

Altered Mesenchymal Stem Cells Mechanotransduction from Oxidized Collagen: Morphological and Biophysical Observations

 , , ,
, , ,
Abstract
1. Introduction
2. Results
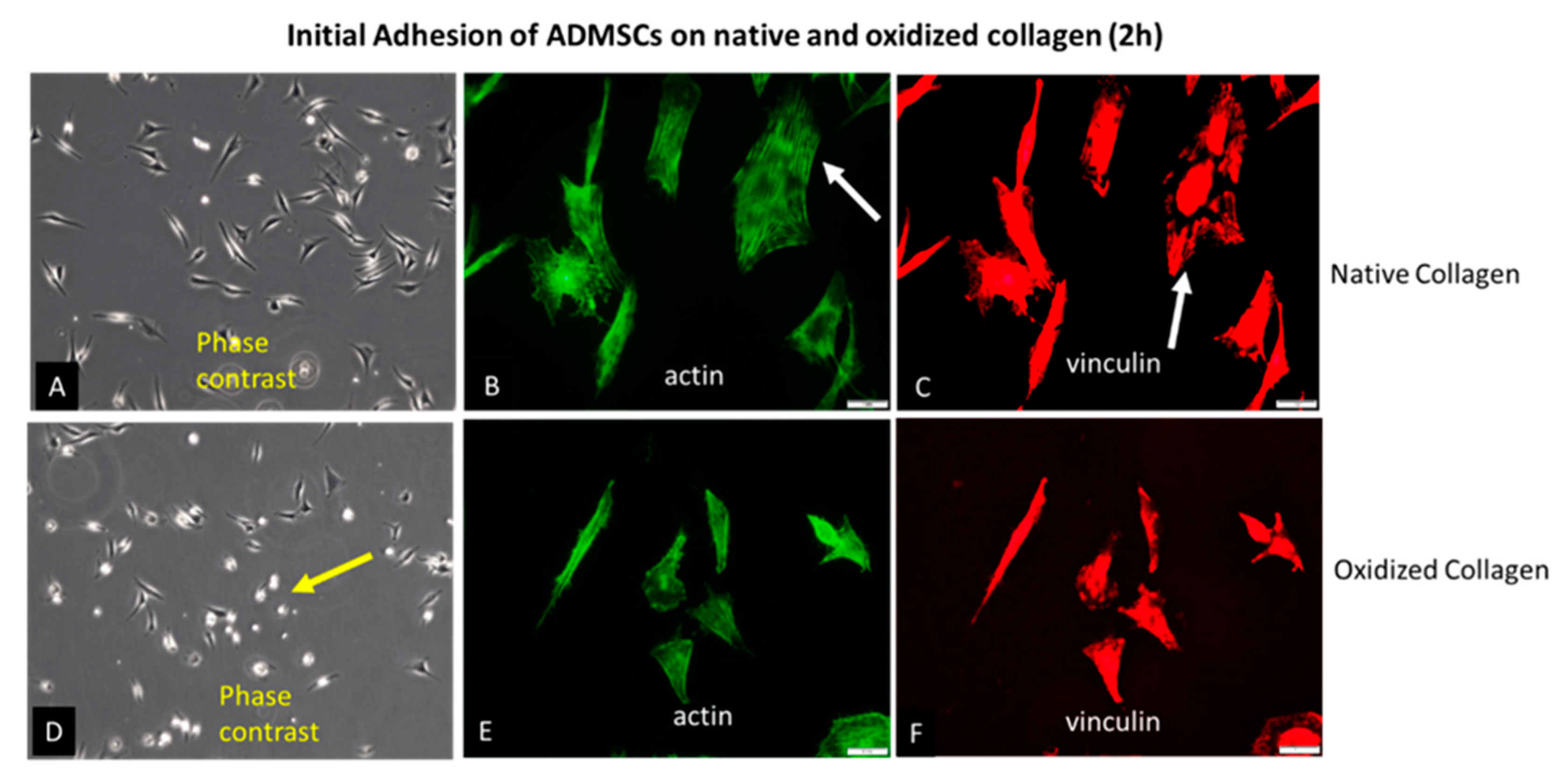
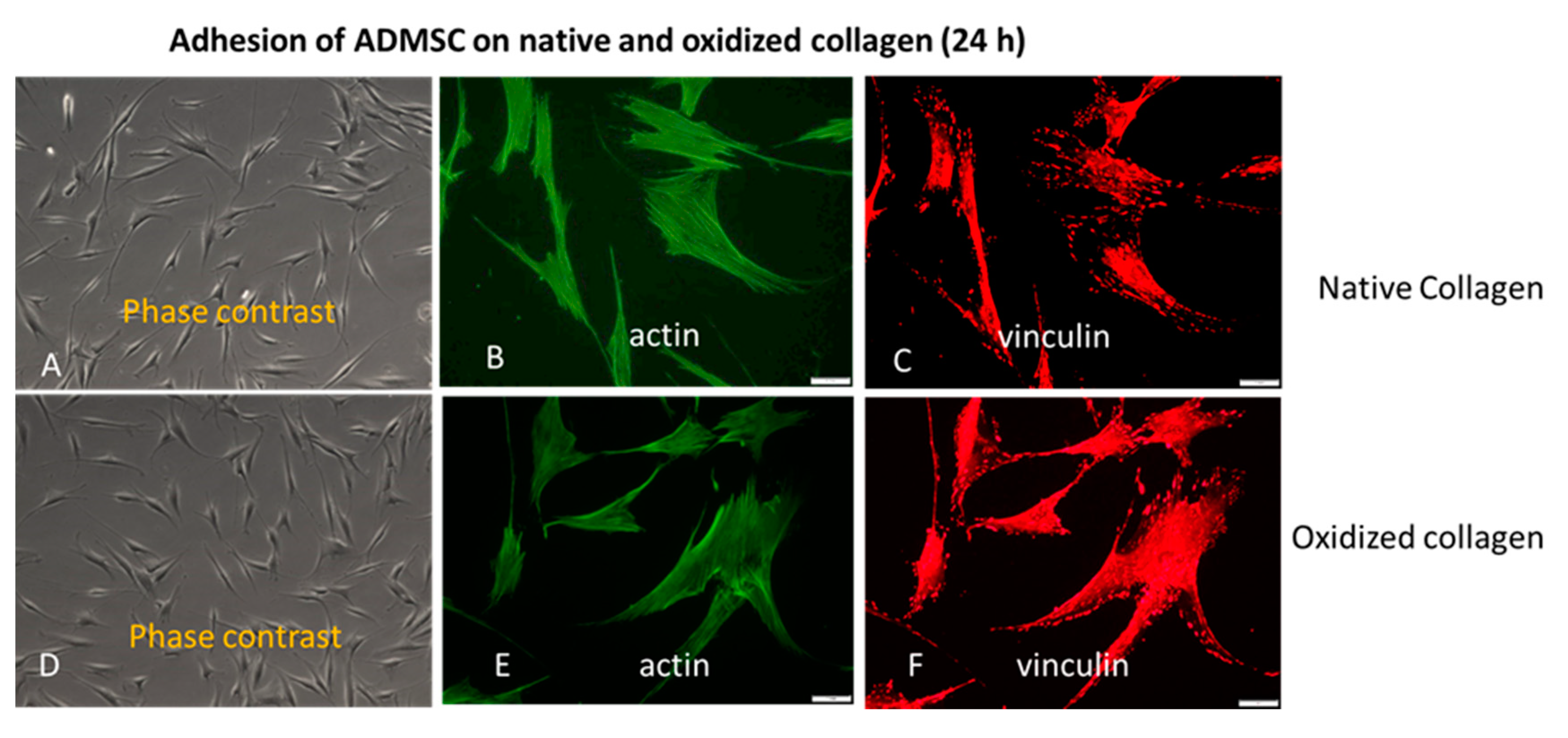
2.1. Initial Cell Attachment
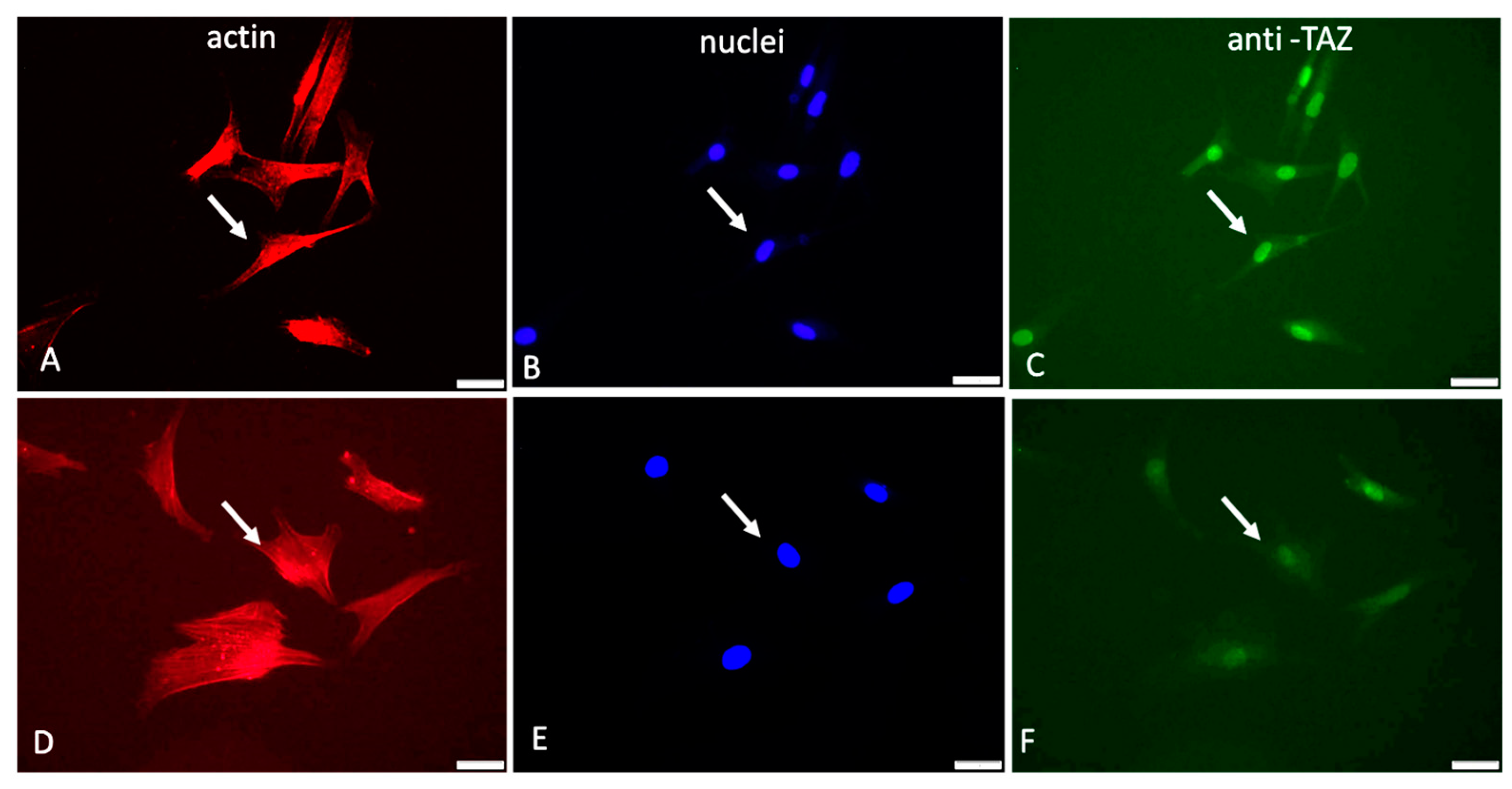
2.2. YAP/TAZ Signaling Events
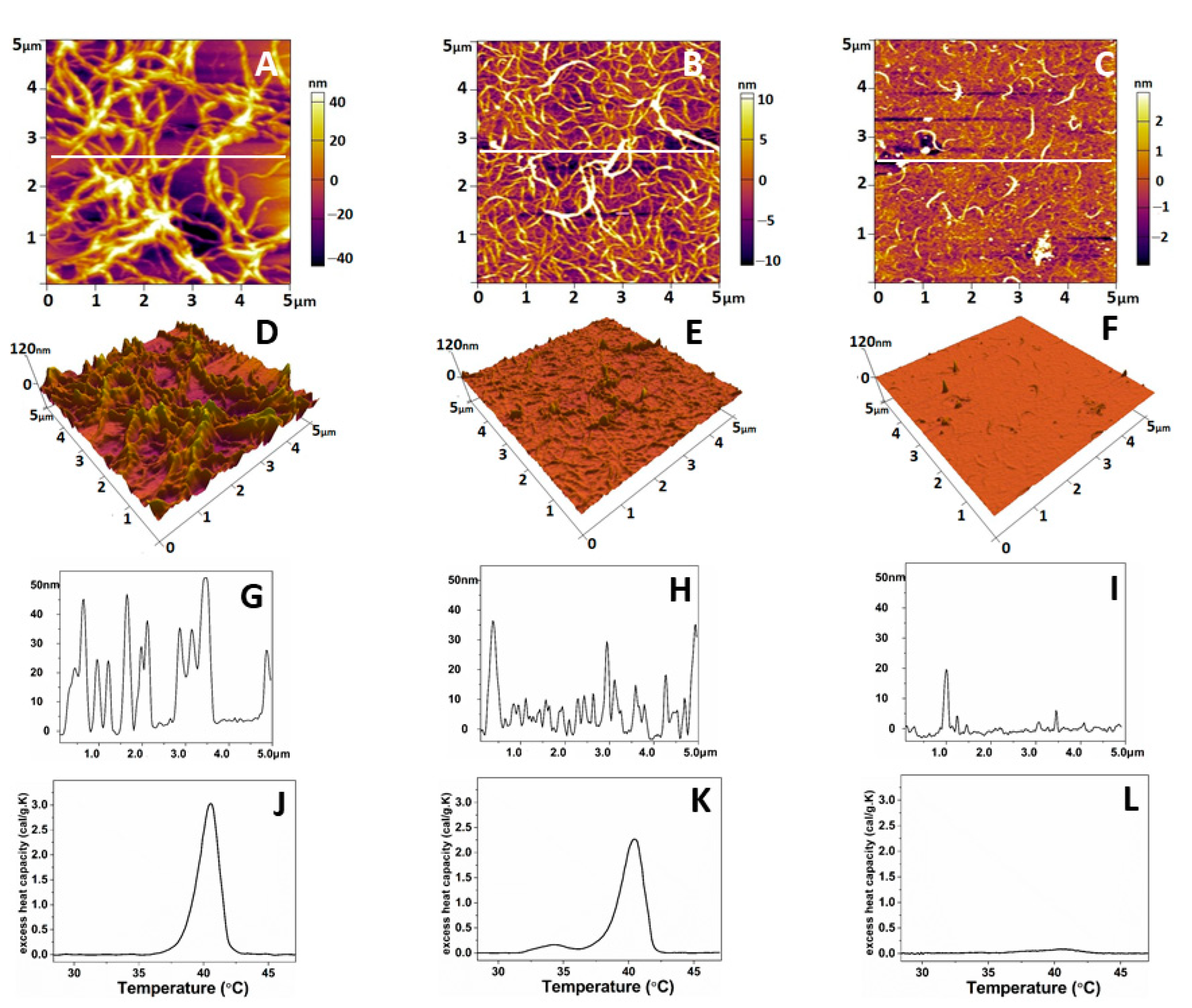
2.3. Comparative Atomic Force Microscopy (AFM) Study
3. Discussion
4. Materials and Methods
4.1. Collagen Preparation
4.2. Collagen Oxidation Procedure
4.3. Cells
4.4. Morphological Study
4.4.1. First Protocol (Cell Spreading and FA Formation)
4.4.2. Second Protocol (YAP/TAZ Signaling Events)
4.5. Image Analysis
4.5.1. Quantitative Analysis of Raw Format Images by ImageJ
4.5.2. Quantification of Overall Morphological Parameters
4.5.3. YAP/TAZ Signaling
4.5.4. Quantification of Focal Adhesions (FA)
4.6. AFM Studies
4.7. DSC Measurements
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burridge, K.; Monaghan Benson, E.; Graham, D.M. Mechanotransduction: From the cell surface to the nucleus via RhoA. Phil. Trans. R. Soc. 2019, 374, 20180229. [Google Scholar] [CrossRef]
- Yamashiroa, Y.; Quoc Thangb, B.; Ramireza, K.; Jae Shina, S.; Kohatae, T.; Ohataf, S.; Anh Vu Nguyena, T.; Ohtsukie, S.; Nagayamaf, K.; Yanagisawaa, H. Matrix mechanotransduction mediated by thrombospondin-1/integrin/YAP in the vascular remodeling. Proc. Natl. Acad. Sci. USA 2020, 117, 9896–9905. [Google Scholar] [CrossRef] [PubMed]
