

Significance of Singlet Oxygen Molecule in Pathologies

 , ,
, ,  and
and
Abstract
1. Introduction
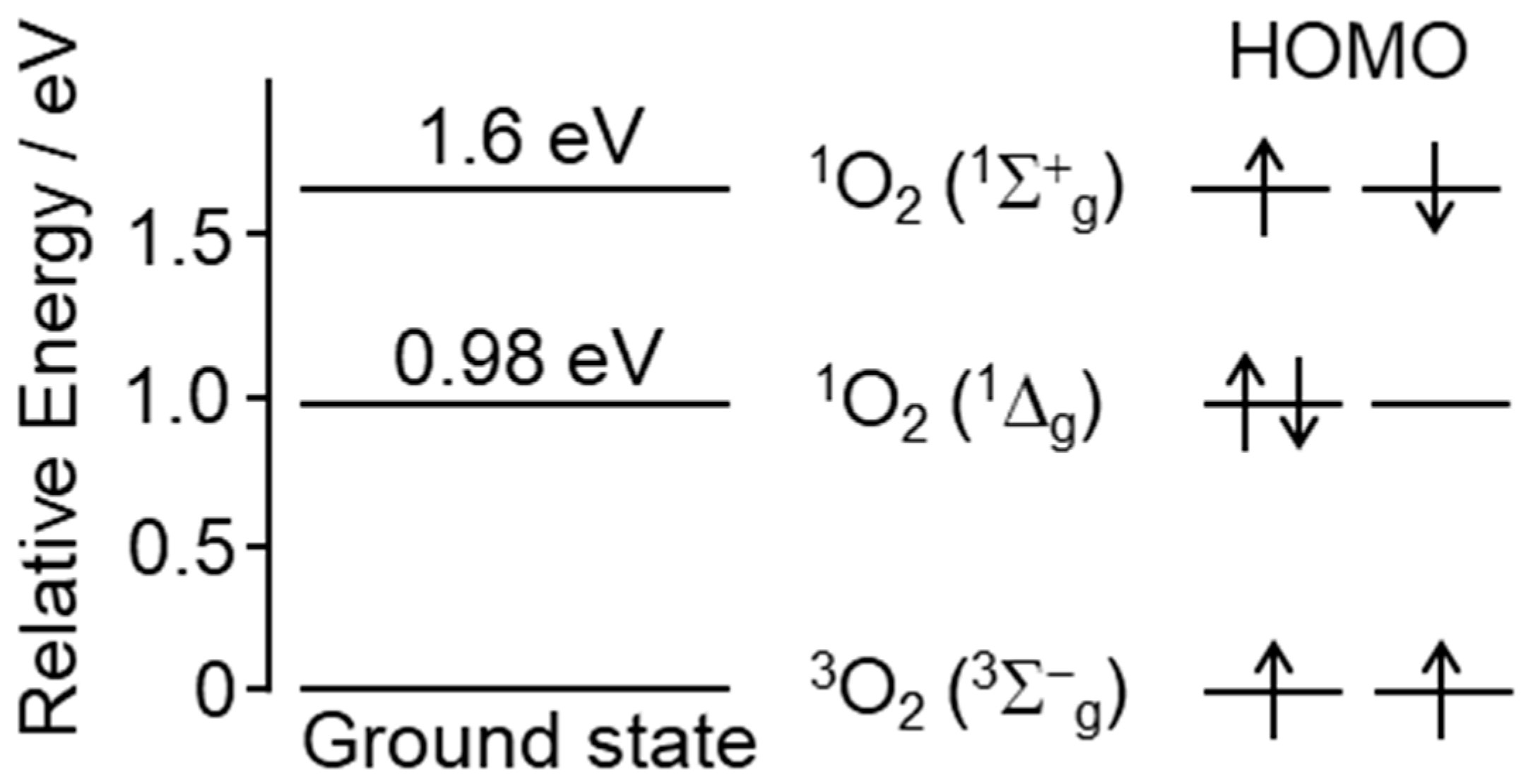
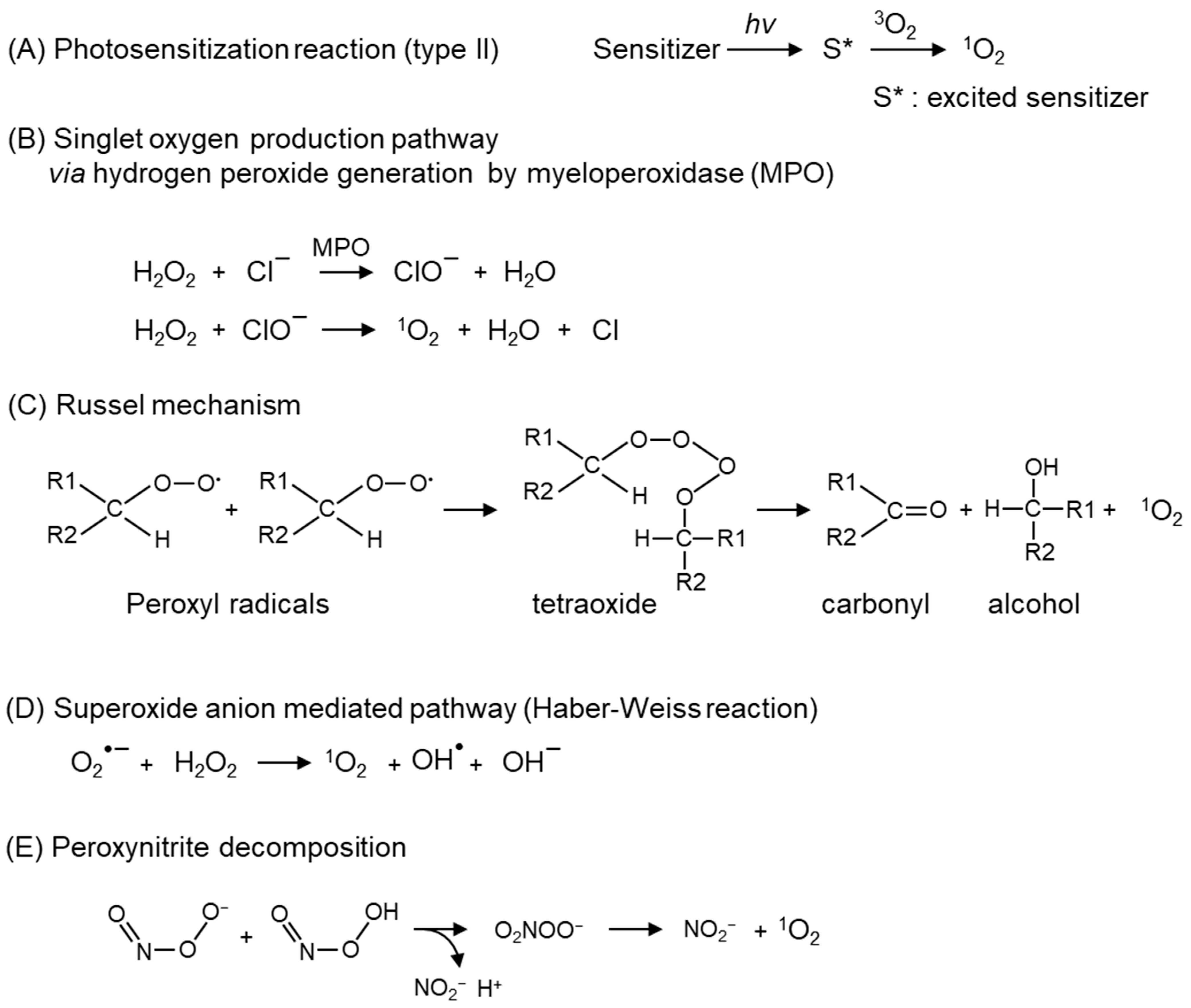
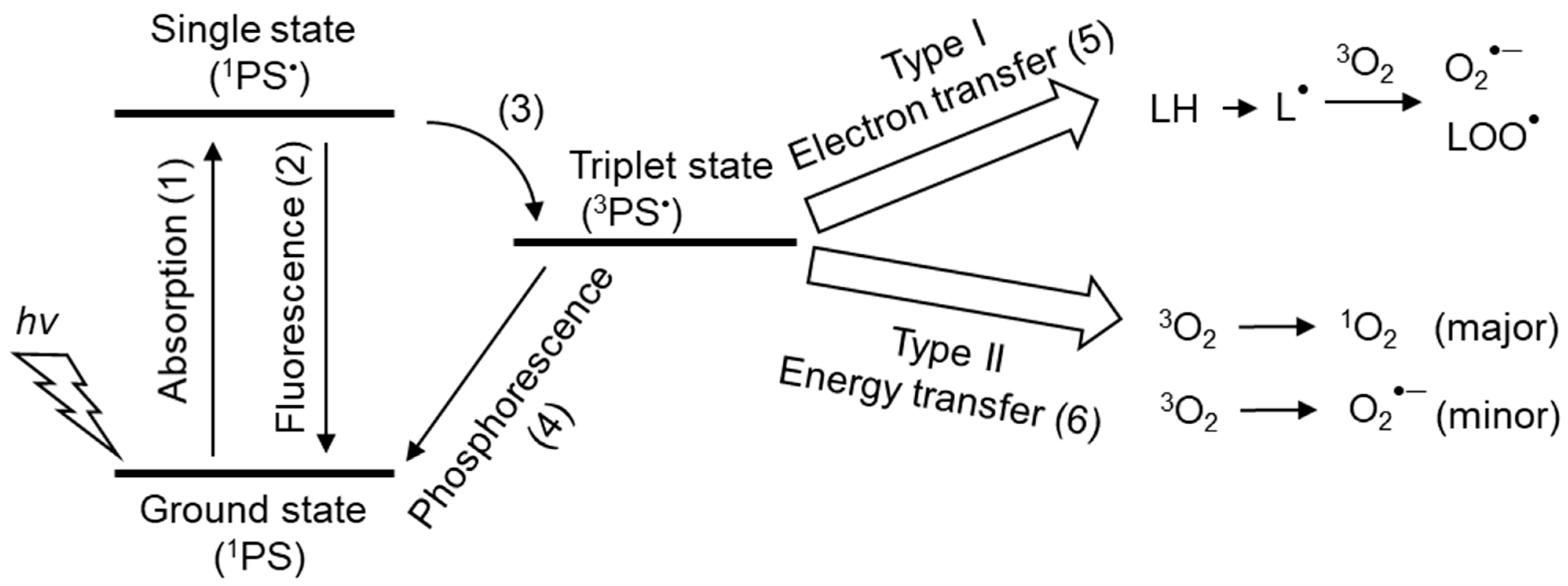
2. Chemical Properties and Possible Production of Singlet Oxygen In Vivo
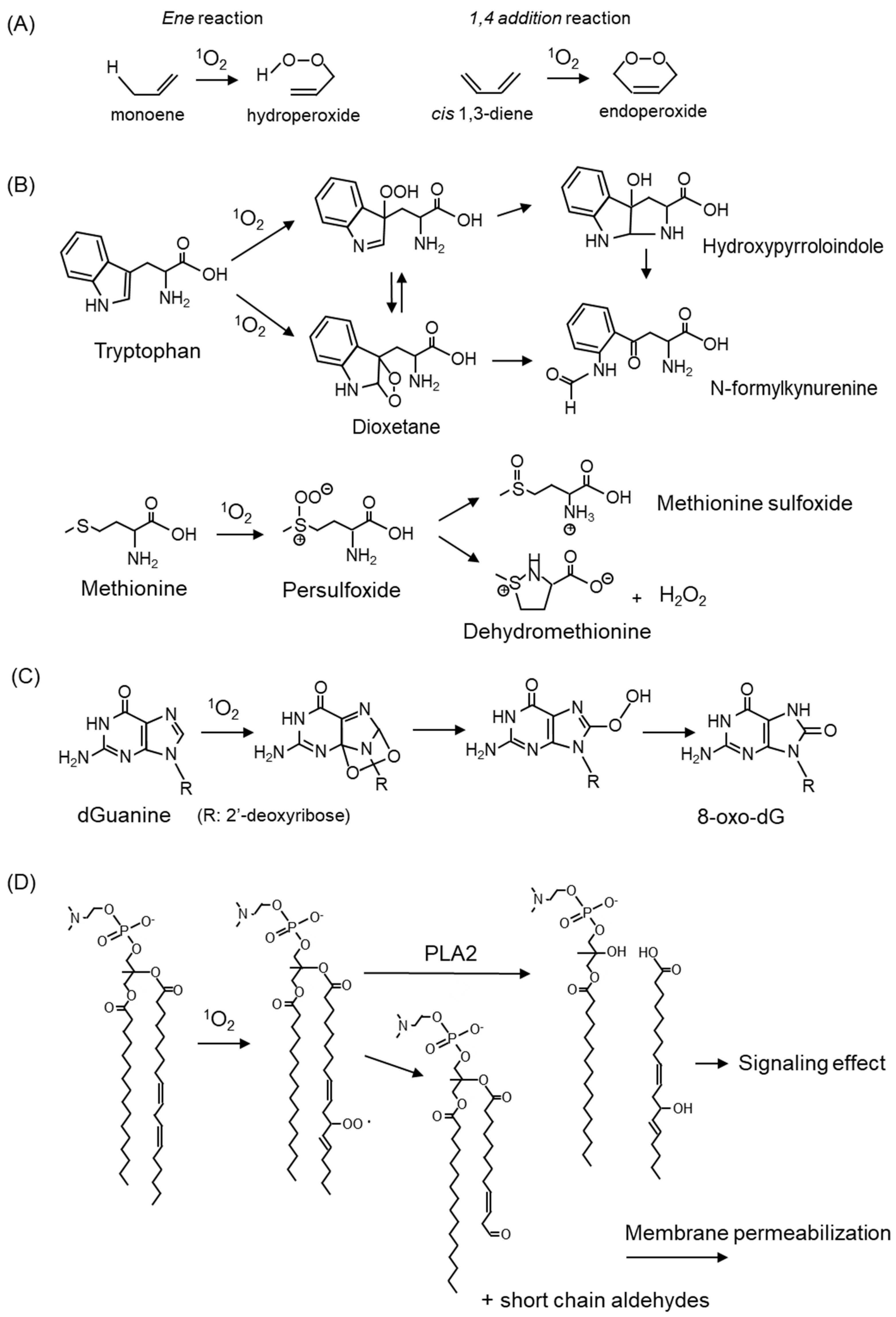
3. Damage to Biomolecules by 1O2-Mediated Oxidation
4. Detection Methods of 1O2 In Vivo and In Vitro
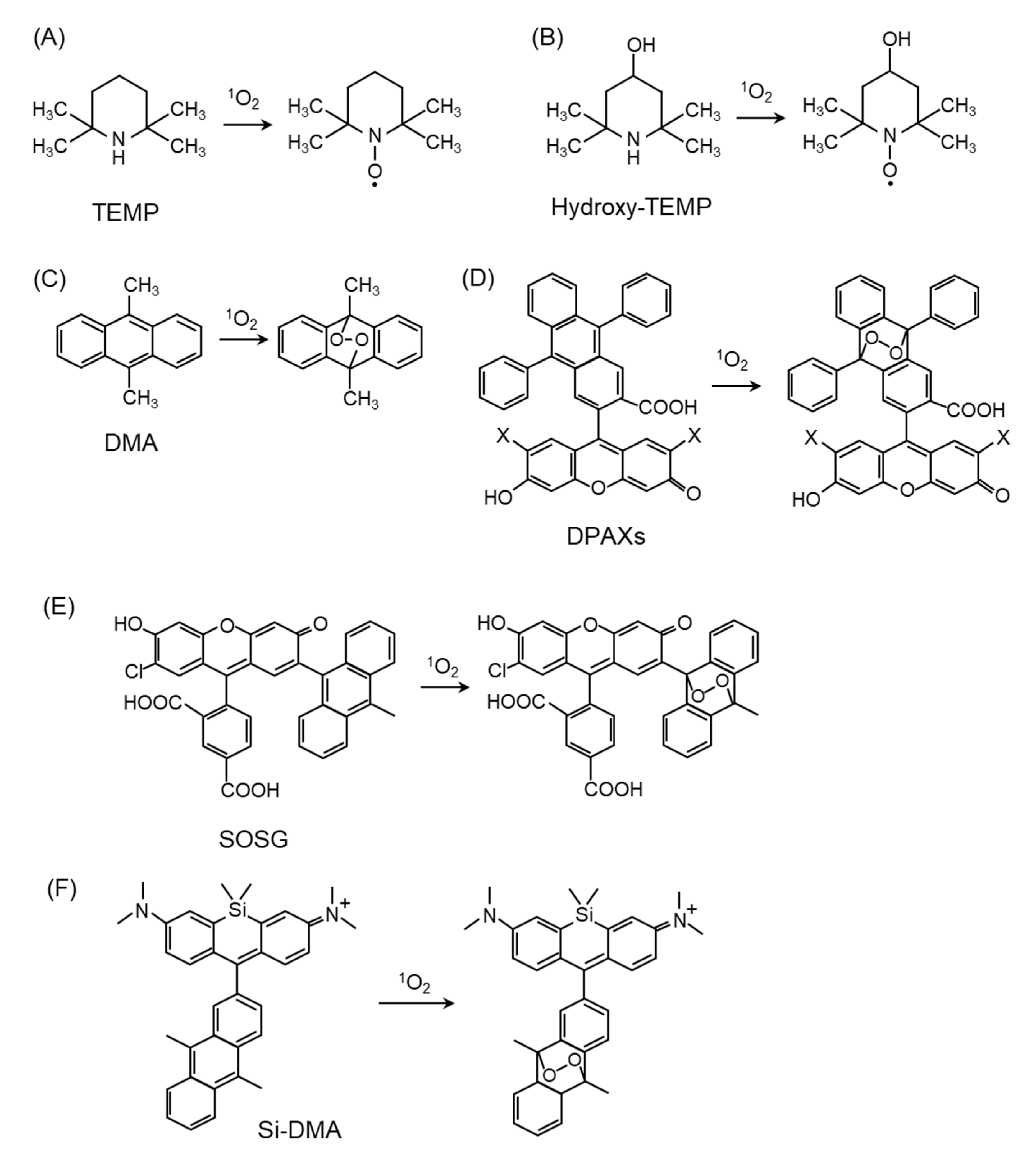
4.1. Direct Detection of 1O2
4.2. 1O2 Detection by Fluorescent Probes
4.3. Detection of 1O2-Mediated Peroxidation Products
5. Diseases Involving 1O2 and Their Pathophysiology
5.1. Skin Diseases
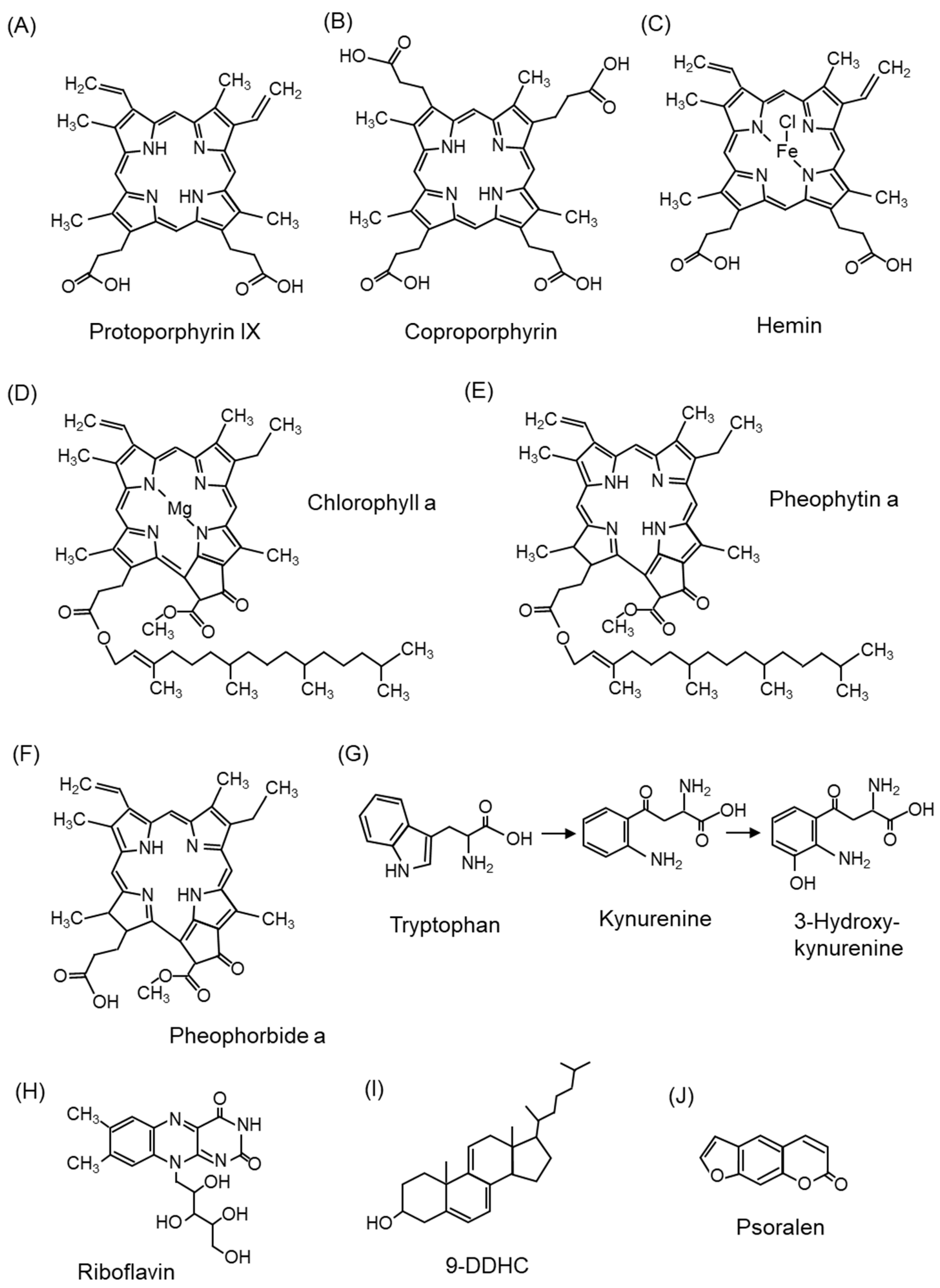
5.1.1. Photosensitizers
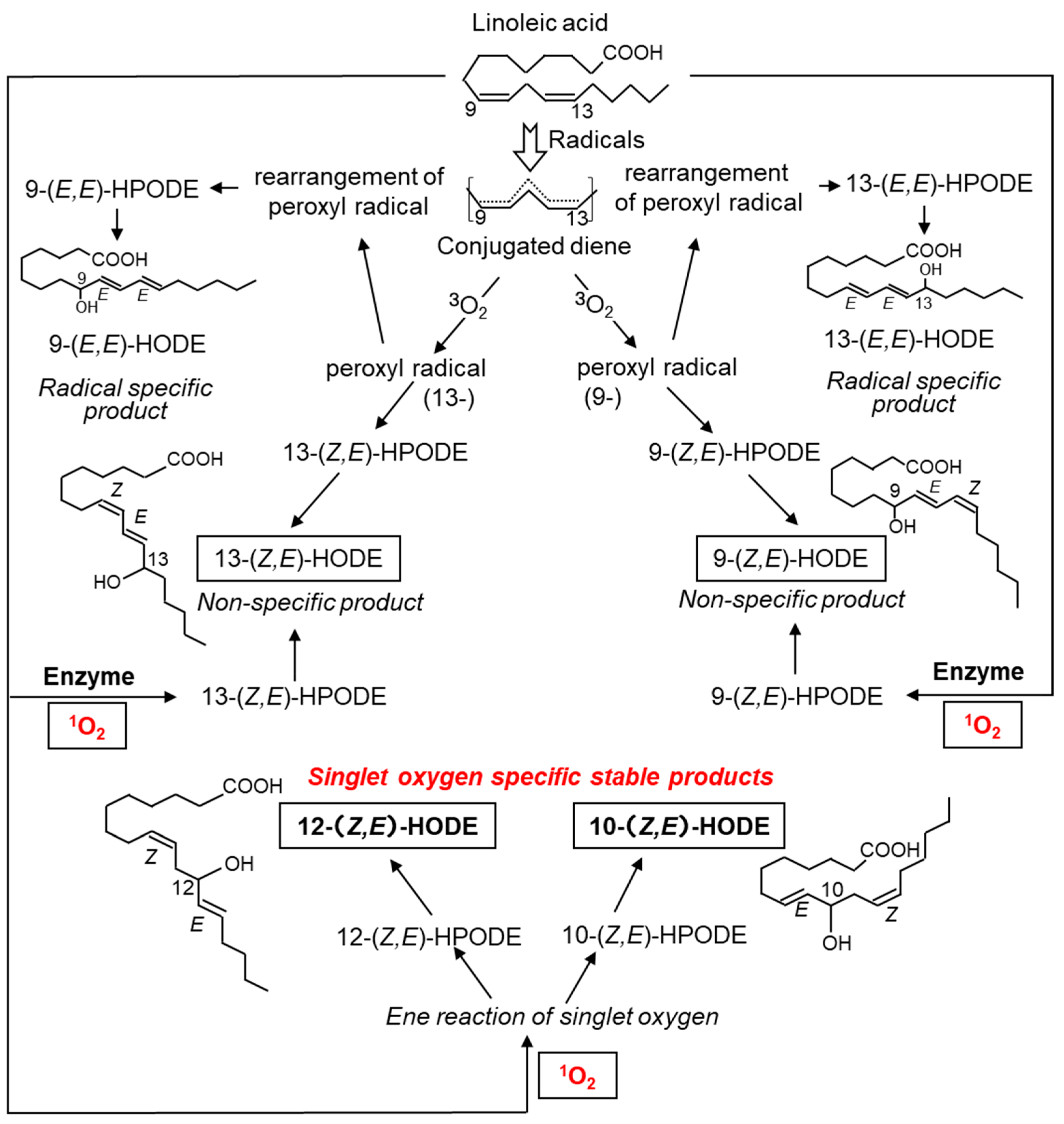
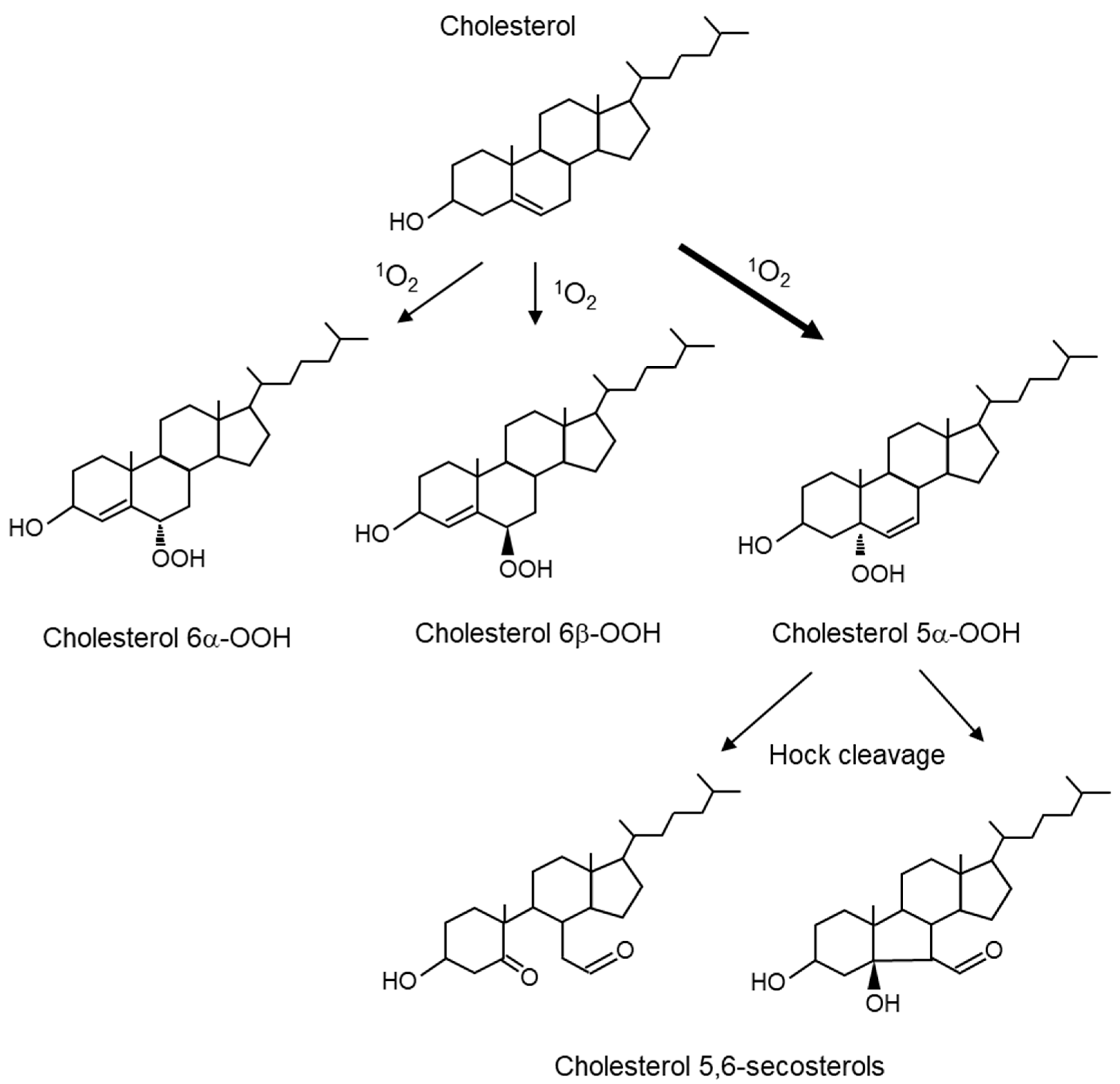
5.1.2. Lipids in the Skin and Lipid Peroxidation Products Mediated by 1O2

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1O2-Mediated Oxidation Product | Sample Species | Disease/Model | Ref. |
|---|---|---|---|
| Squalene monohydroperoxides | Human | Increased by exposure of human skin to tobacco smoke | [131] |
| Human | Sunlight exposure increases SqOOH in sebum | [132] | |
| Human | UV irradiation to human sebum | [133,134,135] | |
| Human | SqOOH reduction in sebum by application of cosmetics | [136] | |
| Human | Human skin surface lipids | [137] | |
| Human | Increased SqOOH in the scalp due to high dandruff | [138] | |
| Human | Detection using human fingerprints | [139] | |
| Human | Increase in SqOOH in the sebum due to ambient dust and ozone | [140] | |
| Human | Less SqOOH in sebum exposed to tobacco heating products and electronic cigarettes compared to cigarette smoke exposure | [129] | |
| Pig | Cigarette smoke exposure to pig skin | [141] | |
| Rabbit | UVA irradiation of rabbit ears | [127] | |
| Guinea pig | Carotene reduces the increase in SqOOH caused by UV irradiation of the skin | [142] | |
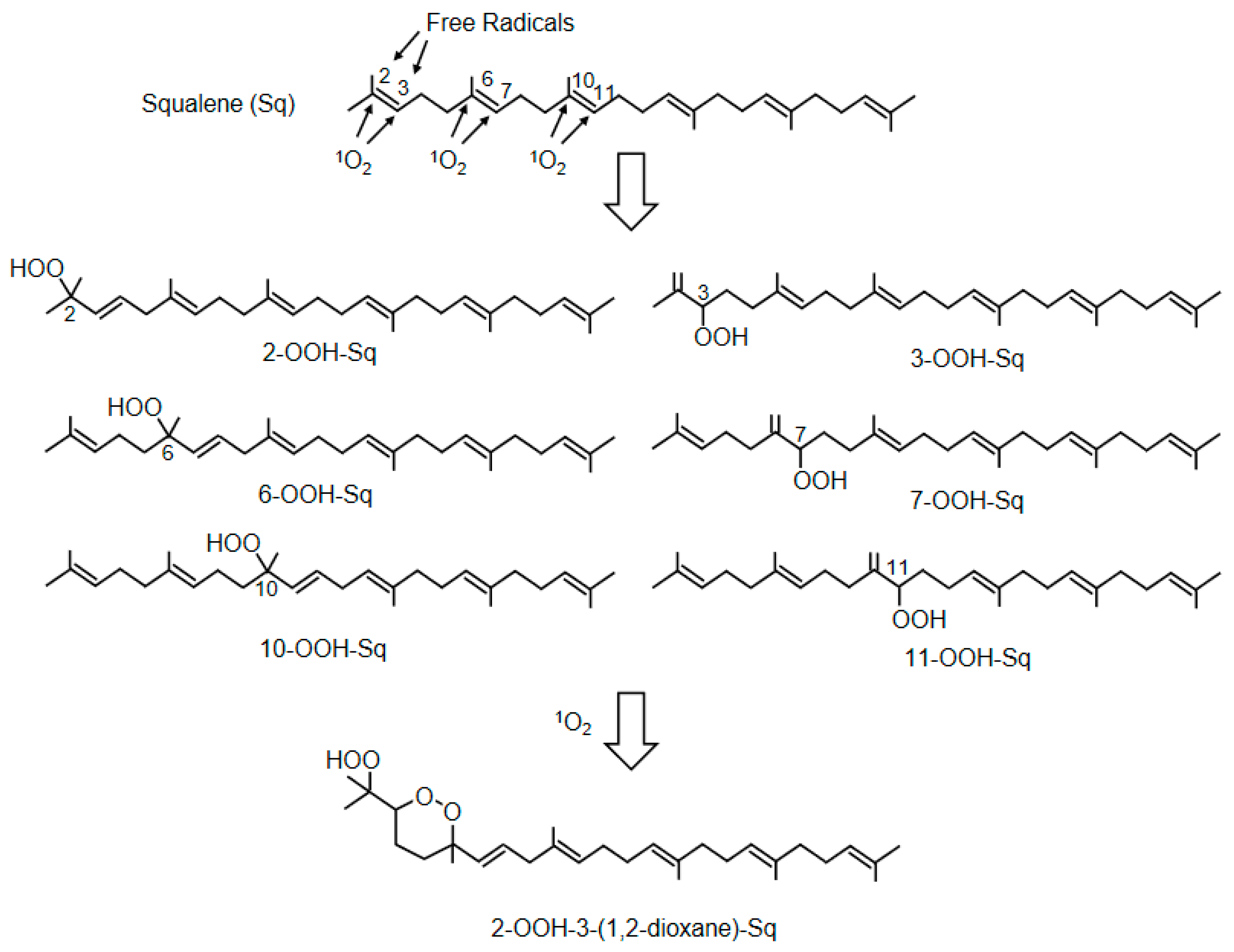
| 2-OOH-3-(1,2-dioxane)-squalene | Human | Identification of cyclic peroxides of SqOOH | [128] |
5.1.3. Skin Aging
5.1.4. Skin Cancer
5.1.5. Porphyria
5.1.6. Smith–Lemli–Opitz Syndrome and Statin-Induced Skin Disorders
5.1.7. Skin Disorders Caused by Exogenous Photosensitizers
5.1.8. Diagnosis and Evaluation of Skin Diseases by Measuring 1O2-Mediated Products
5.2. Ophthalmological Diseases
5.2.1. Structure and Optical Transparency of the Ocular Bulb
5.2.2. Photooxidative Stress
5.2.3. Compounds Protecting against 1O2 in the Macular Pigment and Lens
5.2.4. Blue Light Hazard
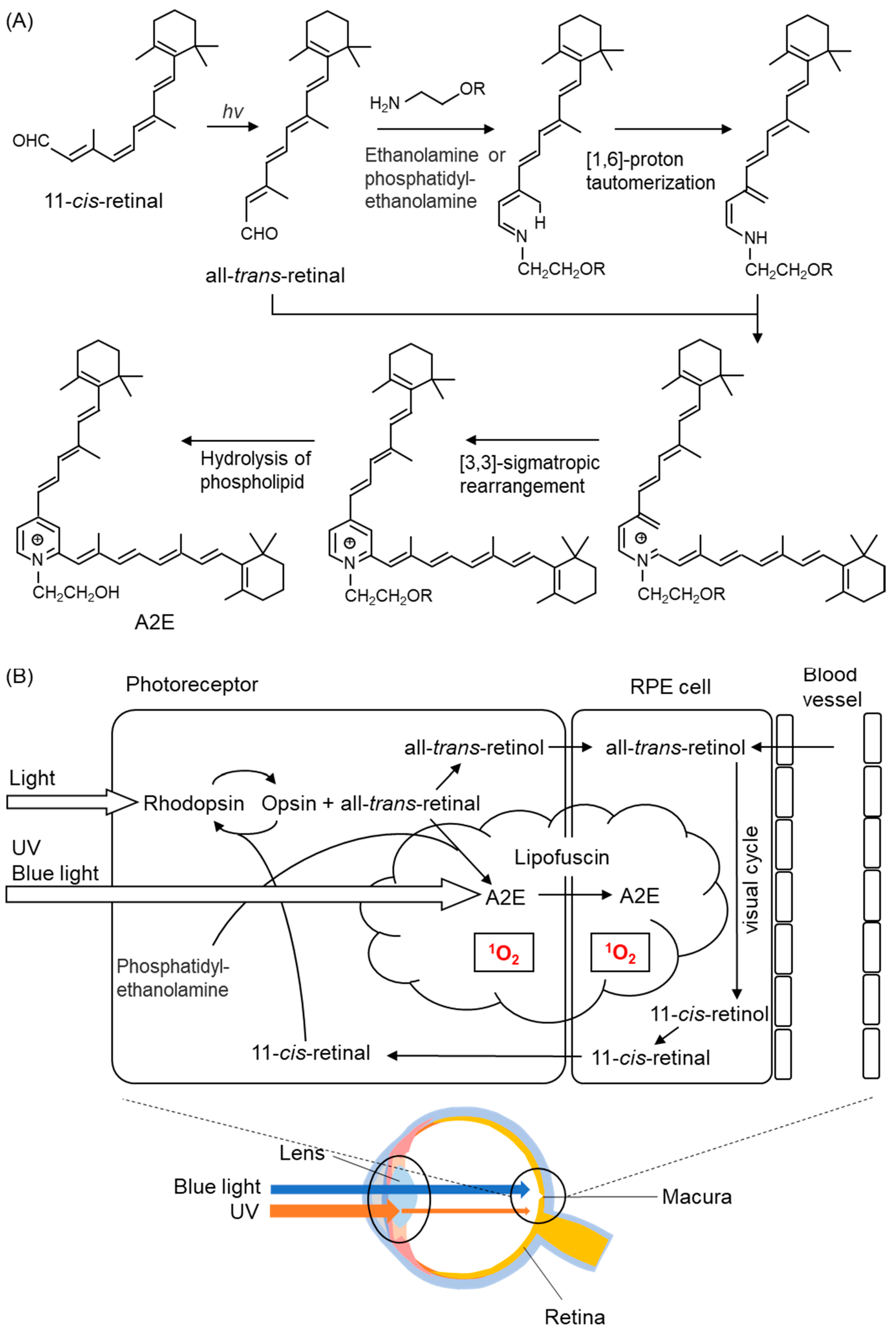
5.2.5. Age-Related Macular Degeneration
5.2.6. Glaucoma
5.2.7. Cataract
5.2.8. 1O2-Derived Peroxidation Products in the Diagnosis and Evaluation of Ophthalmological Diseases
5.3. Diabetes Mellitus
5.3.1. Biomarkers for Diabetes and Diabetic Complications
5.3.2. Diabetic Biomarkers of Lipid Peroxidation
5.4. Bronchial Asthma
6. 1O2 Scavengers
6.1. Carotenoids
6.2. Vitamin E (Tocopherol)
6.3. Other Compounds
7. Summary and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Z.H.; Yoshida, Y.; Saito, Y.; Noguchi, N.; Niki, E. Adaptive response induced by lipid peroxidation products in cell cultures. FEBS Lett. 2006, 580, 479–483. [Google Scholar] [CrossRef]
- Haugaard, N. Cellular mechanisms of oxygen toxicity. Physiol. Rev. 1968, 48, 311–373. [Google Scholar] [CrossRef]
- Gottlieb, S.F. Effect of hyperbaric oxygen on microorganisms. Annu. Rev. Microbiol. 1971, 25, 111–152. [Google Scholar] [CrossRef]
- Gilbert, D.L. Symposium on oxygen. Introduction: Oxygen and life. Anesthesiology 1972, 37, 100–111. [Google Scholar] [CrossRef]
- Chance, B.; Sies, H.; Boveris, A. Hydroperoxide metabolism in mammalian organs. Physiol. Rev. 1979, 59, 527–605. [Google Scholar] [CrossRef] [PubMed]
- Hackbarth, S.; Islam, W.; Fang, J.; Šubr, V.; Röder, B.; Etrych, T.; Maeda, H. Singlet oxygen phosphorescence detection in vivo identifies PDT-induced anoxia in solid tumors. Photochem. Photobiol. Sci. 2019, 18, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Collin, F. Chemical Basis of Reactive Oxygen Species Reactivity and Involvement in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lv, H.; Sun, Y.; Zu, G.; Zhang, X.; Song, Y.; Zhao, F.; Wang, J. New porphyrin photosensitizers-Synthesis, singlet oxygen yield, photophysical properties and application in PDT. Spectrochim. Acta. A Mol. Biomol. Spectrosc. 2022, 279, 121447. [Google Scholar] [CrossRef]
- You, Z.Q.; Hsu, C.P.; Fleming, G.R. Triplet-triplet energy-transfer coupling: Theory and calculation. J. Chem. Phys. 2006, 124, 044506. [Google Scholar] [CrossRef]
- Dzebo, D.; Moth-Poulsen, K.; Albinsson, B. Robust triplet-triplet annihilation photon upconversion by efficient oxygen scavenging. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2017, 16, 1327–1334. [Google Scholar] [CrossRef] [PubMed]
- Turro, N.J. Singlet oxygen and chemiluminescent organic reactions. In Modern Molecular Photochemistry; University Science Books: Melville, NY, USA, 1991; pp. 583–593. [Google Scholar]
- Hatz, S.; Poulsen, L.; Ogilby, P.R. Time-resolved singlet oxygen phosphorescence measurements from photosensitized experiments in single cells: Effects of oxygen diffusion and oxygen concentration. Photochem. Photobiol. 2008, 84, 1284–1290. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.W.; Sinks, L.E.; Breitenbach, T.; Schack, N.B.; Vinogradov, S.A.; Ogilby, P.R. Single cell responses to spatially controlled photosensitized production of extracellular singlet oxygen. Photochem. Photobiol. 2011, 87, 1077–1091. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Kolodieznyi, D.; Creeger, Y.; Ballou, B.; Bruchez, M.P. Subcellular Singlet Oxygen and Cell Death: Location Matters. Front. Chem. 2020, 8, 592941. [Google Scholar] [CrossRef] [PubMed]
- Denys, G.A.; Devoe, N.C.; Gudis, P.; May, M.; Allen, R.C.; Stephens, J.T., Jr. Mechanism of Microbicidal Action of E-101 Solution, a Myeloperoxidase-Mediated Antimicrobial, and Its Oxidative Products. Infect. Immun. 2019, 87, e00261-19. [Google Scholar] [CrossRef]
- Neale, T.J.; Ullrich, R.; Ojha, P.; Poczewski, H.; Verhoeven, A.J.; Kerjaschki, D. Reactive oxygen species and neutrophil respiratory burst cytochrome b558 are produced by kidney glomerular cells in passive Heymann nephritis. Proc. Natl. Acad. Sci. USA 1993, 90, 3645–3649. [Google Scholar] [CrossRef] [PubMed]
- Russell, G.A. Deuterium-isotope Effects in the Autoxidation of Aralkyl Hydrocarbons. Mechanism of the Interaction of PEroxy Radicals1. J. Am. Chem. Soc. 1957, 79, 3871–3877. [Google Scholar] [CrossRef]
- Gonçalves, L.C.P.; Massari, J.; Licciardi, S.; Prado, F.M.; Linares, E.; Klassen, A.; Tavares, M.F.M.; Augusto, O.; Di Mascio, P.; Bechara, E.J.H. Singlet oxygen generation by the reaction of acrolein with peroxynitrite via a 2-hydroxyvinyl radical intermediate. Free Radic. Biol. Med. 2020, 152, 83–90. [Google Scholar] [CrossRef]
- Sun, S.; Bao, Z.; Ma, H.; Zhang, D.; Zheng, X. Singlet oxygen generation from the decomposition of alpha-linolenic acid hydroperoxide by cytochrome c and lactoperoxidase. Biochemistry 2007, 46, 6668–6673. [Google Scholar] [CrossRef]
- Chen, T.; Cohen, D.; Itkin, M.; Malitsky, S.; Fluhr, R. Lipoxygenase functions in 1O2 production during root responses to osmotic stress. Plant Physiol. 2021, 185, 1638–1651. [Google Scholar] [CrossRef]
- Khan, A.U.; Kasha, M. Singlet molecular oxygen in the Haber-Weiss reaction. Proc. Natl. Acad. Sci. USA 1994, 91, 12365–12367. [Google Scholar] [CrossRef]
