
Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action

 ,
, 
Abstract
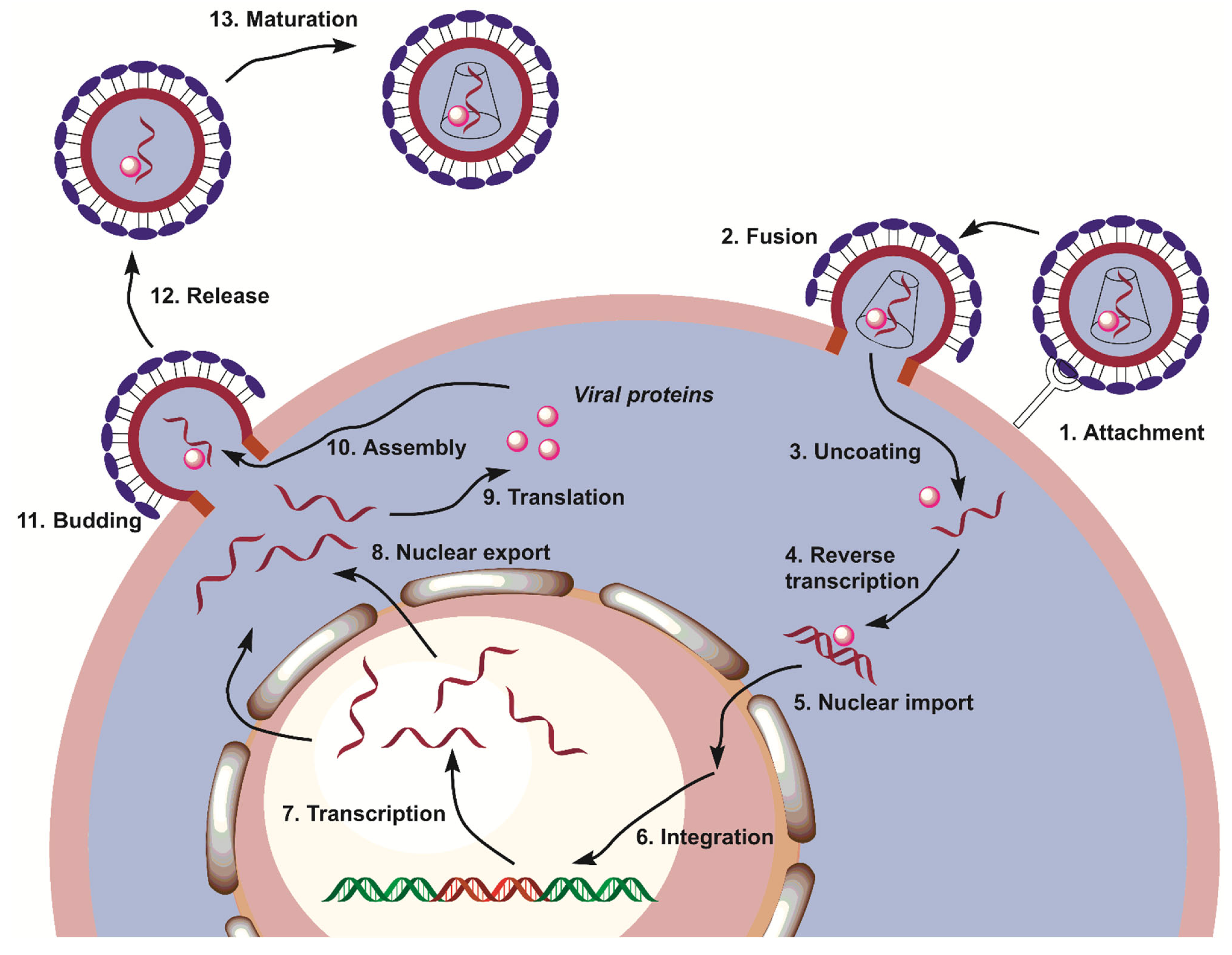
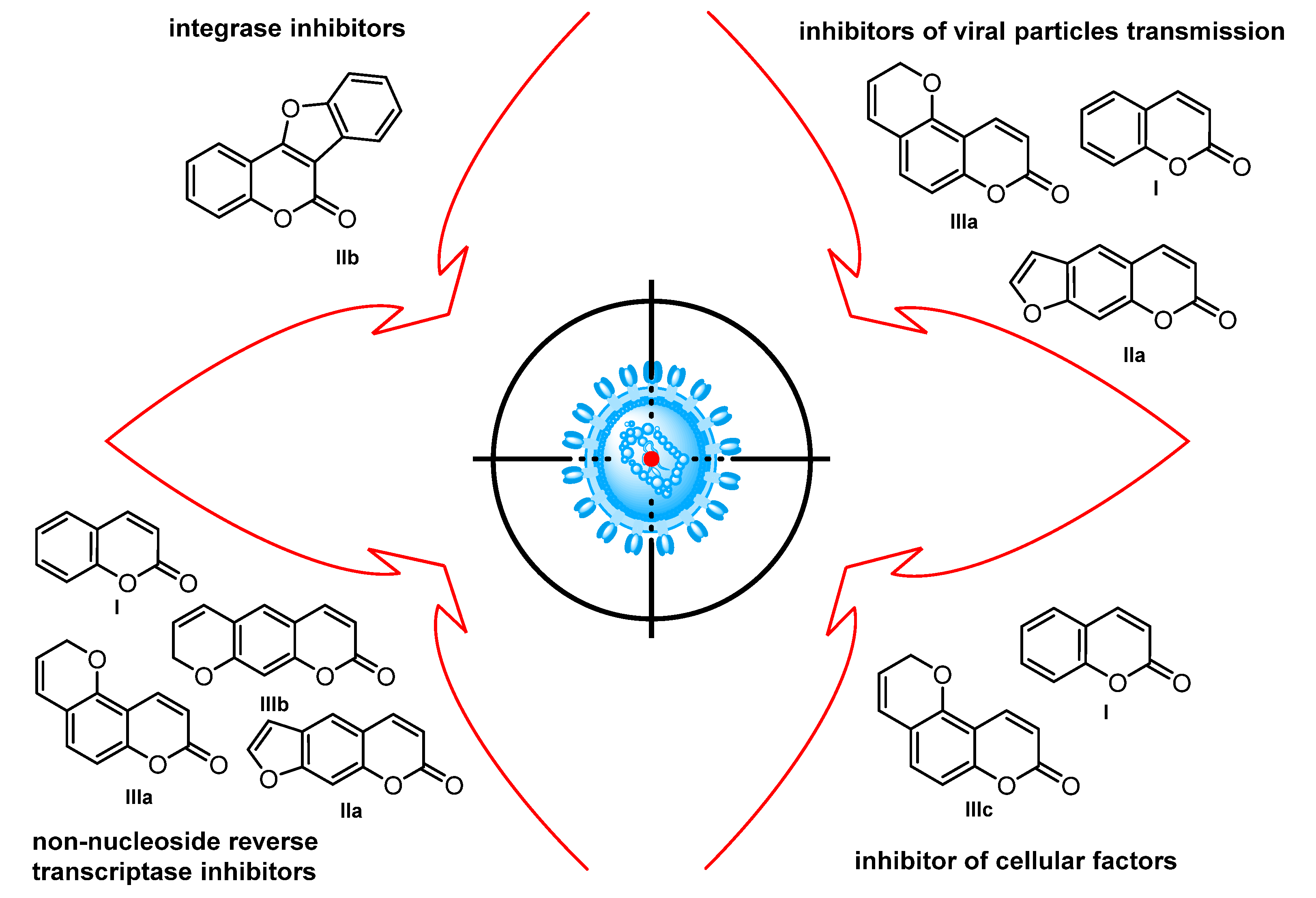
1. Introduction
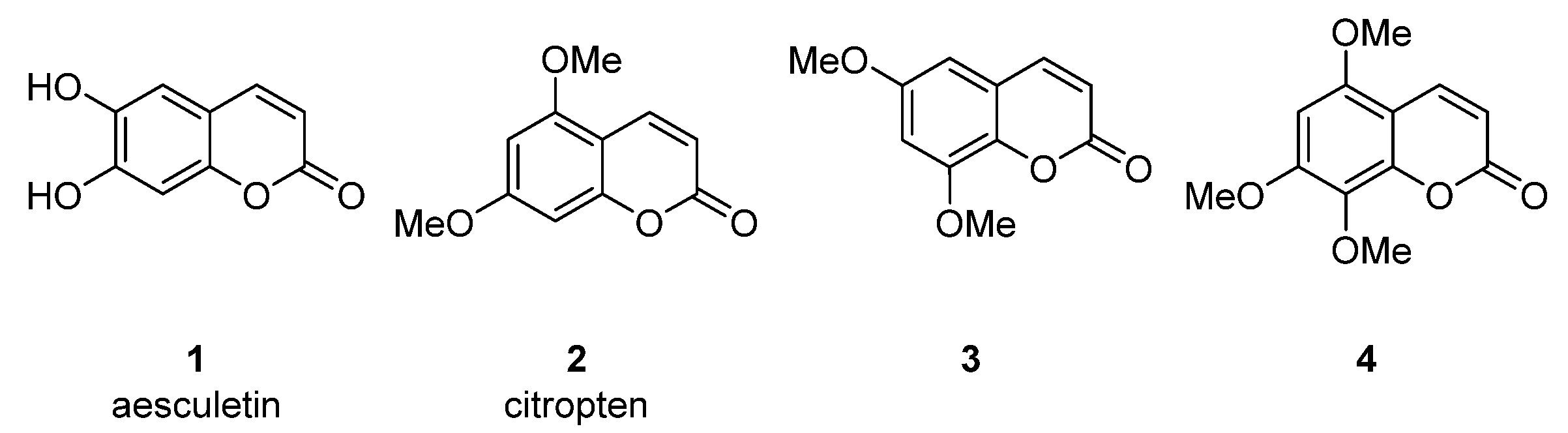
2. Simple Coumarins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
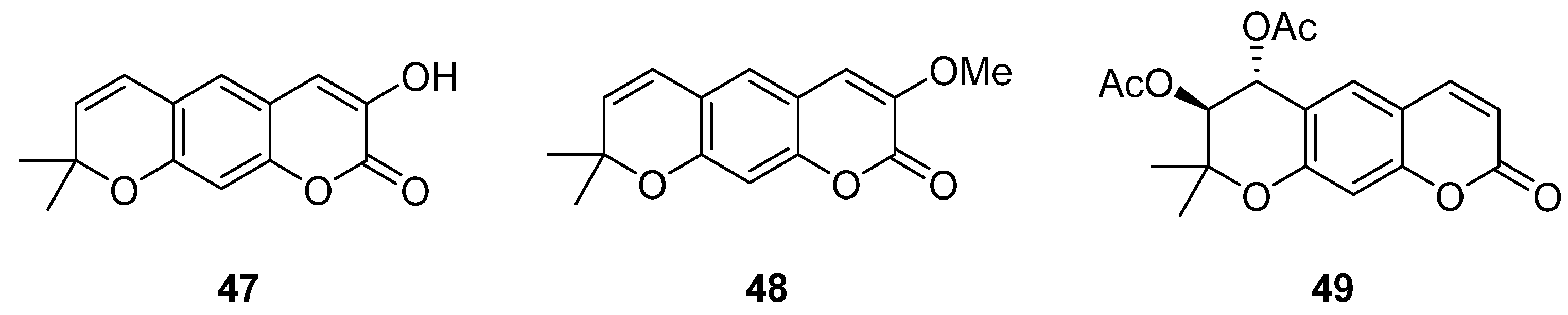{kind=link}
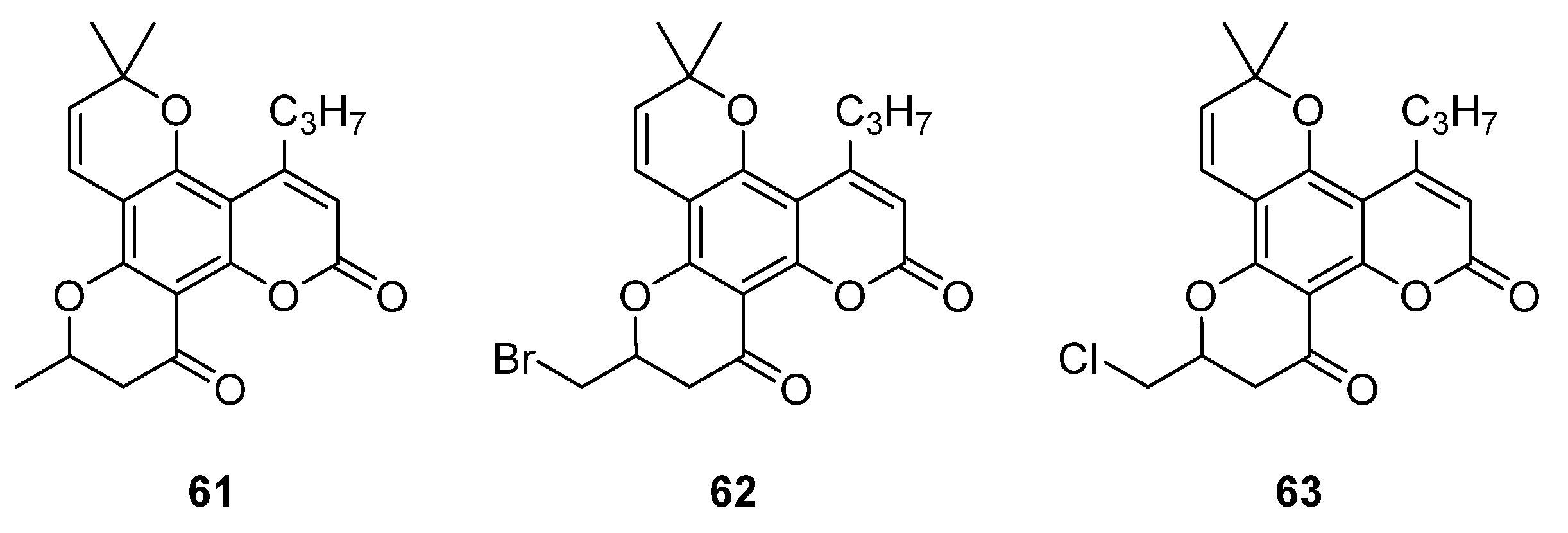{kind=link}
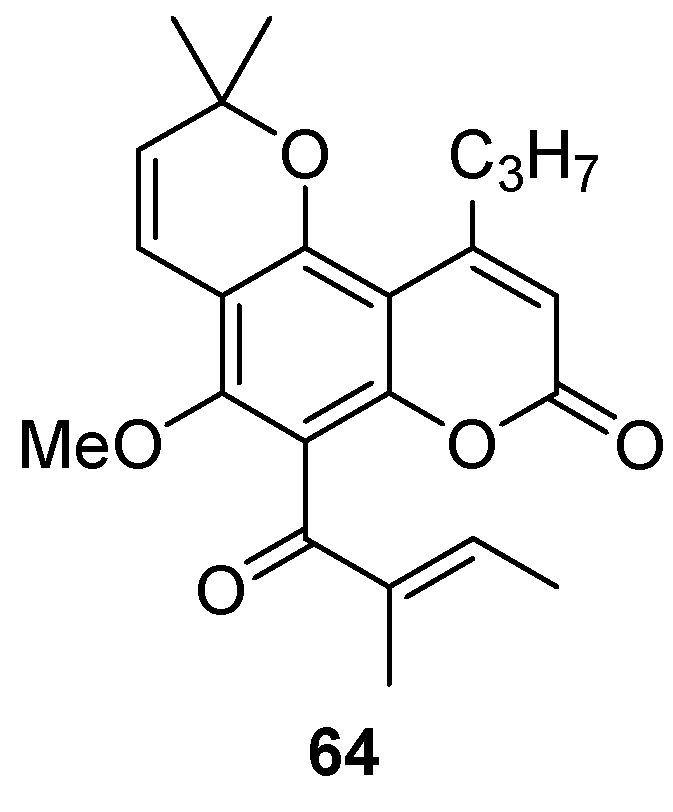{kind=link}
{kind=link}
| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
 auraptene 5 | Triphasia trifolia [24] Zanthoxylum schinifolium [25] | EC50 = 5.7 μM CC50 = 46.8 μM TI = 8.21 [24] |
 anisocoumarin B 6 | Poncirus trifoliata [22] Clausena anisata [31] | EC50 = 18.3 μM CC50 = 813 μM TI = 44.4 [24] |
 isobtusitin 7 | Psiadia dentata [33] | EC10 = 20 μM CC50 = 20 μM [33] |
 farnesiferol C 8 | Ferula assa-foetida [35] Ferula szowitsiana [36] Ferula kopetdaghensis [37] | EC50 = 30 μM [35] CC50 > 150 μM [34] |
| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
| Monoterpenoids | ||
| C3-monoterpenoids 9 | ||
 9 | Poncirus trifoliata [22] | EC50 = 10.7 μM CC50 = 330 μM TI = 31 [22] |
| C6-monoterpenoids 10–16 | ||
 fipsomin 10 | Ficus nipponica [38] Artocarpus heterophyllus [45] | EC50 = 2.93 μM CC50 > 200 μM TI > 68.26 [45] |
 phellodenol C 11 | Phellodendron amurense [39] Artocarpus heterophyllus [45] | EC50 = 8.07 μM CC50 > 200 μM TI > 21.93 [45] |
 isophellodenol C 12 | Heracleum candicans [40] Artocarpus heterophyllus [45] | EC50 = 9.12 μM CC50 > 200 μM TI > 24.78 [45] |
 13 | Manilkara zapota [41] Ruta pinnata [42] Skimmia laureola [43] Acalypha indica [44] | EC50 = 8.69 μM CC50 > 200 μM TI > 38.02 [41] |
 14 | Artocarpus heterophyllus [45] Brombya sp. Nova [46] | EC50 = 5.68 μM CC50 > 200 μM TI > 35.21 [45] |
 (+)-trachypleuranin A 15 | Manilkara zapota [41] Harbouria trachypleura [47] | EC50 = 5.26 μM CC50 > 200 μM TI > 38.02 [41] |
 16 | Manilkara zapota [41] Skimmia laureola [43] Clausena lenis [48] | EC50 = 6.73 μM CC50 > 200 μM TI > 29.72 [41] EC50 = 7.39 μM CC50 > 200 μM TI > 31.9 [48] |
| C8-monoterpenoids 17, 18 | ||
 osthol 17 | Many species. For recent reviews, see [49,50] | EC50 = 0.64 μM CC50 = 48 μM TI = 75.5 [30] |
 O-methylcedrelopsin 18 | Zanthoxylum ailanthoides [23] | EC50 = 7.66 μM CC50 = 77 μM TI = 36.6 [23] |
| Diterpenoids | ||
| C3,C6-diterpenoids 19–27 | ||
 gravelliferone 19 | Manilkara zapota [41] Clausena lansium [51] Helietta apiculata [52] | EC50 = 2.28 μM CC50 > 200 μM TI > 87.7 [41] |
 O-methyl gravelliferone 20 | Manilkara zapota [41] Ruta pinnata [53] | EC50 = 3.49 μM CC50 > 200 μM TI > 35.3 [41] |
 tanizin 21 | Manilkara zapota [41] Artocarpus heterophyllus [45] Helietta apiculata [52] | EC50 = 4.26 μM CC50 > 200 μM TI > 47.0 [41]EC50 = 0.56 μM CC50 > 200 μM TI > 357 [45] |
 manizapotin A 22 | Manilkara zapota [41] | EC50 = 0.12 μM CC50 > 200 μM TI > 1667 [41] |
 manizapotin B 23 | Manilkara zapota [41] | EC50 = 0.33 μM CC50 > 200 μM TI > 606 [41] |
 manizapotin C 24 | Manilkara zapota [41] | EC50 = 0.42 μM CC50 > 200 μM TI > 467 [41] |
 clauselenin B 25 | Clausena lenis [48] | EC50 = 0.68 μM CC50 > 200 μM TI > 294 [48] |
 artoheteronin 26 | Artocarpus heterophyllus [45] | EC50 = 0.18 μM CC50 > 200 μM TI > 1111 [45] |
 anisocoumarin A 27 | Manilkara zapota [41] Artocarpus heterophyllus [45] | EC50 = 0.97 μM CC50 > 200 μMTI > 206 [41] EC50 = 0.33 μM CC50 > 200 μMTI > 606 [45] |
| C6,C8-diterpenoids 28 | ||
 6,8-diprenylumbelliferone 28 | Clausena lenis [48] | EC50 = 1.59 μM CC50 > 200 μM TI > 125 [48] |
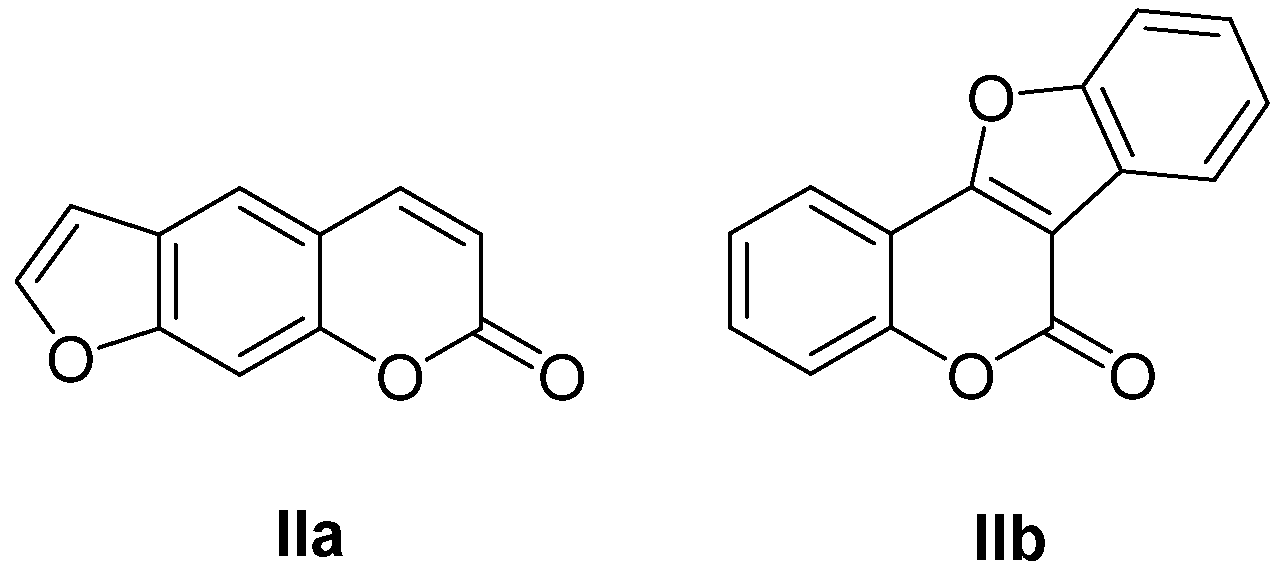
3. Furocoumarins
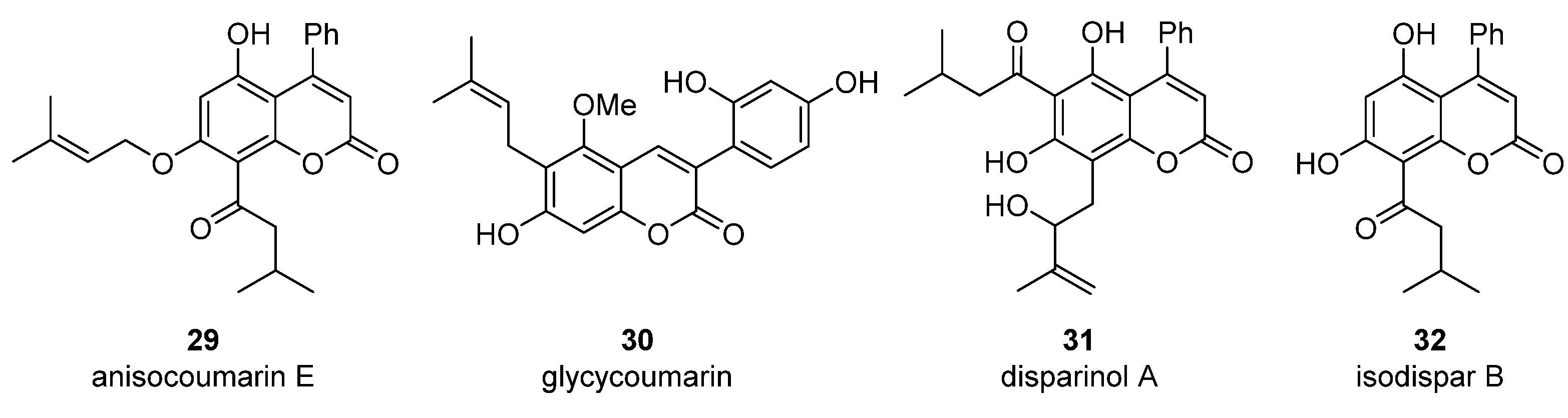
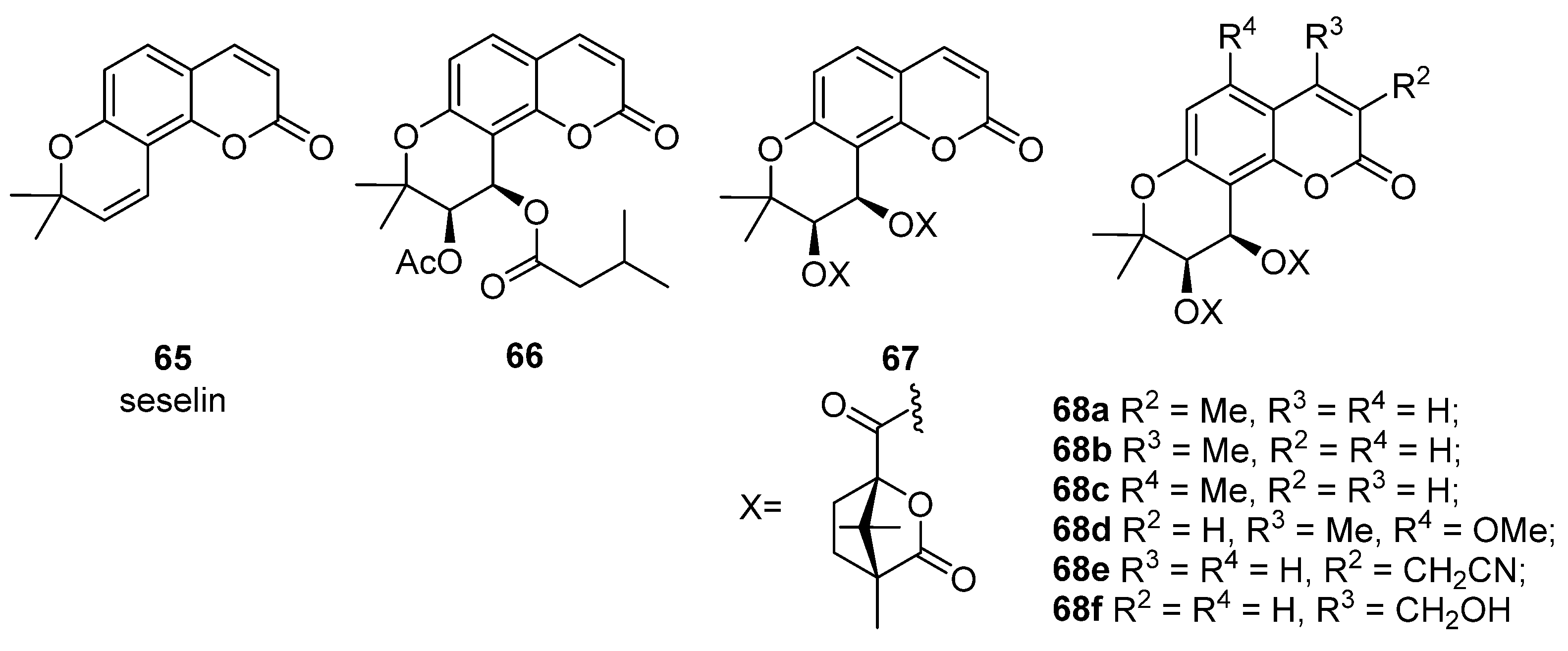
4. Pyranocoumarins
- (1)
- the presence of bulky substituents at the C(4) position;
- (2)
- positions C(10), C(11) and C(12) of the chromanol ring are sensitive to modification;
- (3)
