
Artificial Intelligence-Assisted Meta-Analysis of the Frequency of ACE I/D Polymorphisms in Centenarians and Other Long-Lived Individuals



Abstract
1. Introduction
2. Results
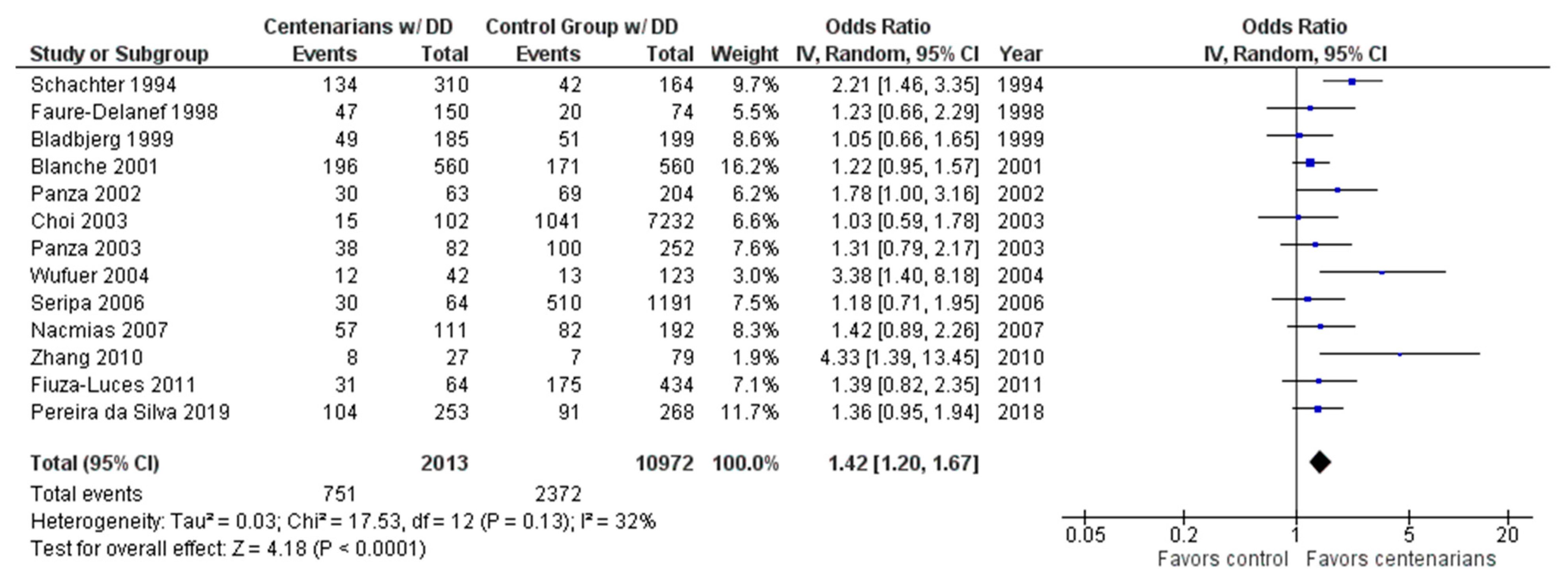
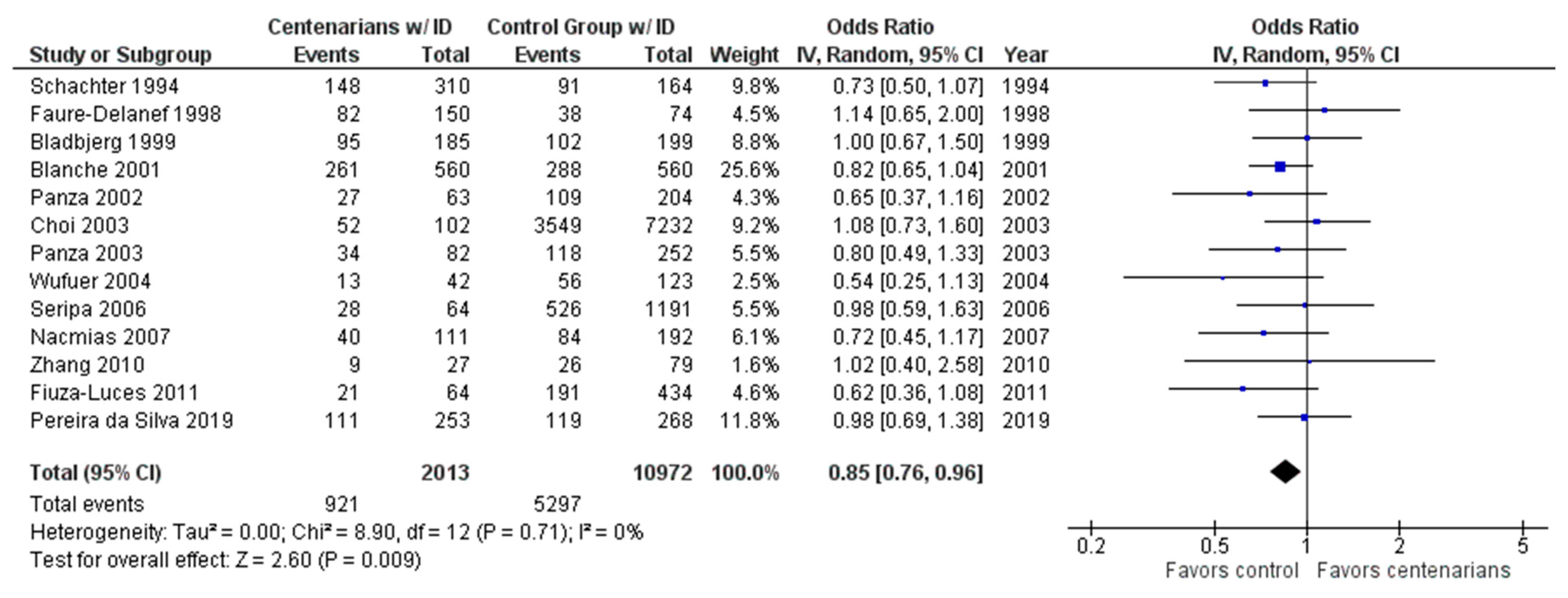
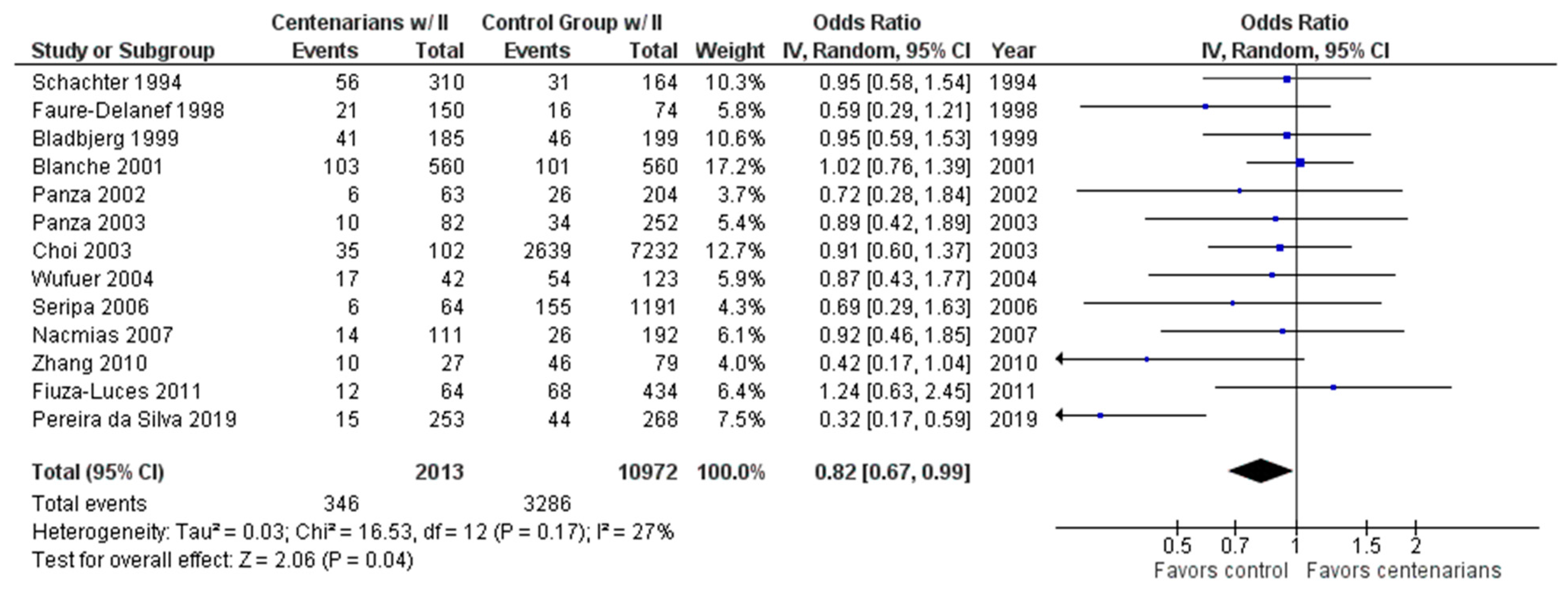
2.1. AI-Assisted Software and Centenarian Results
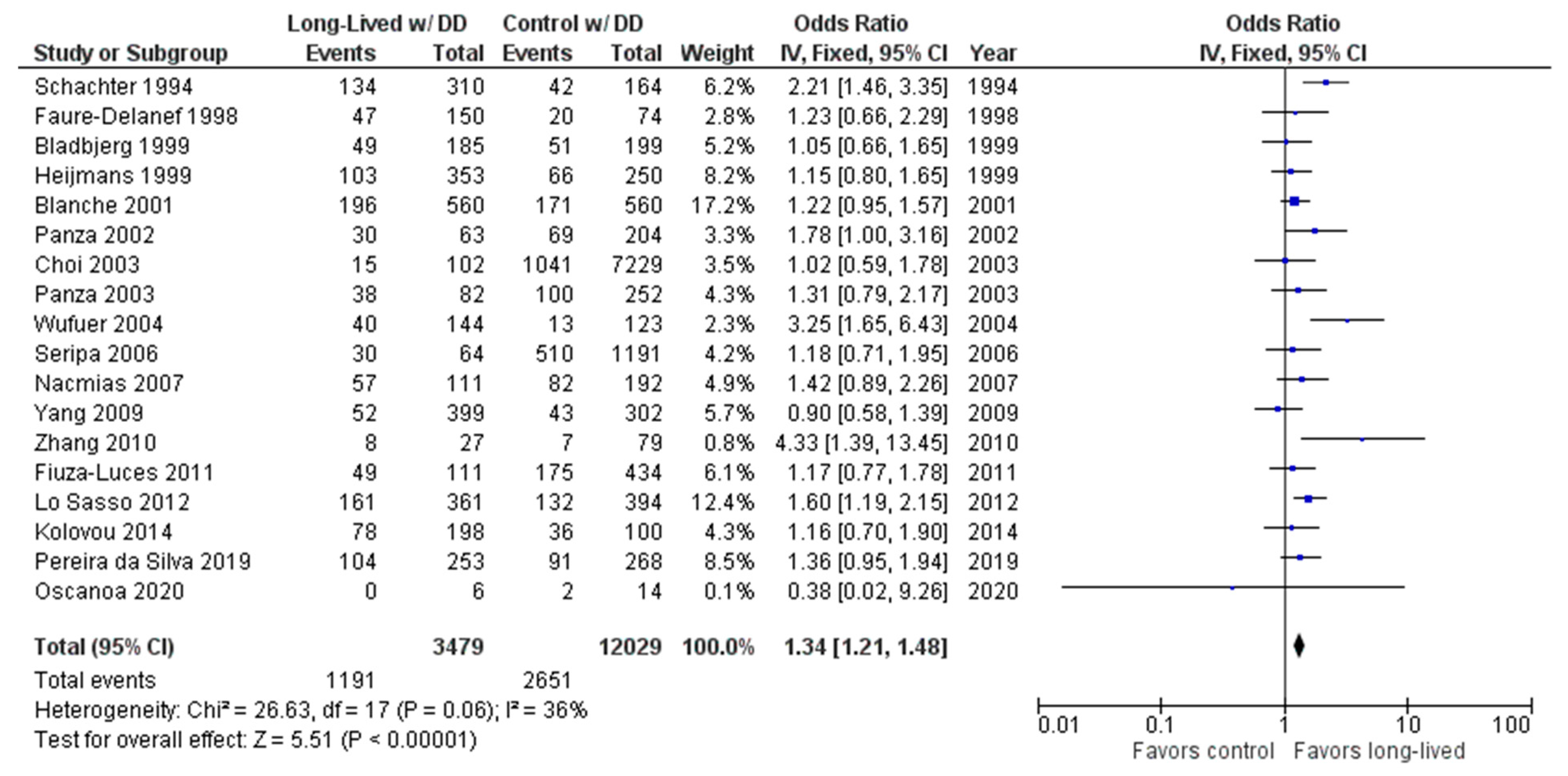
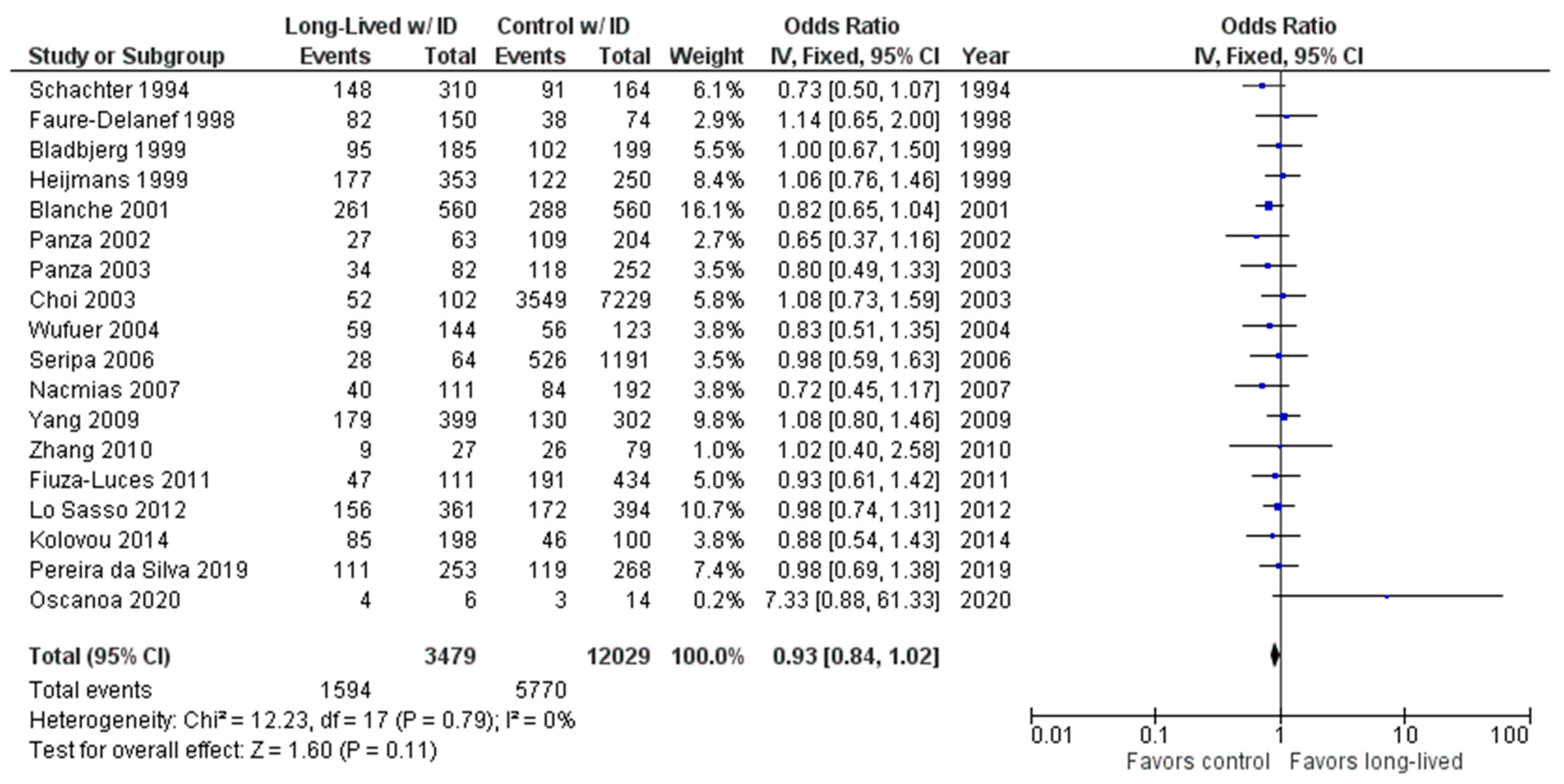
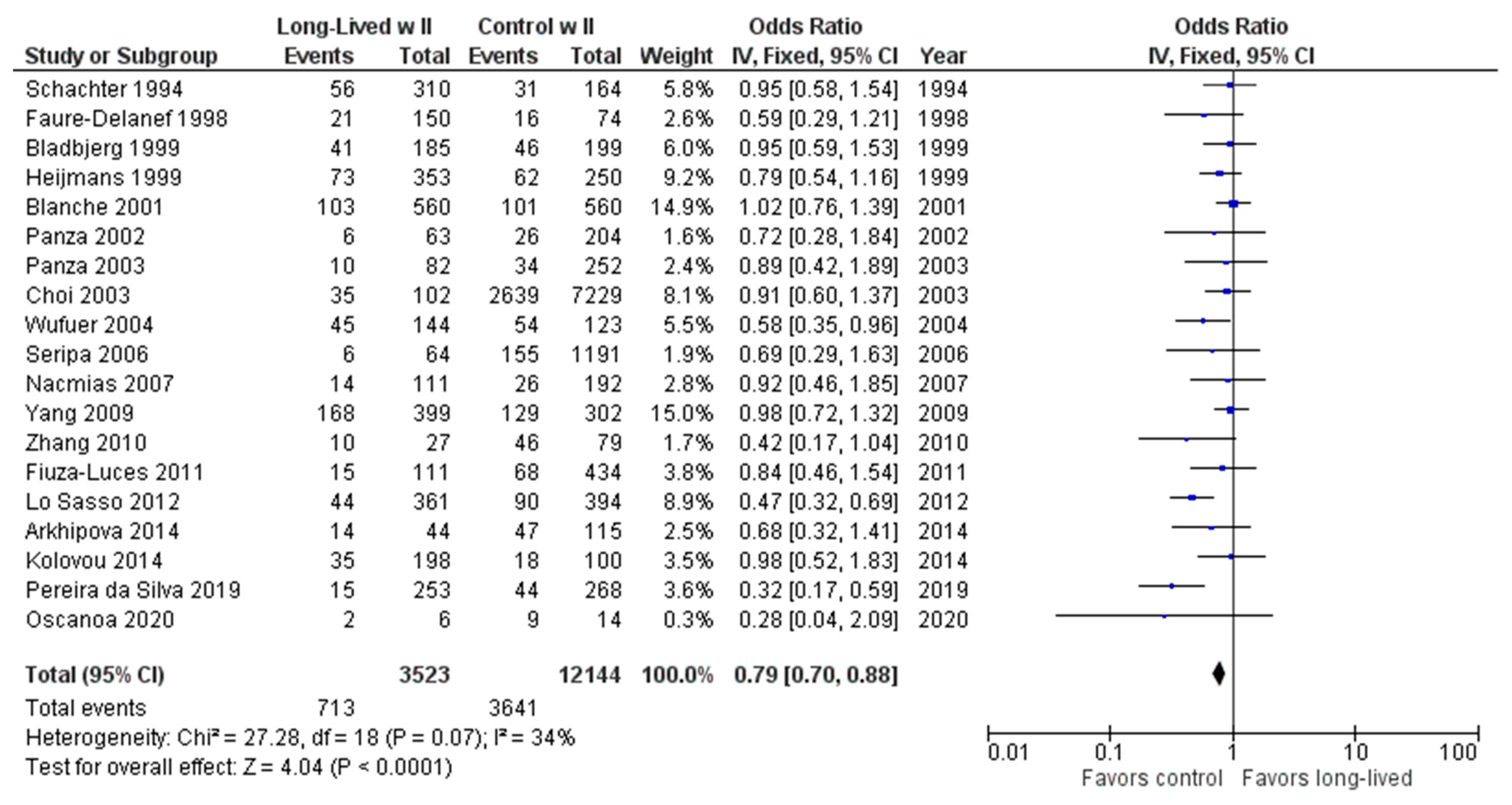
2.2. Long-Lived Results
3. Discussion
4. Materials and Methods
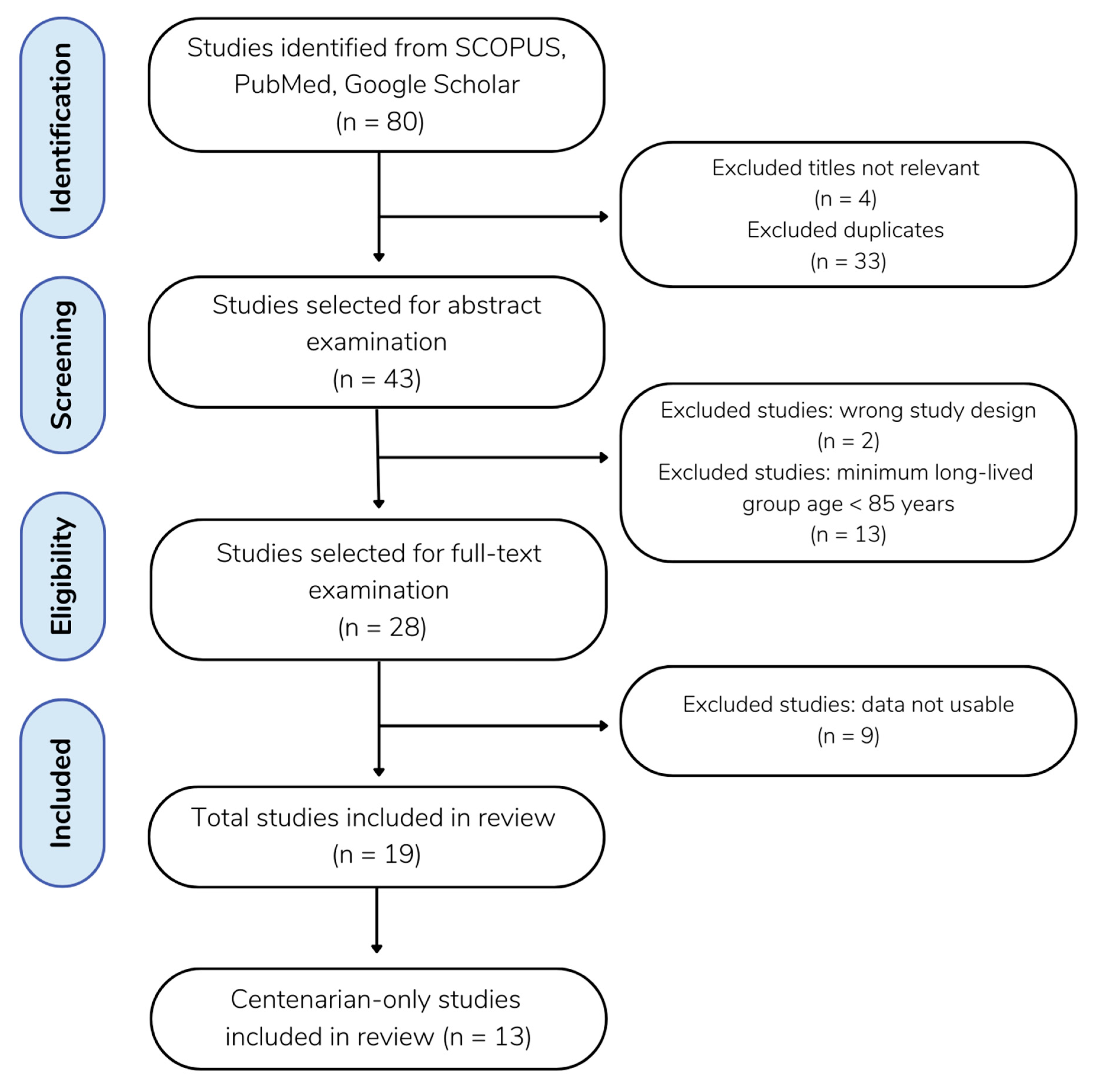
4.1. Nomenclature, PRISMA, and Database Search
4.2. Inclusion Criteria
4.3. Data Extraction
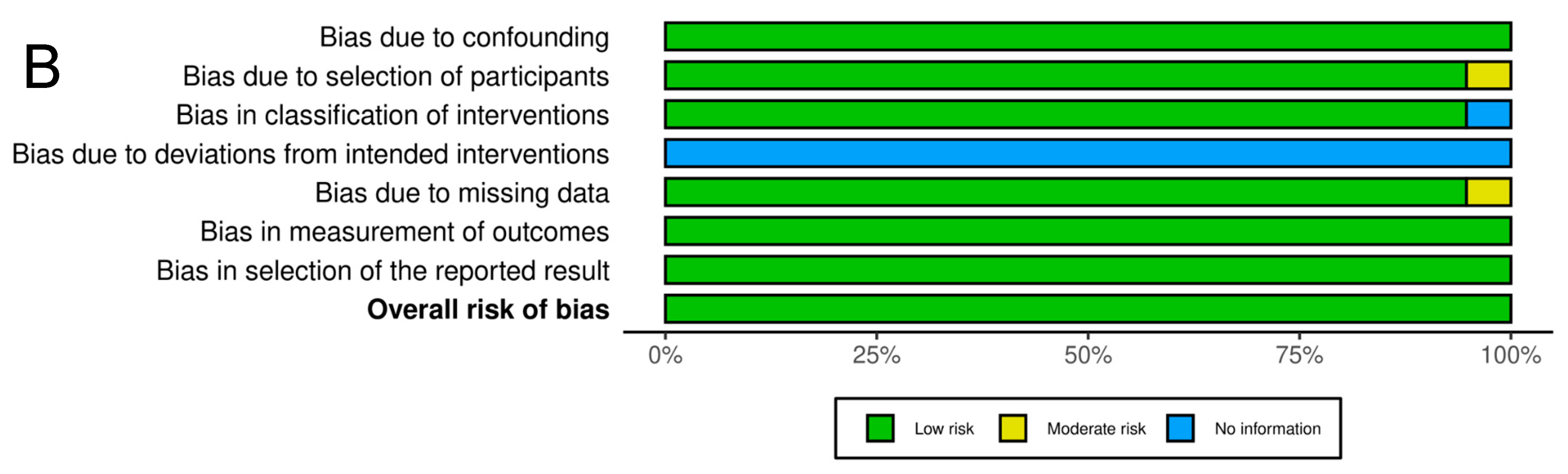
4.4. Data Synthesis, Statistics, and Assessment of the Risk of Bias
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE | angiotensin-converting enzyme |
| I | insertion |
| D | deletion |
| CVD | cardiovascular disease |
| COVID-19 | coronavirus 2019 |
| OR | odds ratio |
| CI | confidence interval |
References
- Montagu, J.D. Length of life in the ancient world: A controlled study. J. R. Soc. Med. 1994, 87, 25–26. [Google Scholar] [CrossRef] [PubMed]
- Batrinos, M.L. The length of life and eugeria in classical Greece. Hormones 2008, 7, 82–83. [Google Scholar] [CrossRef] [PubMed]
- Roser, M.; Ortiz-Ospina, E.; Ritchie, H. Life Expectancy. OurWorldInData.org. 2013. Available online: https://ourworldindata.org/life-expectancy (accessed on 29 November 2022).
- Weinert, B.T.; Timiras, P.S. Invited Review: Theories of aging. J. Appl. Physiol. 2003, 95, 1706–1716. [Google Scholar] [CrossRef]
- Revelas, M.; Thalamuthu, A.; Oldmeadow, C.; Evans, T.-J.; Armstrong, N.J.; Kwok, J.B.; Brodaty, H.; Schofield, P.R.; Scott, R.J.; Sachdev, P.S.; et al. Review and meta-analysis of genetic polymorphisms associated with exceptional human longevity. Mech. Ageing Dev. 2018, 175, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.; Scognamiglio, M.; Fiorito, C.; Benincasa, G.; Napoli, C. Genetic background, epigenetic factors and dietary interventions which influence human longevity. Biogerontology 2019, 20, 605–626. [Google Scholar] [CrossRef] [PubMed]
- Le, D.; Crouch, N.; Villanueva, A.; Ta, P.; Dmitriyev, R.; Tunzi, M.; Murakami, S. Evidence-Based Genetics and Identification of Key Human Alzheimer’s Disease Alleles with Co-morbidities. J. Neurol. Exp. Neurosci. 2020, 6, 20. [Google Scholar] [CrossRef]
