
Genomics, Proteomics, and Metabolomics Approaches to Improve Abiotic Stress Tolerance in Tomato Plant

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
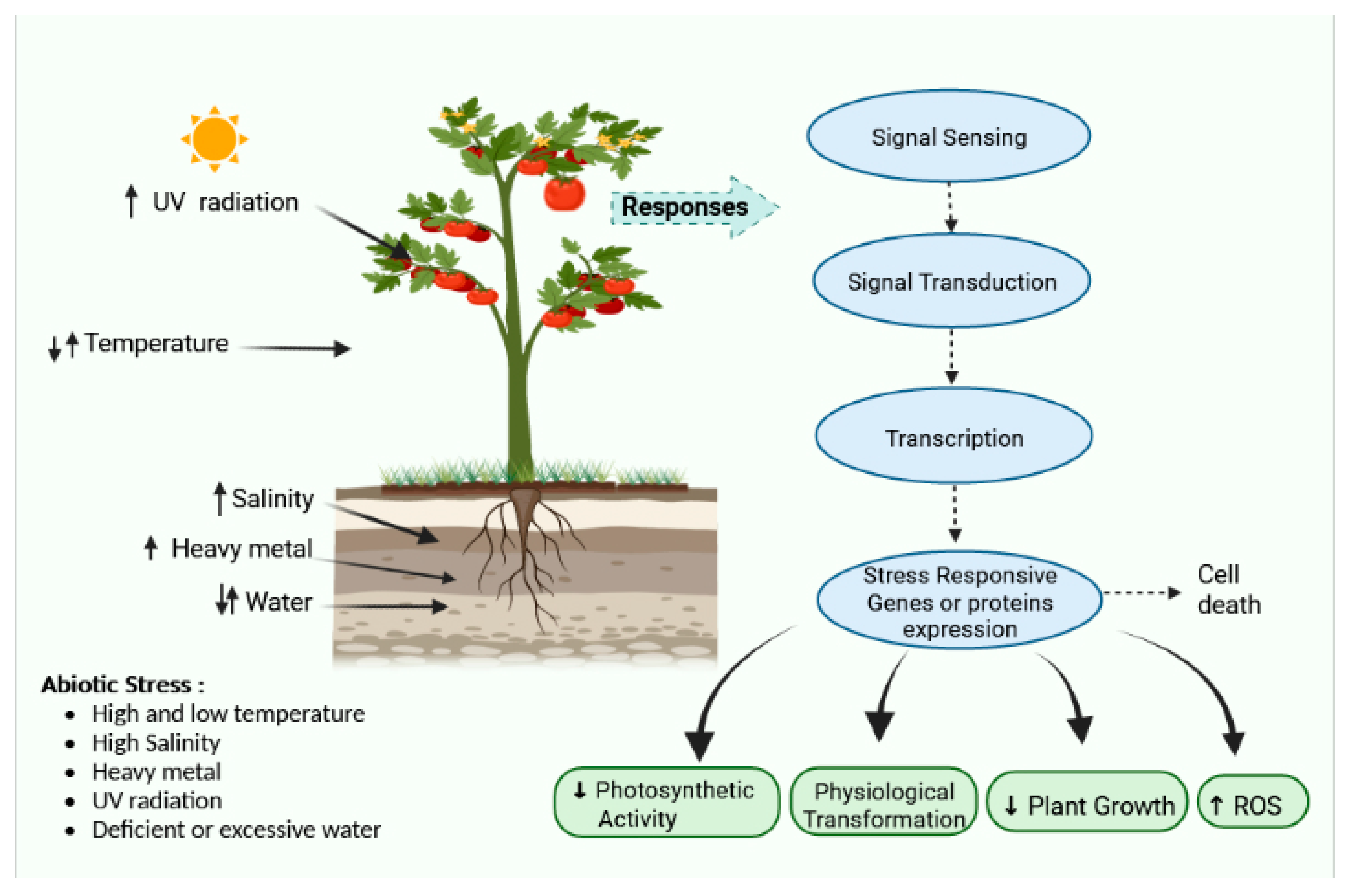
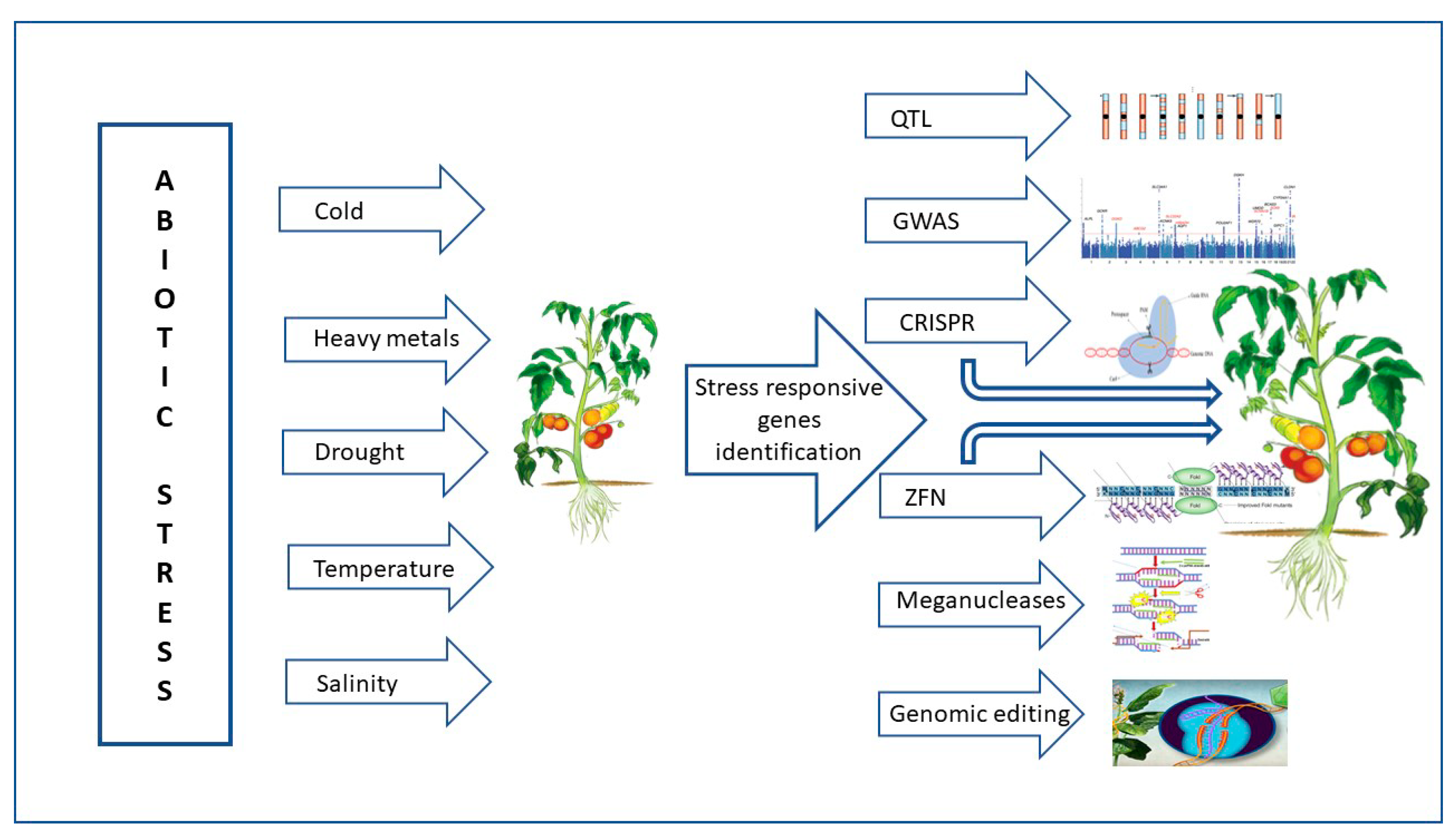
2. Major Abiotic Stress in Tomato
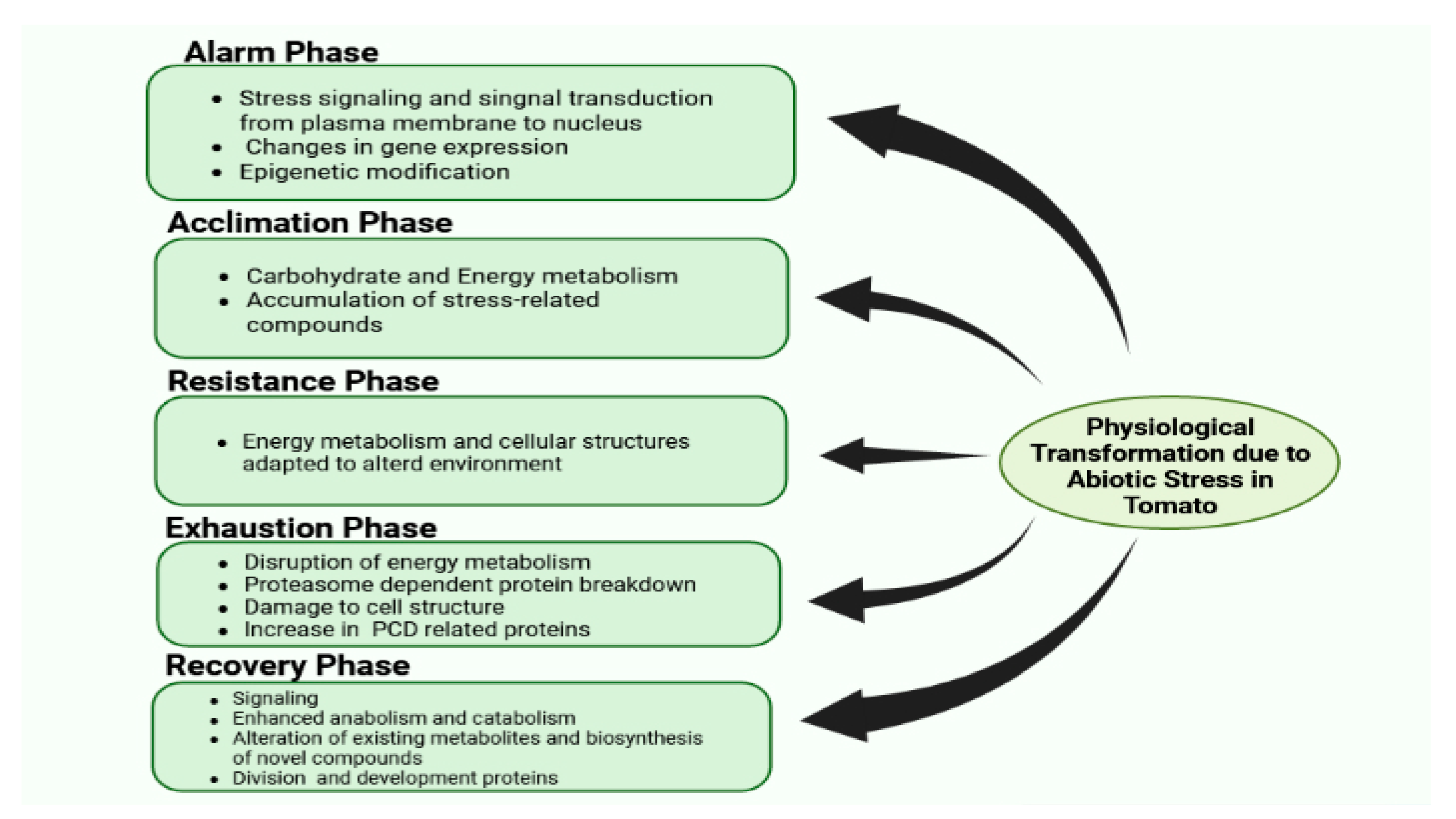
3. Physiological Transformation Due to Abiotic Stress in Tomato
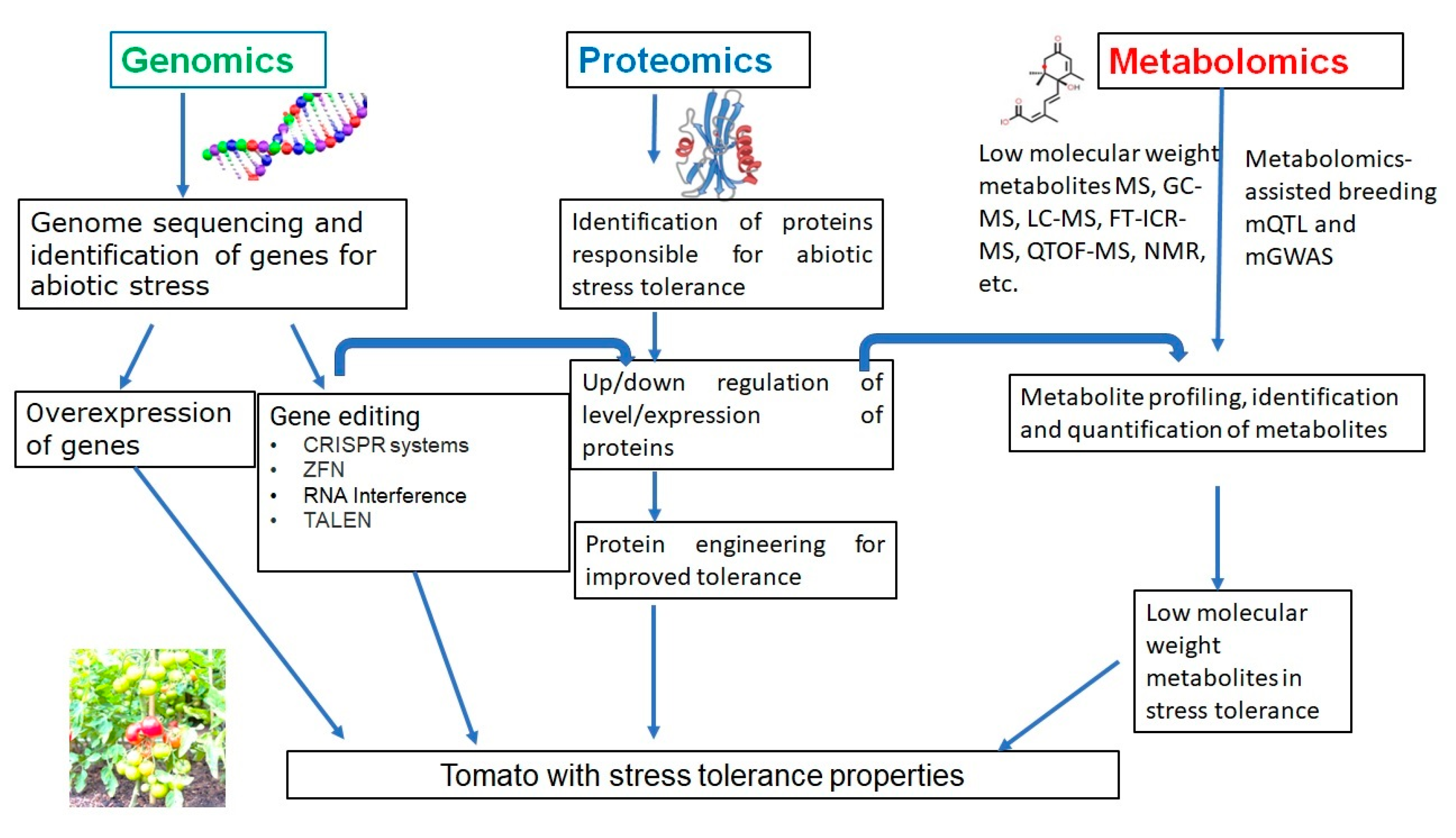
4. Genomics of Abiotic Stress
4.1. Genome Sequencing of Tomato
4.2. Identification and Functional Validation of Genes Associated with Abiotic Stress Tolerance
4.3. Genomic Approaches to Combat Abiotic Stress
5. Proteomics of Abiotic Stress in Tomato
5.1. The Fruit Proteome of Tomato
5.2. Protein Cellular Localization (PCL)
5.3. Role of Protein Isoforms and Post-Translational Modifications (PTMs)
5.4. Proteome Description of Protein Biological Function
6. Metabolomics of Tomato in Stress Management
6.1. Metabolomics Technologies and Advancements
6.2. Metabolomics Applications in Plant Stress Responses
6.3. Plant Metabolites and Ecological Adaptation
6.4. Plant Metabolites in Response to Stresses
6.4.1. Metabolomics of Plant–Microbe Interactions
6.4.2. Metabolomics of Plant–Herbivore Interactions
6.4.3. Impact of Pesticides and Other Chemicals and Epigenetic Modifications on the Metabolomics of Tomato

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Function | Result | References |
|---|---|---|---|
| Melatonin | Prevents damage to proteins and membranes | Tolerance against abiotic stress | [210] |
| ABP19a protein (drought-responsive auxin-binding protein (ABPs) family | Involved in many development processes and drought response | Drought tolerance | [211,212] |
| SpUSP, an annexin-interacting universal stress protein | Reduction in oxidative stress by preventing the generation of ROS, activation of several stress-responsive genes that causes the accumulation of some osmoprotective solutes | Drought tolerance | [213] |
| Solyc04g014600 (Universal stress protein) | Protein profiling of phloem and its exudates | Drought tolerance | [214] |
| SlSRN1 (Solanum lycopersicum stress-related NAC1) | Disease resistance response as well as resistance to drought and oxidative stress | Tolerance against abiotic stress | [215] |
| SlNAC4 | Fruit ripening and carotenoid accumulation | Oxidative stress response | [216] |
| SlNAM2 | Flower-boundary morphogenesis | Inhibits extreme water loss | [217] |
| SlNAC1 | Increasing viral replication and chill tolerance through interactions with replication accessory proteins | Tolerance to lower temperatures and phytopathogens | [218,219] |
| GOBLET | Identifying the borders of compound leaves’ leaflets | Inhibits extreme transpiration | [220] |
| SlMAPK3 | Enhanced germination rate as well as seedling growth; moreover, formed transgenic plants resulted in an improved chlorophyll content and root biomass accumulation under Cd2+ stress | Tolerance to cadmium stress along with drought tolerance | [65] |
| C2H2-Type Zinc Finger Protein | Overall growth as well as the development of plant tissues | Abiotic stress responses | [221] |
7. Conclusions and Future Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Approval
References
- Kumar, M.; Tomar, M.; Bhuyan, D.J.; Punia, S.; Grasso, S.; Sá, A.G.A.; Carciofi, B.A.M.; Arrutia, F.; Changan, S.; Radha; et al. Tomato (Solanum lycopersicum L.) seed: A review on bioactives and biomedical activities. Biomed. Pharmacother. 2021, 142, 112018. [Google Scholar] [CrossRef] [PubMed]
- Ali, Y.; Ibn Sina, A.A.; Khandker, S.S.; Neesa, L.; Tanvir, E.M.; Kabir, A.; Khalil, I.; Gan, S.H. Nutritional Composition and Bioactive Compounds in Tomatoes and Their Impact on Human Health and Disease: A Review. Foods 2020, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, I.; Leça, J.M.; Aguiar, R.; Fernandes, T.; Marques, J.C.; Cordeiro, N. Influence of Crop System Fruit Quality, Carotenoids, Fatty Acids and Phenolic Compounds in Cherry Tomatoes. Agric. Res. 2021, 10, 56–65. [Google Scholar] [CrossRef]
- Mathivanan, S. Abiotic Stress-Induced Molecular and Physiological Changes and Adaptive Mechanisms in Plants. In Abiotic Stress Plants; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q.; Channa, M.M.; Alshegaihi, R.M.; Meng, Q.; Ali, M.; Zaman, W.; Brohi, R.D.; et al. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Chele, K.H. Machine Learning in Plant Metabolomics and the Elucidation of the Biochemistry Underlying the Effects of a Nonmicrobial Biostimulant on Tomato Plants Under Salt Stress Conditions; University of Johannesburg: Johannesburg, South Africa, 2020. [Google Scholar]
- Debnath, M.; Pandey, M.; Bisen, P. An Omics Approach to Understand the Plant Abiotic Stress. Omics 2011, 15, 739–762. [Google Scholar] [CrossRef]
- Rehem, B.C.; Bertolde, F.Z.; de Almeida, A.A.F. Regulation of Gene Expression in Response to Abiotic Stress in Plants; InTech: London, UK, 2012; pp. 13–38. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Klíma, M.; Roy, A.; Prášil, I.T. Biological Networks Underlying Abiotic Stress Tolerance in Temperate Crops—A Proteomic Perspective. Int. J. Mol. Sci. 2015, 16, 20913–20942. [Google Scholar] [CrossRef]
- Mustafa, G.; Akhtar, M.S.; Abdullah, R. Global Concern for Salinity on Various Agro-Ecosystems. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Springer: Singapore, 2019; pp. 1–19. [Google Scholar]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Prášil, I.T.; Renaut, J. Plant Abiotic Stress Proteomics: The Major Factors Determining Alterations in Cellular Proteome. Front. Plant Sci. 2018, 9, 122. [Google Scholar] [CrossRef]
- Hossain, Z.; Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front. Plant Sci. 2012, 3, 310. [Google Scholar] [CrossRef]
- Ahsan, N.; Renaut, J.; Komatsu, S. Recent developments in the application of proteomics to the analysis of plant responses to heavy metals. Proteomics 2009, 9, 2602–2621. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Rajamani, U.; Verma, J.; Subba, P.; Chakraborty, N.; Datta, A.; Chakraborty, S.; Chakraborty, N. Identification of Extracellular Matrix Proteins of Rice (Oryza sativa L.) Involved in Dehydration-Responsive Network: A Proteomic Approach. J. Proteome Res. 2010, 9, 3443–3464. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative Modifications to Cellular Components in Plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Hajheidari, M.; Eivazi, A.; Buchanan, B.B.; Wong, J.H.; Majidi, I.; Salekdeh, G.H. Proteomics Uncovers a Role for Redox in Drought Tolerance in Wheat. J. Proteome Res. 2007, 6, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Zhou, Y.H.; Xia, X.J.; Mao, W.H.; Shi, K.; Yu, J.Q. Brassinosteroid regulates secondary metabolism in tomato towards enhanced tolerance to phenanthrene. Biol. Plant. 2013, 57, 154–158. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elkelish, A.; Soliman, M.; Elansary, H.O.; Zaid, A.; Wani, S.H. Serratia marcescens BM1 Enhances Cadmium Stress Tolerance and Phytoremediation Potential of Soybean Through Modulation of Osmolytes, Leaf Gas Exchange, Antioxidant Machinery, and Stress-Responsive Genes Expression. Antioxidants 2020, 9, 43. [Google Scholar] [CrossRef]
- Mousavi, A.; Hotta, Y. Glycine-Rich Proteins: A Class of Novel Proteins. Appl. Biochem. Biotechnol. 2005, 120, 169–174. [Google Scholar] [CrossRef]
- Rai, G.; Parveen, A.; Jamwal, G.; Basu, U.; Kumar, R.; Rai, P.; Sharma, J.; Alalawy, A.; Al-Duais, M.; Hossain, M.; et al. Leaf Proteome Response to Drought Stress and Antioxidant Potential in Tomato (Solanum lycopersicum L.). Atmosphere 2021, 12, 1021. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Li, R.; Liu, C.; Zhao, R.; Wang, L.; Chen, L.; Yu, W.; Zhang, S.; Sheng, J.; Shen, L. CRISPR/Cas9-Mediated SlNPR1 mutagenesis reduces tomato plant drought tolerance. BMC Plant Biol. 2019, 19, 38. [Google Scholar] [CrossRef] [PubMed]