- Miyamoto, S.; Ronsein, G.E.; Corrêa, T.C.; Martinez, G.R.; Medeiros, M.H.; Di Mascio, P. Direct evidence of singlet molecular oxygen generation from peroxynitrate, a decomposition product of peroxynitrite. Dalton Trans. 2009, 29, 5720–5729. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G. The synergistic effect between hydrogen peroxide and nitrite, two long-lived molecular species from cold atmospheric plasma, triggers tumor cells to induce their own cell death. Redox Biol. 2019, 26, 101291. [Google Scholar] [CrossRef] [PubMed]
- Kanofsky, J.R.; Sima, P. Singlet oxygen production from the reactions of ozone with biological molecules. J. Biol. Chem. 1991, 266, 9039–9042. [Google Scholar] [CrossRef]
- Cerkovnik, J.; Erzen, E.; Koller, J.; Plesnicar, B. Evidence for HOOO radicals in the formation of alkyl hydrotrioxides (ROOOH) and hydrogen trioxide (HOOOH) in the ozonation of C-H bonds in hydrocarbons. J. Am. Chem. Soc. 2002, 124, 404–409. [Google Scholar] [CrossRef]
- Pershin, A.A.; Torbin, A.P.; Zagidullin, M.V.; Mebel, A.M.; Mikheyev, P.A.; Azyazov, V.N. Rate constants for collision-induced emission of O(2)(a(1)Δ(g)) with He, Ne, Ar, Kr, N(2), CO(2) and SF(6) as collisional partners. Phys. Chem. Chem. Phys. 2018, 20, 29677–29683. [Google Scholar] [CrossRef]
- Tao, F.; Gonzalez-Flecha, B.; Kobzik, L. Reactive oxygen species in pulmonary inflammation by ambient particulates. Free Radic. Biol. Med. 2003, 35, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Gurgueira, S.A.; Lawrence, J.; Coull, B.; Murthy, G.G.; González-Flecha, B. Rapid increases in the steady-state concentration of reactive oxygen species in the lungs and heart after particulate air pollution inhalation. Environ. Health Perspect. 2002, 110, 749–755. [Google Scholar] [CrossRef]
- Jensen, R.L.; Arnbjerg, J.; Ogilby, P.R. Reaction of singlet oxygen with tryptophan in proteins: A pronounced effect of the local environment on the reaction rate. J. Am. Chem. Soc. 2012, 134, 9820–9826. [Google Scholar] [CrossRef]
- Nascimento, R.O.; Prado, F.M.; Massafera, M.P.; Di Mascio, P.; Ronsein, G.E. Dehydromethionine is a common product of methionine oxidation by singlet molecular oxygen and hypohalous acids. Free Radic. Biol. Med. 2022, 187, 17–28. [Google Scholar] [CrossRef]
- Fouquerel, E.; Barnes, R.P.; Uttam, S.; Watkins, S.C.; Bruchez, M.P.; Opresko, P.L. Targeted and Persistent 8-Oxoguanine Base Damage at Telomeres Promotes Telomere Loss and Crisis. Mol. Cell 2019, 75, 117–130.e116. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Sun, Y.; Tsai, M.; Zhou, W.; Liu, J. Singlet O(2) Oxidation of a Deprotonated Guanine-Cytosine Base Pair and Its Entangling with Intra-Base-Pair Proton Transfer. Chemphyschem 2018, 19, 2645–2654. [Google Scholar] [CrossRef]
- Kato, S.; Shimizu, N.; Otoki, Y.; Ito, J.; Sakaino, M.; Sano, T.; Takeuchi, S.; Imagi, J.; Nakagawa, K. Determination of acrolein generation pathways from linoleic acid and linolenic acid: Increment by photo irradiation. NPJ Sci. Food 2022, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, A.; Ito, J.; Shimizu, N.; Kato, S.; Kobayashi, E.; Ohnari, H.; Sakata, O.; Naru, E.; Nakagawa, K. Linoleic acid and squalene are oxidized by discrete oxidation mechanisms in human sebum. Ann. N. Y. Acad. Sci. 2021, 1500, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.; Pattison, D.I.; Davies, J.B.; Anderson, R.F.; Lopez-Alarcon, C.; Davies, M.J. Superoxide radicals react with peptide-derived tryptophan radicals with very high rate constants to give hydroperoxides as major products. Free Radic. Biol. Med. 2018, 118, 126–136. [Google Scholar] [CrossRef]
- Moe, M.M.; Tsai, M.; Liu, J. Singlet Oxygen Oxidation of the Radical Cations of 8-Oxo-2′-deoxyguanosine and Its 9-Methyl Analogue: Dynamics, Potential Energy Surface, and Products Mediated by C5-O(2) -Addition. Chempluschem 2021, 86, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Monzo-Beltran, L.; Vazquez-Tarragón, A.; Cerdà, C.; Garcia-Perez, P.; Iradi, A.; Sánchez, C.; Climent, B.; Tormos, C.; Vázquez-Prado, A.; Girbés, J.; et al. One-year follow-up of clinical, metabolic and oxidative stress profile of morbid obese patients after laparoscopic sleeve gastrectomy. 8-oxo-dG as a clinical marker. Redox Biol. 2017, 12, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Ponce, M.A.; Ramirez, J.A.; Galagovsky, L.R.; Gros, E.G.; Erra-Balsells, R. Singlet-oxygen ene reaction with 3β-substituted stigmastanes. An alternative pathway for the classical Schenck rearrangement. J. Chem. Soc. Perkin Trans. 2000, 2, 2351–2358. [Google Scholar] [CrossRef]
- Samuel, D.; Norrell, K.; Hilmey, D.G. Novel ring chemistry of vitamin B6 with singlet oxygen and an activated ene: Isolated products and identified intermediates suggesting an operable [3 + 2] cycloaddition. Org. Biomol. Chem. 2012, 10, 7278–7281. [Google Scholar] [CrossRef]
- Petrou, A.L.; Petrou, P.L.; Ntanos, T.; Liapis, A. A Possible Role for Singlet Oxygen in the Degradation of Various Antioxidants. A Meta-Analysis and Review of Literature Data. Antioxidants 2018, 7, 35. [Google Scholar] [CrossRef]
- Zeinali, N.; Oluwoye, I.; Altarawneh, M.K.; Almatarneh, M.H.; Dlugogorski, B.Z. Probing the Reactivity of Singlet Oxygen with Cyclic Monoterpenes. ACS Omega 2019, 4, 14040–14048. [Google Scholar] [CrossRef]
- Morales, J.; Günther, G.; Zanocco, A.L.; Lemp, E. Singlet oxygen reactions with flavonoids. A theoretical-experimental study. PLoS ONE 2012, 7, e40548. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Tsai, H.C.; Yu, P.L.; Chung, K.R. A Major Facilitator Superfamily Transporter-Mediated Resistance to Oxidative Stress and Fungicides Requires Yap1, Skn7, and MAP Kinases in the Citrus Fungal Pathogen Alternaria alternata. PLoS ONE 2017, 12, e0169103. [Google Scholar] [CrossRef] [PubMed]
- Di Mascio, P.; Martinez, G.R.; Miyamoto, S.; Ronsein, G.E.; Medeiros, M.H.G.; Cadet, J. Singlet Molecular Oxygen Reactions with Nucleic Acids, Lipids, and Proteins. Chem. Rev. 2019, 119, 2043–2086. [Google Scholar] [CrossRef] [PubMed]
- Kanofsky, J.R. Singlet oxygen production by biological systems. Chem. Biol. Interact. 1989, 70, 1–28. [Google Scholar] [CrossRef]
- Dong, S.; Xu, J.; Jia, T.; Xu, M.; Zhong, C.; Yang, G.; Li, J.; Yang, D.; He, F.; Gai, S.; et al. Upconversion-mediated ZnFe2O4 nanoplatform for NIR-enhanced chemodynamic and photodynamic therapy. Chem. Sci. 2019, 10, 4259–4271. [Google Scholar] [CrossRef]
- Choi, J.; Kim, S.Y. Synthesis of near-infrared-responsive hexagonal-phase upconversion nanoparticles with controllable shape and luminescence efficiency for theranostic applications. J. Biomater. Appl. 2022, 37, 646–658. [Google Scholar] [CrossRef]
- Takajo, T.; Kurihara, Y.; Iwase, K.; Miyake, D.; Tsuchida, K.; Anzai, K. Basic Investigations of Singlet Oxygen Detection Systems with ESR for the Measurement of Singlet Oxygen Quenching Activities. Chem. Pharm. Bull. 2020, 68, 150–154. [Google Scholar] [CrossRef]
- Yu, Y.Y.; Quan, W.Z.; Cao, Y.; Niu, Q.; Lu, Y.; Xiao, X.; Cheng, L. Boosting the singlet oxygen production from H2O2 activation with highly dispersed Co-N-graphene for pollutant removal. RSC Adv 2022, 12, 17864–17872. [Google Scholar] [CrossRef]
- Ruiz-González, R.; Bresolí-Obach, R.; Gulías, Ò.; Agut, M.; Savoie, H.; Boyle, R.W.; Nonell, S.; Giuntini, F. NanoSOSG: A Nanostructured Fluorescent Probe for the Detection of Intracellular Singlet Oxygen. Angew Chem. Int. Ed. Engl. 2017, 56, 2885–2888. [Google Scholar] [CrossRef]