- Vining, K.H.; Mooney, D.J. Mechanical forces direct stem cell behavior in development and regeneration. Nat. Rev. Mol. Cell Biol. 2017, 18, 728–742. [Google Scholar] [CrossRef] [PubMed]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of mechanical and cytoskeletal cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef]
- Li, D.; Zhou, J.; Chowdhury, F.; Cheng, J.; Wang, N.; Wang, F. Role of mechanical factors in fate decisions of stem cells. Regen. Med. 2011, 6, 229–240. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.K.; Kim, K.S. Intrinsic and extrinsic mechanical properties related to the differentiation of mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2016, 473, 752–757. [Google Scholar] [CrossRef]
- Hou, Y.; Xie, W.; Yu, L.; Cuellar Camacho, L.; Nie, C.; Zhang, M.; Haag, R.; Wei, Q. Surface Roughness Gradients Reveal Topography-Specific Mechanosensitive Responses in Human Mesenchymal Stem Cells. Small 2020, 16, 1905422. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, G.; Luo, X.; Qiu, J.; Tang, C. Substrate stiffness regulates the proliferation, migration, and differentiation of epidermal cells. Burns 2012, 38, 414–420. [Google Scholar] [CrossRef]
- Shalabi, M.M.; Gortemaker, A.; Van’t Hof, M.A.; Jansen, J.A.; Creugers, N.H.J. Implant surface roughness and bone healing: A systematic review. J. Dent. Res. 2006, 85, 496–500. [Google Scholar] [CrossRef]
- Matos, G.R.M. Surface Roughness of Dental Implant and Osseointegration. Maxillofac. Oral Surg. 2021, 20, 1–4. [Google Scholar] [CrossRef]
- Llopis-Hernandez, V.; Rico, P.; Moratal, D.; Altankov, G.; Salmeron-Sanchez, M. Role of Material-Driven Fibronectin Fibrillogenesis. Acta Biomater. 2013, 77, 74–84. [Google Scholar]