- Gerszberg, A.; Hnatuszko-Konka, K. Tomato tolerance to abiotic stress: A review of most often engineered target sequences. Plant Growth Regul. 2017, 83, 175–198. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Zhao, S.; Liu, J.-F.; Zhao, H.-Y.; Sun, X.-Y.; Wu, T.-R.; Pei, T.; Wang, Y.; Liu, Q.-F.; Yang, H.-H.; et al. Genome-wide identification of Tomato Golden 2-Like transcription factors and abiotic stress related members screening. BMC Plant Biol. 2022, 22, 82. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Ichihashi, Y.; Sinha, N.R. The tomato genome: Implications for plant breeding, genomics and evolution. Genome Biol. 2012, 13, 167. [Google Scholar] [CrossRef]
- Dheer, P.; Rautela, I.; Sharma, V.; Dhiman, M.; Sharma, A.; Sharma, N.; Sharma, M.D. Evolution in crop improvement approaches and future prospects of molecular markers to CRISPR/Cas9 system. Gene 2020, 753, 144795. [Google Scholar] [CrossRef]
- Cambiaso, V.; Pratta, G.R.; da Costa, J.H.P.; Zorzoli, R.; Francis, D.M.; Rodríguez, G.R. Whole genome re-sequencing analysis of two tomato genotypes for polymorphism insight in cloned genes and a genetic map construction. Sci. Hortic. 2019, 247, 58–66. [Google Scholar] [CrossRef]
- Chaudhary, J.; Khatri, P.; Singla, P.; Kumawat, S.; Kumari, A.; R, V.; Vikram, A.; Jindal, S.K.; Kardile, H.; Kumar, R.; et al. Advances in Omics Approaches for Abiotic Stress Tolerance in Tomato. Biology 2019, 8, 90. [Google Scholar] [CrossRef]
- Zhu, M.; Meng, X.; Cai, J.; Li, G.; Dong, T.; Li, Z. Basic leucine zipper transcription factor SlbZIP1 mediates salt and drought stress tolerance in tomato. BMC Plant Biol. 2018, 18, 83. [Google Scholar] [CrossRef]
- Szymanski, J.; Levin, Y.; Savidor, A.; Breitel, D.; Chappell-Maor, L.; Heinig, U.; Töpfer, N.; Aharoni, A. Label-free deep shotgun proteomics reveals protein dynamics during tomato fruit tissues development. Plant J. 2017, 90, 396–417. [Google Scholar] [CrossRef]
- Kausar, R.; Wang, X.; Komatsu, S. Crop Proteomics under Abiotic Stress: From Data to Insights. Plants 2022, 11, 2877. [Google Scholar] [CrossRef] [PubMed]
- Krishna, R.; Karkute, S.G.; Ansari, W.A.; Jaiswal, D.K.; Verma, J.P.; Singh, M. Transgenic tomatoes for abiotic stress tolerance: Status and way ahead. 3 Biotech 2019, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tang, N.; Xian, Z.; Li, Z. Two SnRK2 protein kinases genes play a negative regulatory role in the osmotic stress response in tomato. Plant Cell Tissue Organ Cult. 2015, 122, 421–434. [Google Scholar] [CrossRef]
- Yu, L.; Yan, J.; Yang, Y.; He, L.; Zhu, W. Enhanced Tolerance to Chilling Stress in Tomato by Overexpression of a Mitogen-Activated Protein Kinase, SlMPK7. Plant Mol. Biol. Rep. 2016, 34, 76–88. [Google Scholar] [CrossRef]
- Liu, H.; Yu, C.; Li, H.; Ouyang, B.; Wang, T.; Zhang, J.; Wang, X.; Ye, Z. Overexpression of ShDHN, a dehydrin gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses in tomato. Plant Sci. 2015, 231, 198–211. [Google Scholar] [CrossRef]
- Li, J.-B.; Luan, Y.-S.; Liu, Z. SpWRKY1 mediates resistance to Phytophthora infestans and tolerance to salt and drought stress by modulating reactive oxygen species homeostasis and expression of defense-related genes in tomato. Plant Cell Tissue Organ Cult. 2015, 123, 67–81. [Google Scholar] [CrossRef]
- Shah, S.H.; Ali, S.; Jan, S.A.; Din, J.U.; Ali, G.M. Piercing and incubation method of in planta transformation producing stable transgenic plants by overexpressing DREB1A gene in tomato (Solanum lycopersicum Mill.). Plant Cell Tissue Organ Cult. 2015, 120, 1139–1157. [Google Scholar] [CrossRef]
- Meng, X.; Wang, J.-R.; Wang, G.-D.; Liang, X.-Q.; Li, X.-D.; Meng, Q.-W. An R2R3-MYB gene, LeAN2, positively regulated the thermo-tolerance in transgenic tomato. J. Plant Physiol. 2015, 175, 1–8. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, Q.; Sprague, S.A.; Park, J.; Oh, M.; Rajashekar, C.B.; Koiwa, H.; Nakata, P.A.; Cheng, N.; Hirschi, K.D.; et al. Tomato expressing Arabidopsis glutaredoxin gene AtGRXS17 confers tolerance to chilling stress via modulating cold responsive components. Hortic. Res. 2015, 2, 15051. [Google Scholar] [CrossRef]
- Hu, D.-G.; Ma, Q.-J.; Sun, C.-H.; Sun, M.-H.; You, C.-X.; Hao, Y.-J. Overexpression of MdSOS2L1, a CIPK protein kinase, increases the antioxidant metabolites to enhance salt tolerance in apple and tomato. Physiol. Plant. 2016, 156, 201–214. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; Vandenlangenberg, K.M.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Overexpression of S-adenosyl-l-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant Biotechnol. J. 2014, 12, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Metwali, E.M.R.; Soliman, H.I.A.; Fuller, M.; Almaghrabi, O.A. Improving fruit quality in tomato (Lycopersicum esculentum Mill) under heat stress by silencing the vis 1 gene using small interfering RNA technology. Plant Cell, Tissue Organ Cult. 2015, 121, 153–166. [Google Scholar] [CrossRef]
- Naeem, M.; Shahzad, K.; Saqib, S.; Shahzad, A.; Nasrullah; Younas, M.; Afridi, M.I. The Solanum melongena COP1LIKE manipulates fruit ripening and flowering time in tomato (Solanum lycopersicum). Plant Growth Regul. 2022, 96, 369–382. [Google Scholar] [CrossRef]
- Habib, S.; Lwin, Y.; Li, N. Down-Regulation of SlGRAS10 in Tomato Confers Abiotic Stress Tolerance. Genes 2021, 12, 623. [Google Scholar] [CrossRef] [PubMed]
- Orellana, S.; Yañez, M.; Espinoza, A.; Verdugo, I.; González, E.; Ruiz-Lara, S.; Casaretto, J.A. The transcription factor SlAREB1 confers drought, salt stress tolerance and regulates biotic and abiotic stress-related genes in tomato. Plant Cell Environ. 2010, 33, 2191–2208. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Bao, Y.-F.; Pei, T.; Wu, T.-R.; DU, X.; He, M.-X.; Wang, Y.; Liu, Q.-F.; Yang, H.-H.; Jiang, J.-B.; et al. Silencing the SLB3 transcription factor gene decreases drought stress tolerance in tomato. J. Integr. Agric. 2020, 19, 2699–2708. [Google Scholar] [CrossRef]
- Bvindi, C.; Lee, S.; Tang, L.; Mickelbart, M.V.; Li, Y.; Mengiste, T. Improved pathogen and stress tolerance in tomato mutants of SET domain histone 3 lysine methyltransferases. New Phytol. 2022, 235, 1957–1976. [Google Scholar] [CrossRef]
- Wai, A.H.; Naing, A.H.; Lee, D.-J.; Kil Kim, C.; Chung, M.-Y. Molecular genetic approaches for enhancing stress tolerance and fruit quality of tomato. Plant Biotechnol. Rep. 2020, 14, 515–537. [Google Scholar] [CrossRef]
- Pan, C.; Yang, D.; Zhao, X.; Jiao, C.; Yan, Y.; Lamin-Samu, A.T.; Wang, Q.; Xu, X.; Fei, Z.; Lu, G. Tomato stigma exsertion induced by high temperature is associated with the jasmonate signalling pathway. Plant Cell Environ. 2019, 42, 1205–1221. [Google Scholar] [CrossRef]
- Bineau, E.; Diouf, I.; Carretero, Y.; Duboscq, R.; Bitton, F.; Djari, A.; Zouine, M.; Causse, M. Genetic diversity of tomato response to heat stress at the QTL and transcriptome levels. Plant J. 2021, 107, 1213–1227. [Google Scholar] [CrossRef]
- Sidhu, N.S.; Pruthi, G.; Singh, S.; Bishnoi, R.; Singla, D. Genome-wide identification and analysis of GRAS transcription factors in the bottle gourd genome. Sci. Rep. 2020, 10, 14338. [Google Scholar] [CrossRef] [PubMed]