- position C(12) must contain a hydrogen bond acceptor.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abraham, K.; Wöhrlin, F.; Lindtner, O.; Heinemeyer, G.; Lampen, A. Toxicology and risk assessment of coumarin: Focus on human data. Mol. Nutr. Food Res. 2010, 54, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Heghes, S.C.; Vostinaru, O.; Mogosan, C.; Miere, D.; Iuga, C.A.; Filip, L. Safety Profile of Nutraceuticals Rich in Coumarins: An Update. Front. Pharmacol. 2022, 13, 803338. [Google Scholar] [CrossRef] [PubMed]
- Lake, B.G. Coumarin metabolism, toxicity and carcinogenicity. Food Chem. Toxicol. 1999, 37, 423–453. [Google Scholar] [CrossRef] [PubMed]
- Vella, S.; Schwartländer, B.; Sow, S.P.; Eholie, S.P.; Murphy, R.L. The history of antiretroviral therapy and of its implementation in resource-limited areas of the world. AIDS 2012, 26, 1231–1241. [Google Scholar] [CrossRef]
- Al-Abbasa, N.S.; Shaer, N.A. Combination of coumarin and doxorubicin induces drug-resistant acute myeloid leukemia cell death. Heliyon 2021, 7, e06255. [Google Scholar] [CrossRef]
- Zou, J.; Liu, Y.; Guo, R.; Tang, Y.; Shi, Z.; Zhang, M.; Wu, W.; Chen, Y.; Hou, K. An In Vitro Coumarin-Antibiotic Combination Treatment of Pseudomonas aeruginosa Biofilms. Nat. Prod. Commun. 2021, 16, 1–7. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Osswald, H.L.; Prato, G. Recent Progress in the Development of HIV-1 Protease Inhibitors for the Treatment of HIV/AIDS. J. Med. Chem. 2016, 59, 5172–5208. [Google Scholar] [CrossRef]
- Sharapov, A.D.; Fatykhov, R.F.; Khalymbadzha, I.A.; Sharutin, V.V.; Santra, S.; Zyryanov, G.V.; Chupakhin, O.N.; Ranu, B.C. Mechanochemical synthesis of coumarins via Pechmann condensation under solvent-free conditions: An easy access to coumarins and annulated pyrano-[2,3-f] and [3,2-f]indoles. Green Chem. 2022, 24, 2429–2437. [Google Scholar] [CrossRef]
- Lončarić, M.; Gašo-Sokač, D.; Jokić, S.; Molnar, M. Recent Advances in the Synthesis of Coumarin Derivatives from Different Starting Materials. Biomolecules 2020, 10, 151. [Google Scholar] [CrossRef]
- Serna-Arbeláez, M.S.; Florez-Sampedro, L.; Orozco, L.P.; Ramírez, K.; Galeano, E.; Zapata, W. Natural Products with Inhibitory Activity against Human Immunodeficiency Virus Type 1. Adv. Virol. 2021, 2021, 5552088. [Google Scholar] [CrossRef]
- Cos, P.; Maes, L.; Vlietinck, A.; Pieters, L. Plant-Derived Leading Compounds for Chemotherapy of Human Immunodefiency Virus (HIV) Infection—An Update (1998–2007). Planta Med. 2008, 74, 1323–1337. [Google Scholar] [CrossRef]
- Kostova, I.; Raleva, S.; Genova, P.; Argirova, R. Structure-Activity Relationships of Synthetic Coumarins as HIV-1 Inhibitors. Bioinorg. Chem. Appl. 2006, 2006, 68274. [Google Scholar] [CrossRef]
- Engelman, A.; Cherepanov, P. The structural biology of HIV-1: Mechanistic and therapeutic insights. Nat. Rev. Genet. 2012, 10, 279–290. [Google Scholar] [CrossRef]
- Arts, E.J.; Hazuda, D.J. HIV-1 Antiretroviral Drug Therapy. Cold Spring Harb. Perspect. Med. 2012, 2, a007161. [Google Scholar] [CrossRef]
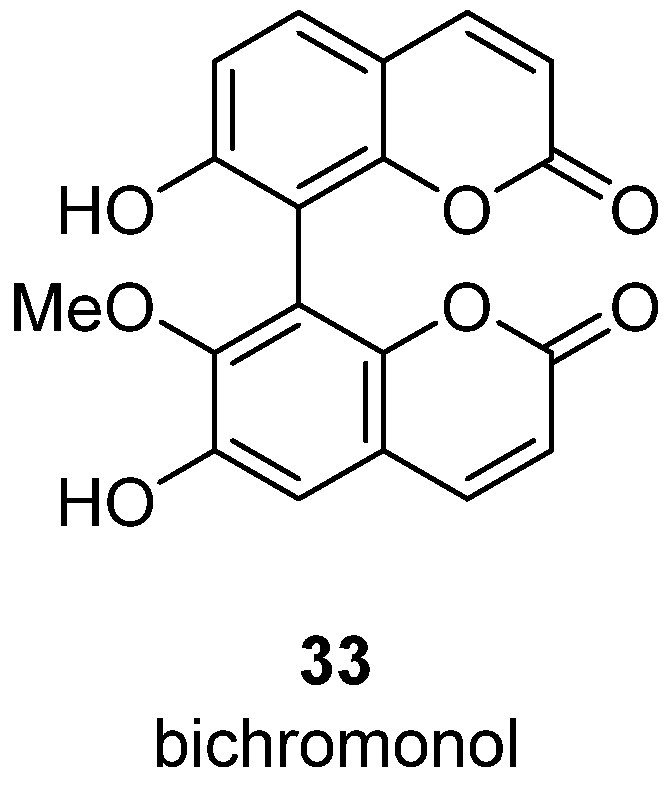
- Fobofou, S.A.T.; Franke, K.; Brandt, W.; Manzin, A.; Madeddu, S.; Serreli, G.; Sanna, G.; Wessjohann, L.A. Bichromonol, a dimeric coumarin with anti-HIV activity from the stem bark of Hypericum roeperianum. Nat. Prod. Res. 2022, 1–7. [Google Scholar] [CrossRef]
- Venugopala, K.N.; Rashmi, V.; Odhav, B. Review on Natural Coumarin Lead Compounds for Their Pharmacological Activity. BioMed. Res. Int. 2013, 2013, 963248. [Google Scholar] [CrossRef]
- Galabov, A.S.; Iosifova, T.; Vassileva, E.; Kostova, I. Antiviral Activity of Some Hydroxycoumarin Derivatives. Z. Für Nat. C 1996, 51, 558–562. [Google Scholar] [CrossRef]
- Bayoumi, S.A.; Rowan, M.G.; Beeching, J.R.; Blagbrough, I.S. Constituents and secondary metabolite natural products in fresh and deteriorated cassava roots. Phytochemistry 2010, 71, 598–604. [Google Scholar] [CrossRef]