- Klein, E.A.; Yin, L.; Kothapalli, D.; Castagnino, P.; Byfield, F.J.; Xu, T.; Levental, I.; Hawthorne, E.; Janmey, P.A.; Assoian, R.K. Cell-cycle control by physiological matrix elasticity and in vivo tissue stiffening. Curr. Biol. 2009, 19, 1511–1518. [Google Scholar] [CrossRef]
- Nedjari, S.; Awaja, F.; Altankov, G. Three Dimensional Honeycomb Patterned Fibrinogen Based Nanofibers Induce Substantial Osteogenic Response of Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 15947. [Google Scholar] [CrossRef]
- Kim, D.; Provenzano, P.P.; Smith, C.L.; Levchenko, A. Matrix nanotopography as a regulator of cell function. J. Cell Biol. 2012, 197, 351. [Google Scholar] [CrossRef]
- Sun, Z.; Guo, S.S.; Fässler, R. Integrin-mediated mechanotransduction. J. Cell Biol. 2016, 215, 445–456. [Google Scholar] [CrossRef]
- Majhy, B.; Priyadarshinia, P.; Sen, A.K. Effect of surface energy and roughness on cell adhesion and growth—Facile surface modification for enhanced cell culture. RSC Adv. 2021, 11, 15467. [Google Scholar] [CrossRef]
- Horbett, T.A. The role of adsorbed proteins in animal cell adhesion. Colloids Surf. B Biointerfaces 1994, 2, 225–240. [Google Scholar] [CrossRef]
- Dupont, S.; Morsut, L.; Aragona, M.; Enzo, E.; Giulitti, S.; Cordenonsi, M.; Zanconato, F.; Le Digabel, J.; Forcato, M.; Bicciato, S.; et al. Role of YAP/TAZ in mechanotransduction. Nature 2011, 474, 179–183. [Google Scholar] [CrossRef]