- Gisbert, C.; Rus, A.M.; Bolarín, M.C.; López-Coronado, J.M.; Arrillaga, I.; Montesinos, C.; Caro, M.; Serrano, R.; Moreno, V. The Yeast HAL1 Gene Improves Salt Tolerance of Transgenic Tomato. Plant Physiol. 2000, 123, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.H.; Kumar, S.A.; Sivan, P.; Katam, R.; Suravajhala, P.; Rao, K.S.; Varshney, R.K.; Kishor, P.B.K. Overexpression of a Plasma Membrane Bound Na+/H+ Antiporter-Like Protein (SbNHXLP) Confers Salt Tolerance and Improves Fruit Yield in Tomato by Maintaining Ion Homeostasis. Front. Plant Sci. 2016, 7, 2027. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hao, J.; Chen, X.; Zhang, X. SlMYB102 expression enhances low-temperature stress resistance in tomato plants. PeerJ 2020, 8, e10059. [Google Scholar] [CrossRef]
- He, Y.; Yang, T.; Yan, S.; Niu, S.; Zhang, Y. Identification and characterization of the BEL1-like genes reveal their potential roles in plant growth and abiotic stress response in tomato. Int. J. Biol. Macromol. 2022, 200, 193–205. [Google Scholar] [CrossRef]
- AbuQamar, S.; Luo, H.; Laluk, K.; Mickelbart, M.V.; Mengiste, T. Crosstalk between biotic and abiotic stress responses in tomato is mediated by the AIM1 transcription factor. Plant J. 2009, 58, 347–360, Erratum in: Plant J. 2009, 60, 929. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2018, 16, 354–366. [Google Scholar] [CrossRef]
- Yin, W.; Hu, Z.; Cui, B.; Guo, X.; Hu, J.; Zhu, Z.; Chen, G. Suppression of the MADS-box gene SlMBP8 accelerates fruit ripening of tomato (Solanum lycopersicum). Plant Physiol. Biochem. 2017, 118, 235–244. [Google Scholar] [CrossRef]
- Xu, X.-X.; Hu, Q.; Yang, W.-N.; Jin, Y. The roles of cell wall invertase inhibitor in regulating chilling tolerance in tomato. BMC Plant Biol. 2017, 17, 195. [Google Scholar] [CrossRef]
- Yang, L.; Huang, W.; Xiong, F.; Xian, Z.; Su, D.; Ren, M.; Li, Z. Silencing of SlPL, which encodes a pectate lyase in tomato, confers enhanced fruit firmness, prolonged shelf-life and reduced susceptibility to grey mould. Plant Biotechnol. J. 2017, 15, 1544–1555. [Google Scholar] [CrossRef]
- Muhammad, T.; Zhang, J.; Ma, Y.; Li, Y.; Zhang, F.; Zhang, Y.; Liang, Y. Overexpression of a Mitogen-Activated Protein Kinase SlMAPK3 Positively Regulates Tomato Tolerance to Cadmium and Drought Stress. Molecules 2019, 24, 556. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhao, S.; Bao, T.; Zhao, P.; Peng, K.; Guo, Q.; Gao, X.; Qin, J. Tomato BZR/BES transcription factor SlBZR1 positively regulates BR signaling and salt stress tolerance in tomato and Arabidopsis. Plant Sci. 2021, 302, 110719. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Liu, G.; Chen, W.; Zhang, F.; Li, H.; Ye, Z.; Zhang, Y. Knockdown of SlNL33 accumulates ascorbate, enhances disease and oxidative stress tolerance in tomato (Solanum lycopersicum). Plant Growth Regul. 2019, 89, 49–58. [Google Scholar] [CrossRef]
- Han, N.; Fan, S.; Zhang, T.; Sun, H.; Zhu, Y.; Gong, H.; Guo, J. SlHY5 is a necessary regulator of the cold acclimation response in tomato. Plant Growth Regul. 2020, 91, 1–12. [Google Scholar] [CrossRef]
- Lu, J.; Sun, M.-H.; Ma, Q.-J.; Kang, H.; Liu, Y.-J.; Hao, Y.-J.; You, C.-X. MdSWEET17, a sugar transporter in apple, enhances drought tolerance in tomato. J. Integr. Agric. 2019, 18, 2041–2051. [Google Scholar] [CrossRef]
- Guo, X.; Zhang, L.; Wang, X.; Zhang, M.; Xi, Y.; Wang, A.; Zhu, J. Overexpression of Saussurea involucrata dehydrin gene SiDHN promotes cold and drought tolerance in transgenic tomato plants. PLoS ONE 2019, 14, e0225090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhao, H.; Shi, J.; Wu, Y.; Jiang, J. Functional characterization of class I SlHSP17.7 gene responsible for tomato cold-stress tolerance. Plant Sci. 2020, 298, 110568. [Google Scholar] [CrossRef]
- Ding, F.; Qiang, X.; Jia, Z.; Li, L.; Hu, J.; Yin, M.; Xia, S.; Chen, B.; Qi, J.; Li, Q.; et al. Knockout of a novel salt responsive gene SlABIG1 enhance salinity tolerance in tomato. Environ. Exp. Bot. 2022, 200, 104903. [Google Scholar] [CrossRef]
- Balyan, S.; Rao, S.; Jha, S.; Bansal, C.; Das, J.R.; Mathur, S. Characterization of novel regulators for heat stress tolerance in tomato from Indian sub-continent. Plant Biotechnol. J. 2020, 18, 2118–2132. [Google Scholar] [CrossRef]
- Pandey, S.; Muthamilarasan, M.; Sharma, N.; Chaudhry, V.; Dulani, P.; Shweta, S.; Jha, S.; Mathur, S.; Prasad, M. Characterization of DEAD-box family of RNA helicases in tomato provides insights into their roles in biotic and abiotic stresses. Environ. Exp. Bot. 2019, 158, 107–116. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, J.; Xu, J.; Li, Y.; Guo, L.; Wang, Z.; Zhang, X.; Zhao, B.; Guo, Y.-D.; Zhang, N. CRISPR/Cas9 targeted mutagenesis of SlLBD40, a lateral organ boundaries domain transcription factor, enhances drought tolerance in tomato. Plant Sci. 2020, 301, 110683. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.; Li, Y.; Li, Z.; Sheng, J.; Shen, L. SlMAPK3 enhances tolerance to salt stress in tomato plants by scavenging ROS accumulation and up-regulating the expression of ethylene signaling related genes. Environ. Exp. Bot. 2022, 193, 104698. [Google Scholar] [CrossRef]
- Mehta, S.; James, D.; Reddy, M.K. Omics Technologies for Abiotic Stress Tolerance in Plants: Current Status and Prospects. In Recent Approaches in Omics for Plant Resilience to Climate Change; Wani, S., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Soda, N.; Wallace, S.; Karan, R. Omics Study for Abiotic Stress Responses in Plants. Adv. Plants Agric. Res. 2015, 2, 28–34. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Palakolanu, S.R.; Chopra, P.; Rajurkar, A.B.; Gupta, R.; Iqbal, N.; Maheshwari, C. Improving drought tolerance in rice: Ensuring food security through multi-dimensional approaches. Physiol. Plant. 2021, 172, 645–668. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Okekeogbu, I.; Sangireddy, S.; Ye, Z.; Li, H.; Bhatti, S.; Hui, D.; McDonald, D.W.; Yang, Y.; Giri, S.; et al. Proteome Modification in Tomato Plants upon Long-Term Aluminum Treatment. J. Proteome Res. 2016, 15, 1670–1684. [Google Scholar] [CrossRef]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Cereal Crop Proteomics: Systemic Analysis of Crop Drought Stress Responses Towards Marker-Assisted Selection Breeding. Front. Plant Sci. 2017, 8, 757. [Google Scholar] [CrossRef]
- Ghosh, D.; Xu, J. Abiotic stress responses in plant roots: A proteomics perspective. Front. Plant Sci. 2014, 5, 6. [Google Scholar] [CrossRef]
- Rocco, M.; D’Ambrosio, C.; Arena, S.; Faurobert, M.; Scaloni, A.; Marra, M. Proteomic analysis of tomato fruits from two ecotypes during ripening. Proteomics 2006, 6, 3781–3791. [Google Scholar] [CrossRef]
- Kok, E.; Lehesranta, S.J.; Van Dijk, J.P.; Helsdingen, J.R.; Dijksma, W.T.P.; Van Hoef, A.M.A.; Koistinen, K.M.; Karenlampi, S.O.; Kuiper, H.A.; Keijer, J. Changes in Gene and Protein Expression during Tomato Ripening—Consequences for the Safety Assessment of New Crop Plant Varieties. Food Sci. Technol. Int. 2008, 14, 503–518. [Google Scholar] [CrossRef]
- Faurobert, M.; Mihr, C.; Bertin, N.; Pawłowski, T.A.; Negroni, L.; Sommerer, N.; Causse, M. Major Proteome Variations Associated with Cherry Tomato Pericarp Development and Ripening. Plant Physiol. 2007, 143, 1327–1346. [Google Scholar] [CrossRef]