- Liu, H.; Carter, P.J.H.; Laan, A.C.; Eelkema, R.; Denkova, A.G. Singlet Oxygen Sensor Green is not a Suitable Probe for (1)O(2) in the Presence of Ionizing Radiation. Sci. Rep. 2019, 9, 8393. [Google Scholar] [CrossRef]
- Girotti, A.W.; Korytowski, W. Cholesterol as a singlet oxygen detector in biological systems. Methods Enzymol. 2000, 319, 85–100. [Google Scholar] [PubMed]
- Miyamoto, S.; Di Mascio, P. Lipid hydroperoxides as a source of singlet molecular oxygen. Subcell Biochem. 2014, 77, 3–20. [Google Scholar] [PubMed]
- Torinuki, W.; Miura, T. Singlet oxygen emission at 1270 nm in protoporphyrin solution excited by argon laser. Tohoku J. Exp. Med. 1983, 140, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Kanofsky, J.R. Direct observation of singlet oxygen phosphorescence at 1270 nm from L1210 leukemia cells exposed to polyporphyrin and light. Arch. Biochem. Biophys. 1991, 286, 70–75. [Google Scholar] [CrossRef]
- Baker, A.; Kanofsky, J.R. Time-resolved studies of singlet-oxygen emission from L1210 leukemia cells labeled with 5-(N-hexadecanoyl)amino eosin. A comparison with a one-dimensional model of singlet-oxygen diffusion and quenching. Photochem. Photobiol. 1993, 57, 720–727. [Google Scholar] [CrossRef]
- Oelckers, S.; Ziegler, T.; Michler, I.; Roder, B. Time-resolved detection of singlet oxygen luminescence in red-cell ghost suspensions: Concerning a signal component that can be attributed to 1O2 luminescence from the inside of a native membrane. J. Photochem. Photobiol. B 1999, 53, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, C.; Schmidt, R. Physical mechanisms of generation and deactivation of singlet oxygen. Chem. Rev. 2003, 103, 1685–1757. [Google Scholar] [CrossRef] [PubMed]
- Niedre, M.; Patterson, M.S.; Wilson, B.C. Direct near-infrared luminescence detection of singlet oxygen generated by photodynamic therapy in cells in vitro and tissues in vivo. Photochem. Photobiol. 2002, 75, 382–391. [Google Scholar] [CrossRef]
- Nosaka, Y.; Nosaka, A.Y. Generation and Detection of Reactive Oxygen Species in Photocatalysis. Chem. Rev. 2017, 117, 11302–11336. [Google Scholar] [CrossRef]
- Lion, Y.; Delmelle, M.; van de Vorst, A. New method of detecting singlet oxygen production. Nature 1976, 263, 442–443. [Google Scholar] [CrossRef]
- Moan, J.; Wold, E. Detection of singlet oxygen production by ESR. Nature 1979, 279, 450–451. [Google Scholar] [CrossRef]
- Dzwigaj, S.; Pezerat, H. Singlet oxygen-trapping reaction as a method of (1)O2 detection: Role of some reducing agents. Free Radic. Res. 1995, 23, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Victória, H.F.V.; Ferreira, D.C.; Filho, J.B.G.; Martins, D.C.S.; Pinheiro, M.V.B.; Sáfar, G.A.M.; Krambrock, K. Detection of singlet oxygen by EPR: The instability of the nitroxyl radicals. Free Radic. Biol. Med. 2022, 180, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.Y.; Choi, D.S. Electron spin resonance and luminescence spectroscopic observation and kinetic study of chemical and physical singlet oxygen quenching by resveratrol in methanol. J. Agric. Food Chem. 2010, 58, 11888–11895. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Maehata, Y.; Kawamura, Y.; Kusubata, M.; Hattori, S.; Tanaka, K.; Miyamoto, C.; Yoshino, F.; Yoshida, A.; Wada-Takahashi, S.; et al. Direct assessments of the antioxidant effects of the novel collagen peptide on reactive oxygen species using electron spin resonance spectroscopy. J. Pharmacol. Sci. 2011, 116, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Soh, N.; Katayama, Y.; Maeda, M. A fluorescent probe for monitoring nitric oxide production using a novel detection concept. Analyst 2001, 126, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Miura, T.; Umezawa, N.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Rational design of fluorescein-based fluorescence probes. Mechanism-based design of a maximum fluorescence probe for singlet oxygen. J. Am. Chem. Soc. 2001, 123, 2530–2536. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Fernandes, E.; Lima, J.L. Fluorescence probes used for detection of reactive oxygen species. J. Biochem. Biophys. Methods 2005, 65, 45–80. [Google Scholar] [CrossRef]
- Corey, E.J.; Taylor, W.C. Study of the peroxidation of organic compounds by externally generated singlet oxygen molecules. J. Am. Chem. Soc. 1964, 86, 3881–3882. [Google Scholar] [CrossRef]
- Francis Wilkinson, W.P.H.; Ross, A.B. Rate Constants for the Decay and Reactions of the Lowest Electronically Excited Singlet State of Molecular Oxygen in Solution. J. Phys. Chem. Ref. 1995, 24, 663–1021. [Google Scholar]
- Umezawa, N.; Tanaka, K.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Novel Fluorescent Probes for Singlet Oxygen. Angew. Chem. Int. Ed. Engl. 1999, 38, 2899–2901. [Google Scholar] [CrossRef]
- Flors, C.; Fryer, M.J.; Waring, J.; Reeder, B.; Bechtold, U.; Mullineaux, P.M.; Nonell, S.; Wilson, M.T.; Baker, N.R. Imaging the production of singlet oxygen in vivo using a new fluorescent sensor, Singlet Oxygen Sensor Green. J. Exp. Bot. 2006, 57, 1725–1734. [Google Scholar] [CrossRef]
- Price, M.; Reiners, J.J.; Santiago, A.M.; Kessel, D. Monitoring singlet oxygen and hydroxyl radical formation with fluorescent probes during photodynamic therapy. Photochem. Photobiol. 2009, 85, 1177–1181. [Google Scholar] [CrossRef]
- Ragas, X.; Jimenez-Banzo, A.; Sanchez-Garcia, D.; Batllori, X.; Nonell, S. Singlet oxygen photosensitisation by the fluorescent probe Singlet Oxygen Sensor Green. Chem. Commun. 2009, 28, 2920–2922. [Google Scholar] [CrossRef]
- Kim, S.; Tachikawa, T.; Fujitsuka, M.; Majima, T. Far-red fluorescence probe for monitoring singlet oxygen during photodynamic therapy. J. Am. Chem. Soc. 2014, 136, 11707–11715. [Google Scholar] [CrossRef]
- Murotomi, K.; Umeno, A.; Sugino, S.; Yoshida, Y. Quantitative kinetics of intracellular singlet oxygen generation using a fluorescence probe. Sci. Rep. 2020, 10, 10616. [Google Scholar] [CrossRef]
- Chen, B.; Yang, Y.; Wang, Y.; Yan, Y.; Wang, Z.; Yin, Q.; Zhang, Q.; Wang, Y. Precise Monitoring of Singlet Oxygen in Specific Endocytic Organelles by Super-pH-Resolved Nanosensors. ACS Appl. Mater. Interfaces 2021, 13, 18533–18544. [Google Scholar] [CrossRef]
- Nath, P.; Hamadna, S.S.; Karamchand, L.; Foster, J.; Kopelman, R.; Amar, J.G.; Ray, A. Intracellular detection of singlet oxygen using fluorescent nanosensors. Analyst 2021, 146, 3933–3941. [Google Scholar] [CrossRef]
- Niki, E. Lipid peroxidation products as oxidative stress biomarkers. Biofactors 2008, 34, 171–180. [Google Scholar] [CrossRef]
- Niki, E. Assessment of antioxidant capacity in vitro and in vivo. Free Radic. Biol. Med. 2010, 49, 503–515. [Google Scholar] [CrossRef]
- Akazawa-Ogawa, Y.; Shichiri, M.; Nishio, K.; Yoshida, Y.; Niki, E.; Hagihara, Y. Singlet-oxygen-derived products from linoleate activate Nrf2 signaling in skin cells. Free Radic. Biol. Med. 2015, 79, 164–175. [Google Scholar] [CrossRef]
- Murotomi, K.; Umeno, A.; Yasunaga, M.; Shichiri, M.; Ishida, N.; Abe, H.; Yoshida, Y.; Nakajima, Y. Switching from singlet-oxygen-mediated oxidation to free-radical-mediated oxidation in the pathogenesis of type 2 diabetes in model mouse. Free Radic. Res. 2015, 49, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Cesarini, E.; Betti, M.; Galluzzi, L.; Galli, L.; Tippins, J.; Zerbinati, C.; Papa, S.; Iuliano, L. 7-Ketocholesterol and 5,6-secosterol induce human endothelial cell dysfunction by differential mechanisms. Steroids 2015, 99, 204–211. [Google Scholar] [CrossRef]