- Meng, Z.; Moroishi, T.; Guan, K. Mechanisms of Hippo pathway regulation. Genes Dev. 2016, 30, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zinatizadeh, M.R.; Miri, S.R.; Zarandi, P.K.; Chalbatani, G.M.; Rapôso, C.; Mirzaei, H.R.; Akbari, M.E.; Mahmoodzadeh, H. The Hippo Tumor Suppressor Pathway (YAP/TAZ/TEAD/MST/LATS) and EGFR-RAS-RAF-MEK in cancer metastasis. Genes Dis. 2019, 8, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Pobbati, A.; Hong, W. A combat with the YAP/TAZ-TEAD oncoproteins for cancer therapy. Theranostics 2020, 10, 3622–3635. [Google Scholar] [CrossRef] [PubMed]
- Janmey, P.A.; Wells, R.G.; Assoian, R.K.; McCulloch, C.A. From tissue mechanics to transcription factors. Differentiation 2013, 86, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Halder, G. The two faces of Hippo: Targeting the Hippo pathway for regenerative medicine and cancer treatment. Nat. Rev. Drug Discov. 2014, 13, 63–79. [Google Scholar] [CrossRef]
- Wang, K.C.; Yeh, Y.T.; Nguyen, P.; Limqueco, E.; Lopez, J.; Thorossian, S.; Guan, K.L.; Li, Y.S.J.; Chien, S. Flow-dependent YAP/TAZ activities regulate endothelial phenotypes and atherosclerosis. Proc. Natl. Acad. Sci. USA 2016, 113, 11525–11530. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Zhong, W. Regulation and mechanism of YAP/TAZ in the mechanical microenvironment of stem cells. Mol. Med. Rep. 2021, 24, 506. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef]
- Mohri, Z.; Del Rio Hernandez, A.; Krams, R. The emerging role of YAP/TAZ in mechanotransduction. J. Thorac. Dis. 2017, 9, E507–E509. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Exp. Cell Res. 2016, 343, 42–53. [Google Scholar] [CrossRef]
