
Genome-Wide Identification of the SQUAMOSA Promoter-Binding Protein-like (SPL) Transcription Factor Family in Sweet Cherry Fruit
 ,
,
Abstract
1. Introduction
2. Results
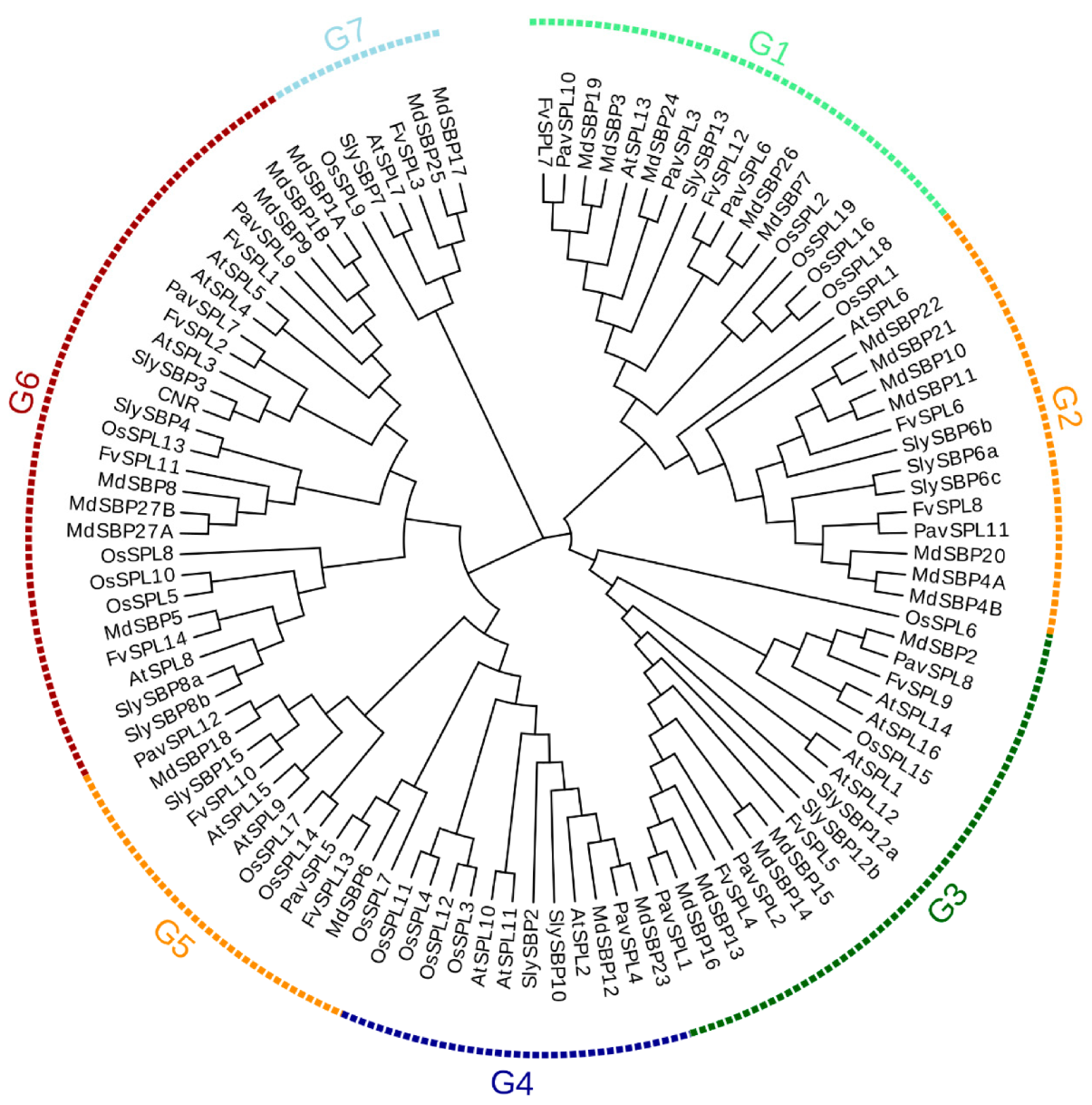
2.1. Identification and Classification of SPL Genes in Sweet Cherry
2.2. Chromosome Distribution and Synteny Analysis of the PavSPL Family
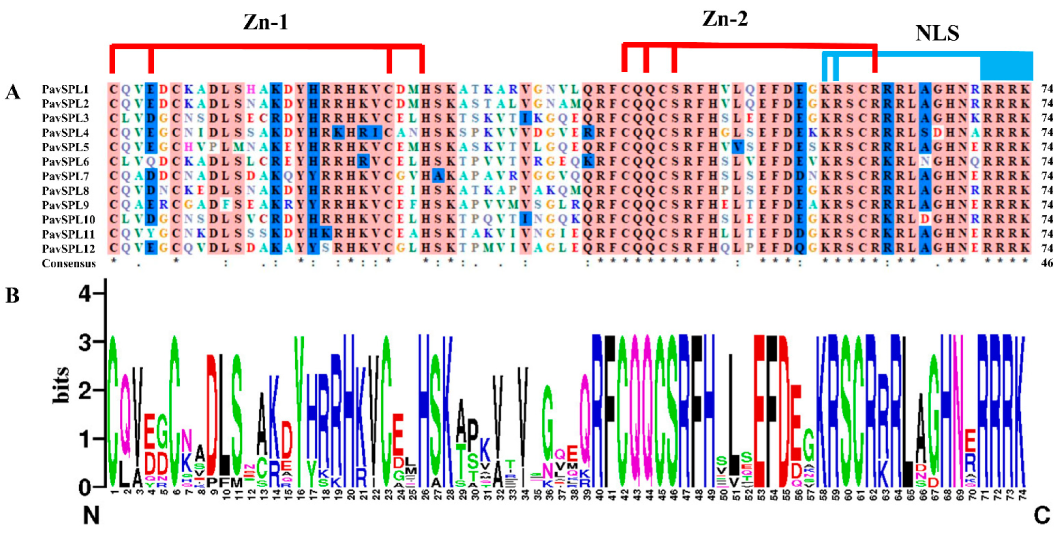
2.3. Sequence Alignments and Phylogenetic Analysis of SPL Genes
2.4. Conserved Motifs and Structure Analysis of the PavSPL Genes
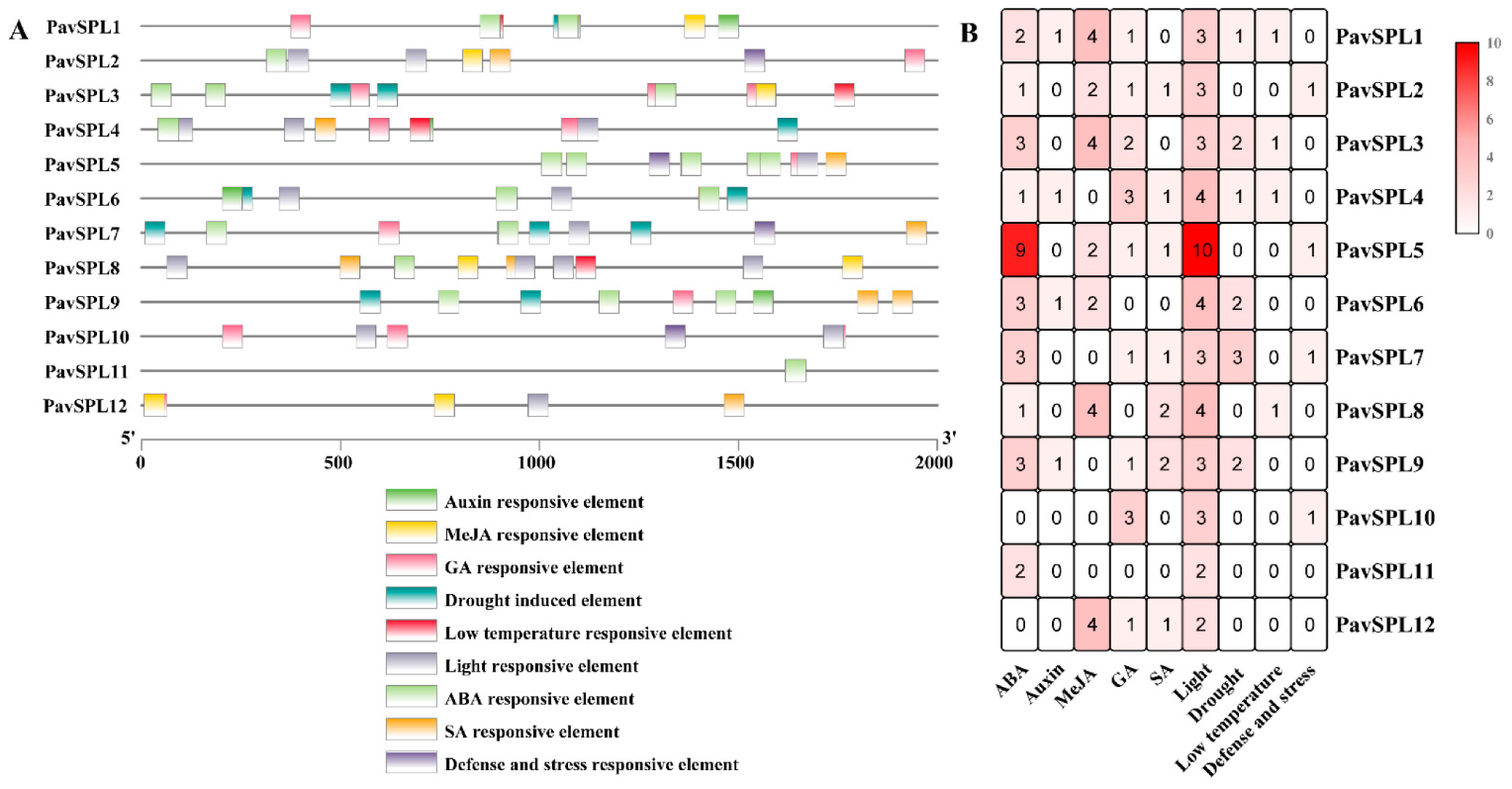
2.5. Cis-Acting Elements in the Promoters of PavSPL Genes
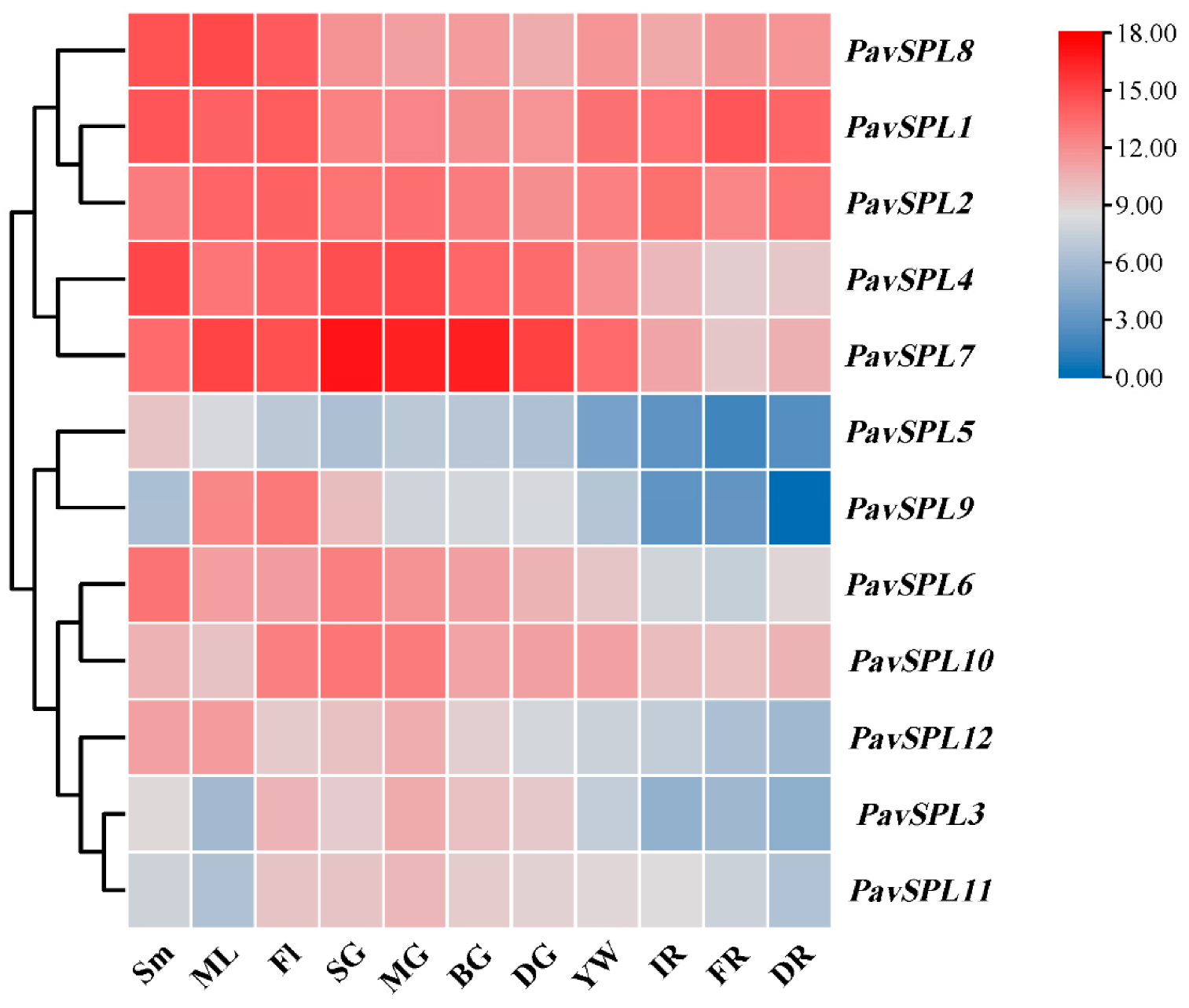
2.6. Expression Analysis of the PavSPL Genes in Different Tissues and in Fruit Development
2.7. Expression Analysis of the PavSPL Genes during Hormone Treatment
2.8. Subcellular Localization of PavSPLs
3. Discussion
3.1. Bioinformatics Analysis of SPL Gene Family in Sweet Cherry
3.2. Expression Profiles of PavSPLs in Sweet Cherry Development
3.3. Expression Profiles of PavSPLs in Plant Responses to Multiple Hormones
4. Materials and Methods
4.1. Plant Materials and Hormone Treatments
4.2. Identification of SPL Genes in Sweet Cherry
4.3. Sequence Alignments, Phylogenetic Analyses, and Gene Structure Analysis
4.4. Cis-Acting Element Prediction and Chromosomal Localization Analysis
4.5. Expression Analysis of PavSPLs
4.6. Subcellular Localization Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riechmann, J.L.; Ratcliffe, O.J. A genomic perspective on plant transcription factors. Curr Opin Plant. Biol. 2000, 3, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 1996, 250, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Cardon, G.; Hohmann, S.; Klein, J.; Nettesheim, K.; Saedler, H.; Huijser, P. Molecular characterisation of the Arabidopsis SBP-box genes. Gene 1999, 237, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Salinas, M.; Xing, S.; Hohmann, S.; Berndtgen, R.; Huijser, P. Genomic organization, phylogenetic comparison and differential expression of the SBP-box family of transcription factors in tomato. Planta 2012, 235, 1171–1184. [Google Scholar] [CrossRef]
- Xiong, J.; Zheng, D.; Zhu, H.; Chen, J.; Na, R.; Cheng, Z. Genome-wide identification and expression analysis of the SPL gene family in woodland strawberry Fragaria vesca. Genome 2018, 61, 675–683. [Google Scholar] [CrossRef]
- Li, J.; Hou, H.; Li, X.; Xiang, J.; Yin, X.; Gao, H.; Zheng, Y.; Bassett, C.L.; Wang, X. Genome-wide identification and analysis of the SBP-box family genes in apple (Malus × domestica Borkh.). Plant. Physiol. Biochem. 2013, 70, 100–114. [Google Scholar] [CrossRef]
- Song, S.; Zhou, H.; Sheng, S.; Cao, M.; Li, Y.; Pang, X. Genome-wide organization and expression profiling of the SBP-Box gene family in Chinese jujube (Ziziphus jujuba Mill.). Int J. Mol. Sci. 2017, 18, 1734. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, X.; Gu, S.; Hu, Z.; Xu, H.; Xu, C. Comparative study of SBP-box gene family in Arabidopsis and rice. Gene 2008, 407, 1–11. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E.; et al. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Jach, G.; Saedler, H.; Huijser, P. Functional dissection of the plant-specific SBP-domain: Overlap of the DNA-binding and nuclear localization domains. J. Mol. Biol. 2005, 352, 585–596. [Google Scholar] [CrossRef]
- Gandikota, M.; Birkenbihl, R.P.; Hohmann, S.; Cardon, G.H.; Saedler, H.; Huijser, P. The miRNA156/157 recognition element in the 3’ UTR of the Arabidopsis SBP box gene SPL3 prevents early flowering by translational inhibition in seedlings. Plant. J. 2007, 49, 683–693. [Google Scholar] [CrossRef]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant. Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef]