- Le, D.; Brown, L.; Malik, K.; Murakami, S. Two Opposing Functions of Angiotensin-Converting Enzyme (ACE) That Links Hypertension, Dementia, and Aging. Int. J. Mol. Sci. 2021, 22, 13178. [Google Scholar] [CrossRef] [PubMed]
- Nia, B.V.; Kang, C.; Tran, M.G.; Lee, D.; Murakami, S. Meta Analysis of Human AlzGene Database: Benefits and Limitations of Using C. elegans for the Study of Alzheimer’s Disease and Co-morbid Conditions. Front. Genet. 2017, 8, 55. [Google Scholar] [CrossRef]
- Murakami, S.; Lacayo, P. Biological and disease hallmarks of Alzheimer’s disease defined by Alzheimer’s disease genes. Front. Aging Neurosci. 2022, 14, 996030. [Google Scholar] [CrossRef]
- Rigat, B.; Hubert, C.; Alhenc-Gelas, F.; Cambien, F.; Corvol, P.; Soubrier, F. An insertion/deletion polymorphism in the angiotensin I-converting enzyme gene accounting for half the variance of serum enzyme levels. J. Clin. Investig. 1990, 86, 1343–1346. [Google Scholar] [CrossRef]
- Tiret, L.; Rigat, B.; Visvikis, S.; Breda, C.; Corvol, P.; Cambien, F.; Soubrier, F. Evidence, from combined segregation and linkage analysis, that a variant of the angiotensin I-converting enzyme (ACE) gene controls plasma ACE levels. Am. J. Hum. Genet. 1992, 51, 197–205. [Google Scholar] [PubMed]
- Tomita, H.; Ina, Y.; Sugiura, Y.; Sato, S.; Kawaguchi, H.; Morishita, M.; Yamamoto, M.; Ueda, R. Polymorphism in the Angiotensin-Converting Enzyme (ACE) Gene and Sarcoidosis. Am. J. Respir. Crit. Care Med. 1997, 156, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Angiotensin-converting enzyme gene insertion/deletion polymorphism and essential hypertension in the Chinese population: A meta-analysis including 21,058 participants. Intern. Med. J. 2012, 42, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, G.; Liu, D.; Fan, X.; Zhu, W.; Liu, X. Angiotensin-Converting Enzyme Insertion/Deletion Polymorphism Contributes to Ischemic Stroke Risk: A Meta-Analysis of 50 Case-Control Studies. PLoS ONE 2012, 7, e46495. [Google Scholar] [CrossRef]
- Zintzaras, E.; Raman, G.; Kitsios, G.; Lau, J. Angiotensin-Converting Enzyme Insertion/Deletion Gene Polymorphic Variant as a Marker of Coronary Artery DiseaseA Meta-analysis. Arch. Intern. Med. 2008, 168, 1077–1089. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Jia, N.; Guo, S.; Chu, S.; Niu, W. Angiotensin-converting enzyme gene deletion allele increases the risk of left ventricular hypertrophy: Evidence from a meta-analysis. Mol. Biol. Rep. 2012, 39, 10063–10075. [Google Scholar] [CrossRef]
