
Adverse Human Health Effects of Chromium by Exposure Route: A Comprehensive Review Based on Toxicogenomic Approach


 ,
,
Abstract
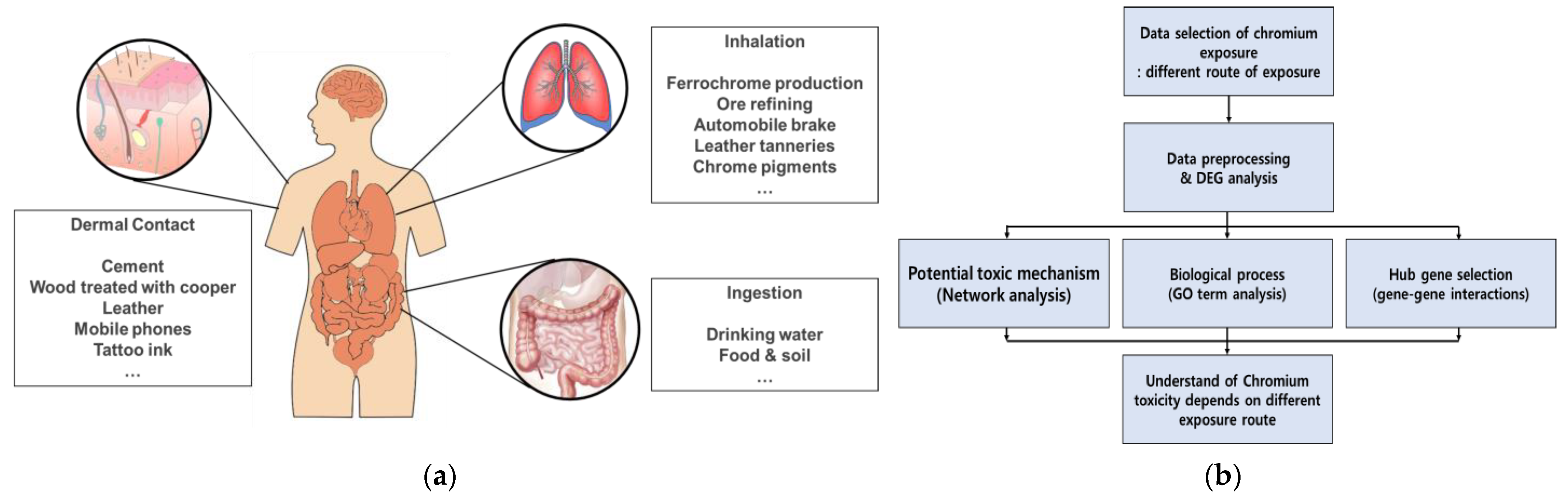
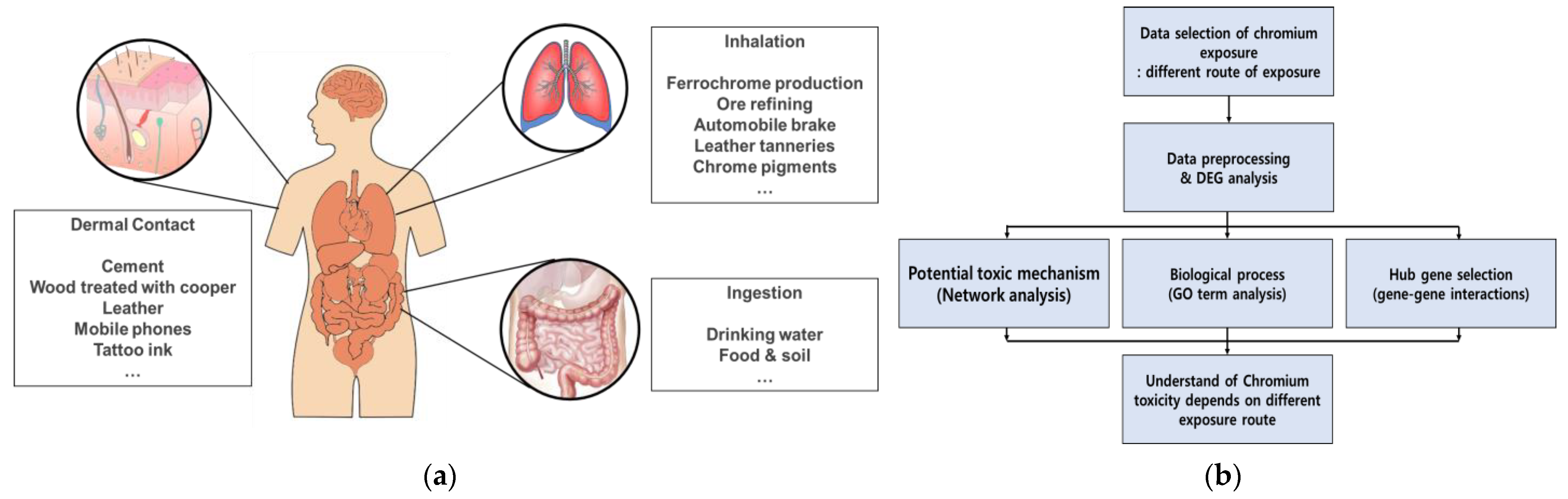
:1. Introduction
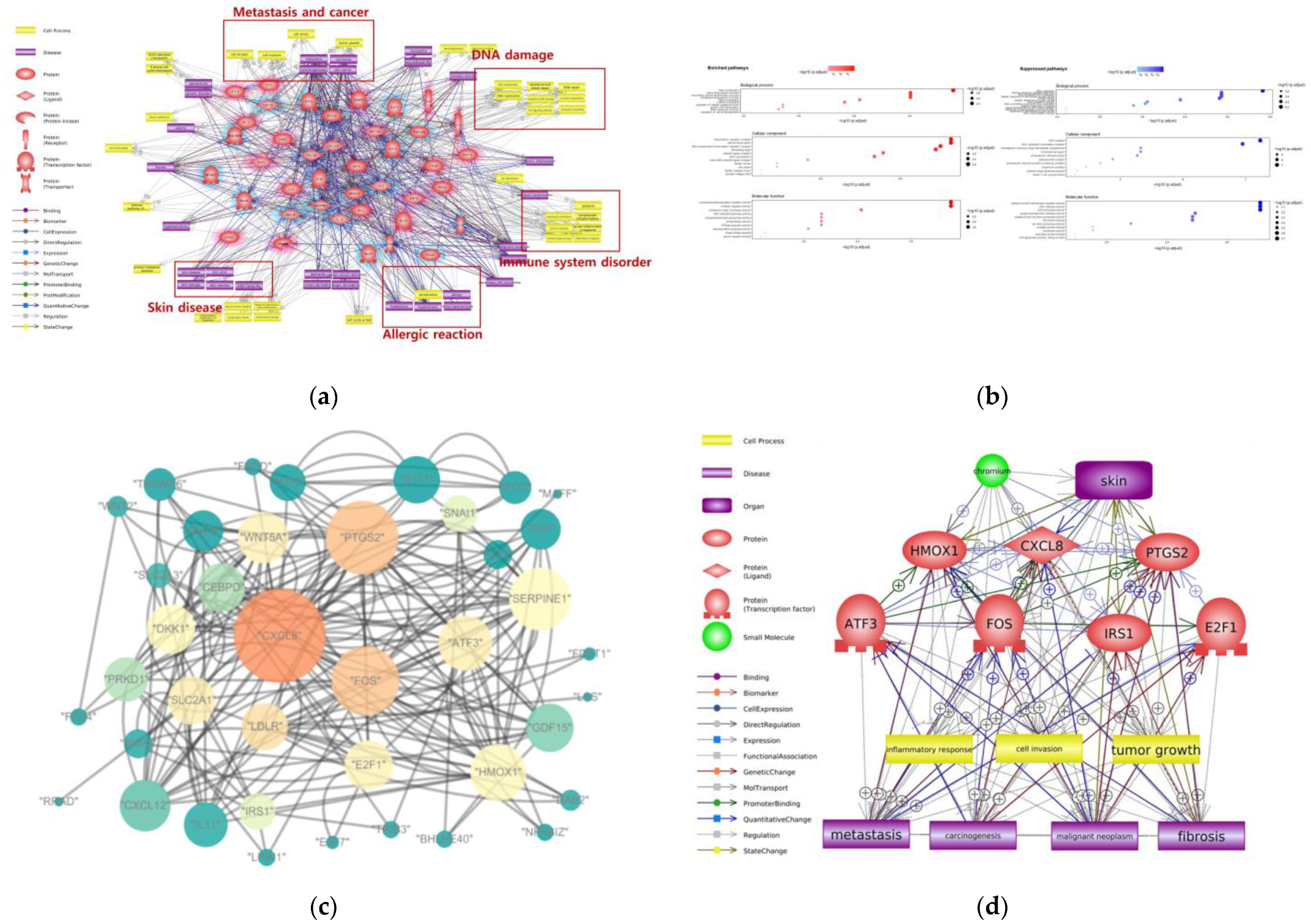
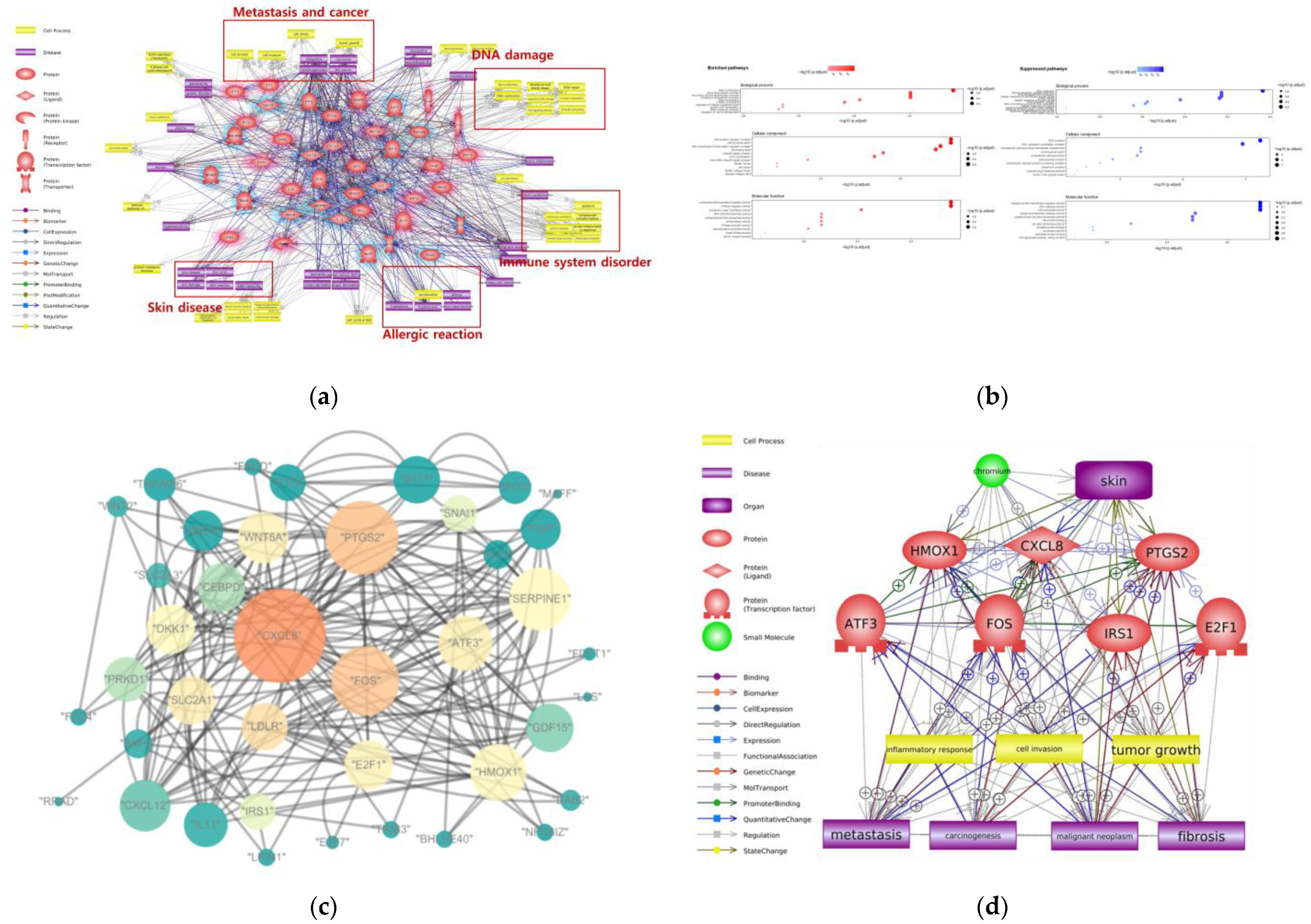
2. Adverse Effects of Cr on Skin
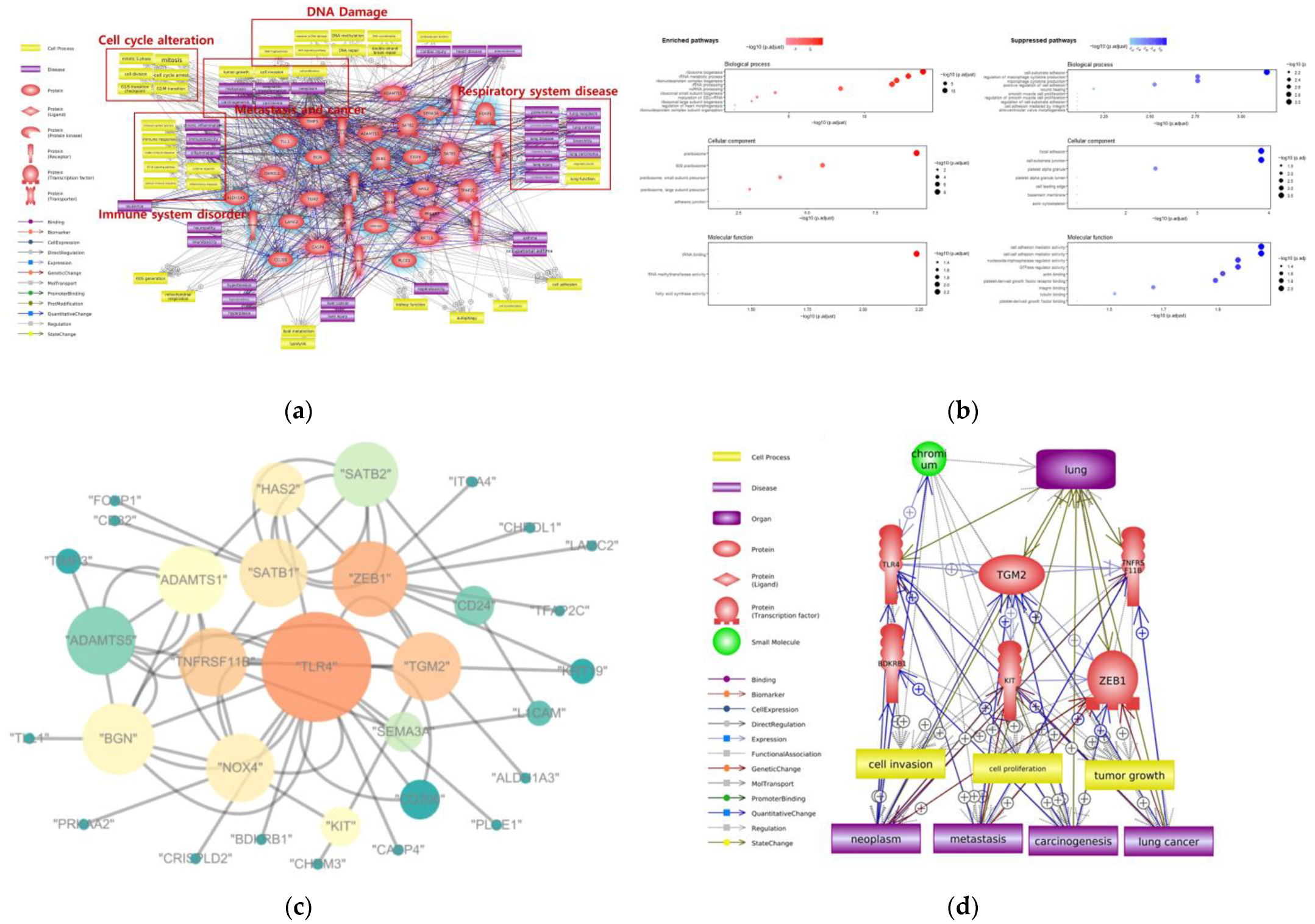
3. Adverse Effects of Cr on Respiratory System
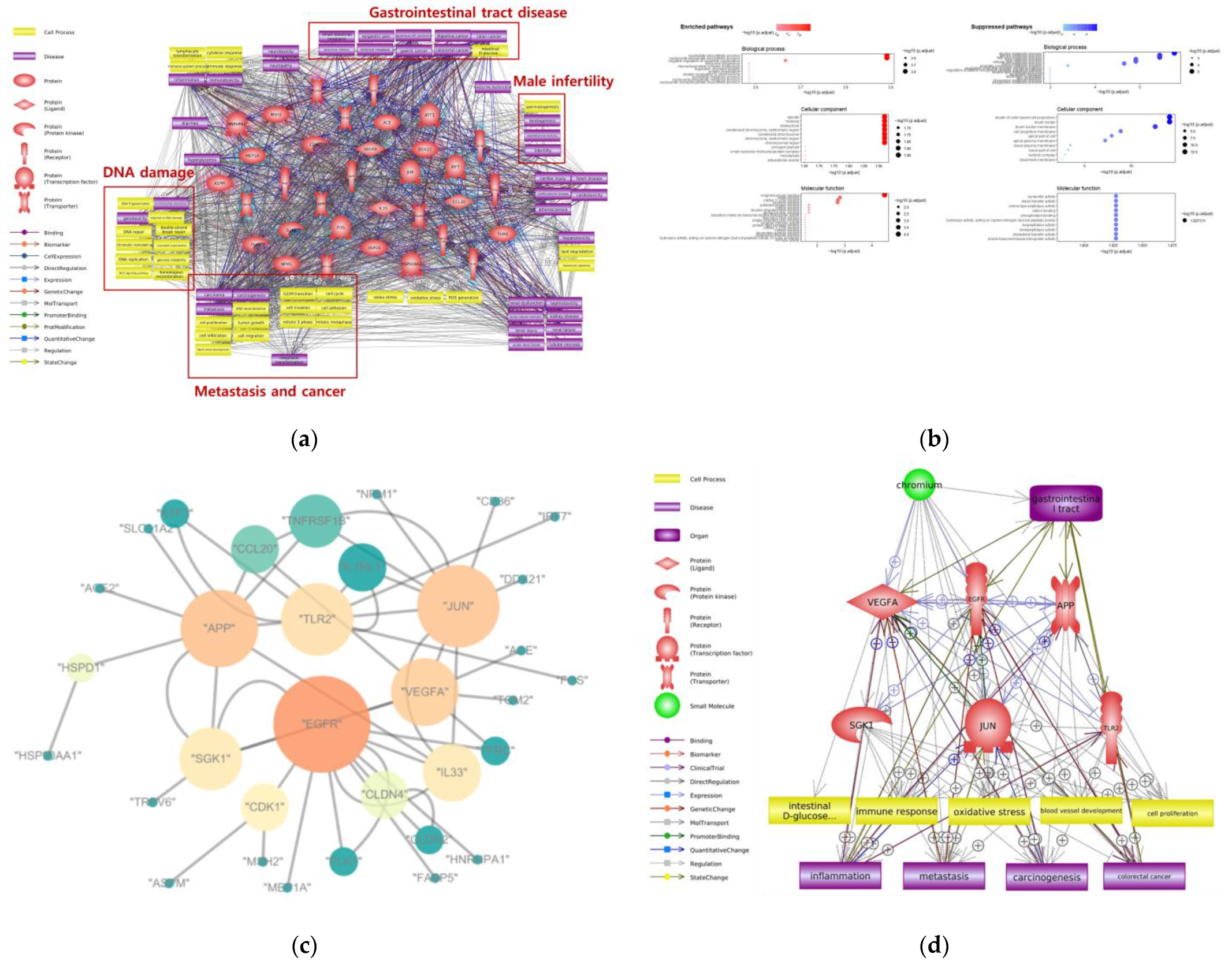
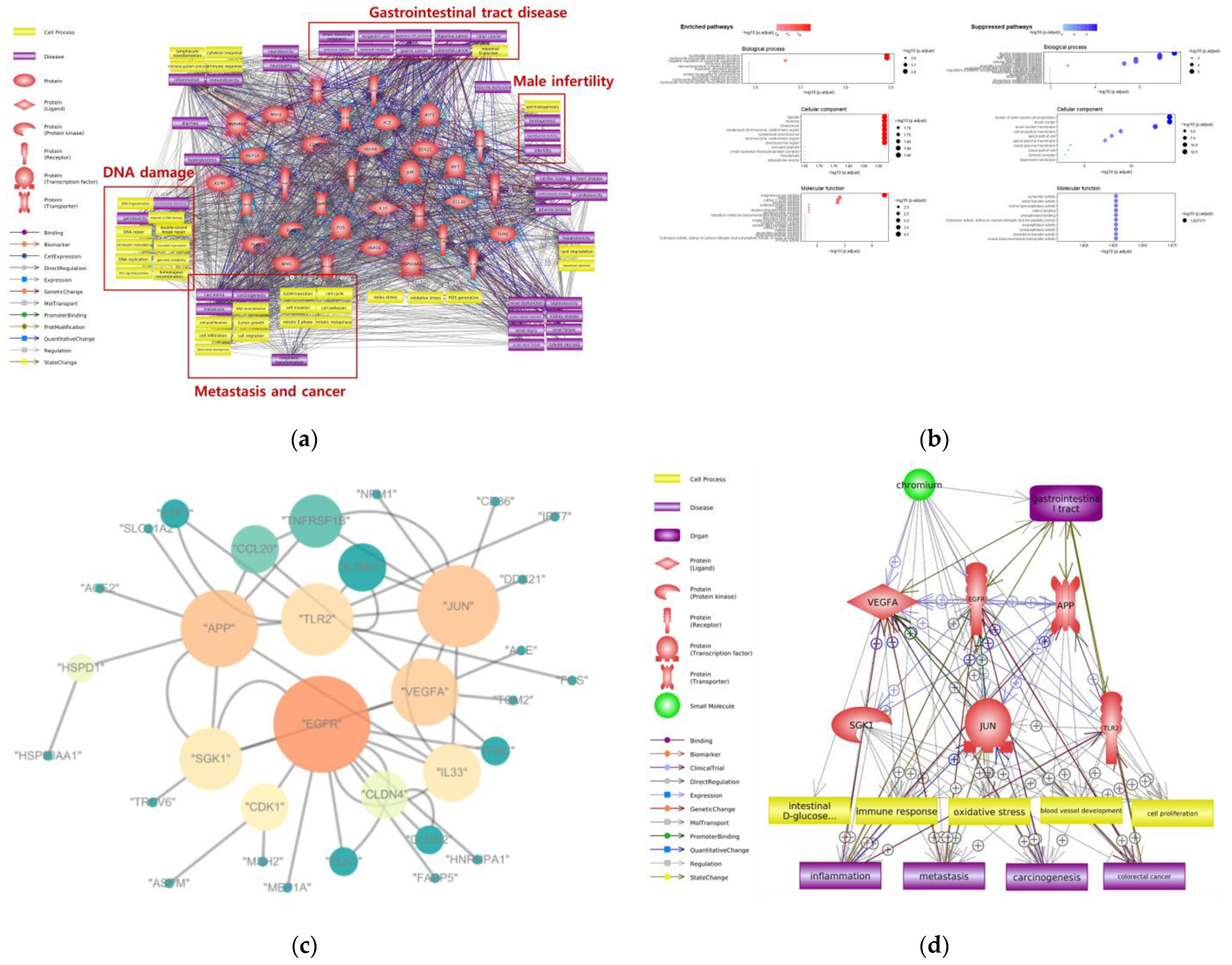
4. Adverse Effects of Cr on Gastrointestinal Tract
5. Adverse Human Health Effects of Cr Depend on the Exposure Route
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pereira, S.C.; Oliveira, P.F.; Oliveira, S.R.; Pereira, M.L.; Alves, M.G. Impact of Environmental and Lifestyle Use of Chromium on Male Fertility: Focus on Antioxidant Activity and Oxidative Stress. Antioxidants 2021, 10, 1365. [Google Scholar] [CrossRef] [PubMed]
- Safonov, A.; Popova, N.; Andrushenko, N.; Boldyrev, K.; Yushin, N.; Zinicovscaia, I. Investigation of materials for reactive permeable barrier in removing cadmium and chromium(VI) from aquifer near a solid domestic waste landfill. Environ. Sci. Pollut. Res. Int. 2021, 28, 4645–4659. [Google Scholar] [CrossRef]
- Forte, I.M.; Indovina, P.; Costa, A.; Iannuzzi, C.A.; Costanzo, L.; Marfella, A.; Montagnaro, S.; Botti, G.; Bucci, E.; Giordano, A. Blood screening for heavy metals and organic pollutants in cancer patients exposed to toxic waste in southern Italy: A pilot study. J. Cell. Physiol. 2020, 235, 5213–5222. [Google Scholar] [CrossRef] [PubMed]
- Zaeemzadeh, N.; Jahanian Sadatmahalleh, S.; Ziaei, S.; Kazemnejad, A.; Movahedinejad, M.; Mottaghi, A.; Mohamadzadeh, N. Comparison of dietary micronutrient intake in PCOS patients with and without metabolic syndrome. J. Ovarian Res. 2021, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Vaiopoulou, E.; Gikas, P. Regulations for chromium emissions to the aquatic environment in Europe and elsewhere. Chemosphere 2020, 254, 126876. [Google Scholar] [CrossRef]
- Batyrova, G.; Kononets, V.; Amanzholkyzy, A.; Tlegenova, Z.; Umarova, G. Chromium as a Risk Factor for Breast Cancer: A Meta-Analysis. Asian Pac. J. Cancer Prev. 2022, 23, 3993–4003. [Google Scholar] [CrossRef]