- Wang, L.; Yu, P.; Lyu, J.; Hu, Y.; Han, C.; Bai, M.Y.; Fan, M. BZR1 physically interacts with SPL9 to regulate the vegetative phase change and cell elongation in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 415. [Google Scholar] [CrossRef]
- Zhang, Y.; Schwarz, S.; Saedler, H.; Huijser, P. SPL8, a local regulator in a subset of gibberellin-mediated developmental processes in Arabidopsis. Plant. Mol. Biol. 2007, 63, 429–439. [Google Scholar] [CrossRef]
- Gou, J.; Felippes, F.F.; Liu, C.; Detlef, W.; Wang, J. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant. Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef]
- Chuck, G.; Whipple, C.; Jackson, D.; Hake, S. The maize SBP-box transcription factor encoded by tasselsheath4 regulates bract development and the establishment of meristem boundaries. Development 2010, 137, 1243–1250. [Google Scholar] [CrossRef]
- Manning, K.; Tor, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat. Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Eriksson, E.M.; Bovy, A.; Manning, K.; Harrison, L.; Andrews, J.; De Silva, J.; Tucker, G.A.; Seymour, G.B. Effect of the colorless non-ripening mutation on cell wall biochemistry and gene expression during tomato fruit development and ripening. Plant. Physiol. 2004, 136, 4184–4197. [Google Scholar] [CrossRef]
- Lai, T.; Wang, X.; Ye, B.; Jin, M.; Chen, W.; Wang, Y.; Zhou, Y.; Blanks, A.M.; Gu, M.; Zhang, P.; et al. Molecular and functional characterization of the SBP-box transcription factor SPL-CNR in tomato fruit ripening and cell death. J. Exp. Bot. 2020, 71, 2995–3011. [Google Scholar] [CrossRef]
- Liu, H.; Shu, Q.; Lin-Wang, K.; Allan, A.C.; Espley, R.V.; Su, J.; Pei, M.; Wu, J. The PyPIF5-PymiR156a-PySPL9-PyMYB114/MYB10 module regulates light-induced anthocyanin biosynthesis in red pear. Mol. Hortic. 2021, 1. [Google Scholar] [CrossRef]