- Bao, M.; Xie, J.; Huck, W.T.S. Recent Advances in Engineering the Stem Cell Microniche in 3D. Adv. Sci. 2018, 5, 1800448. [Google Scholar] [CrossRef]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Heino, J. Cellular signaling by collagen-binding integrins. Adv. Exp. Med. Biol. 2014, 819, 143–155. [Google Scholar]
- Zeltz, C.; Gullberg, D.J. The integrin-collagen connection—A glue for tissue repair? J. Cell Sci. 2016, 129, 1284. [Google Scholar] [CrossRef]
- Myllyharju, J. Intracellular Post-Translational Modifications of Collagens. In Collagen Topics in Current Chemistry; Brinckmann, J., Notbohm, H., Müller, P.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 247. [Google Scholar] [CrossRef]
- Kennett, E.C.; Chuang, C.Y.; Degendorfer, G.; Whitelock, J.M.; Davies, M.J. Mechanisms and consequences of oxidative damage to extracellular matrix. Biochem. Soc. Trans. 2011, 39, 1279–1287. [Google Scholar] [CrossRef]
- Boin, F.; Erre, G.L.; Posadino, A.M.; Cossu, A.; Giordo, R.; Spinetti, G.; Passiu, G.; Emanueli, C.; Pintus, G. Oxidative stress-dependent activation of collagen synthesis is induced in human pulmonary smooth muscle cells by sera from patients with scleroderma-associated pulmonary hypertension. Orphanet J. Rare Dis. 2014, 9, 123. [Google Scholar] [CrossRef]
- Lu, P.; Takai, K.; Weaver, V.M.; Werb, Z. Extracellular matrix degradation and remodeling in development and disease. Cold Spring Harb. Perspect. Biol. 2011, 3, a005058. [Google Scholar] [CrossRef]
- Komsa-Penkova, R.; Stavreva, G.; Belemezova, K.; Kyurkchiev, S.; Todinova, S.; Altankov, G. Mesenchymal Stem-Cell Remodeling of Adsorbed Type-I Collagen-The Effect of Collagen Oxidation. Int. J. Mol. Sci. 2022, 23, 3058. [Google Scholar] [CrossRef]