- Hossini, H.; Shafie, B.; Niri, A.D.; Nazari, M.; Esfahlan, A.J.; Ahmadpour, M.; Nazmara, Z.; Ahmadimanesh, M.; Makhdoumi, P.; Mirzaei, N.; et al. A comprehensive review on human health effects of chromium: Insights on induced toxicity. Environ. Sci. Pollut. Res. Int. 2022, 29, 70686–70705. [Google Scholar] [CrossRef]
- Fontaine, M.; Clement, Y.; Blanc, N.; Demesmay, C. Hexavalent chromium release from leather over time natural ageing vs accelerated ageing according to a multivariate approach. J. Hazard. Mater. 2019, 368, 811–818. [Google Scholar] [CrossRef]
- Tsuchiyama, T.; Tazaki, A.; Al Hossain, M.A.; Yajima, I.; Ahsan, N.; Akhand, A.A.; Hashimoto, K.; Ohgami, N.; Kato, M. Increased levels of renal damage biomarkers caused by excess exposure to trivalent chromium in workers in tanneries. Environ. Res. 2020, 188, 109770. [Google Scholar] [CrossRef]
- Kozlowska, L.; Gromadzinska, J.; Wasowicz, W. Health risk in transport workers. Part II. Dietary compounds as modulators of occupational exposure to chemicals. Int. J. Occup. Med. Environ. Health 2019, 32, 441–464. [Google Scholar] [CrossRef]
- Santonen, T.; Porras, S.P.; Bocca, B.; Bousoumah, R.; Duca, R.C.; Galea, K.S.; Godderis, L.; Goen, T.; Hardy, E.; Iavicoli, I.; et al. HBM4EU chromates study—Overall results and recommendations for the biomonitoring of occupational exposure to hexavalent chromium. Environ. Res. 2022, 204, 111984. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic Mechanisms of Five Heavy Metals: Mercury, Lead, Chromium, Cadmium, and Arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Oginawati, K.; Susetyo, S.H.; Rosalyn, F.A.; Kurniawan, S.B.; Abdullah, S.R.S. Risk analysis of inhaled hexavalent chromium (Cr6+) exposure on blacksmiths from industrial area. Environ. Sci. Pollut. Res. Int. 2021, 28, 14000–14008. [Google Scholar] [CrossRef]
- Ferdous, Z.; Nemmar, A. Health Impact of Silver Nanoparticles: A Review of the Biodistribution and Toxicity Following Various Routes of Exposure. Int. J. Mol. Sci. 2020, 21, 2375. [Google Scholar] [CrossRef]
- Niu, Y.; Jiang, X.; Wang, K.; Xia, J.; Jiao, W.; Niu, Y.; Yu, H. Meta analysis of heavy metal pollution and sources in surface sediments of Lake Taihu, China. Sci. Total Environ. 2020, 700, 134509. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, Y.J.; Seo, Y.R. An Overview of Carcinogenic Heavy Metal: Molecular Toxicity Mechanism and Prevention. J. Cancer Prev. 2015, 20, 232–240. [Google Scholar] [CrossRef]