- Tunc-Ozdemir, M.; Li, B.; Jaiswal, D.K.; Urano, D.; Jones, A.M.; Torres, M.P. Predicted Functional Implications of Phosphorylation of Regulator of G Protein Signaling Protein in Plants. Front. Plant Sci. 2017, 8, 1456. [Google Scholar] [CrossRef] [PubMed]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease Resistance Mechanisms in Plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.-K.; Falvo, J.V.; Gerke, L.C.; Carroll, A.S.; Howson, R.W.; Weissman, J.S.; O’Shea, E.K. Global analysis of protein localization in budding yeast. Nature 2003, 425, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Thul, P.J.; Lindskog, C. The human protein atlas: A spatial map of the human proteome. Protein Sci. 2018, 27, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Mackinder, L.C.; Chen, C.; Leib, R.D.; Patena, W.; Blum, S.R.; Rodman, M.; Ramundo, S.; Adams, C.M.; Jonikas, M.C. A Spatial Interactome Reveals the Protein Organization of the Algal CO2-Concentrating Mechanism. Cell 2017, 171, 133–147.e14. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Li, J.; Ting, A.Y. Proximity labeling: Spatially resolved proteomic mapping for neurobiology. Curr. Opin. Neurobiol. 2018, 50, 17–23. [Google Scholar] [CrossRef]
- Samavarchi-Tehrani, P.; Samson, R.; Gingras, A.-C. Proximity Dependent Biotinylation: Key Enzymes and Adaptation to Proteomics Approaches. Mol. Cell. Proteom. 2020, 19, 757–773. [Google Scholar] [CrossRef]
- Borner, G.H.H. Organellar Maps Through Proteomic Profiling—A Conceptual Guide. Mol. Cell. Proteom. 2020, 19, 1076–1087. [Google Scholar] [CrossRef]
- Wu, X.; Gong, F.; Cao, D.; Hu, X.; Wang, W. Advances in crop proteomics: PTMs of proteins under abiotic stress. Proteomics 2016, 16, 847–865. [Google Scholar] [CrossRef]
- Beynon, R.J. The dynamics of the proteome: Strategies for measuring protein turnover on a proteome-wide scale. Brief. Funct. Genom. Proteom. 2005, 3, 382–390. [Google Scholar] [CrossRef]
- Chakravarti, B.; Mallik, B.; Chakravarti, D.N. Proteomics and Systems Biology: Application in Drug Discovery and Development. Methods Mol. Biol. 2010, 662, 3–28. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, V.; Molisso, D.; Aprile, A.M.; Maggio, A.; Rao, R. Systemin peptide application improves tomato salt stress tolerance and reveals common adaptation mechanisms to biotic and abiotic stress in plants. Environ. Exp. Bot. 2022, 199, 104865. [Google Scholar] [CrossRef]
- Thalhammer, A.; Hincha, D.K. A mechanistic model of COR15 protein function in plant freezing tolerance: Integration of structural and functional characteristics. Plant Signal. Behav. 2014, 9, e977722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Guo, X.; Zhang, Z.; Wang, A.; Zhu, J. Cold-regulated gene LeCOR413PM2 confers cold stress tolerance in tomato plants. Gene 2021, 764, 145097. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, A.; Pandian, S.K.; Ramesh, M. Transgenic indica rice cv. ADT 43 expressing a Δ1-pyrroline-5-carboxylate synthetase (P5CS) gene from Vigna aconitifolia demonstrates salt tolerance. Plant Cell Tissue Organ Cult. 2011, 107, 383–395. [Google Scholar] [CrossRef]
- Ogden, A.J.; Bhatt, J.J.; Brewer, H.M.; Kintigh, J.; Kariuki, S.M.; Rudrabhatla, S.; Adkins, J.N.; Curtis, W.R. Phloem Exudate Protein Profiles during Drought and Recovery Reveal Abiotic Stress Responses in Tomato Vasculature. Int. J. Mol. Sci. 2020, 21, 4461. [Google Scholar] [CrossRef]
- Klay, I.; Pirrello, J.; Riahi, L.; Bernadac, A.; Cherif, A.; Bouzayen, M.; Bouzid, S. Ethylene Response Factor Sl-ERF.B.3 Is Responsive to Abiotic Stresses and Mediates Salt and Cold Stress Response Regulation in Tomato. Sci. World J. 2014, 2014, 167681. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Wei, J.; Pan, Y.; Su, C.; Zhang, X. A tomato proline-, lysine-, and glutamic-rich type gene SpPKE1 positively regulates drought stress tolerance. Biochem. Biophys. Res. Commun. 2018, 499, 777–782. [Google Scholar] [CrossRef]
- Olvera-Carrillo, Y.; Reyes, J.L.; Covarrubias, A.A. Late embryogenesis abundant proteins: Versatile Players in the Plant Adaptation to Water Limiting Environments. Plant Signal. Behav. 2011, 6, 586–589. [Google Scholar] [CrossRef]
- Muñoz-Mayor, A.; Pineda, B.; Garcia-Abellán, J.O.; Antón, T.; Garcia-Sogo, B.; Sánchez-Bel, P.; Flores, F.B.; Atarés, A.; Angosto, T.; Pintor-Toro, J.A.; et al. Overexpression of dehydrin tas14 gene improves the osmotic stress imposed by drought and salinity in tomato. J. Plant Physiol. 2012, 169, 459–468. [Google Scholar] [CrossRef]
- Park, A.R.; Kim, J.; Kim, B.; Ha, A.; Son, J.-Y.; Song, C.W.; Song, H.; Kim, J.-C. Exogenous Bio-Based 2,3-Butanediols Enhanced Abiotic Stress Tolerance of Tomato and Turfgrass under Drought or Chilling Stress. J. Microbiol. Biotechnol. 2022, 32, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Francesca, S.; Najai, S.; Zhou, R.; Decros, G.; Cassan, C.; Delmas, F.; Ottosen, C.-O.; Barone, A.; Rigano, M.M. Phenotyping to dissect the biostimulant action of a protein hydrolysate in tomato plants under combined abiotic stress. Plant Physiol. Biochem. 2022, 179, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Qiao, Z.; Li, Y.; Yang, Z.; Wang, C.; Zhang, Y.; Liu, L.; Wang, B. RING Zinc Finger Proteins in Plant Abiotic Stress Tolerance. Front. Plant Sci. 2022, 13, 877011. [Google Scholar] [CrossRef]
- Lohani, N.; Singh, M.B.; Bhalla, P.L. Biological Parts for Engineering Abiotic Stress Tolerance in Plants. BioDes. Res. 2022, 2022, 9819314. [Google Scholar] [CrossRef]
- Hu, D.-G.; Wang, S.-H.; Luo, H.; Ma, Q.-J.; Yao, Y.-X.; You, C.-X.; Hao, Y.-J. Overexpression of MdVHA-B, a V-ATPase gene from apple, confers tolerance to drought in transgenic tomato. Sci. Hortic. 2012, 145, 94–101. [Google Scholar] [CrossRef]
- Zhao, C.; Shono, M.; Sun, A.; Yi, S.; Li, M.; Liu, J. Constitutive expression of an endoplasmic reticulum small heat shock protein alleviates endoplasmic reticulum stress in transgenic tomato. J. Plant Physiol. 2007, 164, 835–841. [Google Scholar] [CrossRef]
- Macel, M.; van Dam, N.; Keurentjes, J. Metabolomics: The chemistry between ecology and genetics. Mol. Ecol. Resour. 2010, 10, 583–593. [Google Scholar] [CrossRef]
- Ribbenstedt, A.; Ziarrusta, H.; Benskin, J.P. Development, characterization and comparisons of targeted and non-targeted metabolomics methods. PLoS ONE 2018, 13, e0207082. [Google Scholar] [CrossRef]
- Krauss, M.; Singer, H.; Hollender, J. LC–high resolution MS in environmental analysis: From target screening to the identification of unknowns. Anal. Bioanal. Chem. 2010, 397, 943–951. [Google Scholar] [CrossRef]
- Flamini, R.; De Rosso, M.; De Marchi, F.; Vedova, A.D.; Panighel, A.; Gardiman, M.; Maoz, I.; Bavaresco, L. An innovative approach to grape metabolomics: Stilbene profiling by suspect screening analysis. Metabolomics 2013, 9, 1243–1253. [Google Scholar] [CrossRef]
- Zheng, F.; Zhao, X.; Zeng, Z.; Wang, L.; Lv, W.; Wang, Q.; Xu, G. Development of a plasma pseudotargeted metabolomics method based on ultra-high-performance liquid chromatography–mass spectrometry. Nat. Protoc. 2020, 15, 2519–2537. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B.; Assmann, S.M.; Chen, S. Plant single-cell and single-cell-type metabolomics. Trends Plant Sci. 2014, 19, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Abouleila, Y.; Shimizu, Y.; Hiyama, E.; Emara, S.; Mashaghi, A.; Hankemeier, T. Single-cell metabolomics by mass spectrometry: Advances, challenges, and future applications. TrAC Trends Anal. Chem. 2019, 120, 115436. [Google Scholar] [CrossRef]
- Wada, H.; Hatakeyama, Y.; Onda, Y.; Nonami, H.; Nakashima, T.; Erra-Balsells, R.; Morita, S.; Hiraoka, K.; Tanaka, F.; Nakano, H. Multiple strategies for heat adaptation to prevent chalkiness in the rice endosperm. J. Exp. Bot. 2019, 70, 1299–1311. [Google Scholar] [CrossRef]
- Nakashima, T.; Wada, H.; Morita, S.; Erra-Balsells, R.; Hiraoka, K.; Nonami, H. Single-Cell Metabolite Profiling of Stalk and Glandular Cells of Intact Trichomes with Internal Electrode Capillary Pressure Probe Electrospray Ionization Mass Spectrometry. Anal. Chem. 2016, 88, 3049–3057. [Google Scholar] [CrossRef] [PubMed]