- Li, X.; Hou, Y.; Xie, X.; Li, H.; Li, X.; Zhu, Y.; Zhai, L.; Zhang, C.; Bian, S. A blueberry MIR156a-SPL12 module coordinates the accumulation of chlorophylls and anthocyanins during fruit ripening. J. Exp. Bot. 2020, 71, 5976–5989. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Gao, H.; Chen, H.; Fu, C. The involvement of papaya CpSBP1 in modulating fruit softening and carotenoid accumulation by repressing CpPME1/2 and CpPDS4. Sci. Hortic. 2019, 256. [Google Scholar] [CrossRef]
- Zhao, X.; Qu, D.; Wang, L.; Gao, Y.; An, N.; Wang, A.; Li, Y.; Yang, J.; Wu, F.; Su, H. Genome-wide identification of cysteine-rich receptor-like kinases in sweet cherry reveals that PaCRK1 enhances sweet cherry resistance to salt stress. Plant. Cell Rep. 2022, 41, 2037–2088. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, P.; Dai, S.; Sun, Y.; Yuan, B.; Kai, W.; Pei, Y.; He, S.; Liang, B.; Zhang, Y.; et al. PacCYP707A2 negatively regulates cherry fruit ripening while PacCYP707A1 mediates drought tolerance. J. Exp. Bot. 2015, 66, 3765–3774. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Fan, X.; Mattheis, J.P. Responses of ‘Bing’ and ‘Rainier’ sweet cherries to ethylene and 1-methylcyclopropene. J. Amer. Soc. Hort. Sci. 2002, 127, 831–835. [Google Scholar] [CrossRef]
- Tijero, V.; Teribia, N.; Munoz, P.; Munne-Bosch, S. Implication of abscisic acid on ripening and quality in sweet cherries: Differential effects during pre- and post-harvest. Front. Plant. Sci. 2016, 7, 602. [Google Scholar] [CrossRef]
- Kumar, R.; Khurana, A.; Sharma, A.K. Role of plant hormones and their interplay in development and ripening of fleshy fruits. J. Exp. Bot. 2014, 65, 4561–4575. [Google Scholar] [CrossRef]