- Nie, W.; Zang, Y.; Chen, J.; Liu, T.; Xiao, L.; Xiu, Q. Angiotensin-converting enzyme I/D polymorphism is associated with pneumonia risk: A meta-analysis. J. Renin-Angiotensin-Aldosterone Syst. 2013, 15, 585–592. [Google Scholar] [CrossRef]
- Yamamoto, N.; Ariumi, Y.; Nishida, N.; Yamamoto, R.; Bauer, G.; Gojobori, T.; Shimotohno, K.; Mizokami, M. SARS-CoV-2 infections and COVID-19 mortalities strongly correlate with ACE1 I/D genotype. Gene 2020, 758, 144944. [Google Scholar] [CrossRef]
- Schächter, F.; Faure-Delanef, L.; Guénot, F.; Rouger, H.; Froguel, P.; Lesueur-Ginot, L.; Cohen, D. Genetic associations with human longevity at the APOE and ACE loci. Nat. Genet. 1994, 6, 29–32. [Google Scholar] [CrossRef]
- Garatachea, N.; Marín, P.J.; Lucia, A. The ACE DD genotype and D-allele are associated with exceptional longevity: A meta-analysis. Ageing Res. Rev. 2013, 12, 1079–1087. [Google Scholar] [CrossRef]
- Choi, Y.-H.; Kim, J.-H.; Kim, D.K.; Lee, M.S.; Kim, C.H.; Park, S.C. Distributions of ACE and APOE Polymorphisms and Their Relations with Dementia Status in Korean Centenarians. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2003, 58, M227–M231. [Google Scholar] [CrossRef] [PubMed]
- Oscanoa, T.J.; Cieza, E.C.; Lizaraso-Soto, F.A.; Guevara, M.L.; Fujita, R.M.; Romero-Ortuño, R. Angiotensin-Converting Enzyme (ACE) genetic variation and longevity in Peruvian older people: A cross-sectional study. Ann. Hum. Biol. 2020, 47, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Augustin, C.; Bethel, A.; Gill, D.; Anzaroot, S.; Brun, J.; DeWilde, B.; Minnich, R.; Garside, R.; Masuda, Y.; et al. Using machine learning to advance synthesis and use of conservation and environmental evidence. Conserv. Biol. 2018, 32, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Faure-Delanef, L.; Baudin, B.; Bénézteau-Burnat, B.; Beaudoin, J.-C.; Giboudeau, J.; Cohen, D. Plasma concentration, kinetic constants, and gene polymorphism of angiotensin I-converting enzyme in centenarians. Clin. Chem. 1998, 44, 2083–2087. [Google Scholar] [CrossRef] [PubMed]
- Bladbjerg, E.M.; Andersen-Ranberg, K.; de Maat, M.P.; Kristensen, S.R.; Jeune, B.; Gram, J.; Jespersen, J. Longevity is independent of common variations in genes associated with cardiovascular risk. Thromb. Haemost. 1999, 82, 1100–1105. [Google Scholar] [PubMed]