- Heyman, H.M.; Dubery, I.A. The potential of mass spectrometry imaging in plant metabolomics: A review. Phytochem. Rev. 2016, 15, 297–316. [Google Scholar] [CrossRef]
- Kulkarni, P.; Wilschut, R.A.; Verhoeven, K.J.F.; van der Putten, W.H.; Garbeva, P. LAESI mass spectrometry imaging as a tool to differentiate the root metabolome of native and range-expanding plant species. Planta 2018, 248, 1515–1523. [Google Scholar] [CrossRef]
- Misra, B.B. New software tools, databases, and resources in metabolomics: Updates from 2020. Metabolomics 2021, 17, 49. [Google Scholar] [CrossRef]
- Obata, T.; Fernie, A.R. The use of metabolomics to dissect plant responses to abiotic stresses. Cell. Mol. Life Sci. 2012, 69, 3225–3243. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Feng, Z.; Ji, S.; Ping, J.; Cui, D. Recent advances in metabolomics for studying heavy metal stress in plants. TrAC Trends Anal. Chem. 2021, 143, 116402. [Google Scholar] [CrossRef]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental biotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 493–502. [Google Scholar] [CrossRef]
- Borges, C.V.; Minatel, I.O.; Gomez-Gomez, H.A.; Lima, G.P.P. Medicinal Plants: Influence of Environmental Factors on the Content of Secondary Metabolites. In Medicinal Plants and Environmental Challenges; Springer: Cham, Switzerland, 2017; pp. 259–277. [Google Scholar]
- Sharma, A. Gene Expression Analysis in Medicinal Plants Under Abiotic Stress Conditions. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 407–414. [Google Scholar]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Hurry, V.; Hüner, N.P. Interaction of glycine betaine and plant hormones: Protection of the photosynthetic apparatus during abiotic stress. In Photosynthesis: Structures, Mechanisms, and Applications; Springer: Cham, Switzerland, 2017; pp. 185–202. [Google Scholar]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Fiesel, P.D.; Parks, H.M.; Last, R.L.; Barry, C.S. Fruity, sticky, stinky, spicy, bitter, addictive, and deadly: Evolutionary signatures of metabolic complexity in the Solanaceae. Nat. Prod. Rep. 2022, 39, 1438–1464. [Google Scholar] [CrossRef]
- Pigatto, A.G.; Blanco, C.C.; Mentz, L.A.; Soares, G.L. Tropane alkaloids and calystegines as chemotaxonomic markers in the Solanaceae. An. Acad. Bras. Cienc. 2015, 87, 2139–2149. [Google Scholar] [CrossRef]
- Richter, U.; Sonnewald, U.; Dräger, B. Calystegines in potatoes with genetically engineered carbohydrate metabolism. J. Exp. Bot. 2007, 58, 1603–1615. [Google Scholar] [CrossRef]
- Goldmann, A.; Message, B.; Tepfer, D.; Molyneux, R.J.; Duclos, O.; Boyer, F.-D.; Pan, Y.T.; Elbein, A.D. Biological Activities of the Nortropane Alkaloid, Calystegine B2, and Analogs: Structure–Function Relationships. J. Nat. Prod. 1996, 59, 1137–1142. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Fan, P.; Leong, B.J.; Last, R.L. Tip of the trichome: Evolution of acylsugar metabolic diversity in Solanaceae. Curr. Opin. Plant Biol. 2019, 49, 8–16. [Google Scholar] [CrossRef]
- Ito, S.-I.; Ihara, T.; Tamura, H.; Tanaka, S.; Ikeda, T.; Kajihara, H.; Dissanayake, C.; Abdel-Motaal, F.F.; El-Sayed, M.A. α-Tomatine, the major saponin in tomato, induces programmed cell death mediated by reactive oxygen species in the fungal pathogen Fusarium oxysporum. FEBS Lett. 2007, 581, 3217–3222. [Google Scholar] [CrossRef]
- Friedman, M. Tomato Glycoalkaloids: Role in the Plant and in the Diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef] [PubMed]
- Sandrock, R.W.; VanEtten, H.D. The relevance of tomatinase activity in pathogens of tomato: Disruption of the β2-tomatinase gene in Colletotrichum coccodes and Septoria lycopersici and heterologous expression of the Septoria lycopersiciβ2-tomatinase in Nectria haematococca, a pathogen of tomato fruit. Physiol. Mol. Plant Pathol. 2001, 58, 159–171. [Google Scholar] [CrossRef]
- Mhlongo, M.; Piater, L.; Steenkamp, P.; Labuschagne, N.; Dubery, I. Metabolomic Evaluation of Tissue-Specific Defense Responses in Tomato Plants Modulated by PGPR-Priming against Phytophthora capsici Infection. Plants 2021, 10, 1530. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, P.; Vinale, F.; Woo, S.L.; Pascale, A.; Lorito, M.; Piccolo, A. Metabolomics by Proton High-Resolution Magic-Angle-Spinning Nuclear Magnetic Resonance of Tomato Plants Treated with Two Secondary Metabolites Isolated from Trichoderma. J. Agric. Food Chem. 2016, 64, 3538–3545. [Google Scholar] [CrossRef]
- Camañes, G.; Scalschi, L.; Vicedo, B.; González-Bosch, C.; García-Agustín, P. An untargeted global metabolomic analysis reveals the biochemical changes underlying basal resistance and priming in Solanum lycopersicum, and identifies 1-methyltryptophan as a metabolite involved in plant responses to Botrytis cinerea and Pseudomonas syringae. Plant J. 2015, 84, 125–139. [Google Scholar] [CrossRef]
- Akram, W.; Anjum, T.; Ali, B. Phenylacetic Acid Is ISR Determinant Produced by Bacillus fortis IAGS162, Which Involves Extensive Re-modulation in Metabolomics of Tomato to Protect against Fusarium Wilt. Front. Plant Sci. 2016, 7, 498. [Google Scholar] [CrossRef]
- Laura, A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla, E. Phenolic Compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Woodhead Publishing: Sawston, UK, 2019; pp. 253–271. [Google Scholar]
- Kumar, D. Chapter 4—Defense Strategies in Plants against Insect Herbivores. In Advances in Agricultural Entomology; Kosmos Publishers: Stuttgart, Germany, 2019; Volume 7, pp. 67–119. [Google Scholar]
- Gerlin, L.; Escourrou, A.; Cassan, C.; Macia, F.M.; Peeters, N.; Genin, S.; Baroukh, C. Unravelling physiological signatures of tomato bacterial wilt and xylem metabolites exploited by Ralstonia solanacearum. Environ. Microbiol. 2021, 23, 5962–5978. [Google Scholar] [CrossRef]
- Zeiss, D.R.; Mhlongo, M.I.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A. Metabolomic Profiling of the Host Response of Tomato (Solanum lycopersicum) Following Infection by Ralstonia solanacearum. Int. J. Mol. Sci. 2019, 20, 3945. [Google Scholar] [CrossRef]
- Mas, F.; Vereijssen, J.; Suckling, D.M. Influence of the Pathogen Candidatus Liberibacter solanacearum on Tomato Host Plant Volatiles and Psyllid Vector Settlement. J. Chem. Ecol. 2014, 40, 1197–1202. [Google Scholar] [CrossRef]
- López-Gresa, M.P.; Lisón, P.; Kim, H.K.; Choi, Y.H.; Verpoorte, R.; Rodrigo, I.; Conejero, V.; Bellés, J.M. Metabolic fingerprinting of Tomato Mosaic Virus infected Solanum lycopersicum. J. Plant Physiol. 2012, 169, 1586–1596. [Google Scholar] [CrossRef]
- Ito, S.-I.; Eto, T.; Tanaka, S.; Yamauchi, N.; Takahara, H.; Ikeda, T. Tomatidine and lycotetraose, hydrolysis products of α-tomatine by Fusarium oxysporum Tomatinase, suppress induced defense responses in tomato cells. FEBS Lett. 2004, 571, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Ökmen, B.; Etalo, D.W.; Joosten, M.H.A.J.; Bouwmeester, H.J.; Vos, R.C.H.; Collemare, J.; Wit, P.J.G.M. Detoxification of α-tomatine by Cladosporium fulvum is required for full virulence on tomato. New Phytol. 2013, 198, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Roumani, M.; Le Bot, J.; Boisbrun, M.; Magot, F.; Péré, A.; Robin, C.; Hilliou, F.; Larbat, R. Transcriptomics and Metabolomics Analyses Reveal High Induction of the Phenolamide Pathway in Tomato Plants Attacked by the Leafminer Tuta absoluta. Metabolites 2022, 12, 484. [Google Scholar] [CrossRef] [PubMed]