- Chockchaisawasdee, S.; Golding, J.B.; Vuong, Q.V.; Papoutsis, K.; Stathopoulos, C.E. Sweet cherry: Composition, postharvest preservation, processing and trends for its future use. Trends Food Sci. Tech. 2016, 55, 72–83. [Google Scholar] [CrossRef]
- Correia, S.; Schouten, R.; Silva, A.P.; Goncalves, B. Factors affecting quality and health promoting compounds during growth and postharvest life of sweet cherry (Prunus avium L.). Front. Plant. Sci 2017, 8, 2166. [Google Scholar] [CrossRef]
- Zhang, D.; Han, Z.; Li, J.; Qin, H.; Zhou, L.; Wang, Y.; Zhu, X.; Ma, Y.; Fang, W. Genome-wide analysis of the SBP-box gene family transcription factors and their responses to abiotic stresses in tea (Camellia sinensis). Genomics 2020, 112, 2194–2202. [Google Scholar] [CrossRef]
- Hou, H.; Yan, X.; Sha, T.; Yan, Q.; Wang, X. The SBP-box gene VpSBP11 from Chinese wild Vitis is involved in floral transition and affects leaf development. Int J. Mol. Sci. 2017, 18, 1493. [Google Scholar] [CrossRef]
- Liu, M.; Sun, W.; Ma, Z.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide identification of the SPL gene family in tartary buckwheat (Fagopyrum tataricum) and expression analysis during fruit development stages. BMC Plant. Biol. 2019, 19, 299. [Google Scholar] [CrossRef]
- Tan, H.; Song, X.; Duan, W.; Wang, Y.; Hou, X. Genome-wide analysis of the SBP-box gene family in Chinese cabbage (Brassica rapa subsp. pekinensis). Genome 2015, 58, 463–477. [Google Scholar] [CrossRef]
- Cai, C.; Guo, W.; Zhang, B. Genome-wide identification and characterization of SPL transcription factor family and their evolution and expression profiling analysis in cotton. Sci Rep. 2018, 8, 762. [Google Scholar] [CrossRef]
- Shen, X.; Zhao, K.; Liu, L.; Zhang, K.; Yuan, H.; Liao, X.; Wang, Q.; Guo, X.; Li, F.; Li, T. A role for PacMYBA in ABA-regulated anthocyanin biosynthesis in red-colored sweet cherry cv. Hong Deng (Prunus avium L.). Plant. Cell Physiol. 2014, 55, 862–880. [Google Scholar] [CrossRef]
