006. [Google Scholar] [CrossRef]
- Penke, Z.; Morice, E.; Veyrac, A.; Gros, A.; Chagneau, C.; LeBlanc, P.; Samson, N.; Baumgärtel, K.; Mansuy, I.M.; Davis, S.; et al. Zif268/Egr1 gain of function facilitates hippocampal synaptic plasticity and long-term spatial recognition memory. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130159. [Google Scholar] [CrossRef] [PubMed]
- Fernández-García, S.; Sancho-Balsells, A.; Longueville, S.; Hervé, D.; Gruart, A.; Delgado-García, J.M.; Alberch, J.; Giralt, A. Astrocytic BDNF and TrkB regulate severity and neuronal activity in mouse models of temporal lobe epilepsy. Cell Death Dis. 2020, 11, 411. [Google Scholar] [CrossRef]
- Zhan, J.; Komal, R.; Keenan, W.T.; Hattar, S.; Fernandez, D.C. Non-invasive Strategies for Chronic Manipulation of DREADD-controlled Neuronal Activity. JoVE 2019, 150, e59439. [Google Scholar]
- Montalban, E.; Al-Massadi, O.; Sancho-Balsells, A.; Brito, V.; de Pins, B.; Alberch, J.; Ginés, S.; Girault, J.-A.; Giralt, A. Pyk2 in the amygdala modulates chronic stress sequelae via PSD-95-related micro-structural changes. Transl. Psychiatry 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Giralt, A.; Brito, V.; Pardo, M.; Rubio, S.E.; Marion-Poll, L.; Martín-Ibáñez, R.; Zamora-Moratalla, A.; Bosch, C.; Ballesteros, J.J.; Blasco, E.; et al. Helios modulates the maturation of a CA1 neuronal subpopulation required for spatial memory formation. Exp. Neurol. 2020, 323, 113095. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GROUP | ||||
|---|---|---|---|---|
| TIME | STRESSOR | CNT | STS | LTS |
| Day 0 | Isolation | Isolation | ||
| Day 1 | Home cage inclination (1 h) | |||
| Day 2 | Food deprivation (24 h) | |||
| Day 3 | Restraint (1 h) | |||
| Day 4 | Water deprivation (24 h) | |||
| Day 5 | Light–dark cycle alterations (24 h) | |||
| Day 6 | Forced Swim (5 min) | |||
| Day 7 | Exposition to rat sawdust (4 h) | |||
| Day 8 | Home cage inclination (1 h) | |||
| Day 9 | Water deprivation (24 h) | |||
| Day 10 | Restraint (1 h) | |||
| Day 11 | Light–dark cycle alterations (24 h) | |||
| Day 12 | Food deprivation (24 h) | |||
| Day 13 | Restraint (1 h) | |||
| Day 14 | Exposition to rat sawdust (4 h) | |||
| Day 15 | Home cage inclination (1 h) | |||
| Day 16 | Light–dark cycle alterations (24 h) | |||
| Day 17 | Forced Swim (5 min) | |||
| Day 18 | Exposition to rat sawdust (4 h) | |||
| Day 19 | Water deprivation (24 h) | |||
| Day 20 | Forced Swim (5 min) | |||
| Day 21 | Exposition to rat sawdust (4 h) | |||
| Day 22 | Home cage inclination (1 h) | |||
| Day 23 | Food deprivation (24 h) | |||
| Day 24 | Water deprivation (24 h) | |||
| Day 25 | Light–dark cycle alterations (24 h) | |||
| Day 26 | Food deprivation (24 h) | Isolation | ||
| Day 27 | Restraint (1 h) | +4-HT | +4-HT | +4-HT |
| Day 28 | Forced Swim (5 min) | +4-HT | +4-HT | +4-HT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho-Balsells, A.; Borràs-Pernas, S.; Brito, V.; Alberch, J.; Girault, J.-A.; Giralt, A. Cognitive and Emotional Symptoms Induced by Chronic Stress Are Regulated by EGR1 in a Subpopulation of Hippocampal Pyramidal Neurons. Int. J. Mol. Sci. 2023, 24, 3833. https://doi.org/10.3390/ijms24043833
Sancho-Balsells A, Borràs-Pernas S, Brito V, Alberch J, Girault J-A, Giralt A. Cognitive and Emotional Symptoms Induced by Chronic Stress Are Regulated by EGR1 in a Subpopulation of Hippocampal Pyramidal Neurons. International Journal of Molecular Sciences. 2023; 24(4):3833. https://doi.org/10.3390/ijms24043833
Chicago/Turabian StyleSancho-Balsells, Anna, Sara Borràs-Pernas, Verónica Brito, Jordi Alberch, Jean-Antoine Girault, and Albert Giralt. 2023. "Cognitive and Emotional Symptoms Induced by Chronic Stress Are Regulated by EGR1 in a Subpopulation of Hippocampal Pyramidal Neurons" International Journal of Molecular Sciences 24, no. 4: 3833. https://doi.org/10.3390/ijms24043833
APA StyleSancho-Balsells, A., Borràs-Pernas, S., Brito, V., Alberch, J., Girault, J.-A., & Giralt, A. (2023). Cognitive and Emotional Symptoms Induced by Chronic Stress Are Regulated by EGR1 in a Subpopulation of Hippocampal Pyramidal Neurons. International Journal of Molecular Sciences, 24(4), 3833. https://doi.org/10.3390/ijms24043833