- Dantas, L.S.; Chaves-Filho, A.B.; Coelho, F.R.; Genaro-Mattos, T.C.; Tallman, K.A.; Porter, N.A.; Augusto, O.; Miyamoto, S. Cholesterol secosterol aldehyde adduction and aggregation of Cu,Zn-superoxide dismutase: Potential implications in ALS. Redox Biol. 2018, 19, 105–115. [Google Scholar] [CrossRef]
- Niki, E.; Yoshida, Y. Biomarkers for oxidative stress: Measurement, validation, and application. J. Med. Invest. 2005, 52, 228–230. [Google Scholar] [CrossRef]
- Yoshida, Y.; Niki, E. Bio-markers of lipid peroxidation in vivo: Hydroxyoctadecadienoic acid and hydroxycholesterol. Biofactors 2006, 27, 195–202. [Google Scholar] [CrossRef]
- Yoshida, Y.; Hayakawa, M.; Habuchi, Y.; Itoh, N.; Niki, E. Evaluation of lipophilic antioxidant efficacy in vivo by the biomarkers hydroxyoctadecadienoic acid and isoprostane. Lipids 2007, 42, 463–472. [Google Scholar] [CrossRef]
- Yoshida, Y.; Kodai, S.; Takemura, S.; Minamiyama, Y.; Niki, E. Simultaneous measurement of F2-isoprostane, hydroxyoctadecadienoic acid, hydroxyeicosatetraenoic acid, and hydroxycholesterols from physiological samples. Anal. Biochem. 2008, 379, 105–115. [Google Scholar] [CrossRef]
- Yoshida, Y.; Umeno, A.; Shichiri, M. Lipid peroxidation biomarkers for evaluating oxidative stress and assessing antioxidant capacity in vivo. J. Clin. Biochem. Nutr. 2013, 52, 9–16. [Google Scholar] [CrossRef]
- Yoshida, Y.; Umeno, A.; Akazawa, Y.; Shichiri, M.; Murotomi, K.; Horie, M. Chemistry of lipid peroxidation products and their use as biomarkers in early detection of diseases. J. Oleo Sci. 2015, 64, 347–356. [Google Scholar] [CrossRef]
- Umeno, A.; Shichiri, M.; Ishida, N.; Hashimoto, Y.; Abe, K.; Kataoka, M.; Yoshino, K.; Hagihara, Y.; Aki, N.; Funaki, M.; et al. Singlet oxygen induced products of linoleates, 10- and 12-(Z,E)-hydroxyoctadecadienoic acids (HODE), can be potential biomarkers for early detection of type 2 diabetes. PLoS ONE 2013, 8, e63542. [Google Scholar] [CrossRef] [PubMed]
- Umeno, A.; Yoshino, K.; Hashimoto, Y.; Shichiri, M.; Kataoka, M.; Yoshida, Y. Multi-Biomarkers for Early Detection of Type 2 Diabetes, Including 10- and 12-(Z,E)-Hydroxyoctadecadienoic Acids, Insulin, Leptin, and Adiponectin. PLoS ONE 2015, 10, e0130971. [Google Scholar] [CrossRef] [PubMed]
- Umeno, A.; Tanito, M.; Kaidzu, S.; Takai, Y.; Horie, M.; Yoshida, Y. Comprehensive measurements of hydroxylinoleate and hydroxyarachidonate isomers in blood samples from primary open-angle glaucoma patients and controls. Sci. Rep. 2019, 9, 2171. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, U.; Spiteller, G. The course of enzymatically induced lipid peroxidation in homogenized porcine kidney tissue. Z. Naturforsch C J. Biosci. 1998, 53, 431–437. [Google Scholar] [CrossRef]
- Horie, M.; Miura, T.; Hirakata, S.; Hosoyama, A.; Sugino, S.; Umeno, A.; Murotomi, K.; Yoshida, Y.; Koike, T. Comparative analysis of the intestinal flora in type 2 diabetes and nondiabetic mice. Exp. Anim. 2017, 66, 405–416. [Google Scholar] [CrossRef]
- Ogawa, H.; Azuma, M.; Umeno, A.; Shimizu, M.; Murotomi, K.; Yoshida, Y.; Nishioka, Y.; Tsuneyama, K. Singlet oxygen -derived nerve growth factor exacerbates airway hyperresponsiveness in a mouse model of asthma with mixed inflammation. Allergol. Int. 2022, 71, 395–404. [Google Scholar] [CrossRef]
- Adachi, J.; Asano, M.; Naito, T.; Ueno, Y.; Imamichi, H.; Tatsuno, Y. Cholesterol hydroperoxides in erythrocyte membranes of alcoholic patients. Alcohol. Clin. Exp. Res. 1999, 23, 96s–100s. [Google Scholar] [CrossRef]
- Yamazaki, S.; Ozawa, N.; Hiratsuka, A.; Watanabe, T. Quantitative determination of cholesterol 5alpha-, 7alpha-, and 7beta-hydroperoxides in rat skin. Free Radic. Biol. Med. 1999, 27, 110–118. [Google Scholar] [CrossRef]
- Minami, Y.; Yokoi, S.; Setoyama, M.; Bando, N.; Takeda, S.; Kawai, Y.; Terao, J. Combination of TLC blotting and gas chromatography-mass spectrometry for analysis of peroxidized cholesterol. Lipids 2007, 42, 1055–1063. [Google Scholar] [CrossRef]
- Minami, Y.; Kawabata, K.; Kubo, Y.; Arase, S.; Hirasaka, K.; Nikawa, T.; Bando, N.; Kawai, Y.; Terao, J. Peroxidized cholesterol-induced matrix metalloproteinase-9 activation and its suppression by dietary beta-carotene in photoaging of hairless mouse skin. J. Nutr. Biochem. 2009, 20, 389–398. [Google Scholar] [CrossRef]
- Terao, J.; Minami, Y.; Bando, N. Singlet molecular oxygen-quenching activity of carotenoids: Relevance to protection of the skin from photoaging. J. Clin. Biochem. Nutr. 2011, 48, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wentworth, P., Jr.; Nieva, J.; Takeuchi, C.; Galve, R.; Wentworth, A.D.; Dilley, R.B.; DeLaria, G.A.; Saven, A.; Babior, B.M.; Janda, K.D.; et al. Evidence for ozone formation in human atherosclerotic arteries. Science 2003, 302, 1053–1056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Powers, E.T.; Nieva, J.; Huff, M.E.; Dendle, M.A.; Bieschke, J.; Glabe, C.G.; Eschenmoser, A.; Wentworth, P., Jr.; Lerner, R.A.; et al. Metabolite-initiated protein misfolding may trigger Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 4752–4757. [Google Scholar] [CrossRef] [PubMed]
- Elshafei, M.E.; Minamiyama, Y.; Ichikawa, H. Singlet oxygen from endoperoxide initiates an intracellular reactive oxygen species release in HaCaT keratinocytes. J. Clin. Biochem. Nutr. 2022, 71, 198–205. [Google Scholar] [CrossRef]
- Umar, S.A.; Shahid, N.H.; Nazir, L.A.; Tanveer, M.A.; Divya, G.; Archoo, S.; Raghu, S.R.; Tasduq, S.A. Pharmacological Activation of Autophagy Restores Cellular Homeostasis in Ultraviolet-(B)-Induced Skin Photodamage. Front. Oncol. 2021, 11, 726066. [Google Scholar] [CrossRef]
- Mokrzyński, K.; Krzysztyńska-Kuleta, O.; Zawrotniak, M.; Sarna, M.; Sarna, T. Fine Particulate Matter-Induced Oxidative Stress Mediated by UVA-Visible Light Leads to Keratinocyte Damage. Int. J. Mol. Sci. 2021, 22, 10645. [Google Scholar] [CrossRef] [PubMed]
- Requena, M.B.; Russignoli, P.E.; Vollet-Filho, J.D.; Salvio, A.G.; Fortunato, T.C.; Pratavieira, S.; Bagnato, V.S. Use of dermograph for improvement of PpIX precursor’s delivery in photodynamic therapy: Experimental and clinical pilot studies. Photodiagnosis Photodyn. Ther. 2020, 29, 101599. [Google Scholar] [CrossRef]
- Pinto, M.A.F.; Ferreira, C.B.R.; de Lima, B.E.S.; Molon, Â.C.; Ibarra, A.M.C.; Cecatto, R.B.; Dos Santos Franco, A.L.; Rodrigues, M. Effects of 5-ALA mediated photodynamic therapy in oral cancer stem cells. J. Photochem. Photobiol. B 2022, 235, 112552. [Google Scholar] [CrossRef]
- Arakane, K.; Ryu, A.; Hayashi, C.; Masunaga, T.; Shinmoto, K.; Mashiko, S.; Nagano, T.; Hirobe, M. Singlet oxygen (1 delta g) generation from coproporphyrin in Propionibacterium acnes on irradiation. Biochem. Biophys. Res. Commun. 1996, 223, 578–582. [Google Scholar] [CrossRef]
- Patwardhan, S.V.; Richter, C.; Vogt, A.; Blume-Peytavi, U.; Canfield, D.; Kottner, J. Measuring acne using Coproporphyrin III, Protoporphyrin IX, and lesion-specific inflammation: An exploratory study. Arch. Dermatol. Res. 2017, 309, 159–167. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, X.; Wang, Z.; Sun, J.; He, Z.; Sun, B.; Luo, C. Molecularly Self-Engineered Nanoamplifier for Boosting Photodynamic Therapy via Cascade Oxygen Elevation and Lipid ROS Accumulation. ACS Appl. Mater Interfaces 2022, 14, 38497–38505. [Google Scholar] [CrossRef] [PubMed]