- Tsoucas, D.; Dong, R.; Chen, H.; Zhu, Q.; Guo, G.; Yuan, G.C. Accurate estimation of cell-type composition from gene expression data. Nat. Commun. 2019, 10, 2975. [Google Scholar] [CrossRef]
- Wijesooriya, K.; Jadaan, S.A.; Perera, K.L.; Kaur, T.; Ziemann, M. Urgent need for consistent standards in functional enrichment analysis. PLoS Comput. Biol. 2022, 18, e1009935. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Zhao, M.; Xu, J.; Li, A.; Mei, Y.; Ge, X.; Liu, X.; Wei, L.; Xu, Q. Multiple exposure pathways and urinary chromium in residents exposed to chromium. Environ. Int. 2020, 141, 105753. [Google Scholar] [CrossRef]
- Kan, Y.; Ohashi, T.; Kobayashi, E.; Kumagai, A.; Uhara, H. Cement and Retarder-Associated Systemic Contact Dermatitis in a Chromium-Negative Patch-Tested Patient. Dermatitis 2021, 32, e76–e78. [Google Scholar] [CrossRef] [PubMed]
- Hagvall, L.; Pour, M.D.; Feng, J.; Karma, M.; Hedberg, Y.; Malmberg, P. Skin permeation of nickel, cobalt and chromium salts in ex vivo human skin, visualized using mass spectrometry imaging. Toxicol. In Vitro 2021, 76, 105232. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, E.; Cervinka, M. The role of intracellular zinc in chromium(VI)-induced oxidative stress, DNA damage and apoptosis. Chem. Biol. Interact. 2006, 162, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.; Summer, B.; Sander, C.A.; Przybilla, B.; Thomas, M.; Naumann, T. Intolerance of osteosynthesis material: Evidence of dichromate contact allergy with concomitant oligoclonal T-cell infiltrate and TH1-type cytokine expression in the peri-implantar tissue. Allergy 2000, 55, 969–972. [Google Scholar] [CrossRef]
- Leroyer, C.; Dewitte, J.D.; Bassanets, A.; Boutoux, M.; Daniel, C.; Clavier, J. Occupational asthma due to chromium. Respiration 1998, 65, 403–405. [Google Scholar] [CrossRef]
- Pavesi, T.; Moreira, J.C. Mechanisms and individuality in chromium toxicity in humans. J. Appl. Toxicol. 2020, 40, 1183–1197. [Google Scholar] [CrossRef]
- Guo, L.; Xiao, Y.; Wang, Y. Hexavalent chromium-induced alteration of proteomic landscape in human skin fibroblast cells. J. Proteome Res. 2013, 12, 3511–3518. [Google Scholar] [CrossRef]
- Liu, F.J.; Barchowsky, A.; Opresko, P.L. The Werner syndrome protein suppresses telomeric instability caused by chromium (VI) induced DNA replication stress. PLoS ONE 2010, 5, e11152. [Google Scholar] [CrossRef]