- Wu, T.-S.; Tsang, Z.-J.; Wu, P.-L.; Lin, F.-W.; Li, C.-Y.; Teng, C.-M.; Lee, K.-H. New constituents and antiplatelet aggregation and anti-HIV principles of Artemisia capillaris. Bioorganic Med. Chem. 2001, 9, 77–83. [Google Scholar] [CrossRef]
- Kuo, P.-C.; Hwang, T.-L.; Lin, Y.-T.; Kuo, Y.-C.; Leu, Y.-L. Chemical constituents from Lobelia chinensis and their anti-virus and anti-inflammatory bioactivities. Arch. Pharmacal Res. 2011, 34, 715–722. [Google Scholar] [CrossRef]
- Chan, Y.-Y.; Li, C.-H.; Shen, Y.-C.; Wu, T.-S. Anti-inflammatory Principles from the Stem and Root Barks of Citrus medica. Chem. Pharm. Bull. 2010, 58, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Wang, R.-R.; Cai, X.-H.; Zheng, Y.-T.; Luo, X.-D. Anti-human Immunodeficiency Virus-1 Constituents of the Bark of Poncirus trifoliata. Chem. Pharm. Bull. 2010, 58, 971–975. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-J.; Lee, K.-H.; Tsai, I.-L.; Chen, I.-S. Two new sesquiterpenoids and anti-HIV principles from the root bark of Zanthoxylum ailanthoides. Bioorganic Med. Chem. 2005, 13, 5915–5920. [Google Scholar] [CrossRef] [PubMed]
- Likhitwitayawuid, K.; Supudompol, B.; Sritularak, B.; Lipipun, V.; Rapp, K.; Schinazi, R.F. Phenolics with Anti-HSV and Anti-HIV Activities from Artocarpus gomezianus., Mallotus pallidus., and Triphasia trifolia. Pharm. Biol. 2005, 43, 651–657. [Google Scholar] [CrossRef]
- Epifano, F.; Genovese, S.; Marcotullio, M.; Curini, M. Chemistry and Pharmacology of Collinin, Active Principle of Zanthoxylum spp. Mini-Rev. Med. Chem. 2008, 8, 1203–1208. [Google Scholar] [CrossRef]
- Adianti, M.; Aoki, C.; Komoto, M.; Deng, L.; Shoji, I.; Wahyuni, T.S.; Lusida, M.I.; Fuchino, S.H.; Kawahara, N.; Hotta, H. Anti-hepatitis C virus compounds obtained from Glycyrrhiza uralensis and other Glycyrrhiza species. Microbiol. Immunol. 2014, 58, 180–187. [Google Scholar] [CrossRef]
- Chang, C.-T.; Doong, S.-L.; Tsai, I.-L.; Chen, I.-S. Coumarins and anti-HBV constituents from Zanthoxylum schinifolium. Phytochemistry 1997, 45, 1419–1422. [Google Scholar] [CrossRef]
- Lv, H.-N.; Wang, S.; Zeng, K.-W.; Li, J.; Guo, X.-Y.; Ferreira, D.; Zjawiony, J.K.; Tu, P.-F.; Jiang, Y. Anti-inflammatory Coumarin and Benzocoumarin Derivatives from Murraya alata. J. Nat. Prod. 2015, 78, 279–285. [Google Scholar] [CrossRef]
- Lee, C.-L.; Chiang, L.-C.; Cheng, L.-H.; Liaw, C.-C.; El-Razek, M.H.A.; Chang, F.-R.; Wu, Y.-C. Influenza A (H(1)N(1)) Antiviral and Cytotoxic Agents from Ferula assa-foetida. J. Nat. Prod. 2009, 72, 1568–1572. [Google Scholar] [CrossRef]
- Zhou, P.; Takaishi, Y.; Duan, H.; Chen, B.; Honda, G.; Itoh, M.; Takeda, Y.; Kodzhimatov, O.K.; Lee, K.-H. Coumarins and bicoumarin from Ferula sumbul: Anti-HIV activity and inhibition of cytokine release. Phytochemistry 2000, 53, 689–697. [Google Scholar] [CrossRef]
- Ngadjui, T.B.; Ayafor, J.F.; Sondengam, B.L.; Connolly, J.D. Coumarins from Clausena anisate. Phytochemistry 1989, 28, 585–589. [Google Scholar] [CrossRef]
- Hatano, T.; Yasuhara, T.; Miyamoto, K.; Okuda, T. Anti-human immunodeficiency virus phenolics from licorice. Chem. Pharm. Bull. 1988, 36, 2286–2288. [Google Scholar] [CrossRef]
- Fortin, H.; Tomasi, S.; Jaccard, P.; Robin, V.; Boustie, J. A Prenyloxycoumarin from Psiadia dentata. Chem. Pharm. Bull. 2001, 49, 619–621. [Google Scholar] [CrossRef]
- Kasaian, J.; Mohammadi, A. Biological activities of farnesiferol C: A review. J. Asian Nat. Prod. Res. 2017, 20, 27–35. [Google Scholar] [CrossRef]
- Sistani, P.; Dehghan, G.; Sadeghi, L. Structural and kinetic insights into HIV-1 reverse transcriptase inhibition by farnesiferol C. Int. J. Biol. Macromol. 2021, 174, 309–318. [Google Scholar] [CrossRef]
- Turabelidze, D.G.; Kemertelidze, É.P. Farnesiferol C from the roots of Ferula szovitsiana. Chem. Nat. Compd. 1976, 12, 589–589. [Google Scholar] [CrossRef]