- Qi, X.; Liu, C.; Song, L.; Li, M. PaMADS7, a MADS-box transcription factor, regulates sweet cherry fruit ripening and softening. Plant. Sci. 2020, 301, 110634. [Google Scholar] [CrossRef]
- Jin, W.; Wang, H.; Li, M.; Wang, J.; Yang, Y.; Zhang, X.; Yan, G.; Zhang, H.; Liu, J.; Zhang, K. The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant. Biotechnol. J. 2016, 14, 2120–2133. [Google Scholar] [CrossRef]
- Chen, W.; Yu, Z.; Kong, J.; Wang, H.; Li, Y.; Zhao, M.; Wang, X.; Zheng, Q.; Shi, N.; Zhang, P.; et al. Comparative WGBS identifies genes that influence non-ripe phenotype in tomato epimutant Colourless non-ripening. Sci. China Life Sci. 2018, 61, 244–252. [Google Scholar] [CrossRef]
- Cui, M.; Wang, C.; Zhang, W.; Pervaiz, T.; Haider, M.S.; Tang, W.; Fang, J. Characterization of Vv-miR156: Vv-SPL pairs involved in the modulation of grape berry development and ripening. Mol. Genet. Genom. 2018, 293, 1333–1354. [Google Scholar] [CrossRef]
- Leng, P.; Yuan, B.; Guo, Y. The role of abscisic acid in fruit ripening and responses to abiotic stress. J. Exp. Bot. 2014, 65, 4577–4588. [Google Scholar] [CrossRef]
- Guo, J.; Wang, S.; Yu, X.; Dong, R.; Li, Y.; Mei, X.; Shen, Y. Polyamines regulate strawberry fruit ripening by abscisic acid, auxin, and ethylene. Plant. Physiol. 2018, 177, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Santner, A.; Calderon-Villalobos, L.I.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, N.; Ponce, C.; Arellano, M.; Time, A.; Sagredo, B.; Donoso, J.M.; Meisel, L.A. Gibberellic acid modifies the transcript abundance of ABA pathway orthologs and modulates sweet cherry (Prunus avium) fruit ripening in early- and mid-season varieties. Plants (Basel) 2020, 9, 1796. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Yan, S.; Jing, Y.; Yang, R.; Zhang, Y.; Zhou, Y.; Zhu, Y.; Sun, J. MIR156-targeted SPL9 is phosphorylated by SnRK2s and interacts with ABI5 to enhance ABA responses in Arabidopsis. Front. Plant. Sci. 2021, 12, 708573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, X.; Li, J.; Cai, H.; Deng, X.W.; Li, L. MicroRNA408 is critical for the HY5-SPL7 gene network that mediates the coordinated response to light and copper. Plant. Cell 2014, 26, 4933–4953. [Google Scholar] [CrossRef]
- Wang, Y.; Zhai, Z.; Sun, Y.; Feng, C.; Peng, X.; Zhang, X.; Xiao, Y.; Zhou, X.; Wang, W.; Jiao, J.; et al. Genome-wide identification of the B-BOX genes that respond to multiple ripening related signals in sweet cherry fruit. Int. J. Mol. Sci. 2021, 22, 1622. [Google Scholar] [CrossRef]
- Jia, H.F.; Chai, Y.M.; Li, C.L.; Lu, D.; Luo, J.J.; Qin, L.; Shen, Y.Y. Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant. Physiol. 2011, 157, 188–199. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Protein /AA | Chrom. | Chr Start | Chr End | MW (Da) |
|---|---|---|---|---|---|---|
| PavSPL1 | Pav_sc0000065.1_g080.1 | 1034 | Chr1 | 6612478 | 6619865 | 115029.50 |
| PavSPL2 | Pav_sc0000065.1_g070.1 | 1009 | Chr1 | 6620405 | 6626786 | 112292.00 |
| PavSPL3 | Pav_sc0000091.1_g140.1 | 316 | Chr1 | 18093845 | 18097381 | 34624.93 |
| PavSPL4 | Pav_sc0000091.1_g350.1 | 443 | Chr1 | 18230101 | 18234634 | 48283.35 |
| PavSPL5 | Pav_sc0000120.1_g190.1 | 434 | Chr2 | 12784392 | 12787547 | 47451.18 |
| PavSPL6 | Pav_sc0001080.1_g880.1 | 217 | Chr3 | 21876290 | 21878055 | 25097.53 |
| PavSPL7 | Pav_sc0001305.1_g910.1 | 162 | Chr4 | 3003020 | 3004866 | 18563.10 |
| PavSPL8 | Pav_sc0000600.1_g500.1 | 1069 | Chr4 | 5991293 | 5997912 | 118373.64 |
| PavSPL9 | Pav_sc0000975.1_g210.1 | 162 | Chr4 | 13389781 | 13391094 | 18440.52 |
| PavSPL10 | Pav_sc0000740.1_g280.1 | 401 | Chr5 | 11637365 | 11640898 | 43899.78 |