- VonHandorf, A.; Sanchez-Martin, F.J.; Biesiada, J.; Zhang, H.; Zhang, X.; Medvedovic, M.; Puga, A. Chromium disrupts chromatin organization and CTCF access to its cognate sites in promoters of differentially expressed genes. Epigenetics 2018, 13, 363–375. [Google Scholar] [CrossRef]
- Zhao, Y.; Cholewa, J.; Shang, H.; Yang, Y.; Ding, X.; Wang, Q.; Su, Q.; Zanchi, N.E.; Xia, Z. Advances in the Role of Leucine-Sensing in the Regulation of Protein Synthesis in Aging Skeletal Muscle. Front. Cell Dev. Biol. 2021, 9, 646482. [Google Scholar] [CrossRef]
- Wise, S.S.; Holmes, A.L.; Wise, J.P., Sr. Hexavalent chromium-induced DNA damage and repair mechanisms. Rev. Environ. Health 2008, 23, 39–57. [Google Scholar] [CrossRef]
- Toebak, M.J.; Pohlmann, P.R.; Sampat-Sardjoepersad, S.C.; von Blomberg, B.M.; Bruynzeel, D.P.; Scheper, R.J.; Rustemeyer, T.; Gibbs, S. CXCL8 secretion by dendritic cells predicts contact allergens from irritants. Toxicol. In Vitro 2006, 20, 117–124. [Google Scholar] [CrossRef]
- Astakhova, A.; Chistyakov, D.; Thomas, D.; Geisslinger, G.; Brune, B.; Sergeeva, M.; Namgaladze, D. Inhibitors of Oxidative Phosphorylation Modulate Astrocyte Inflammatory Responses through AMPK-Dependent Ptgs2 mRNA Stabilization. Cells 2019, 8, 1185. [Google Scholar] [CrossRef]
- Jiang, X.; Xie, H.; Dou, Y.; Yuan, J.; Zeng, D.; Xiao, S. Expression and function of FRA1 protein in tumors. Mol. Biol. Rep. 2020, 47, 737–752. [Google Scholar] [CrossRef]
- Ade, N.; Leon, F.; Pallardy, M.; Peiffer, J.L.; Kerdine-Romer, S.; Tissier, M.H.; Bonnet, P.A.; Fabre, I.; Ourlin, J.C. HMOX1 and NQO1 genes are upregulated in response to contact sensitizers in dendritic cells and THP-1 cell line: Role of the Keap1/Nrf2 pathway. Toxicol. Sci. 2009, 107, 451–460. [Google Scholar] [CrossRef]
- Hao, Z.F.; Ao, J.H.; Zhang, J.; Su, Y.M.; Yang, R.Y. ATF3 activates Stat3 phosphorylation through inhibition of p53 expression in skin cancer cells. Asian Pac. J. Cancer Prev. 2013, 14, 7439–7444. [Google Scholar] [CrossRef]
- Sadagurski, M.; Nofech-Mozes, S.; Weingarten, G.; White, M.F.; Kadowaki, T.; Wertheimer, E. Insulin receptor substrate 1 (IRS-1) plays a unique role in normal epidermal physiology. J. Cell. Physiol. 2007, 213, 519–527. [Google Scholar] [CrossRef]
- Xu, J.; Liu, H.; Yang, Y.; Wang, X.; Liu, P.; Li, Y.; Meyers, C.; Banerjee, N.S.; Wang, H.K.; Cam, M.; et al. Genome-Wide Profiling of Cervical RNA-Binding Proteins Identifies Human Papillomavirus Regulation of RNASEH2A Expression by Viral E7 and E2F1. mBio 2019, 10, e02687-18. [Google Scholar] [CrossRef]