- Kamilov, K.M.; Nikonov, G.K. Coumarins of Ferula kopetdaghensis and the structure of kopeolin and kopeoside. Chem. Nat. Compd. 1973, 9, 294–298. [Google Scholar] [CrossRef]
- Akihara, Y.; Ohta, E.; Nehira, T.; Ômura, H.; Ohta, S. New Prenylated ortho-Dihydroxycoumarins from the Fruits of Ficus nipponica. Chem. Biodivers. 2017, 14, e1700196. [Google Scholar] [CrossRef]
- Wu, T.S.; Hsu, M.Y.; Kuo, P.C.; Sreenivasulu, B.; Damu, A.G.; Su, C.R.; Li, C.Y.; Chang, H.C. Constituents from the leaves of Phellodendron amurense var. wilsonii and their bioactivity. J. Nat. Prod. 2003, 66, 1207–1211. [Google Scholar] [CrossRef]
- Nakamori, T.; Taniguchi, M.; Shibano, M.; Wang, N.-H.; Baba, K. Chemical studies on the root of Heracleum candicans WALL. J. Nat. Med. 2008, 62, 403–412. [Google Scholar] [CrossRef]
- Liu, Y.-P.; Yan, G.; Guo, J.-M.; Liu, Y.-Y.; Li, Y.-J.; Zhao, Y.-Y.; Qiang, L.; Fu, Y.-H. Prenylated Coumarins from the Fruits of Manilkara zapota with Potential Anti-inflammatory Effects and Anti-HIV Activities. J. Agric. Food Chem. 2019, 67, 11942–11947. [Google Scholar] [CrossRef]
- Gonzalez, A.G.; Reyes, R.E.; Espino, M.R. Two new coumarins from Ruta pinnata. Phytochemistry 1977, 16, 2033–2035. [Google Scholar] [CrossRef]
- Atta-ur-Rahman; Sultana, N.; Khan, M.R.; Choudhary, M.I. Triterpene and coumarins from Skimmia laureola. Nat. Prod. Lett. 2002, 16, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, P.; Sundari, S.; Selvaraj, S.; Natarajan, A.; Suganya, R.; Jayaprakash, R.; Kasthuri, K.; Kumaresan, S. GC-MS, Phytochemical Analysis and In Silico Approaches of a Medicinal Plant Acalypha indica. J. Sci. Res. 2022, 14, 671–684. [Google Scholar] [CrossRef]
- Tao, L.; Zhuo, Y.-T.; Qiao, Z.-H.; Li, J.; Tang, H.-X.; Yu, Q.-M.; Liu, Y.-Y.; Liu, Y.-P. Prenylated coumarins from the fruits of Artocarpus heterophyllus with their potential anti-inflammatory and anti-HIV activities. Nat. Prod. Res. 2022, 36, 2526–2533. [Google Scholar] [CrossRef]
- Halstead, C.W.; Forster, P.I.; Waterman, P.G. Novel Metabolites from the Stem Bark of Brombya sp. Nova (Gap Creek) (Rutaceae). Nat. Prod. Commun. 2006, 1, 351–356. [Google Scholar] [CrossRef]
- Guz, N.R.; Lorenz, P.; Stermitz, F.R. New coumarins from Harbouria trachypleura: Isolation and synthesis. Tetrahedron Lett. 2001, 42, 6491–6494. [Google Scholar] [CrossRef]
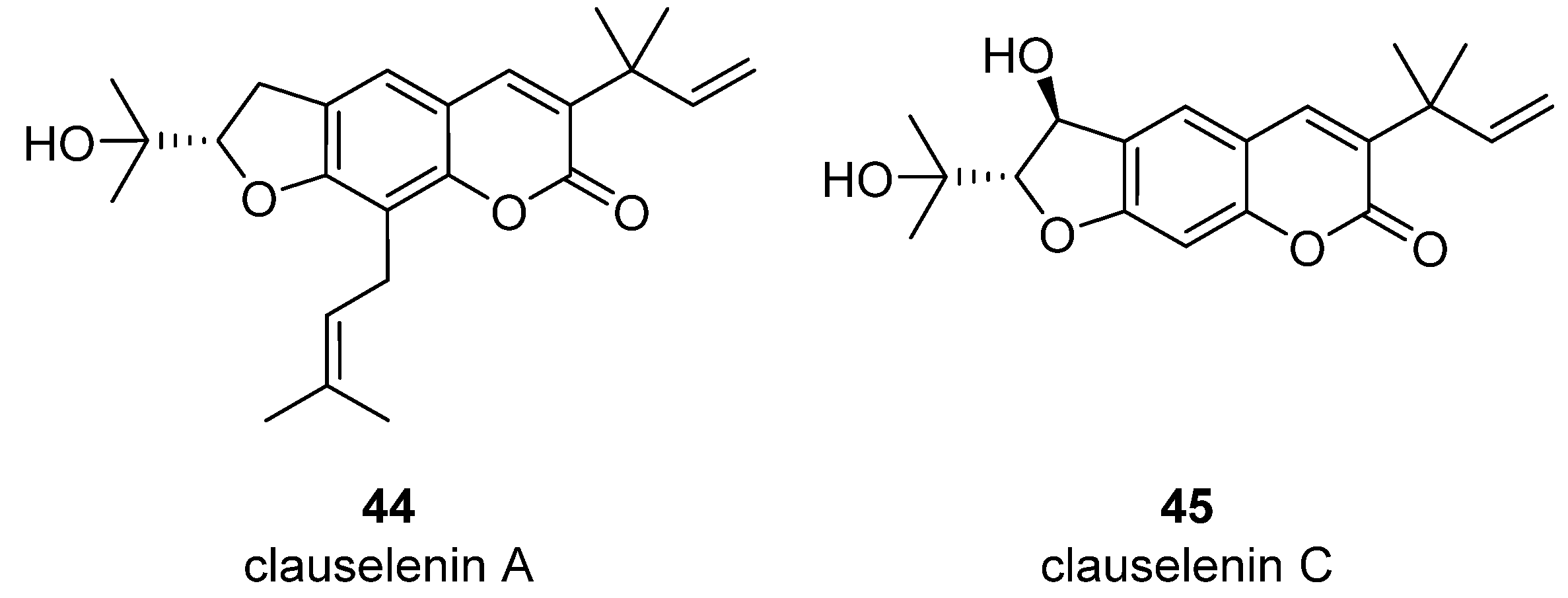
- Liu, Y.-P.; Yan, G.; Xie, Y.-T.; Lin, T.-C.; Zhang, W.; Li, J.; Wu, Y.-J.; Zhou, J.-Y.; Fu, Y.-H. Bioactive prenylated coumarins as potential anti-inflammatory and anti-HIV agents from Clausena lenis. Bioorganic Chem. 2020, 97, 103699. [Google Scholar] [CrossRef]
- Sun, M.; Sun, M.; Zhang, J. Osthole: An overview of its sources, biological activities, and modification development. Med. Chem. Res. 2021, 30, 1767–1794. [Google Scholar] [CrossRef]
- Zafar, S.; Sarfraz, I.; Rasul, A.; Shah, M.A.; Hussain, G.; Zahoor, M.K.; Shafiq, N.; Riaz, A.; Selamoglu, Z.; Sarker, S.D. Osthole: A Multifunctional Natural Compound with Potential Anticancer, Antioxidant and Anti-inflammatory Activities. Mini-Rev. Med. Chem. 2021, 21, 2747–2763. [Google Scholar] [CrossRef]
- Kumar, V.; Vallipuram, K.; Adebajo, A.C.; Reisch, J. 2,7-dihydroxy-3-formyl-1-(3′-methyl-2′-butenyl) carbazole from Clausena lansium. Phytochemistry 1995, 40, 1563−1565. [Google Scholar] [CrossRef]
- Fernandes, T.S.; Copetti, D.; Carmo, G.D.; Neto, A.T.; Pedroso, M.; Silva, U.F.; Mostardeiro, M.A.; Burrow, R.E.; Dalcol, I.I.; Morel, A.F. Phytochemical analysis of bark from Helietta apiculata Benth and antimicrobial activities. Phytochemistry 2017, 141, 131–139. [Google Scholar] [CrossRef]
- González, A.; Estévez, R.; Jaraiz, I. Structure of sabandinin and other coumarins isolated from the roots of Ruta pinnata. Phytochemistry 1971, 10, 1621–1626. [Google Scholar] [CrossRef]