- Blanché, H.; Cabanne, L.; Sahbatou, M.; Thomas, G. A study of French centenarians: Are ACE and APOE associated with longevity? Comptes Rendus L’académie Sci. Ser. III-Sci. De La Vie 2001, 324, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Solfrizzi, V.; D‘Introno, A.; Capurso, C.; Basile, A.M.; Colacicco, A.M.; Sabba, M.; Noya, R.; Capurso, A. Apolipo-protein and angiotensin converting enzyme genes: Regional differences and extreme longevity in Europe. Arch. Gerontol. Geriatr. 2002, 8, 247–251. [Google Scholar] [CrossRef]
- Panza, F.; Solfrizzi, V.; D’Introno, A.; Colacicco, A.M.; Capurso, C.; Kehoe, P.G.; Capurso, A. Angiotensin I converting enzyme (ACE) gene polymorphism in centenarians: Different allele frequencies between the North and South of Europe. Exp. Gerontol. 2003, 38, 1015–1020. [Google Scholar] [CrossRef]
- Wufuer, M.; Fang, M.-W.; Cheng, Z.-H.; Qiu, C.-C. Polymorphism of angiotensin converting enzyme gene and natural longevity in the Xinjiang Uygur people: An association study. Zhonghua Yi Xue Za Zhi 2004, 84, 1603–1606. (In Chinese) [Google Scholar]
- Seripa, D.; Franceschi, M.; Matera, M.G.; Panza, F.; Kehoe, P.; Gravina, C.; Orsitto, G.; Solfrizzi, V.; Di Minno, G.; Dallapiccola, B.; et al. Sex Differences in the Association of Apolipoprotein E and Angiotensin-Converting Enzyme Gene Polymorphisms With Healthy Aging and Longevity: A Population-Based Study From Southern Italy. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2006, 61, 918–923. [Google Scholar] [CrossRef]
- Nacmias, B.; Bagnoli, S.; Tedde, A.; Cellini, E.; Bessi, V.; Guarnieri, B.; Ortensi, L.; Piacentini, S.; Bracco, L.; Sorbi, S. Angiotensin converting enzyme insertion/deletion polymorphism in sporadic and familial Alzheimer’s disease and longevity. Arch. Gerontol. Geriatr. 2007, 45, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-F.; Ling, J.-H.; Wang, A.-J. Relationship between shen-deficiency syndrome and polymorphism of angiotensin converting enzyme gene in longevous elders in Bama area of Guangxi municipality. Zhongguo Zhong Xi Yi Jie He Za Zhi Zhongguo Zhongxiyi Jiehe Zazhi 2010, 30, 259–263. (In Chinese) [Google Scholar] [PubMed]
- Fiuza-Luces, C.; Ruiz, J.R.; Rodríguez-Romo, G.; Santiago, C.; Gómez-Gallego, F.; Cano-Nieto, A.; Garatachea, N.; Rodríguez-Moreno, I.; Morán, M.; Lucia, A. Is the ACE I/D polymorphism associated with extreme longevity? A study on a Spanish cohort. J. Renin-Angiotensin-Aldosterone Syst. 2011, 12, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Pereira da Silva, A.; Matos, A.; Aguiar, L.; Ramos-Marques, N.; Ribeiro, R.; Gil, Â.; Gorjão-Clara, J.; Bicho, M. Hypertension and longevity: Role of genetic polymorphisms in renin-angiotensin-aldosterone system and endothelial nitric oxide synthase. Mol. Cell. Biochem. 