- Kurt, O.K.; Basaran, N. Occupational Exposure to Metals and Solvents: Allergy and Airway Diseases. Curr. Allergy Asthma Rep. 2020, 20, 38. [Google Scholar] [CrossRef]
- Alvarez, C.C.; Bravo Gomez, M.E.; Hernandez Zavala, A. Hexavalent chromium: Regulation and health effects. J. Trace Elem. Med. Biol. 2021, 65, 126729. [Google Scholar] [CrossRef]
- Behrens, T.; Ge, C.; Vermeulen, R.; Kendzia, B.; Olsson, A.; Schuz, J.; Kromhout, H.; Pesch, B.; Peters, S.; Portengen, L.; et al. Occupational exposure to nickel and hexavalent chromium and the risk of lung cancer in a pooled analysis of case-control studies (SYNERGY). Int. J. Cancer 2023, 152, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Seidler, A.; Jahnichen, S.; Hegewald, J.; Fishta, A.; Krug, O.; Ruter, L.; Strik, C.; Hallier, E.; Straube, S. Systematic review and quantification of respiratory cancer risk for occupational exposure to hexavalent chromium. Int. Arch. Occup. Environ. Health 2013, 86, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Shi, X. Gene expression profile in response to chromium-induced cell stress in A549 cells. Mol. Cell. Biochem. 2001, 222, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Laubli, H.; Borsig, L. Altered Cell Adhesion and Glycosylation Promote Cancer Immune Suppression and Metastasis. Front. Immunol. 2019, 10, 2120. [Google Scholar] [CrossRef] [PubMed]
- Sousa, B.; Pereira, J.; Paredes, J. The Crosstalk Between Cell Adhesion and Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 1933. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Liu, W.; Yin, C.; Chu, H.; Zhang, M. Cucurbitacin E ameliorates lipopolysaccharide-evoked injury, inflammation and MUC5AC expression in bronchial epithelial cells by restraining the HMGB1-TLR4-NF-kappaB signaling. Mol. Immunol. 2019, 114, 571–577. [Google Scholar] [CrossRef]
- Zhang, X.; Shan, P.; Jiang, G.; Cohn, L.; Lee, P.J. Toll-like receptor 4 deficiency causes pulmonary emphysema. J. Clin. Investig. 2006, 116, 3050–3059. [Google Scholar] [CrossRef]
- Li, M.; Song, D.; Chen, X.; Wang, X.; Xu, L.; Yang, M.; Yang, J.; Kalvakolanu, D.V.; Wei, X.; Liu, X.; et al. RSL3 triggers glioma stem cell differentiation via the Tgm2/AKT/ID1 signaling axis. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166529. [Google Scholar] [CrossRef]