- Bedoya, L.M.; Beltrán, M.; Sancho, R.; Olmedo, D.A.; Sánchez-Palomino, S.; del Olmo, E.; López-Pérez, J.L.; Muñoz, E.; San Feliciano, A.; Alcamí, J. 4-Phenylcoumarins as HIV transcription inhibitors. Bioorg. Med. Chem. Lett. 2005, 15, 4447–4450. [Google Scholar] [CrossRef]
- El-Gogary, T.M.; Koehler, G.J. Interaction of psoralens with DNA-bases (II): An ab initio quantum chemical, density functional theory and second-order MØller-Plesset perturbational study. Mol. Struc. THEOCHEM 2009, 895, 57–64. [Google Scholar] [CrossRef]
- El-Gogary, T.M. Interaction of angelicin with DNA-bases (III) DFT and ab initio second-order Moeller-Plesset study. Int. J. Quantum Chem. 2009, 110, 1445–1454. [Google Scholar] [CrossRef]
- Stern, R.S.; Nichols, K.T.; Väkevä, L.H. Malignant Melanoma in Patients Treated for Psoriasis with Methoxsalen (Psoralen) and Ultraviolet A Radiation (PUVA). N. Engl. J. Med. 1997, 336, 1041–1045. [Google Scholar] [CrossRef]
- Shikishima, Y.; Takaishi, Y.; Honda, G.; Ito, M.; Takeda, Y.; Kodzhimatov, O.K.; Ashurmetov, O.; Lee, K.-H. Chemical Constituents of Prangos tschimganica; Structure Elucidation and Absolute Configuration of Coumarin and Furanocoumarin Derivatives with Anti-HIV Activity. Chem. Pharm. Bull. 2001, 49, 877–880. [Google Scholar] [CrossRef]
- Sancho, R.; Márquez, N.; Gómez-Gonzalo, M.; Calzado, M.A.; Bettoni, G.; Coiras, M.; Alcami, J.; López-Cabrera, M.; Appendino, G.B.; Muñoz, E. Imperatorin Inhibits HIV-1 Replication through an Sp1-dependent Pathway. J. Biol. Chem. 2004, 279, 37349–37359. [Google Scholar] [CrossRef]
- Nebo, L.; Varela, R.M.; Molinillo, J.M.G.; Sampaio, O.M.; Severino, V.G.; Cazal, C.M.; Fernandes, M.F.D.G.; Fernandes, J.B.; Macías, F.A. Phytotoxicity of alkaloids, coumarins and flavonoids isolated from 11 species belonging to the Rutaceae and Meliaceae families. Phytochem. Lett. 2014, 8, 226–232. [Google Scholar] [CrossRef]
- Heinke, R.; Franke, K.; Porzel, A.; Wessjohann, L.A.; Awadh, A.N.A.; Schmidt, J. Furanocoumarins from Dorstenia foetida. Phytochemistry 2011, 72, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Ban, H.S.; Lim, S.S.; Suzuki, K.; Jung, S.H.; Lee, S.; Lee, Y.S.; Shin, K.H.; Ohuchi, K. Inhibitory Effects of Furanocoumarins Isolated from the Roots of Angelica dahurica on Prostaglandin E2 Production. Planta Med. 2003, 69, 408–412. [Google Scholar] [PubMed]
- Kostova, I.; Ivanova, A.; Mikhova, B.; Klaiber, I. Alkaloids and Coumarins from Ruta graveolens. Monatsh. Chem. 1999, 130, 703–707. [Google Scholar] [CrossRef]
- Liu, H.; Li, F.; Li, C.-J.; Yang, J.-Z.; Li, L.; Chen, N.-H.; Zhang, D.-M. Bioactive furanocoumarins from stems of Clausena lansium. Phytochemistry 2014, 107, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Harkar, S.; Razdan, T.; Waight, E. Steroids, chromone and coumarins from angelica officinalis. Phytochemistry 1984, 23, 419–426. [Google Scholar] [CrossRef]
- Liu, X.; Nam, J.-W.; Song, Y.S.; Viswanath, A.N.I.; Pae, A.N.; Kil, Y.-S.; Kim, H.-D.; Park, J.H.; Seo, E.-K.; Chang, M. Psoralidin, a coumestan analogue, as a novel potent estrogen receptor signaling molecule isolated from Psoralea corylifolia. Bioorganic Med. Chem. Lett. 2014, 24, 1403–1406. [Google Scholar] [CrossRef]
- Jang, D.S.; Kim, J.M.; Lee, Y.M.; Kim, Y.S.; Kim, J.-H.; Kim, J.S. Puerariafuran, a New Inhibitor of Advanced Glycation End Products (AGEs) Isolated from the Roots of Pueraria lobata. Chem. Pharm. Bull. 2006, 54, 1315–1317. [Google Scholar] [CrossRef]
- Yadav, P.P.; Ahmad, G.; Maurya, R. Furanoflavonoids from Pongamia pinnata fruits. Phytochemistry 2004, 65, 439–443. [Google Scholar] [CrossRef]
- Macías, F.A.; Simonet, A.M.; Galindo, J.C.; Castellano, D. Bioactive phenolics and polar compounds from Melilotus messanensis. Phytochemistry 1999, 50, 35–46. [Google Scholar] [CrossRef]
- Tuskaev, V.A. Synthesis and biological activity of coumestan derivatives (Review). Pharm. Chem. J. 2013, 47, 1–11. [Google Scholar] [CrossRef]