2018, 455, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Paolisso, G.; Tagliamonte, M.R.; De Lucia, D.; Palmieri, F.; Manzella, D.; Rinaldi, C.; Bossone, A.; Colaizzo, D.; Margaglione, M.; Varricchio, M. ACE Gene Polymorphism and Insulin Action in Older Subjects and Healthy Centenarians. J. Am. Geriatr. Soc. 2001, 49, 610–614. [Google Scholar] [CrossRef]
- Arkhipova, N.; Popova, E.; Grigorieva, L. The angiotension converting enzyme (ACE) gene I/D polymorphism in different ethnic groups of geriatric age living in the Far North. Health 2014, 6, 426–431. [Google Scholar] [CrossRef]
- Kolovou, G.; Kolovou, V.; Vasiliadis, I.; Giannakopoulou, V.; Mihas, C.; Bilianou, H.; Kollia, A.; Papadopoulou, E.; Marvaki, A.; Goumas, G.; et al. The Frequency of 4 Common Gene Polymorphisms in Nonagenarians, Centenarians, and Average Life Span Individuals. Angiology 2014, 65, 210–215. [Google Scholar] [CrossRef]
- Lo Sasso, B.; Bellia, C.; Tomaiuolo, R.; Zarrilli, F.; Scorza, M.; Caruso, A.; Agnello, L.; Bazza, F.; Carru, C.; Zinellu, A.; et al. Polimorfismo I/D del gene per l’enzima di conversione dell’angiotensina (ACE): Gene della longevità o fattore di rischio nella patologia ipertensiva? Biochim. Clin. 2013, 37, 461–464. [Google Scholar]
- Yang, J.-K.; Gong, Y.-Y.; Xie, L.; Lian, S.-G.; Yang, J.; Xu, L.-Y.; Gao, S.-J.; Zhang, Y.-P. Lack of genetic association between the angiotensin-converting enzyme gene insertion/deletion polymorphism and longevity in a Han Chinese population. J. Renin-Angiotensin-Aldosterone Syst. 2009, 10, 115–118. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Westendorp, R.G.; Knook, D.L.; Kluft, C.; Slagboom, P. Angiotensin I–converting enzyme and plasminogen activator inhibitor-1 gene variants: Risk of mortality and fatal cardiovascular disease in an elderly population-based cohort. J. Am. Coll. Cardiol. 1999, 34, 1176–1183. [Google Scholar] [CrossRef]
- Willcox, D.C.; Willcox, B.J.; Poon, L.W. Centenarian Studies: Important Contributors to Our Understanding of the Aging Process and Longevity. Curr. Gerontol. Geriatr. Res. 2010, 2010, 484529. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Igarashi, A.; Kamata, M.; Nakagawa, H. Angiotensin-converting Enzyme Degrades Alzheimer Amyloid β-Peptide (Aβ); Retards Aβ Aggregation, Deposition, Fibril Formation; and Inhibits Cytotoxicity. J. Biol. Chem. 2001, 276, 47863–47868. [Google Scholar] [CrossRef]