- Magin, C.M.; Alge, D.L.; Anseth, K.S. Bio-inspired 3D microenvironments: A new dimension in tissue engineering. Biomed. Mater. 2016, 11, 022001. [Google Scholar] [CrossRef] [PubMed]
- Komsa-Penkova, R.; Koynova, R.; Kostov, G.; Tenchov, B. Discrete reduction of type I collagen thermal stability upon oxidation. Biophys. Chem. 2000, 83, 185–195. [Google Scholar] [CrossRef]
- Horzum, U.; Ozdil, B.; Pesen-Okvur, D. Step-by-step quantitative analysis of focal adhesions. MethodsX 2014, 1, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Saalfeld, S. CLAHE (Contrast Limited Adaptive Histogram Equalization). 2009. Available online: http://rsbweb.nih.gov/ij/plugins/clahe/index.html (accessed on 8 January 2023).
- Sage, D.; Neumann, F.R.; Hediger, F.; Gasser, S.M.; Unser, M. Automatic tracking of individual fluorescence particles: Application to the study of chromosome dynamics. IEEE Trans. Image Process. 2005, 14, 1372–1383. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, D.H.; Shah, S.R.; Kim, H.N.; Kshitiz; Kim, P.; Quiñones-Hinojosa, A.; Levchenko, A. Switch-like enhancement of epithelial-mesenchymal transition by YAP through feedback regulation of WT1 and Rho-family GTPases. Nat. Commun. 2019, 10, 2797. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Thampatty, B.P.; Lin, J.S.; Im, H.J. Mechanoregulation of gene expression in fibroblasts. Gene 2007, 391, 1–15. [Google Scholar] [CrossRef]
- Wolfenson, H.; Yang, B.; Sheetz, M.P. Steps in Mechanotransduction Pathways that Control Cell Morphology. Annu. Rev. Physiol. 2019, 81, 585. [Google Scholar] [CrossRef]