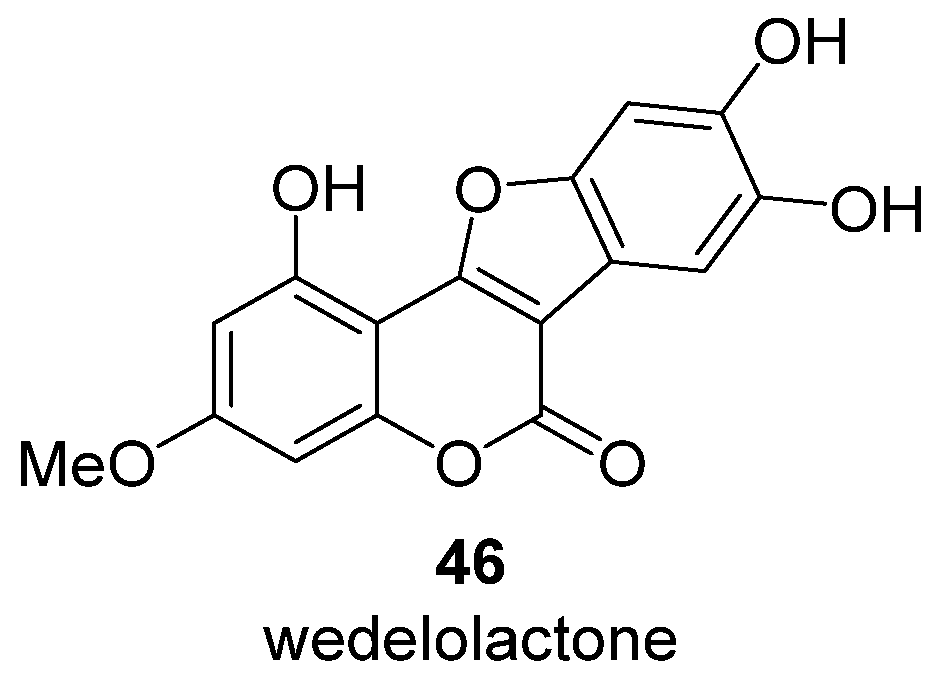
- Tewtrakul, S.; Subhadhirasakul, S.; Cheenpracha, S.; Karalai, C. HIV-1 protease and HIV-1 integrase inhibitory substances from Eclipta prostrata. Phytotherapy Res. 2007, 21, 1092–1095. [Google Scholar] [CrossRef]
- Lee, T.T.-Y.; Kashiwada, Y.; Huang, L.; Snider, J.; Cosentino, M.; Lee, K.-H. Suksdorfin: An anti-HIV principle from Lomatium suksdorfii, its structure-activity correlation with related coumarins, and synergistic effects with anti-AIDS nucleosides. Bioorganic Med. Chem. 1994, 2, 1051–1056. [Google Scholar] [CrossRef]
- Ishikawa, T. Anti HIV-1 Active Calophyllum Coumarins: Distribution, Chemistry, and Activity. Heterocycles 2000, 53, 453–474. [Google Scholar] [CrossRef]
- Huang, L.; Kashiwada, Y.; Cosentino, L.M.; Fan, S.; Lee, K.-H. 3′,4′-Di-o-(−)-camphanoyl-(+)-ciskhellactone and related compounds: A. new class of potent anti-HIV agents. Bioorg. Med. Chem. Lett. 1994, 4, 593–598. [Google Scholar] [CrossRef]
- Chen, L.-W.; Cheng, M.-J.; Peng, C.-F.; Chen, I.-S. Secondary Metabolites and Antimycobacterial Activities from the Roots of Ficus nervosa. Chem. Biodivers. 2010, 7, 1814–1821. [Google Scholar] [CrossRef]
- Su, X.-H.; Zhang, M.-L.; Li, L.-G.; Huo, C.-H.; Gu, Y.-C.; Shi, Q.-W. Chemical Constituents of the Plants of the Genus Calophyllum. Chem. Biodivers. 2008, 5, 2579–2608. [Google Scholar] [CrossRef]
- Flavin, M.T.; Rizzo, J.D.; Khilevich, A.; Kucherenko, A.; Sheinkman, A.K.; Vilaychack, V.; Lin, L.; Chen, W.; Greenwood, E.M.; Pengsuparp, T.; et al. Synthesis, Chromatographic Resolution, and Anti-Human Immunodeficiency Virus Activity of (±)-Calanolide A and Its Enantiomers. J. Med. Chem. 1996, 39, 1303–1313. [Google Scholar] [CrossRef]
- Matthée, G.; Wright, A.D.; König, G.M. HIV Reverse Transcriptase Inhibitors of Natural Origin. Planta Med. 1999, 65, 493–506. [Google Scholar] [CrossRef]
- Kashman, Y.; Gustafson, K.R.; Fuller, R.W.; Ii, J.H.C.; McMahon, J.B.; Currens, M.J.; Buckheit, R.W.; Hughes, S.H.; Cragg, G.M.; Boyd, M.R. HIV inhibitory natural products. Part 7. The calanolides, a novel HIV-inhibitory class of coumarin derivatives from the tropical rainforest tree, Calophyllum lanigerum. J. Med. Chem. 1992, 35, 2735–2743. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Eggleston, D.S.; Haltiwanger, R.C.; Bean, M.F.; Taylor, P.B.; Caranfa, M.J.; Breen, A.L.; Bartus, H.R. The inophyllums, novel inhibitors of HIV-1 reverse transcriptase isolated from the Malaysian tree, Calophyllum inophyllum Linn. J. Med. Chem. 1993, 36, 4131–4138. [Google Scholar] [CrossRef]
- Yu, D.; Suzuki, M.; Xie, L.; Morris-Natschke, S.L.; Lee, K.-H. Recent progress in the development of coumarin derivatives as potent anti-HIV agents. Med. Res. Rev. 2003, 23, 322–345. [Google Scholar] [CrossRef] [PubMed]
- Cardellina, J.H.; Bokesch, H.R.; McKee, T.C.; Boyd, M.R. Resolution and comparative anti-HIV evaluation of the enantiomers of calanolides A and B. Bioorganic Med. Chem. Lett. 1995, 5, 1011–1014. [Google Scholar] [CrossRef]
- Usach, I.; Melis, V.; Peris, J.-E. Non-nucleoside reverse transcriptase inhibitors: A review on pharmacokinetics, pharmacodynamics, safety and tolerability. J. Int. AIDS Soc. 2013, 16, 18567. [Google Scholar] [CrossRef] [PubMed]
- Creagh, T.; Ruckle, J.L.; Tolbert, D.T.; Giltner, J.; Eiznhamer, D.A.; Dutta, B.; Flavin, M.T.; Xu, Z.-Q. Safety and Pharmacokinetics of Single Doses of (+)-Calanolide A, a Novel, Naturally Occurring Nonnucleoside Reverse Transcriptase Inhibitor, in Healthy, Human Immunodeficiency Virus-Negative Human Subjects. Antimicrob. Agents Chemother. 2001, 45, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Liu, L.; Xue, H.; Han, C.; Wang, L.; Chen, Z.; Liu, G. Chemical Library and Structure–Activity Relationships of 11-Demethyl-12-oxo Calanolide A Analogues as Anti-HIV-1 Agents. J. Med. Chem. 2008, 51, 1432–1446. [Google Scholar] [CrossRef]