- Kölsch, H.; Jessen, F.; Freymann, N.; Kreis, M.; Hentschel, F.; Maier, W.; Heun, R. ACE I/D polymorphism is a risk factor of Alzheimer’s disease but not of vascular dementia. Neurosci. Lett. 2005, 377, 37–39. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.J.; Cortina-Borja, M.; Warden, D.R.; Smith, A.D.; Sleegers, K.; Prince, J.A.; van Duijn, C.M.; Kehoe, P.G. Large Meta-Analysis Establishes the ACE Insertion-Deletion Polymorphism as a Marker of Alzheimer’s Disease. Am. J. Epidemiol. 2005, 162, 305–317. [Google Scholar] [CrossRef]
- Murakami, S.; Cabana, K.; Anderson, D. Chapter 25—Current advances in the study of oxidative stress and age-related memory impairment in C. elegans. In Molecular Aspects of Oxidative Stress on Cell Signaling in Vertebrates and In-Vertebrates; Farooqui, T., Farooqui, A., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 347–360. [Google Scholar]
- Murakami, S. Chapter 12—Age-dependent modulation of learning and memory in C. elegans. In Handbook of Behavioral Neuroscience; Part of Invertebrate Learning and Memory; Menzel, R., Benjamin, P.R., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2013; pp. 140–150. [Google Scholar]
- Machino, K.; Link, C.D.; Wang, S.; Murakami, H.; Murakami, S. A semi-automated motion-tracking analysis of locomotion speed in the C. elegans transgenics overexpressing beta-amyloid in neurons. Front. Genet. 2014, 5, 202. [Google Scholar] [CrossRef] [PubMed]
- Costerousse, O.; Allegrini, J.; Lopez, M.; Alhenc-Gelas, F. Angiotensin I-converting enzyme in human circulating mononuclear cells: Genetic polymorphism of expression in T-lymphocytes. Biochem. J. 1993, 290, 33–40. [Google Scholar] [CrossRef]
- Dato, S.; Crocco, P.; De Rango, F.; Iannone, F.; Maletta, R.; Bruni, A.C.; Saiardi, A.; Rose, G.; Passarino, G. IP6K3 and IPMK variations in LOAD and longevity: Evidence for a multifaceted signaling network at the crossroad between neurodegeneration and survival. Mech. Ageing Dev. 2021, 195, 111439. [Google Scholar] [CrossRef]
- Balmorez, T.; Sakazaki, A.; Murakami, S. Genetic Networks of Alzheimer’s Disease, Aging and Longevity in Humans. medRxiv 2023. [Google Scholar] [CrossRef]