- Xie, G.; Yang, H.; Ma, D.; Sun, Y.; Chen, H.; Hu, X.; Jiang, Y.Z.; Shao, Z.M. Integration of whole-genome sequencing and functional screening identifies a prognostic signature for lung metastasis in triple-negative breast cancer. Int. J. Cancer 2019, 145, 2850–2860. [Google Scholar] [CrossRef]
- Luan, F.; Li, X.; Cheng, X.; Huangfu, L.; Han, J.; Guo, T.; Du, H.; Wen, X.; Ji, J. TNFRSF11B activates Wnt/beta-catenin signaling and promotes gastric cancer progression. Int. J. Biol. Sci. 2020, 16, 1956–1971. [Google Scholar] [CrossRef]
- Lopez-Martin, A.; Ballestin, C.; Garcia-Carbonero, R.; Castano, A.; Lopez-Rios, F.; Lopez-Encuentra, A.; Sanchez-Cespedes, M.; Castellano, D.; Bartolome, A.; Cortes-Funes, H.; et al. Prognostic value of KIT expression in small cell lung cancer. Lung Cancer 2007, 56, 405–413. [Google Scholar] [CrossRef]
- Larsen, J.E.; Nathan, V.; Osborne, J.K.; Farrow, R.K.; Deb, D.; Sullivan, J.P.; Dospoy, P.D.; Augustyn, A.; Hight, S.K.; Sato, M.; et al. ZEB1 drives epithelial-to-mesenchymal transition in lung cancer. J. Clin. Investig. 2016, 126, 3219–3235. [Google Scholar] [CrossRef]
- Beaumont, J.J.; Sedman, R.M.; Reynolds, S.D.; Sherman, C.D.; Li, L.H.; Howd, R.A.; Sandy, M.S.; Zeise, L.; Alexeeff, G.V. Cancer mortality in a Chinese population exposed to hexavalent chromium in drinking water. Epidemiology 2008, 19, 12–23. [Google Scholar] [CrossRef]
- Villa, E.; Ali, E.S.; Sahu, U.; Ben-Sahra, I. Cancer Cells Tune the Signaling Pathways to Empower de Novo Synthesis of Nucleotides. Cancers 2019, 11, 688. [Google Scholar] [CrossRef]
- Charni-Natan, M.; Aloni-Grinstein, R.; Osher, E.; Rotter, V. Liver and Steroid Hormones-Can a Touch of p53 Make a Difference? Front. Endocrinol. 2019, 10, 374. [Google Scholar] [CrossRef]
- Zatulovskiy, E.; Zhang, S.; Berenson, D.F.; Topacio, B.R.; Skotheim, J.M. Cell growth dilutes the cell cycle inhibitor Rb to trigger cell division. Science 2020, 369, 466–471. [Google Scholar] [CrossRef]
- Schulthess, G.; Lipka, G.; Compassi, S.; Boffelli, D.; Weber, F.E.; Paltauf, F.; Hauser, H. Absorption of monoacylglycerols by small intestinal brush border membrane. Biochemistry 1994, 33, 4500–4508. [Google Scholar] [CrossRef]
- Mazzolini, R.; Dopeso, H.; Mateo-Lozano, S.; Chang, W.; Rodrigues, P.; Bazzocco, S.; Alazzouzi, H.; Landolfi, S.; Hernandez-Losa, J.; Andretta, E.; et al. Brush border myosin Ia has tumor suppressor activity in the intestine. Proc. Natl. Acad. Sci. USA 2012, 109, 1530–1535. [Google Scholar] [CrossRef]