- Uliana, M.P.; da Cruz Rodrigues, A.; Ono, B.A.; Pratavieira, S.; de Oliveira, K.T.; Kurachi, C. Photodynamic Inactivation of Microorganisms Using Semisynthetic Chlorophyll a Derivatives as Photosensitizers. Molecules 2022, 27, 5769. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Azami, C.; Koyama, K.; Rutherford, A.W.; Rappaport, F.; Boussac, A. Modification of the pheophytin redox potential in Thermosynechococcus elongatus Photosystem II with PsbA3 as D1. Biochim. Biophys. Acta 2014, 1837, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.; Braun, M.S.; Wink, M. Chlorophyll and Chlorophyll Derivatives Interfere with Multi-Drug Resistant Cancer Cells and Bacteria. Molecules 2019, 24, 2968. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Poojary, M.M.; Skibsted, L.H.; Lund, M.N. Cleavage of Disulfide Bonds in Cystine by UV-B Illumination Mediated by Tryptophan or Tyrosine as Photosensitizers. J. Agric. Food Chem. 2020, 68, 6900–6909. [Google Scholar] [CrossRef]
- Ionita, G.; Matei, I. Application of Riboflavin Photochemical Properties in Hydrogel Synthesis. In Biophysical Chemistry Advance Applications; InTech Open: London, UK, 2020. [Google Scholar]
- Wangsuwan, S.; Meephansan, J. Comparative Study Of Photodynamic Therapy With Riboflavin-Tryptophan Gel And 13% 5-Aminolevulinic Acid In The Treatment Of Mild To Moderate Acne Vulgaris. Clin. Cosmet. Investig. Dermatol. 2019, 12, 805–814. [Google Scholar] [CrossRef]
- Chignell, C.F.; Kukielczak, B.M.; Sik, R.H.; Bilski, P.J.; He, Y.Y. Ultraviolet A sensitivity in Smith-Lemli-Opitz syndrome: Possible involvement of cholesta-5,7,9(11)-trien-3 beta-ol. Free Radic. Biol. Med. 2006, 41, 339–346. [Google Scholar] [CrossRef]
- Bode, C.W.; Hänsel, W. 5-(3-Phenylpropoxy)psoralen and 5-(4-phenylbutoxy)psoralen: Mechanistic studies on phototoxicity. Pharmazie 2005, 60, 225–228. [Google Scholar]
- Lee, K.P.; Girijala, R.L.; Chon, S.Y. Phytophotodermatitis due to a Citrus-Based Hand Sanitizer: A Case Report. Korean J. Fam. Med. 2022, 43, 271–273. [Google Scholar] [CrossRef]
- Pappas, A. Epidermal surface lipids. Dermatoendocrinology 2009, 1, 72–76. [Google Scholar] [CrossRef]
- Chapkin, R.S.; Ziboh, V.A.; Marcelo, C.L.; Voorhees, J.J. Metabolism of essential fatty acids by human epidermal enzyme preparations: Evidence of chain elongation. J. Lipid Res. 1986, 27, 945–954. [Google Scholar] [CrossRef]
- Ni Raghallaigh, S.; Bender, K.; Lacey, N.; Brennan, L.; Powell, F.C. The fatty acid profile of the skin surface lipid layer in papulopustular rosacea. Br. J. Dermatol. 2012, 166, 279–287. [Google Scholar]
- Shimizu, N.; Ito, J.; Kato, S.; Otoki, Y.; Goto, M.; Eitsuka, T.; Miyazawa, T.; Nakagawa, K. Oxidation of squalene by singlet oxygen and free radicals results in different compositions of squalene monohydroperoxide isomers. Sci. Rep. 2018, 8, 9116. [Google Scholar] [CrossRef] [PubMed]
- Chiba, K.; Kawakami, K.; Sone, T.; Onoue, M. Characteristics of skin wrinkling and dermal changes induced by repeated application of squalene monohydroperoxide to hairless mouse skin. Skin Pharmacol. Appl. Skin Physiol. 2003, 16, 242–251. [Google Scholar] [CrossRef]
- Chiba, K.; Yoshizawa, K.; Makino, I.; Kawakami, K.; Onoue, M. Comedogenicity of squalene monohydroperoxide in the skin after topical application. J. Toxicol. Sci. 2000, 25, 77–83. [Google Scholar] [CrossRef]
- Khalifa, S.; Enomoto, M.; Kato, S.; Nakagawa, K. Novel Photoinduced Squalene Cyclic Peroxide Identified, Detected, and Quantified in Human Skin Surface Lipids. Antioxidants 2021, 10, 1760. [Google Scholar] [CrossRef]
- Dalrymple, A.; McEwan, M.; Brandt, M.; Bielfeldt, S.; Bean, E.J.; Moga, A.; Coburn, S.; Hardie, G. A novel clinical method to measure skin staining reveals activation of skin damage pathways by cigarette smoke. Skin Res. Technol. 2022, 28, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Minami, Y.; Yokoyama, K.; Bando, N.; Kawai, Y.; Terao, J. Occurrence of singlet oxygen oxygenation of oleic acid and linoleic acid in the skin of live mice. Free Radic. Res. 2008, 42, 197–204. [Google Scholar] [CrossRef]
- Egawa, M.; Kohno, Y.; Kumano, Y. Oxidative effects of cigarette smoke on the human skin. Int. J. Cosmet. Sci. 1999, 21, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Ibusuki, D.; Suzuki, Y.; Yamashita, S.; Higuchi, O.; Oikawa, S.; Miyazawa, T. Ion-trap tandem mass spectrometric analysis of squalene monohydroperoxide isomers in sunlight-exposed human skin. J. Lipid Res. 2007, 48, 2779–2787. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake-Mudiyanselage, S.; Tavakkol, A.; Polefka, T.G.; Nabi, Z.; Elsner, P.; Thiele, J.J. Vitamin E delivery to human skin by a rinse-off product: Penetration of alpha-tocopherol versus wash-out effects of skin surface lipids. Skin Pharmacol. Physiol. 2005, 18, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Ekanayake Mudiyanselage, S.; Hamburger, M.; Elsner, P.; Thiele, J.J. Ultraviolet a induces generation of squalene monohydroperoxide isomers in human sebum and skin surface lipids in vitro and in vivo. J. Invest. Dermatol. 2003, 120, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Auffray, B. Protection against singlet oxygen, the main actor of sebum squalene peroxidation during sun exposure, using Commiphora myrrha essential oil. Int. J. Cosmet. Sci. 2007, 29, 23–29. [Google Scholar] [CrossRef]
- Lens, M.; Podesta Marty, M.H. Assessment of the kinetics of the antioxidative capacity of topical antioxidants. J. Drugs Dermatol. 2011, 10, 262–267. [Google Scholar] [PubMed]
- Shimizu, N.; Ito, J.; Kato, S.; Eitsuka, T.; Saito, T.; Nishida, H.; Miyazawa, T.; Nakagawa, K. Evaluation of squalene oxidation mechanisms in human skin surface lipids and shark liver oil supplements. Ann. N. Y. Acad. Sci. 2019, 1457, 158–165. [Google Scholar] [CrossRef]
- Jourdain, R.; Moga, A.; Vingler, P.; El Rawadi, C.; Pouradier, F.; Souverain, L.; Bastien, P.; Amalric, N.; Breton, L. Exploration of scalp surface lipids reveals squalene peroxide as a potential actor in dandruff condition. Arch. Dermatol. Res. 2016, 308, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Mountfort, K.A.; Bronstein, H.; Archer, N.; Jickells, S.M. Identification of oxidation products of squalene in solution and in latent fingerprints by ESI-MS and LC/APCI-MS. Anal. Chem. 2007, 79, 2650–2657. [Google Scholar] [CrossRef]
- Curpen, S.; Francois-Newton, V.; Moga, A.; Hosenally, M.; Petkar, G.; Soobramaney, V.; Ruchaia, B.; Lutchmanen Kolanthan, V.; Roheemun, N.; Sokeechand, B.N.; et al. A novel method for evaluating the effect of pollution on the human skin under controlled conditions. Skin Res. Technol. 2020, 26, 50–60. [Google Scholar] [CrossRef]
- Bielfeldt, S.; Jung, K.; Laing, S.; Moga, A.; Wilhelm, K.P. Anti-pollution effects of two antioxidants and a chelator-Ex vivo electron spin resonance and in vivo cigarette smoke model assessments in human skin. Skin Res. Technol. 2021, 27, 1092–1099. [Google Scholar] [CrossRef]
- Someya, K.; Totsuka, Y.; Murakoshi, M.; Kitano, H.; Miyazawa, T. The antioxidant effect of palm fruit carotene on skin lipid peroxidation in guinea pigs as estimated by chemiluminescence-HPLC method. J. Nutr. Sci. Vitaminol. 1994, 40, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Pena, A.M.; Baldeweck, T.; Decencière, E.; Koudoro, S.; Victorin, S.; Raynaud, E.; Ngo, B.; Bastien, P.; Brizion, S.; Tancrède-Bohin, E. In vivo multiphoton multiparametric 3D quantification of human skin aging on forearm and face. Sci. Rep. 2022, 12, 14863. [Google Scholar] [CrossRef] [PubMed]
- Scharffetter, K.; Wlaschek, M.; Hogg, A.; Bolsen, K.; Schothorst, A.; Goerz, G.; Krieg, T.; Plewig, G. UVA irradiation induces collagenase in human dermal fibroblasts in vitro and in vivo. Arch. Dermatol. Res. 1991, 283, 506–511. [Google Scholar] [CrossRef]