- Egan, B.M.; Scharf, A.; Pohl, F.; Kornfeld, K. Control of aging by the renin–angiotensin system: A review of C. elegans, Drosophila, and mammals. Front. Pharmacol. 2022, 13, 938650. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Antos, A.; Kwong, M.; Balmorez, T.; Villanueva, A.; Murakami, S. Unusually High Risks of COVID-19 Mortality with Age-Related Comorbidities: An Adjusted Meta-Analysis Method to Improve the Risk Assessment of Mortality Using the Comorbid Mortality Data. Infect. Dis. Rep. 2021, 13, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]
- Sterne, J.A.C.; Hernán, M.A.; Reeves, B.C.; Savović, J.; Berkman, N.D.; Viswanathan, M.; Henry, D.; Altman, D.G.; Ansari, M.T.; Boutron, I.; et al. ROBINS-I: A tool for assessing risk of bias in non-randomised studies of interventions. BMJ 2016, 355, i4919. [Google Scholar] [CrossRef]
- McGuinness, L.A.; Higgins, J.P.T. Risk-of-bias VISualization (robvis): An R package and Shiny web app for visualizing risk-of-bias assessments. Res. Synth. Methods 2021, 12, 55–61. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Centenarian | Control | Ethnicity | Ref | |||
|---|---|---|---|---|---|---|---|
| # of Subjects | Age (Years) Mean ± SD | # of Subjects | Age (Years) Range | ||||
| Schachter et al. | 1994 | 310 | 100.71 ± 0.13 | 164 | 20–70 | Caucasian | [20] |
| Faure-Delanef et al. | 1998 | 150 | 100.6 | 74 | 20–70 | Caucasian | [25] |
| Bladbjerg et al. | 1999 | 185 | >100 | 199 | 20–64 | Caucasian | [26] |
| Blanche et al. | 2001 | 560 | 103.1 | 560 | 18–70 | Caucasian | [27] |
| Panza et al. | 2002 | 63 | 100.3 ± 1.9 | 204 | 19–80 | Caucasian | [28] |
| Choi et al. | 2003 | 102 | 102.4 ± 2.6 | 7229 | 20–85 | Korean | [22] |
| Panza et al. | 2003 | 82 | 100 ± 2 | 252 | 19–70 | Caucasian | [29] |
| Wufuer et al. | 2004 | 42 | 103.0 ± 3.0 | 53 | 65–70 | Uyghur | [30] |
| Seripa et al. | 2006 | 64 | 100.28 ± 1.86 | 1191 | 22–59 | Caucasian | [31] |
| Nacmias et al. | 2007 | 111 | 102.4 ± 2.6 | 192 | 84 ± 18.2 | Caucasian | [32] |
| Zhang et al. | 2010 | 27 | >100 | 79 | 60–70 | Han Chinese | [33] |
| Fiuza-Luces et al. | 2011 | 65 | 101.7 ± 1.8 | 283 | 21.2 ± 2.0 | Caucasian | [34] |
| Pereira da Silva et al. | 2018 | 253 | 100.26 ± 1.98 | 268 | 67.51 ± 3.25 | Caucasian | [35] |
| Year | Long-Lived | Control | Ethnicity | Ref | |||
|---|---|---|---|---|---|---|---|
| # of Subjects | Age (Years) Mean ± SD | # of Subjects | Age (Years) Range/Mean ± SD | ||||
| Schachter et al. | 1994 | 310 | 100.71 ± 0.13 | 164 | 20–70 | Caucasian | [20] |