- Xue, H.; Lu, X.; Zheng, P.; Liu, L.; Han, C.; Hu, J.; Liu, Z.; Ma, T.; Li, Y.; Wang, L.; et al. Highly Suppressing Wild-Type HIV-1 and Y181C Mutant HIV-1 Strains by 10-Chloromethyl-11-demethyl-12-oxo-calanolide A with Druggable Profile. J. Med. Chem. 2010, 53, 1397–1401. [Google Scholar] [CrossRef]
- Wang, L.; Chen, H.S.; Tao, P.Z.; Li, Y.; Bai, Y.; Hu, J.P.; Ma, T.; Xing, Z.T.; Peng, Z.G.; Zhou, C.M.; et al. Improved synthesis and pharmaceutical evaluation of racemic 11-demethyl calanolide A. Acta Pharm. Sin. 2008, 43, 707–718. [Google Scholar]
- Kimura, S.; Ito, C.; Jyoko, N.; Segawa, H.; Kuroda, J.; Okada, M.; Adachi, S.; Nakahata, T.; Yuasa, T.; Filho, V.C.; et al. Inhibition of leukemic cell growth by a novel anti-cancer drug (GUT-70) from Calophyllum Brasiliense that acts by induction of apoptosis. Int. J. Cancer 2004, 113, 158–165. [Google Scholar] [CrossRef]
- Ito, C.; Itoigawa, M.; Mishina, Y.; Filho, V.C.; Enjo, F.; Tokuda, H.; Nishino, H.; Furukawa, H. Chemical Constituents of Calophyllum brasiliense. 2. Structure of Three New Coumarins and Cancer Chemopreventive Activity of 4-Substituted Coumarins. J. Nat. Prod. 2003, 66, 368–371. [Google Scholar] [CrossRef]
- Matsuda, K.; Hattori, S.; Kariya, R.; Komizu, Y.; Kudo, E.; Goto, H.; Taura, M.; Ueoka, R.; Kimura, S.; Okada, S. Inhibition of HIV-1 entry by the tricyclic coumarin GUT-70 through the modification of membrane fluidity. Biochem. Biophys. Res. Commun. 2015, 457, 288–294. [Google Scholar] [CrossRef]
- Kudo, E.; Taura, M.; Matsuda, K.; Shimamoto, M.; Kariya, R.; Goto, H.; Hattori, S.; Kimura, S.; Okada, S. Inhibition of HIV-1 replication by a tricyclic coumarin GUT-70 in acutely and chronically infected cells. Bioorganic Med. Chem. Lett. 2012, 23, 606–609. [Google Scholar] [CrossRef]
- Xie, L.; Takeuchi, Y.; Cosentino, L.M.; Lee, K.-H. Anti-AIDS Agents. 37. Synthesis and Structure−Activity Relationships of (3′R,4′R)-(+)-cis-Khellactone Derivatives as Novel Potent Anti-HIV Agents. J. Med. Chem. 1999, 42, 2662–2672. [Google Scholar] [CrossRef]
- Xie, L.; Takeuchi, Y.; Cosentino, L.M.; McPhail, A.T.; Lee, K.-H. Anti-AIDS Agents. 42. Synthesis and Anti-HIV Activity of Disubstituted (3´R,4´R)-3´,4´-Di-O-(S)-camphanoyl-(+)-cis-khellactone Analogues. J. Med. Chem. 2001, 44, 664–671. [Google Scholar] [CrossRef]
- Xie, L.; Guo, H.-F.; Hong, L.; Zhuang, X.-M.; Zhang, A.-M.; Wu, G.; Ruan, J.-X.; Zhou, T.; Yu, D.; Qian, K.; et al. Development and Preclinical Studies of Broad-Spectrum Anti-HIV Agent (3′R,4′R)-3-Cyanomethyl-4-methyl-3′,4′-di-O-(S)-camphanoyl-(+)-cis-khellactone (3-Cyanomethyl-4-methyl-DCK). J. Med. Chem. 2008, 51, 7689–7696. [Google Scholar] [CrossRef]
- Amaral, J.C.; da Silva, M.M.; da Silva, M.F.G.F.; Alves, T.C.; Ferreira, A.G.; Forim, M.R.; Fernandes, J.B.; Pina, E.S.; Lopes, A.A.; Pereira, M.S.; et al. Advances in the Biosynthesis of Pyranocoumarins: Isolation and 13C-Incorporation Analysis by High-Performance Liquid Chromatography−Ultraviolet−Solid-Phase Extraction−Nuclear Magnetic Resonance Data. J. Nat. Prod. 2020, 83, 1409–1415. [Google Scholar] [CrossRef]
- Khandy, M.T.; Sofronova, A.K.; Gorpenchenko, T.Y.; Chirikova, N.K. Plant Pyranocoumarins: Description, Biosynthesis, Application. Plants 2022, 11, 3135. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Albohy, A.; Abdulrazik, B.S.; Bayoumi, S.A.L.; Malak, L.G.; Khallaf, I.S.A.; Bringmann, G.; Farag, S.F. Natural coumarins as potential anti-SARS-CoV-2 agents supported by docking analysis. RSC Adv. 2021, 11, 16970–16979. [Google Scholar] [CrossRef]












| Compound | Plant Source | Anti-HIV Activity and Toxicity |
|---|---|---|
 Psoralen 34 | Prangos tschimganica [58], Ruta graveolens [60], Dorstenia foetida [61] | EC50 = 0.54 μM CC50 = 103 μM TI = 191 [58] |
 Bergapten 35 | Zanthoxylum ailanthoides [23], Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62,63] | EC50 = 1.63 μM CC50 = 115 μM TI = 69.9 [58] |