- de Falco, B.; Manzo, D.; Incerti, G.; Garonna, A.P.; Ercolano, M.; Lanzotti, V. Metabolomics approach based on NMR spectroscopy and multivariate data analysis to explore the interaction between the leafminer Tuta absoluta and tomato (Solanum lycopersicum). Phytochem. Anal. 2019, 30, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-M.; Li, X.-W.; He, T.-J.; Li, P.-J.; Liu, Y.; Zhou, S.-X.; Wu, Q.-C.; Chen, T.-T.; Lu, Y.-B.; Hou, Y.-M. Comparative biochemical and transcriptome analyses in tomato and eggplant reveal their differential responses to Tuta absoluta infestation. Genomics 2021, 113, 2108–2121. [Google Scholar] [CrossRef] [PubMed]
- Papantoniou, D.; Vergara, F.; Weinhold, A.; Quijano, T.; Khakimov, B.; Pattison, D.I.; Bak, S.; van Dam, N.M.; Martínez-Medina, A. Cascading Effects of Root Microbial Symbiosis on the Development and Metabolome of the Insect Herbivore Manduca sexta L. Metabolites 2021, 11, 731. [Google Scholar] [CrossRef]
- Białczyk, J.; Lechowski, Z.; Libik, A. The protective action of tannins against glasshouse whitefly in tomato seedlings. J. Agric. Sci. 1999, 133, 197–201. [Google Scholar] [CrossRef]
- Ament, K.; Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Jasmonic Acid Is a Key Regulator of Spider Mite-Induced Volatile Terpenoid and Methyl Salicylate Emission in Tomato. Plant Physiol. 2004, 135, 2025–2037. [Google Scholar] [CrossRef]
- Degenhardt, D.C.; Refi-Hind, S.; Stratmann, J.W.; Lincoln, D.E. Systemin and jasmonic acid regulate constitutive and herbivore-induced systemic volatile emissions in tomato, Solanum lycopersicum. Phytochemistry 2010, 71, 2024–2037. [Google Scholar] [CrossRef]
- Formenti, L.; Rasmann, S. Mycorrhizal Fungi Enhance Resistance to Herbivores in Tomato Plants with Reduced Jasmonic Acid Production. Agronomy 2019, 9, 131. [Google Scholar] [CrossRef]
- A Ryan, C. The systemin signaling pathway: Differential activation of plant defensive genes. Biochim. Biophys. Acta 2000, 1477, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Paré, P.W.; Tumlinson, J.H. Plant Volatiles as a Defense against Insect Herbivores. Plant Physiol. 1999, 121, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, G.; Tian, L.; Peng, Z.; Xie, W.; Wu, Q.; Wang, S.; Zhou, X.; Zhang, Y. The Salicylic Acid-Mediated Release of Plant Volatiles Affects the Host Choice of Bemisia tabaci. Int. J. Mol. Sci. 2016, 17, 1048. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Peng, Z.; Tong, H.; Yang, F.; Xing, G.; Wang, L.; Zheng, J.; Zhang, Y.; Su, Q. Tomato Plant Flavonoids Increase Whitefly Resistance and Reduce Spread of Tomato yellow leaf curl virus. J. Econ. Èntomol. 2019, 112, 2790–2796. [Google Scholar] [CrossRef]
- Vos, C.; Claerhout, S.; Mkandawire, R.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi reduce root-knot nematode penetration through altered root exudation of their host. Plant Soil 2012, 354, 335–345. [Google Scholar] [CrossRef]
- Dong, L.; Li, X.; Huang, L.; Gao, Y.; Zhong, L.; Zheng, Y.; Zuo, Y. Lauric acid in crown daisy root exudate potently regulates root-knot nematode chemotaxis and disrupts Mi-flp-18 expression to block infection. J. Exp. Bot. 2014, 65, 131–141. [Google Scholar] [CrossRef]
- Gómez, S.; Ferrieri, R.A.; Schueller, M.; Orians, C.M. Methyl jasmonate elicits rapid changes in carbon and nitrogen dynamics in tomato. New Phytol. 2010, 188, 835–844. [Google Scholar] [CrossRef]
- Gómez, S.; Steinbrenner, A.D.; Osorio, S.; Schueller, M.; Ferrieri, R.A.; Fernie, A.R.; Orians, C.M. From shoots to roots: Transport and metabolic changes in tomato after simulated feeding by a specialist lepidopteran. èntomol. Exp. Appl. 2012, 144, 101–111. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Gómez, S.; Osorio, S.; Fernie, A.R.; Orians, C.M. Herbivore-Induced Changes in Tomato (Solanum lycopersicum) Primary Metabolism: A Whole Plant Perspective. J. Chem. Ecol. 2011, 37, 1294–1303. [Google Scholar] [CrossRef]
- Nascimento, V.L.; Pereira, A.M.; Siqueira, J.A.; Pereira, A.S.; Silva, V.F.; Costa, L.C.; Ribeiro, D.M.; Zsögön, A.; Nunes-Nesi, A.; Araújo, W.L. Exogenous ethylene reduces growth via alterations in central metabolism and cell wall composition in tomato (Solanum lycopersicum). J. Plant Physiol. 2021, 263, 153460. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef]
- Shi, Y.; Zhao, Y.; Yao, Q.; Liu, F.; Li, X.; Jin, X.; Zhang, Y.; Ahammed, G.J. Comparative Physiological and Transcriptomic Analyses Reveal Mechanisms of Exogenous Spermidine-Induced Tolerance to Low-Iron Stress in Solanum lycopersicum L. Antioxidants 2022, 11, 1260. [Google Scholar] [CrossRef] [PubMed]
- Shakir, S.K.; Irfan, S.; Akhtar, B.; Rehman, S.U.; Daud, M.K.; Taimur, N.; Azizullah, A. Pesticide-induced oxidative stress and antioxidant responses in tomato (Solanum lycopersicum) seedlings. Ecotoxicology 2018, 27, 919–935. [Google Scholar] [CrossRef] [PubMed]
- Hatamleh, A.A.; Danish, M.; Al-Dosary, M.A.; El-Zaidy, M.; Ali, S. Physiological and oxidative stress responses of Solanum lycopersicum (L.) (tomato) when exposed to different chemical pesticides. RSC Adv. 2022, 12, 7237–7252. [Google Scholar] [CrossRef]
- Luna, E.; Flandin, A.; Cassan, C.; Prigent, S.; Chevanne, C.; Kadiri, C.F.; Gibon, Y.; Pétriacq, P. Metabolomics to Exploit the Primed Immune System of Tomato Fruit. Metabolites 2020, 10, 96. [Google Scholar] [CrossRef]
- Ji, X.; Li, J.; Meng, Z.; Zhang, S.; Dong, B.; Qiao, K. Synergistic Effect of Combined Application of a New Fungicide Fluopimomide with a Biocontrol Agent Bacillus methylotrophicus TA-1 for Management of Gray Mold in Tomato. Plant Dis. 2019, 103, 1991–1997. [Google Scholar] [CrossRef]
- Chen, J.; Li, Q.X.; Song, B. Chemical Nematicides: Recent Research Progress and Outlook. J. Agric. Food Chem. 2020, 68, 12175–12188. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, H.; Zong, X.; Wang, X.; Wu, C. Effects of soil treated fungicide fluopimomide on tomato (Solanum lycopersicum L.) disease control and plant growth. Open Life Sci. 2022, 17, 800–810. [Google Scholar] [CrossRef]
- Sharaf, E.F.; A Farrag, A. Induced resistance in tomato plants by IAA against Fusarium oxysporum lycopersici. Pol. J. Microbiol. 2004, 53, 111–116. [Google Scholar]
- Pretali, L.; Bernardo, L.; Butterfield, T.S.; Trevisan, M.; Lucini, L. Botanical and biological pesticides elicit a similar Induced Systemic Response in tomato (Solanum lycopersicum) secondary metabolism. Phytochemistry 2016, 130, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Fei, Z.; Chen, Y.-R.; Zheng, Y.; Huang, M.; Vrebalov, J.; McQuinn, R.; Gapper, N.; Liu, B.; Xiang, J.; et al. Single-base resolution methylomes of tomato fruit development reveal epigenome modifications associated with ripening. Nat. Biotechnol. 2013, 31, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: The mark and its mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tang, K.; Datsenka, T.U.; Liu, W.; Lv, S.; Lang, Z.; Wang, X.; Gao, J.; Wang, W.; Nie, W.; et al. Critical function of DNA methyltransferase 1 in tomato development and regulation of the DNA methylome and transcriptome. J. Integr. Plant Biol. 2019, 61, 1224–1242. [Google Scholar] [CrossRef]
- Osorio, S.; Alba, R.; Damasceno, C.M.; Lopez-Casado, G.; Lohse, M.; Zanor, M.I.; Tohge, T.; Usadel, B.; Rose, J.K.; Fei, Z.; et al. Systems Biology of Tomato Fruit Development: Combined Transcript, Protein, and Metabolite Analysis of Tomato Transcription Factor (nor, rin) and Ethylene Receptor (Nr) Mutants Reveals Novel Regulatory Interactions. Plant Physiol. 2011, 157, 405–425. [Google Scholar] [CrossRef]
- Manning, K.; Tör, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat. Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Quadrana, L.; Almeida, J.; Asís, R.; Duffy, T.; Dominguez, P.G.; Bermúdez, L.; Conti, G.; Da Silva, J.V.C.; Peralta, I.E.; Colot, V.; et al. Natural occurring epialleles determine vitamin E accumulation in tomato fruits. Nat. Commun. 2014, 5, 3027. [Google Scholar] [CrossRef]