| Faure-Delanef et al. | 1998 | 150 | 100.6 | 74 | 20–70 | Caucasian | [25] |
| Bladbjerg et al. | 1999 | 185 | >100 | 199 | 20–64 | Caucasian | [26] |
| Heijmans et al. | 1999 | 353 | 85–100 | 250 | 18–40 | Caucasian | [41] |
| Blanche et al. | 2001 | 560 | 103.1 | 560 | 18–70 | Caucasian | [27] |
| Panza et al. | 2002 | 63 | 100.3 ± 1.9 | 204 | 19–80 | Caucasian | [28] |
| Choi et al. | 2003 | 102 | 102.4 ± 2.6 | 7229 | 20–85 | Korean | [22] |
| Panza et al. | 2003 | 82 | 100 ± 2 | 252 | 19–70 | Caucasian | [29] |
| Wufuer et al. | 2004 | 42 | 103.0 ± 3.0 | 53 | 65–70 | Uyghur | [30] |
| Seripa et al. | 2006 | 64 | 100.28 ± 1.86 | 1191 | 22–59 | Caucasian | [31] |
| Nacmias et al. | 2007 | 111 | 102.4 ± 2.6 | 192 | 84 ± 18.2 | Caucasian | [32] |
| Yang et al. | 2009 | 399 | >90 | 302 | <60 | Han Chinese | [40] |
| Zhang et al. | 2010 | 27 | >100 | 79 | 60–70 | Han Chinese | [33] |
| Fiuza-Luces et al. | 2011 | 65 | 101.7 ± 1.8 | 283 | 21.2 ± 2.0 | Caucasian | [34] |
| Lo Sasso et al. | 2012 | 361 | 97 | 394 | 39 | Caucasian | [39] |
| Arkhipova et al. | 2014 | 44 | >90 | 115 | 60–74 | Russian, Yakut | [37] |
| Kolovou et al. | 2014 | 198 | ≥90 | 105 | 18–79 | Caucasian | [38] |
| Pereira da Silva et al. | 2018 | 253 | 100.26 ± 1.98 | 268 | 67.51 ± 3.25 | Caucasian | [35] |
| Oscanoa et al. | 2020 | 6 | >85 | 14 | <65 | Peruvian | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Murakami, S. Artificial Intelligence-Assisted Meta-Analysis of the Frequency of ACE I/D Polymorphisms in Centenarians and Other Long-Lived Individuals. Int. J. Mol. Sci. 2023, 24, 3411. https://doi.org/10.3390/ijms24043411
Li L, Murakami S. Artificial Intelligence-Assisted Meta-Analysis of the Frequency of ACE I/D Polymorphisms in Centenarians and Other Long-Lived Individuals. International Journal of Molecular Sciences. 2023; 24(4):3411. https://doi.org/10.3390/ijms24043411
Chicago/Turabian StyleLi, Lingxuan, and Shin Murakami. 2023. "Artificial Intelligence-Assisted Meta-Analysis of the Frequency of ACE I/D Polymorphisms in Centenarians and Other Long-Lived Individuals" International Journal of Molecular Sciences 24, no. 4: 3411. https://doi.org/10.3390/ijms24043411
APA StyleLi, L., & Murakami, S. (2023). Artificial Intelligence-Assisted Meta-Analysis of the Frequency of ACE I/D Polymorphisms in Centenarians and Other Long-Lived Individuals. International Journal of Molecular Sciences, 24(4), 3411. https://doi.org/10.3390/ijms24043411