- Macedo, F.; Ladeira, K.; Longatto-Filho, A.; Martins, S.F. Gastric Cancer and Angiogenesis: Is VEGF a Useful Biomarker to Assess Progression and Remission? J. Gastric Cancer 2017, 17, 1–10. [Google Scholar] [CrossRef]
- Nautiyal, J.; Kanwar, S.S.; Majumdar, A.P. EGFR(s) in aging and carcinogenesis of the gastrointestinal tract. Curr. Protein Pept. Sci. 2010, 11, 436–450. [Google Scholar] [CrossRef]
- Arvidsson, Y.; Andersson, E.; Bergstrom, A.; Andersson, M.K.; Altiparmak, G.; Illerskog, A.C.; Ahlman, H.; Lamazhapova, D.; Nilsson, O. Amyloid precursor-like protein 1 is differentially upregulated in neuroendocrine tumours of the gastrointestinal tract. Endocr. Relat. Cancer 2008, 15, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gu, S.; Nasir, O.; Foller, M.; Ackermann, T.F.; Klingel, K.; Kandolf, R.; Kuhl, D.; Stournaras, C.; Lang, F. SGK1-dependent intestinal tumor growth in APC-deficient mice. Cell. Physiol. Biochem. 2010, 25, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, J.; Zhang, W.; Gan, J.; Hu, C.; Huang, G.; Zhang, Y. lncRNA GAS5 enhances G1 cell cycle arrest via binding to YBX1 to regulate p21 expression in stomach cancer. Sci. Rep. 2015, 5, 10159. [Google Scholar] [CrossRef] [PubMed]
- Tattoli, I.; Petitta, C.; Scirocco, A.; Ammoscato, F.; Cicenia, A.; Severi, C. Microbiota, innate immune system, and gastrointestinal muscle: Ongoing studies. J. Clin. Gastroenterol. 2012, 46, S6–S11. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Hameed, A.; Hasnain, S. Chromium-resistant bacteria and cyanobacteria: Impact on Cr(VI) reduction potential and plant growth. J. Ind. Microbiol. Biotechnol. 2005, 32, 615–621. [Google Scholar] [CrossRef]
- Shrivastava, H.Y.; Ravikumar, T.; Shanmugasundaram, N.; Babu, M.; Unni Nair, B. Cytotoxicity studies of chromium(III) complexes on human dermal fibroblasts. Free Radic. Biol. Med. 2005, 38, 58–69. [Google Scholar] [CrossRef]
- Rager, J.E.; Suh, M.; Chappell, G.A.; Thompson, C.M.; Proctor, D.M. Review of transcriptomic responses to hexavalent chromium exposure in lung cells supports a role of epigenetic mediators in carcinogenesis. Toxicol. Lett. 2019, 305, 40–50. [Google Scholar] [CrossRef]
- Tumolo, M.; Ancona, V.; De Paola, D.; Losacco, D.; Campanale, C.; Massarelli, C.; Uricchio, V.F. Chromium Pollution in European Water, Sources, Health Risk, and Remediation Strategies: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 5438. [Google Scholar] [CrossRef]