- Lang, Z.; Wang, Y.; Tang, K.; Tang, D.; Datsenka, T.; Cheng, J.; Zhang, Y.; Handa, A.K.; Zhu, J.-K. Critical roles of DNA demethylation in the activation of ripening-induced genes and inhibition of ripening-repressed genes in tomato fruit. Proc. Natl. Acad. Sci. USA 2017, 114, E4511–E4519. [Google Scholar] [CrossRef]
- Zhang, B.; Tieman, D.M.; Jiao, C.; Xu, Y.; Chen, K.; Fei, Z.; Giovannoni, J.J.; Klee, H.J. Chilling-induced tomato flavor loss is associated with altered volatile synthesis and transient changes in DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, 12580–12585. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Ding, X.; Liu, X.; Jiang, G.; Li, Z.; Song, Y.; Zhang, D.; Jiang, Y.; Duan, X. SlJMJ7 orchestrates tomato fruit ripening via crosstalk between H3K4me3 and DML2-mediated DNA demethylation. New Phytol. 2022, 233, 1202–1219. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, G.; Liu, X.; Ding, X.; Zhang, D.; Wang, X.; Zhou, Y.; Yan, H.; Li, T.; Wu, K.; et al. Histone demethylase SlJMJ6 promotes fruit ripening by removing H3K27 methylation of ripening-related genes in tomato. New Phytol. 2020, 227, 1138–1156. [Google Scholar] [CrossRef]
- Bouyer, D.; Roudier, F.; Heese, M.; Andersen, E.D.; Gey, D.; Nowack, M.K.; Goodrich, J.; Renou, J.P.; Grini, P.E.; Colot, V.; et al. Polycomb Repressive Complex 2 Controls the Embryo-to-Seedling Phase Transition. PLoS Genet. 2011, 7, e1002014. [Google Scholar] [CrossRef] [PubMed]
- Lü, P.; Yu, S.; Zhu, N.; Chen, Y.-R.; Zhou, B.; Pan, Y.; Tzeng, D.; Fabi, J.P.; Argyris, J.; Garcia-Mas, J.; et al. Genome encode analyses reveal the basis of convergent evolution of fleshy fruit ripening. Nat. Plants 2018, 4, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Kit, A.H.; Boureau, L.; Stammitti-Bert, L.; Rolin, D.; Teyssier, E.; Gallusci, P. Functional analysis of SlEZ1 a tomato Enhancer of zeste (E(z)) gene demonstrates a role in flower development. Plant Mol. Biol. 2010, 74, 201–213. [Google Scholar] [CrossRef]
- Boureau, L.; How-Kit, A.; Teyssier, E.; Drevensek, S.; Rainieri, M.; Joubès, J.; Stammitti, L.; Pribat, A.; Bowler, C.; Hong, Y.; et al. A CURLY LEAF homologue controls both vegetative and reproductive development of tomato plants. Plant Mol. Biol. 2016, 90, 485–501. [Google Scholar] [CrossRef]
- Xiao, L.; Liang, H.; Jiang, G.; Ding, X.; Liu, X.; Sun, J.; Jiang, Y.; Song, L.; Duan, X. Proteome-wide identification of non-histone lysine methylation in tomato during fruit ripening. J. Adv. Res. 2022, 42, 177–188. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, A.; Wei, H.; Hao, P.; Zhang, Q.; Tian, M.; Yang, X.; Cheng, S.; Fu, X.; Ma, L.; et al. Genome-wide identification and expression patterns analysis of the RPD3/HDA1 gene family in cotton. BMC Genom. 2020, 21, 643. [Google Scholar] [CrossRef]
- Guo, J.-E.; Hu, Z.; Yu, X.; Li, A.; Li, F.; Wang, Y.; Tian, S.; Chen, G. A histone deacetylase gene, SlHDA3, acts as a negative regulator of fruit ripening and carotenoid accumulation. Plant Cell Rep. 2018, 37, 125–135. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, J.; Zhang, J.; Wu, P.-Y.; Yang, S.; Wu, K. Identification and characterization of histone deacetylases in tomato (Solanum lycopersicum). Front. Plant Sci. 2014, 5, 760. [Google Scholar] [CrossRef]
- Yu, X.; Gao, Q.; Chen, G.; Guo, J.-E.; Guo, X.; Tang, B.; Hu, Z. SlHDA5, a Tomato Histone Deacetylase Gene, Is Involved in Responding to Salt, Drought, and ABA. Plant Mol. Biol. Rep. 2018, 36, 36–44. [Google Scholar] [CrossRef]
- Guo, J.-E.; Hu, Z.; Li, F.; Zhang, L.; Yu, X.; Tang, B.; Chen, G. Silencing of histone deacetylase SlHDT3 delays fruit ripening and suppresses carotenoid accumulation in tomato. Plant Sci. 2017, 265, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-E. Histone deacetylase gene SlHDT1 regulates tomato fruit ripening by affecting carotenoid accumulation and ethylene biosynthesis. Plant Sci. 2022, 318, 111235. [Google Scholar] [CrossRef] [PubMed]
- Cigliano, R.A.; Sanseverino, W.; Cremona, G.; Ercolano, M.R.; Conicella, C.; Consiglio, F.M. Genome-wide analysis of histone modifiers in tomato: Gaining an insight into their developmental roles. BMC Genom. 2013, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Riaz, A.; Chachar, S.; Ding, Y.; Du, H.; Gu, X. Epigenetic Modifications of mRNA and DNA in Plants. Mol. Plant 2020, 13, 14–30. [Google Scholar] [CrossRef]
- Shao, Y.; Wong, C.E.; Shen, L.; Yu, H. N6-methyladenosine modification underlies messenger RNA metabolism and plant development. Curr. Opin. Plant Biol. 2021, 63, 102047. [Google Scholar] [CrossRef]
- Yang, D.; Xu, H.; Liu, Y.; Li, M.; Ali, M.; Xu, X.; Lu, G. RNA N6-Methyladenosine Responds to Low-Temperature Stress in Tomato Anthers. Front. Plant Sci. 2021, 12, 687826. [Google Scholar] [CrossRef]
- Hu, J.; Cai, J.; Umme, A.; Chen, Y.; Xu, T.; Kang, H. Unique features of mRNA m6A methylomes during expansion of tomato (Solanum lycopersicum) fruits. Plant Physiol. 2022, 188, 2215–2227. [Google Scholar] [CrossRef]
- Zhou, L.; Tian, S.; Qin, G. RNA methylomes reveal the m6A-mediated regulation of DNA demethylase gene SlDML2 in tomato fruit ripening. Genome Biol. 2019, 20, 1. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef]
- Oliver, M.J.; Jain, R.; Balbuena, T.S.; Agrawal, G.; Gasulla, F.; Thelen, J.J. Proteome analysis of leaves of the desiccation-tolerant grass, Sporobolus stapfianus, in response to dehydration. Phytochemistry 2011, 72, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q.; Dai, S. Drought-Responsive Mechanisms in Plant Leaves Revealed by Proteomics. Int. J. Mol. Sci. 2016, 17, 1706. [Google Scholar] [CrossRef] [PubMed]
- Loukehaich, R.; Wang, T.; Ouyang, B.; Ziaf, K.; Li, H.; Zhang, J.; Lu, Y.; Ye, Z. SpUSP, an annexin-interacting universal stress protein, enhances drought tolerance in tomato. J. Exp. Bot. 2012, 63, 5593–5606. [Google Scholar] [CrossRef] [PubMed]
- Irfan, M.; Kumar, P.; Ahmad, I.; Datta, A. Unraveling the role of tomato Bcl-2-associated athanogene (BAG) proteins during abiotic stress response and fruit ripening. Sci. Rep. 2021, 11, 21734. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Ouyang, Z.; Zhang, Y.; Li, X.; Hong, Y.; Huang, L.; Liu, S.; Zhang, H.; Li, D.; Song, F. Tomato NAC Transcription Factor SlSRN1 Positively Regulates Defense Response against Biotic Stress but Negatively Regulates Abiotic Stress Response. PLoS ONE 2014, 9, e102067. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, G.; Zhang, J.; Zhang, Y.; Xie, Q.; Zhao, Z.; Pan, Y.; Hu, Z. The abiotic stress-responsive NAC-type transcription factor SlNAC4 regulates salt and drought tolerance and stress-related genes in tomato (Solanum lycopersicum). Plant Cell Rep. 2014, 33, 1851–1863. [Google Scholar] [CrossRef]
- Hendelman, A.; Stav, R.; Zemach, H.; Arazi, T. The tomato NAC transcription factor SlNAM2 is involved in flower-boundary morphogenesis. J. Exp. Bot. 2013, 64, 5497–5507. [Google Scholar] [CrossRef]
- Ma, N.-N.; Zuo, Y.-Q.; Liang, X.-Q.; Yin, B.; Wang, G.-D.; Meng, Q.-W. The multiple stress-responsive transcription factor SlNAC1 improves the chilling tolerance of tomato. Physiol. Plant. 2013, 149, 474–486. [Google Scholar] [CrossRef]
- Selth, L.A.; Dogra, S.C.; Rasheed, M.S.; Healy, H.; Randles, J.W.; Rezaian, M.A. A NAC Domain Protein Interacts with Tomato leaf curl virus Replication Accessory Protein and Enhances Viral Replication. Plant Cell 2005, 17, 311–325. [Google Scholar] [CrossRef]