- Elango, J.; Hou, C.; Bao, B.; Wang, S.; Maté Sánchez de Val, J.E.; Wenhui, W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function. Polymers 2022, 14, 876. [Google Scholar] [CrossRef]
- Ruggiero, F.; Champliaud, M.F.; Garrone, R.; Aumailley, M. Interactions between Cells and Collagen V Molecules or Single Chains Involve Distinct Mechanisms. Exp. Cell Res. 1994, 210, 215–223. [Google Scholar] [CrossRef]
- Burridge, K.; Feramisco, J.R. Microinjection and localization of a 130K protein in living fibroblasts: A relationship to actin and fibronectin. Cell 1980, 19, 587–595. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Parameters | Col | Col-Oxi | p |
|---|---|---|---|
| Cell Spreading Area (μm2) | 216.0 | 179.6 | p > 0.05 |
| Cell Shape Index (CSI) | 0.25 | 0.33 | p < 0.05 |
| Cell Aspect Ratio (CAR) | 3.33 | 2.61 | p > 0.05 |
| Cell | Col | Col-Oxi | p |
|---|---|---|---|
| Number of FA | 317 | 143 | p < 0.05 |
| Total FA Area (μm2) | 1085 | 406 | p < 0.05 |
| Mean Area per FA (μm2) | 3.66 | 2.84 | p > 0.05 |
| Parameters | Col | Col-Oxi | p |
|---|---|---|---|
| Nuclear TAZ | 229.3 | 178.2 | p < 0.05 |
| Cytosolic TAZ | 1.3 | 12.4 | p < 0.05 |
| Ratio TAZ Nuclei/TAZ Cytosol | 176.2 | 14.4 | p < 0.05 |
| Nuclear Parameters | Col | Col-Oxi | p |
|---|---|---|---|
| Nuclear Area per cell (μM2) | 17.01 | 21.2 | p < 0.05 |
| Nuclear Shape Index (NSI) | 0.81 | 0.86 | p > 0.05 |
| Nuclear Aspect Ratio (NAR) | 1.68 | 1.34 | p > 0.05 |
| Samples | RRMS (nm) | Ea (MPa) |
|---|---|---|