- Nakamura, T.; Noma, A.; Shimada, S.; Ishii, N.; Bando, N.; Kawai, Y.; Terao, J. Non-selective distribution of isomeric cholesterol hydroperoxides to microdomains in cell membranes and activation of matrix metalloproteinase activity in a model of dermal cells. Chem. Phys. Lipids 2013, 174, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to the guanine moiety of DNA: Mechanistic aspects and formation in cells. Acc. Chem. Res. 2008, 41, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Photochem. Photobiol. 2015, 91, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Ravanat, J.L.; Di Mascio, P.; Martinez, G.R.; Medeiros, M.H.; Cadet, J. Singlet oxygen induces oxidation of cellular DNA. J. Biol. Chem. 2000, 275, 40601–40604. [Google Scholar] [CrossRef] [PubMed]
- Yagura, T.; Schuch, A.P.; Garcia, C.C.M.; Rocha, C.R.R.; Moreno, N.C.; Angeli, J.P.F.; Mendes, D.; Severino, D.; Bianchini Sanchez, A.; Di Mascio, P.; et al. Direct participation of DNA in the formation of singlet oxygen and base damage under UVA irradiation. Free Radic. Biol. Med. 2017, 108, 86–93. [Google Scholar] [CrossRef]
- Cadet, J.; Sage, E.; Douki, T. Ultraviolet radiation-mediated damage to cellular DNA. Mutat. Res. 2005, 571, 3–17. [Google Scholar] [CrossRef]
- Mouret, S.; Baudouin, C.; Charveron, M.; Favier, A.; Cadet, J.; Douki, T. Cyclobutane pyrimidine dimers are predominant DNA lesions in whole human skin exposed to UVA radiation. Proc. Natl. Acad. Sci. USA 2006, 103, 13765–13770. [Google Scholar] [CrossRef]
- Prat, F.; Hou, C.-C.; Foote, C.S. Determination of the Quenching Rate Constants of Singlet Oxygen by Derivatized Nucleosides in Nonaqueous Solution. J. Am. Chem. Soc. 1997, 119, 5051–5052. [Google Scholar] [CrossRef]
- David, S.S.; O’Shea, V.L.; Kundu, S. Base-excision repair of oxidative DNA damage. Nature 2007, 447, 941–950. [Google Scholar] [CrossRef] [PubMed]
- Hsu, G.W.; Ober, M.; Carell, T.; Beese, L.S. Error-prone replication of oxidatively damaged DNA by a high-fidelity DNA polymerase. Nature 2004, 431, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Greenman, C.; Stephens, P.; Smith, R.; Dalgliesh, G.L.; Hunter, C.; Bignell, G.; Davies, H.; Teague, J.; Butler, A.; Stevens, C.; et al. Patterns of somatic mutation in human cancer genomes. Nature 2007, 446, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Barnes, D.E. Repair of endogenous DNA damage. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 127–133. [Google Scholar] [CrossRef]
- Greinert, R.; Volkmer, B.; Henning, S.; Breitbart, E.W.; Greulich, K.O.; Cardoso, M.C.; Rapp, A. UVA-induced DNA double-strand breaks result from the repair of clustered oxidative DNA damages. Nucleic Acids Res. 2012, 40, 10263–10273. [Google Scholar] [CrossRef] [PubMed]
- Agar, N.S.; Halliday, G.M.; Barnetson, R.S.; Ananthaswamy, H.N.; Wheeler, M.; Jones, A.M. The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: A role for UVA in human skin carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4954–4959. [Google Scholar] [CrossRef]
- Kunisada, M.; Sakumi, K.; Tominaga, Y.; Budiyanto, A.; Ueda, M.; Ichihashi, M.; Nakabeppu, Y.; Nishigori, C. 8-Oxoguanine formation induced by chronic UVB exposure makes Ogg1 knockout mice susceptible to skin carcinogenesis. Cancer Res. 2005, 65, 6006–6010. [Google Scholar] [CrossRef]
- Kunisada, M.; Kumimoto, H.; Ishizaki, K.; Sakumi, K.; Nakabeppu, Y.; Nishigori, C. Narrow-band UVB induces more carcinogenic skin tumors than broad-band UVB through the formation of cyclobutane pyrimidine dimer. J. Invest. Dermatol. 2007, 127, 2865–2871. [Google Scholar] [CrossRef]
- Zhang, R.B.; Eriksson, L.A. A triplet mechanism for the formation of cyclobutane pyrimidine dimers in UV-irradiated DNA. J. Phys. Chem. B 2006, 110, 7556–7562. [Google Scholar] [CrossRef]
- You, Y.; Zhu, F.; Li, Z.; Zhang, L.; Xie, Y.; Chinnathambi, A.; Alahmadi, T.A.; Lu, B. Phyllanthin prevents diethylnitrosamine (DEN) induced liver carcinogenesis in rats and induces apoptotic cell death in HepG2 cells. Biomed. Pharmacother. 2021, 137, 111335. [Google Scholar] [CrossRef]
- Yuan, H.; Guo, L.; Su, Q.; Su, X.; Wen, Y.; Wang, T.; Yang, P.; Xu, M.; Li, F. Afterglow Amplification for Fast and Sensitive Detection of Porphyria in Whole Blood. ACS Appl. Mater. Interfaces 2021, 13, 27991–27998. [Google Scholar] [CrossRef]
- Ventura, P.; Brancaleoni, V.; Di Pierro, E.; Graziadei, G.; Macrì, A.; Carmine Guida, C.; Nicolli, A.; Rossi, M.T.; Granata, F.; Fiorentino, V.; et al. Clinical and molecular epidemiology of erythropoietic protoporphyria in Italy. Eur. J. Dermatol. 2020, 30, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M. Erythropoietic Protoporphyria and X-Linked Protoporphyria: Pathophysiology, genetics, clinical manifestations, and management. Mol. Genet. Metab. 2019, 128, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Mathews-Roth, M.M.; Pathak, M.A.; Fitzpatrick, T.B.; Harber, L.C.; Kass, E.H. Beta-carotene as a photoprotective agent in erythropoietic protoporphyria. N. Engl. J. Med. 1970, 282, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids that Exhibit both Anti- and Pro-Oxidative Mechanisms-A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef]
- Mathews-Roth, M.M.; Pathak, M.A.; Fitzpatrick, T.B.; Harber, L.H.; Kass, E.H. Beta carotene therapy for erythropoietic protoporphyria and other photosensitivity diseases. Arch. Dermatol. 1977, 113, 1229–1232. [Google Scholar] [CrossRef]
- Maitra, D.; Bragazzi Cunha, J.; Elenbaas, J.S.; Bonkovsky, H.L.; Shavit, J.A.; Omary, M.B. Porphyrin-Induced Protein Oxidation and Aggregation as a Mechanism of Porphyria-Associated Cell Injury. Cell Mol Gastroenterol. Hepatol. 2019, 8, 535–548. [Google Scholar] [CrossRef]
- Maitra, D.; Carter, E.L.; Richardson, R.; Rittié, L.; Basrur, V.; Zhang, H.; Nesvizhskii, A.I.; Osawa, Y.; Wolf, M.W.; Ragsdale, S.W.; et al. Oxygen and Conformation Dependent Protein Oxidation and Aggregation by Porphyrins in Hepatocytes and Light-Exposed Cells. Cell Mol. Gastroenterol. Hepatol. 2019, 8, 659–682.e651. [Google Scholar] [CrossRef]
- Maitra, D.; Elenbaas, J.S.; Whitesall, S.E.; Basrur, V.; D’Alecy, L.G.; Omary, M.B. Ambient Light Promotes Selective Subcellular Proteotoxicity after Endogenous and Exogenous Porphyrinogenic Stress. J. Biol. Chem. 2015, 290, 23711–23724. [Google Scholar] [CrossRef]
- Tomita, H.; Hines, K.M.; Herron, J.M.; Li, A.; Baggett, D.W.; Xu, L. 7-Dehydrocholesterol-derived oxysterols cause neurogenic defects in Smith-Lemli-Opitz syndrome. Elife 2022, 11, e67141. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, C.; Ma, L.; Zhang, Y.; Liu, Z.; Chen, L.; Wang, R.; Luan, Y.; Rao, Y. Measurement of 7-dehydrocholesterol and cholesterol in hair can be used in the diagnosis of Smith-Lemli-Opitz syndrome. J. Lipid Res. 2022, 63, 100228. [Google Scholar] [CrossRef]
- Koczok, K.; Horváth, L.; Korade, Z.; Mezei, Z.A.; Szabó, G.P.; Porter, N.A.; Kovács, E.; Mirnics, K.; Balogh, I. Biochemical and Clinical Effects of Vitamin E Supplementation in Hungarian Smith-Lemli-Opitz Syndrome Patients. Biomolecules 2021, 11, 1228. [Google Scholar] [CrossRef] [PubMed]
- Quiec, D.; Mazière, C.; Auclair, M.; Santus, R.; Gardette, J.; Redziniak, G.; Franchi, J.; Dubertret, L.; Mazière, J.C. Lovastatin enhances the photocytotoxicity of UVA radiation towards cultured N.C.T.C. 2544 human keratinocytes: Prevention by cholesterol supplementation and by a cathepsin inhibitor. Biochem. J. 1995, 310 Pt 1, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Alrashidi, A.; Rhodes, L.E.; Sharif, J.C.H.; Kreeshan, F.C.; Farrar, M.D.; Ahad, T. Systemic drug photosensitivity-Culprits, impact and investigation in 122 patients. Photodermatol. Photoimmunol. Photomed. 2020, 36, 441–451. [Google Scholar] [CrossRef]