| PavSPL11 | Pav_sc0000358.1_g620.1 | 559 | Chr5 | 15069897 | 15073721 | 61412.26 |
| PavSPL12 | Pav_sc0001280.1_g530.1 | 383 | Chr6 | 17835832 | 17840241 | 41097.72 |
| Gene 1 | Gene 2 | Ka | Ks | Ka/Ks | Gene Duplication |
|---|---|---|---|---|---|
| PavSPL3 | PavSPL6 | 0.462 | 1.551 | 0.298 | Segmental |
| PavSPL3 | PavSPL10 | 0.406 | 2.133 | 0.190 | Segmental |
| PavSPL6 | PavSPL10 | 0.400 | NaN | NaN | Segmental |
| PavSPL7 | PavSPL9 | 0.343 | 1.379 | 0.249 | Segmental |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Wang, Y.; Xiao, Y.; Zhang, X.; Du, B.; Turupu, M.; Wang, C.; Yao, Q.; Gai, S.; Huang, J.; et al. Genome-Wide Identification of the SQUAMOSA Promoter-Binding Protein-like (SPL) Transcription Factor Family in Sweet Cherry Fruit. Int. J. Mol. Sci. 2023, 24, 2880. https://doi.org/10.3390/ijms24032880
Sun Y, Wang Y, Xiao Y, Zhang X, Du B, Turupu M, Wang C, Yao Q, Gai S, Huang J, et al. Genome-Wide Identification of the SQUAMOSA Promoter-Binding Protein-like (SPL) Transcription Factor Family in Sweet Cherry Fruit. International Journal of Molecular Sciences. 2023; 24(3):2880. https://doi.org/10.3390/ijms24032880
Chicago/Turabian StyleSun, Yueting, Yanyan Wang, Yuqin Xiao, Xiang Zhang, Bingyang Du, Maihemuti Turupu, Chao Wang, Qisheng Yao, Shilin Gai, Jing Huang, and et al. 2023. "Genome-Wide Identification of the SQUAMOSA Promoter-Binding Protein-like (SPL) Transcription Factor Family in Sweet Cherry Fruit" International Journal of Molecular Sciences 24, no. 3: 2880. https://doi.org/10.3390/ijms24032880
APA StyleSun, Y., Wang, Y., Xiao, Y., Zhang, X., Du, B., Turupu, M., Wang, C., Yao, Q., Gai, S., Huang, J., Tong, S., & Li, T. (2023). Genome-Wide Identification of the SQUAMOSA Promoter-Binding Protein-like (SPL) Transcription Factor Family in Sweet Cherry Fruit. International Journal of Molecular Sciences, 24(3), 2880. https://doi.org/10.3390/ijms24032880