 Imperatorin 36 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62], ,Clausena lansium [64], | EC50 <0.37 μM CC50 > 370 μM TI > 1000 [30] |
 Oxypeucedanin 37 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] | EC50 = 3.67 μM CC50 = 81.8 μM TI = 22.2 [30] |
 Heraclenin 38 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] | EC50 = 8.29 μM CC50 > 70.3 μM TI = 8.48 [30] |
 Oxypeucedanin Hydrate 39 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] | EC50 = 3.29 μM CC50 = 6.94 μM TI = 2.11 [30] |
 Heraclenol 40 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] | EC50 = 0.38 μM CC50 = 329 μM TI = 870 [30] |
 Oxypeucedanin methylate 41 | Ferula sumbul [30], Prangos tschimganica [58] | EC50 = 105 μM CC50 = 314 μM TI = 3.00 [30] |
 Saxalin 42 | Prangos tschimganica [58], Angelica officinalis [65] | EC50 = 6.99 μM CC50 = 81.7 μM TI = 11.7 [58] |
 Pabulenol 43 | Ferula sumbul [30], Prangos tschimganica [58], gen. Angelica [62] | EC50 = 22.3 μM CC50 = 58.4 μM TI = 2.61 [58] |
| Compound | Inhibition of HIV-1 PRP, μM | Anti-HIV Activity in CEM SS Cells and Toxicity |
|---|---|---|
 calanolide A 50 | IC50 = 0.07 [78] | IC50 = 0.1 μM CC50 = 20 μM TI = 200 [73,79] |
 calanolide B 51 | IC50 = 0.4 μM CC50 = 15 μM TI = 37 [73,76,79] | |
 calanolide C 52 | no significant activity CC50 = 30 μM [79] | |
 inophyllum A 53 | IC50 = 30 μM [80] | |
 inophyllum B 54 | IC50 = 0.038 [73] | IC50 = 1.4 μM CC50 = 55 μM TI = 39 [73,80] |
 inophyllum C 55 | low activity CC50 = 18 μM [80] | |
 inophyllum D 56 | low activity CC50 = 15 μM [80] | |
 inophyllum E 57 | low activity CC50 = 6.2 μM [80] | |
 inophyllum P 58 | IC50 = 0.130 [73] | IC50 = 1.6 μM CC50 = 55 μM TI = 16 [80,81] |
 (+)-oxocalanolide 59 | IC50 HIV-1 (IIIB) = 1.0 μM, CC50 HIV-1 (IIIB) > 10 μM IC50 HIV-1 (RF) = 0.9 μM, CC50 HIV-1 (RF) > 10 μM IC50 HIV-1 (SK1) = 0.17 μM, CC50 HIV-1 (SK1) > 10 μM IC50 HIV-2 (ROD) = 15.9 μM, CC50 HIV-1 (ROD) > 13.8 μM [81] | |
 (–)-oxocalanolide 60 | IC50 HIV-1 (IIIB)= 1.88 μM, CC50 HIV-1 (IIIB) > 10 μM IC50 HIV-1 (RF) = 3.41 μM, CC50 HIV-1 (RF) > 10 μM IC50 HIV-1 (SK1) = 0.27 μM, CC50 (SK1) = 55 μM [81] |
| Cell Line | CC50, μM | EC50, μM | TI (CC50/EC50) |
|---|---|---|---|
| U1 (PMA) | 8.44 | 3.48 | 2.43 |
| U1 (TNF-α) | 8.44 | 4.32 | 1.95 |
| Molt-4 | >10 | 3.41 | >2.93 |
| TZM-bl | >10 | – | – |
| Compound | Anti-HIV Activity, EC50 μM (TI) | Compound | Anti-HIV Activity, EC50 μM (TI) |
|---|---|---|---|
| Seselin 65 | 25.5 (12.9) [22] | 68c | 2.4 × 10–7 (4 × 108) [92] |
| Suksdorfin 66 | 1.3 (79) [72] | 68d | 7.2 × 10–6 (2.1 × 107) [93] |
| DCK 67 | 2.5 × 10–4 (1.4 × 105) [92] | 68e | 2.4 × 10–3 (1.5 × 104) [94] |
| 68a | 5.3 × 10–5 (2 × 106) [92] | 68f | 4 × 10–3 (6 × 103) [94] |
| 68b | 1.8 × 10–6 (6.9 × 107) [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharapov, A.D.; Fatykhov, R.F.; Khalymbadzha, I.A.; Zyryanov, G.V.; Chupakhin, O.N.; Tsurkan, M.V. Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action. Int. J. Mol. Sci. 2023, 24, 2839. https://doi.org/10.3390/ijms24032839
Sharapov AD, Fatykhov RF, Khalymbadzha IA, Zyryanov GV, Chupakhin ON, Tsurkan MV. Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action. International Journal of Molecular Sciences. 2023; 24(3):2839. https://doi.org/10.3390/ijms24032839
Chicago/Turabian StyleSharapov, Ainur D., Ramil F. Fatykhov, Igor A. Khalymbadzha, Grigory V. Zyryanov, Oleg N. Chupakhin, and Mikhail V. Tsurkan. 2023. "Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action" International Journal of Molecular Sciences 24, no. 3: 2839. https://doi.org/10.3390/ijms24032839
APA StyleSharapov, A. D., Fatykhov, R. F., Khalymbadzha, I. A., Zyryanov, G. V., Chupakhin, O. N., & Tsurkan, M. V. (2023). Plant Coumarins with Anti-HIV Activity: Isolation and Mechanisms of Action. International Journal of Molecular Sciences, 24(3), 2839. https://doi.org/10.3390/ijms24032839