- Chen, H.L.; Chang, H.M.; Wu, H.J.; Lin, Y.C.; Chang, Y.H.; Chang, Y.C.; Lee, W.H.; Chang, C.T. Effect of hydrophilic and lipophilic statins on early onset cataract: A nationwide case-control study. Regul. Toxicol. Pharmacol. 2021, 124, 104970. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, B.W.; Lin, L.N.; Zan, X.L.; Zhang, G.C.; Chen, G.S.; Ji, J.Y.; Ma, W.H. Key factors affecting photoactivated fungicidal activity of sodium pheophorbide a against Pestalotiopsis neglecta. Pestic. Biochem. Physiol. 2020, 167, 104584. [Google Scholar] [CrossRef]
- Kobayashi, T.; Sugaya, K.; Onose, J.I.; Abe, N. Peppermint (Mentha piperita L.) extract effectively inhibits cytochrome P450 3A4 (CYP3A4) mRNA induction in rifampicin-treated HepG2 cells. Biosci. Biotechnol. Biochem. 2019, 83, 1181–1192. [Google Scholar] [CrossRef]
- Puschner, B.; Chen, X.; Read, D.; Affolter, V.K. Alfalfa hay induced primary photosensitization in horses. Vet. J. 2016, 211, 32–38. [Google Scholar] [CrossRef]
- Bayoumy, A.B.; Crouwel, F.; Chanda, N.; Florin, T.H.J.; Buiter, H.J.C.; Mulder, C.J.J.; de Boer, N.K.H. Advances in Thiopurine Drug Delivery: The Current State-of-the-Art. Eur. J. Drug Metab. Pharmacokinet. 2021, 46, 743–758. [Google Scholar] [CrossRef]
- Euvrard, S.; Kanitakis, J.; Claudy, A. Skin cancers after organ transplantation. N. Engl. J. Med. 2003, 348, 1681–1691. [Google Scholar] [CrossRef]
- Bignon, E.; Marazzi, M.; Besancenot, V.; Gattuso, H.; Drouot, G.; Morell, C.; Eriksson, L.A.; Grandemange, S.; Dumont, E.; Monari, A. Ibuprofen and ketoprofen potentiate UVA-induced cell death by a photosensitization process. Sci. Rep. 2017, 7, 8885. [Google Scholar] [CrossRef] [PubMed]
- Zawadzka, M.; Ràcz, B.; Ambrosini, D.; Görbitz, C.H.; Morth, J.P.; Wilkins, A.L.; Østeby, A.; Elgstøen, K.B.P.; Lundanes, E.; Rise, F.; et al. Searching for a UV-filter in the eyes of high-flying birds. Sci. Rep. 2021, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.L.; Lim, H.W.; Mohammad, T.F. Sunscreens and Photoaging: A Review of Current Literature. Am. J. Clin. Dermatol. 2021, 22, 819–828. [Google Scholar] [CrossRef]
- Behar-Cohen, F.; Martinsons, C.; Viénot, F.; Zissis, G.; Barlier-Salsi, A.; Cesarini, J.P.; Enouf, O.; Garcia, M.; Picaud, S.; Attia, D. Light-emitting diodes (LED) for domestic lighting: Any risks for the eye? Prog. Retin. Eye Res. 2011, 30, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.X.; Zhou, W.C.; Zhu, X.G. Mitochondria as Potential Targets and Initiators of the Blue Light Hazard to the Retina. Oxid. Med. Cell. Longev. 2019, 2019, 6435364. [Google Scholar] [CrossRef] [PubMed]
- Kaarniranta, K.; Blasiak, J.; Liton, P.; Boulton, M.; Klionsky, D.J.; Sinha, D. Autophagy in age-related macular degeneration. Autophagy 2023, 19, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Różanowska, M.B.; Pawlak, A.; Różanowski, B. Products of Docosahexaenoate Oxidation as Contributors to Photosensitising Properties of Retinal Lipofuscin. Int. J. Mol. Sci. 2021, 22, 3525. [Google Scholar] [CrossRef]
- Höhn, A.; Grune, T. Lipofuscin: Formation, effects and role of macroautophagy. Redox Biol. 2013, 1, 140–144. [Google Scholar] [CrossRef]
- Singh Kushwaha, S.; Patro, N.; Kumar Patro, I. A Sequential Study of Age-Related Lipofuscin Accumulation in Hippocampus and Striate Cortex of Rats. Ann. Neurosci. 2018, 25, 223–233. [Google Scholar] [CrossRef]
- Gao, Z.; Liao, Y.; Chen, C.; Liao, C.; He, D.; Chen, J.; Ma, J.; Liu, Z.; Wu, Y. Conversion of all-trans-retinal into all-trans-retinal dimer reflects an alternative metabolic/antidotal pathway of all-trans-retinal in the retina. J. Biol. Chem. 2018, 293, 14507–14519. [Google Scholar] [CrossRef]
- Parish, C.A.; Hashimoto, M.; Nakanishi, K.; Dillon, J.; Sparrow, J. Isolation and one-step preparation of A2E and iso-A2E, fluorophores from human retinal pigment epithelium. Proc. Natl. Acad. Sci. USA 1998, 95, 14609–14613. [Google Scholar] [CrossRef] [PubMed]
- Marie, M.; Bigot, K.; Angebault, C.; Barrau, C.; Gondouin, P.; Pagan, D.; Fouquet, S.; Villette, T.; Sahel, J.A.; Lenaers, G.; et al. Light action spectrum on oxidative stress and mitochondrial damage in A2E-loaded retinal pigment epithelium cells. Cell Death Dis. 2018, 9, 287. [Google Scholar] [CrossRef] [PubMed]
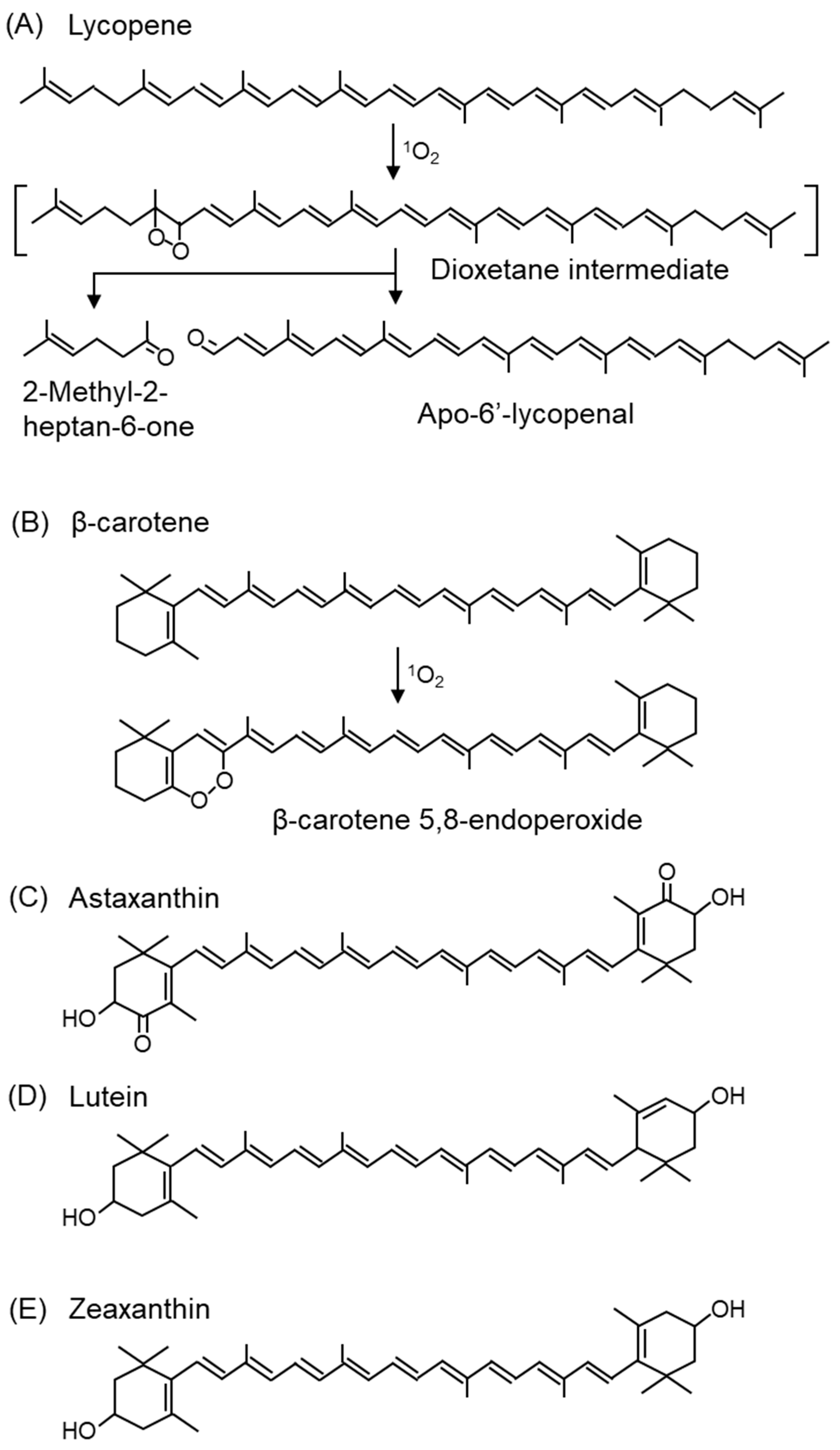
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [PubMed]
- Lem, D.W.; Davey, P.G.; Gierhart, D.L.; Rosen, R.B. A Systematic Review of Carotenoids in the Management of Age-Related Macular Degeneration. Antioxidants 2021, 10, 1255. [Google Scholar] [CrossRef] [PubMed]
- Shyam, R.; Gorusupudi, A.; Nelson, K.; Horvath, M.P.; Bernstein, P.S. RPE65 has an additional function as the lutein to meso-zeaxanthin isomerase in the vertebrate eye. Proc. Natl. Acad. Sci. USA 2017, 114, 10882–10887. [Google Scholar] [CrossRef] [PubMed]
- Johra, F.T.; Bepari, A.K.; Bristy, A.T.; Reza, H.M. A Mechanistic Review of β-Carotene, Lutein, and Zeaxanthin in Eye Health and Disease. Antioxidants 2020, 9, 1046. [Google Scholar] [CrossRef] [PubMed]
- Mizdrak, J.; Hains, P.G.; Truscott, R.J.; Jamie, J.F.; Davies, M.J. Tryptophan-derived ultraviolet filter compounds covalently bound to lens proteins are photosensitizers of oxidative damage. Free Radic. Biol. Med. 2008, 44, 1108–1119. [Google Scholar] [CrossRef]
- Ouyang, X.; Yang, J.; Hong, Z.; Wu, Y.; Xie, Y.; Wang, G. Mechanisms of blue light-induced eye hazard and protective measures: A review. Biomed. Pharmacother. 2020, 130, 110577. [Google Scholar] [CrossRef]
- Boulton, M.; Rozanowska, M.; Rozanowski, B.; Wess, T. The photoreactivity of ocular lipofuscin. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2004, 3, 759–764. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Khaw, P.T. Primary open-angle glaucoma. Lancet 2004, 363, 1711–1720. [Google Scholar] [CrossRef]
- Foster, A.; Resnikoff, S. The impact of Vision 2020 on global blindness. Eye 2005, 19, 1133–1135. [Google Scholar] [CrossRef] [PubMed]
- Tanito, M.; Kaidzu, S.; Takai, Y.; Ohira, A. Status of systemic oxidative stresses in patients with primary open-angle glaucoma and pseudoexfoliation syndrome. PLoS ONE 2012, 7, e49680. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, J.A.; Murphy, C.G. Outflow obstruction in pigmentary and primary open angle glaucoma. Arch. Ophthalmol. 1992, 110, 1769–1778. [Google Scholar] [CrossRef] [PubMed]
- Himori, N.; Yamamoto, K.; Maruyama, K.; Ryu, M.; Taguchi, K.; Yamamoto, M.; Nakazawa, T. Critical role of Nrf2 in oxidative stress-induced retinal ganglion cell death. J. Neurochem. 2013, 127, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Maruyama, K.; Yamamoto, K.; Omodaka, K.; Yasuda, M.; Himori, N.; Ryu, M.; Nishiguchi, K.M.; Nakazawa, T. The role of calpain in an in vivo model of oxidative stress-induced retinal ganglion cell damage. Biochem. Biophys. Res. Commun. 2014, 451, 510–515. [Google Scholar] [CrossRef]
- Inomata, Y.; Nakamura, H.; Tanito, M.; Teratani, A.; Kawaji, T.; Kondo, N.; Yodoi, J.; Tanihara, H. Thioredoxin inhibits NMDA-induced neurotoxicity in the rat retina. J. Neurochem. 2006, 98, 372–385. [Google Scholar] [CrossRef]
- Munemasa, Y.; Ahn, J.H.; Kwong, J.M.; Caprioli, J.; Piri, N. Redox proteins thioredoxin 1 and thioredoxin 2 support retinal ganglion cell survival in experimental glaucoma. Gene Ther. 2009, 16, 17–25. [Google Scholar] [CrossRef]