| Collagen Native | 27.95 ± 5.1 | 56.6 ± 8 |
| Col-Oxidized | 5.51 ± 0.8 | 66.8 ± 5 |
| Col Denatured | 1.04 ± 0.6 | 3610 ± 59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komsa-Penkova, R.; Yordanova, A.; Tonchev, P.; Kyurkchiev, S.; Todinova, S.; Strijkova, V.; Iliev, M.; Dimitrov, B.; Altankov, G. Altered Mesenchymal Stem Cells Mechanotransduction from Oxidized Collagen: Morphological and Biophysical Observations. Int. J. Mol. Sci. 2023, 24, 3635. https://doi.org/10.3390/ijms24043635
Komsa-Penkova R, Yordanova A, Tonchev P, Kyurkchiev S, Todinova S, Strijkova V, Iliev M, Dimitrov B, Altankov G. Altered Mesenchymal Stem Cells Mechanotransduction from Oxidized Collagen: Morphological and Biophysical Observations. International Journal of Molecular Sciences. 2023; 24(4):3635. https://doi.org/10.3390/ijms24043635
Chicago/Turabian StyleKomsa-Penkova, Regina, Adelina Yordanova, Pencho Tonchev, Stanimir Kyurkchiev, Svetla Todinova, Velichka Strijkova, Mario Iliev, Borislav Dimitrov, and George Altankov. 2023. "Altered Mesenchymal Stem Cells Mechanotransduction from Oxidized Collagen: Morphological and Biophysical Observations" International Journal of Molecular Sciences 24, no. 4: 3635. https://doi.org/10.3390/ijms24043635
APA StyleKomsa-Penkova, R., Yordanova, A., Tonchev, P., Kyurkchiev, S., Todinova, S., Strijkova, V., Iliev, M., Dimitrov, B., & Altankov, G. (2023). Altered Mesenchymal Stem Cells Mechanotransduction from Oxidized Collagen: Morphological and Biophysical Observations. International Journal of Molecular Sciences, 24(4), 3635. https://doi.org/10.3390/ijms24043635