- Berger, Y.; Harpaz-Saad, S.; Brand, A.; Melnik, H.; Sirding, N.; Alvarez, J.P.; Zinder, M.; Samach, A.; Eshed, Y.; Ori, N. The NAC-domain transcription factor GOBLET specifies leaflet boundaries in compound tomato leaves. Development 2009, 136, 823–832. [Google Scholar] [CrossRef]
- Ming, N.; Ma, N.; Jiao, B.; Lv, W.; Meng, Q. Genome Wide Identification of C2H2-Type Zinc Finger Proteins of Tomato and Expression Analysis Under Different Abiotic Stresses. Plant Mol. Biol. Rep. 2020, 38, 75–94. [Google Scholar] [CrossRef]




| S. No. | Gene/Origin | Function | Expression/Regulation | Results | References |
|---|---|---|---|---|---|
| 1 | SDG34 | Response to stress | Expression of negative stress response regulators and transcriptional repressors | Improvement of stress and pathogen tolerance | [51] |
| 2 | SlGRAS10 | Increasing osmotic potential, flavonoid production, and the ROS scavenging mechanism to increase abiotic stress tolerance | Downregulation | [48] | |
| 3 | BEL1-like genes | Numerous biological processes in plants are regulated by transcription factors, which are members of the superfamily of three-amino-loop-extension (TALE) proteins | Displayed various tissue-specific expression patterns and reacted to heat, cold, and drought stress | Plant growth and abiotic stress response | [59] |
| 4 | SlAIM1 | Salt and oxidative stress tolerance | Salt and oxidative stress tolerance is increased by SlAIM1 overexpression, but these two abiotic stimuli are made more sensitive by SlAIM1 silencing | Resistance to abiotic stress | [52,60] |
| 5 | TFs, s Cycling Dof Factor AtCDF3, AtDREB1a, NAC transcription factor JUNGBRUNNEN1 (AtJUB1) and AP2/ERF-like transcription factor CcHRD | Increases abiotic stress tolerance of tomatoes, including cold, salt, and drought stress | Overexpression | Stress tolerance | [52,61] |
| 6 | SlMBP8, SlHB2, SlAGO4A | Tolerance to salt, drought stress | Overexpression | Tolerance to salt, drought stress | [62] |
| 7 | INVINH1 | Tolerance to cold stress | Tolerance to cold stress | [63] | |
| 8 | SlMBP8 | more tolerance to drought and salt stress | Gene silencing | More tolerance to drought and salt stress | [62] |
| 9 | SI PL | Resistance to pathogenic Botrytis cinerea and prolonged shelf life | Resistance to pathogenic Botrytis cinerea | [64] | |
| 10 | SlbZIP1 | Salt and drought stress tolerance | Expression | Salt and drought stress tolerance | [3] |
| 11 | SlMAPK3, SlMPK7 i | Resistance to chilling, cadmium, and drought stresses | Overexpression | Resistance to chilling, cadmium, and drought stresses | [65] |
| 12 | PpSnRK1α) | Accelerated metabolism of reactive oxygen species via upregulating antioxidase gene expression and antioxidant enzyme activity | Overexpression | Salt resistance | [66] |
| 13 | SlBZR1D | Salt tolerance and upregulated the expression of multiple stress-related genes | Overexpression and upregulation | Salt tolerance | [33] |
| 14 | SlNL33 | Ascorbate accumulation | suppressed expression | Stress tolerance | [67] |
| 15 | SlHY5 | Cold tolerance | Overexpression | [68] | |
| 16 | MdSWEET17 | Drought stress response and the regulation of fructose. | Expressed in tomatoes | Drought stress | [69] |
| 17 | SiDHN | Saussurea involucrata dehydrin gene overexpression | Overexpression | Cold and drought tolerance | [70] |
| 16 | SlHSP17.7 | Controlling Calcium Signaling and Phosphatidylglycerol Metabolism | Overexpression | Cold tolerance | [71] |
| 17 | SlABIG1 | Salt stress negative regulator gene | Knockout | Salt tolerance | [72] |
| 18 | Solyc03g020030 | Proteinase inhibitor-II | Gene silencing | Thermotolerance | [73] |
| 19 | SlDEAD23 and SlDEAD35 | Abiotic and biotic stress responses | Overexpression | Enhanced tolerance to salt and cold | [74] |
| 20 | SlGRAS10 | Improved the expression of superoxide dismutase, peroxidase, and catalase to lessen the impact of reactive oxygen species | Downregulation by RNA interference | Abiotic stress tolerance | [48] |
| 21 | SlLBD40 | A negative regulator of drought tolerance, it was implicated in JA signaling. | CRISPR/Cas9 targeted mutagenesis (knockout) | Drought tolerance | [75] |
| 22 | SlMAPK3i | Removing ROS buildup and increasing the expression of genes associated with ethylene signaling | Over-expression | Salt stress tolerance | [76] |
| S. No. | Protein | Function | Result | References |
|---|---|---|---|---|
| 1 | Systemin peptide | Sodium exclusion, antioxidant activity, protease inhibitor | Lower palatability for herbivores and high salt stress tolerance | [97] |
| 2 | COR15 protein | Folds and adheres to the chloroplast membranes to support leaf cells during freezing | Cold tolerance | [98,99] |
| 3 | P5CS protein | Involved in proline biosynthesis and inducible upon salt shock in drought-resistance | Drought and salt tolerance | [100,101] |
| 4 | Transcription factor of the ERF (ethylene-responsive factor) family | Reduces cell injury and enhances tolerance against cold stress | Cold, heat, and flood tolerance | [102] |
| 5 | SpPKE1 | Interact with an F-box protein associated with drought tolerance | Drought tolerance | [103] |
| 6 | LEA proteins | Prevents membrane leakage, membrane and protein stabilization | Water balance and ion sequestration maintenance | [104] |
| 7 | Dehydrins (DHNs) | Enhances tolerance to salinity and drought stress | Salinity and drought stress | [105] |
| 8 | 2,3-butanediol | Drought and chilling response | reduced harmful effects of abiotic stresses | [106] |
| 9 | E42 and LA3120 | Stimulates plant growth and reduces stresses | Better plant growth under water stress | [107] |
| 10 | RING zinc finger | Plant growth and reducing abiotic stresses | Reduced stresses on plant | [108] |
| 11 | Golden 2-Like | Plant development and reducing abiotic stress | Reduced drought stress tolerance by lowering SOD, peroxidase | [28] |
| 12 | Histidine kinase | Reduces abiotic stresses | Maintaining cellular Na+ homeostasis | [109] |
| 13 | SlbZIP1 | Reduces ABA-mediated stress | Reduced salt and drought stress | [33] |
| 14 | MdVHA-B | Better tolerance to drought stress | Increased tolerance | [110] |
| 15 | LeHSP21.5 | Improved tolerance to tunicamycin-ER stress inducer | Increased tolerance | [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naik, B.; Kumar, V.; Rizwanuddin, S.; Chauhan, M.; Choudhary, M.; Gupta, A.K.; Kumar, P.; Kumar, V.; Saris, P.E.J.; Rather, M.A.; et al. Genomics, Proteomics, and Metabolomics Approaches to Improve Abiotic Stress Tolerance in Tomato Plant. Int. J. Mol. Sci. 2023, 24, 3025. https://doi.org/10.3390/ijms24033025
Naik B, Kumar V, Rizwanuddin S, Chauhan M, Choudhary M, Gupta AK, Kumar P, Kumar V, Saris PEJ, Rather MA, et al. Genomics, Proteomics, and Metabolomics Approaches to Improve Abiotic Stress Tolerance in Tomato Plant. International Journal of Molecular Sciences. 2023; 24(3):3025. https://doi.org/10.3390/ijms24033025
Chicago/Turabian StyleNaik, Bindu, Vijay Kumar, Sheikh Rizwanuddin, Mansi Chauhan, Megha Choudhary, Arun Kumar Gupta, Pankaj Kumar, Vivek Kumar, Per Erik Joakim Saris, Muzamil Ahmad Rather, and et al. 2023. "Genomics, Proteomics, and Metabolomics Approaches to Improve Abiotic Stress Tolerance in Tomato Plant" International Journal of Molecular Sciences 24, no. 3: 3025. https://doi.org/10.3390/ijms24033025
APA StyleNaik, B., Kumar, V., Rizwanuddin, S., Chauhan, M., Choudhary, M., Gupta, A. K., Kumar, P., Kumar, V., Saris, P. E. J., Rather, M. A., Bhuyan, S., Neog, P. R., Mishra, S., & Rustagi, S. (2023). Genomics, Proteomics, and Metabolomics Approaches to Improve Abiotic Stress Tolerance in Tomato Plant. International Journal of Molecular Sciences, 24(3), 3025. https://doi.org/10.3390/ijms24033025