- Lou, J.; Yu, S.; Feng, L.; Guo, X.; Wang, M.; Branco, A.T.; Li, T.; Lemos, B. Environmentally induced ribosomal DNA (rDNA) instability in human cells and populations exposed to hexavalent chromium [Cr (VI)]. Environ. Int. 2021, 153, 106525. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Route | Potential Toxicity Mechanism | Biological Process (BP) | Hub Genes |
|---|---|---|---|
| Dermal contact | DNA damage, metastasis, immune system disorder, allergic reaction | RNA modification, sterol biosynthesis process, DNA replication, intrinsic apoptotic signaling pathway | CXCL8, PTGS2, FOS, HMOX1, ATF3, IRS1, E2F1 |
| Inhalation | DNA damage, metastasis, cell cycle alteration, immune system disorder | Ribosome biogenesis, rRNA metabolic process, cell–substrate adhesion, macrophage cytokine production | TLR4, TGM2, TNFRSF11B, BDKRB1, KIT, ZEB1 |
| Ingestion | DNA damage, metastasis | Nucleotide biosynthesis process, nucleotide phosphate biosynthesis process, alcohol metabolic process, steroid metabolic process | VEGFA, EGFR, APP, SGK1, JUN, TLR2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, D.Y.; Lee, S.M.; Jang, Y.; Lee, J.; Lee, C.M.; Cho, E.-M.; Seo, Y.R. Adverse Human Health Effects of Chromium by Exposure Route: A Comprehensive Review Based on Toxicogenomic Approach. Int. J. Mol. Sci. 2023, 24, 3410. https://doi.org/10.3390/ijms24043410
Shin DY, Lee SM, Jang Y, Lee J, Lee CM, Cho E-M, Seo YR. Adverse Human Health Effects of Chromium by Exposure Route: A Comprehensive Review Based on Toxicogenomic Approach. International Journal of Molecular Sciences. 2023; 24(4):3410. https://doi.org/10.3390/ijms24043410
Chicago/Turabian StyleShin, Dong Yeop, Sang Min Lee, Yujin Jang, Jun Lee, Cheol Min Lee, Eun-Min Cho, and Young Rok Seo. 2023. "Adverse Human Health Effects of Chromium by Exposure Route: A Comprehensive Review Based on Toxicogenomic Approach" International Journal of Molecular Sciences 24, no. 4: 3410. https://doi.org/10.3390/ijms24043410
APA StyleShin, D. Y., Lee, S. M., Jang, Y., Lee, J., Lee, C. M., Cho, E.-M., & Seo, Y. R. (2023). Adverse Human Health Effects of Chromium by Exposure Route: A Comprehensive Review Based on Toxicogenomic Approach. International Journal of Molecular Sciences, 24(4), 3410. https://doi.org/10.3390/ijms24043410