- Umeno, A.; Tanito, M.; Kaidzu, S.; Takai, Y.; Yoshida, Y. Involvement of free radical-mediated oxidation in the pathogenesis of pseudoexfoliation syndrome detected based on specific hydroxylinoleate isomers. Free Radic. Biol. Med. 2020, 147, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Umeno, A.; Yoshida, Y.; Kaidzu, S.; Tanito, M. Positive Association between Aqueous Humor Hydroxylinoleate Levels and Intraocular Pressure. Molecules 2022, 27, 2215. [Google Scholar] [CrossRef]
- Schafheimer, N.; King, J. Tryptophan cluster protects human γD-crystallin from ultraviolet radiation-induced photoaggregation in vitro. Photochem. Photobiol. 2013, 89, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Hayakawa, M.; Habuchi, Y.; Niki, E. Evaluation of the dietary effects of coenzyme Q in vivo by the oxidative stress marker, hydroxyoctadecadienoic acid and its stereoisomer ratio. Biochim. Biophys. Acta 2006, 1760, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Hirashima, Y.; Doshi, M.; Hayashi, N.; Endo, S.; Akazawa, Y.; Shichiri, M.; Yoshida, Y. Plasma platelet-activating factor-acetyl hydrolase activity and the levels of free forms of biomarker of lipid peroxidation in cerebrospinal fluid of patients with aneurysmal subarachnoid hemorrhage. Neurosurgery 2012, 70, 602–609. [Google Scholar] [CrossRef]
- Fan Gaskin, J.C.; Shah, M.H.; Chan, E.C. Oxidative Stress and the Role of NADPH Oxidase in Glaucoma. Antioxidants 2021, 10, 238. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Medina, J.J.; Rubio-Velazquez, E.; Lopez-Bernal, M.D.; Cobo-Martinez, A.; Zanon-Moreno, V.; Pinazo-Duran, M.D.; Del-Rio-Vellosillo, M. Glaucoma and Antioxidants: Review and Update. Antioxidants 2020, 9, 1031. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, S.; Cao, W.; Sun, X. The Association of Oxidative Stress Status with Open-Angle Glaucoma and Exfoliation Glaucoma: A Systematic Review and Meta-Analysis. J. Ophthalmol. 2019, 2019, 1803619. [Google Scholar] [CrossRef]
- Neale, R.E.; Purdie, J.L.; Hirst, L.W.; Green, A.C. Sun exposure as a risk factor for nuclear cataract. Epidemiology 2003, 14, 707–712. [Google Scholar] [CrossRef]
- Garźon-Chavez, D.R.; Quentin, E.; Harrison, S.L.; Parisi, A.V.; Butler, H.J.; Downs, N.J. The geospatial relationship of pterygium and senile cataract with ambient solar ultraviolet in tropical Ecuador. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2018, 17, 1075–1083. [Google Scholar] [CrossRef]
- Dillon, J.; Atherton, S.J. Time resolved spectroscopic studies on the intact human lens. Photochem. Photobiol. 1990, 51, 465–468. [Google Scholar] [CrossRef]
- Kessel, L.; Eskildsen, L.; Lundeman, J.H.; Jensen, O.B.; Larsen, M. Optical effects of exposing intact human lenses to ultraviolet radiation and visible light. BMC Ophthalmol. 2011, 11, 41. [Google Scholar] [CrossRef]
- Davies, M.J.; Truscott, R.J. Photo-oxidation of proteins and its role in cataractogenesis. J. Photochem. Photobiol. B 2001, 63, 114–125. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Zhao, B.; Andley, U.P.; Mason, R.P.; Roberts, J.E. Immunological detection of N-formylkynurenine in porphyrin-mediated photooxided lens α-crystallin. Photochem. Photobiol. 2011, 87, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Benedek, G.B. Theory of transparency of the eye. Appl. Opt. 1971, 10, 459–473. [Google Scholar] [CrossRef] [PubMed]
- Ávila, F.; Ravello, N.; Zanocco, A.L.; Gamon, L.F.; Davies, M.J.; Silva, E. 3-Hydroxykynurenine bound to eye lens proteins induces oxidative modifications in crystalline proteins through a type I photosensitizing mechanism. Free Radic. Biol. Med. 2019, 141, 103–114. [Google Scholar] [CrossRef]
- Parker, N.R.; Jamie, J.F.; Davies, M.J.; Truscott, R.J. Protein-bound kynurenine is a photosensitizer of oxidative damage. Free Radic. Biol. Med. 2004, 37, 1479–1489. [Google Scholar] [CrossRef]
- Ratter-Rieck, J.M.; Maalmi, H.; Trenkamp, S.; Zaharia, O.P.; Rathmann, W.; Schloot, N.C.; Straßburger, K.; Szendroedi, J.; Herder, C.; Roden, M. Leukocyte Counts and T-Cell Frequencies Differ Between Novel Subgroups of Diabetes and Are Associated With Metabolic Parameters and Biomarkers of Inflammation. Diabetes 2021, 70, 2652–2662. [Google Scholar] [CrossRef] [PubMed]
- Penno, G.; Solini, A.; Orsi, E.; Bonora, E.; Fondelli, C.; Trevisan, R.; Vedovato, M.; Cavalot, F.; Zerbini, G.; Lamacchia, O.; et al. Insulin resistance, diabetic kidney disease, and all-cause mortality in individuals with type 2 diabetes: A prospective cohort study. BMC Med. 2021, 19, 66. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xu, Y.; Liu, X.; Nie, Y.; Zhao, Z. Urinary heme oxygenase-1 as a potential biomarker for early diabetic nephropathy. Nephrology 2017, 22, 58–64. [Google Scholar] [CrossRef]
- Xu, G.W.; Yao, Q.H.; Weng, Q.F.; Su, B.L.; Zhang, X.; Xiong, J.H. Study of urinary 8-hydroxydeoxyguanosine as a biomarker of oxidative DNA damage in diabetic nephropathy patients. J. Pharm. Biomed. Anal. 2004, 36, 101–104. [Google Scholar] [CrossRef]
- Maschirow, L.; Khalaf, K.; Al-Aubaidy, H.A.; Jelinek, H.F. Inflammation, coagulation, endothelial dysfunction and oxidative stress in prediabetes--Biomarkers as a possible tool for early disease detection for rural screening. Clin. Biochem. 2015, 48, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Zhou, Y.H.; Wu, C.; Shao, Y.; Lu, C.L.; Wang, Q.Y. The changes in miR-130b levels in human serum and the correlation with the severity of diabetic nephropathy. Diabetes Metab. Res. Rev. 2015, 31, 717–724. [Google Scholar] [CrossRef]
- Sánchez-Gómez, F.J.; Espinosa-Díez, C.; Dubey, M.; Dikshit, M.; Lamas, S. S-glutathionylation: Relevance in diabetes and potential role as a biomarker. Biol. Chem. 2013, 394, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.; Wang, J.; Xie, C.; Yang, Y.; Tian, J.W.; Xue, Y.M.; Hou, F.F. Increased plasma advanced oxidation protein products is an early marker of endothelial dysfunction in type 2 diabetes patients without albuminuria 2. J. Diabetes 2014, 6, 417–426. [Google Scholar] [CrossRef]
- Ceriello, A.; Testa, R.; Genovese, S. Clinical implications of oxidative stress and potential role of natural antioxidants in diabetic vascular complications. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Paneni, F.; Costantino, S.; Cosentino, F. Molecular mechanisms of vascular dysfunction and cardiovascular biomarkers in type 2 diabetes. Cardiovasc. Diagn. Ther. 2014, 4, 324–332. [Google Scholar] [PubMed]
- Molnar, J.; Garamvolgyi, Z.; Herold, M.; Adanyi, N.; Somogyi, A.; Rigo, J., Jr. Serum selenium concentrations correlate significantly with inflammatory biomarker high-sensitive CRP levels in Hungarian gestational diabetic and healthy pregnant women at mid-pregnancy. Biol. Trace Elem. Res. 2008, 121, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Yu, J.Y.; Jenkins, A.J.; Nankervis, A.J.; Hanssen, K.F.; Henriksen, T.; Lorentzen, B.; Garg, S.K.; Menard, M.K.; Hammad, S.M.; et al. Trace elements as predictors of preeclampsia in type 1 diabetic pregnancy. Nutr. Res. 2015, 35, 421–430. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Spinelli, S.; Vitale, G.; Trichilo, V.; Loddo, S.; Marino, A. High Glucose Concentrations Affect Band 3 Protein in Human Erythrocytes. Antioxidants 2020, 9, 365. [Google Scholar] [CrossRef]
- Chai, J.F.; Kao, S.L.; Wang, C.; Lim, V.J.; Khor, I.W.; Dou, J.; Podgornaia, A.I.; Chothani, S.; Cheng, C.Y.; Sabanayagam, C.; et al. Genome-Wide Association for HbA1c in Malay Identified Deletion on SLC4A1 that Influences HbA1c Independent of Glycemia. J. Clin. Endocrinol. Metab. 2020, 105, dgaa658. [Google Scholar] [CrossRef]
- Zhekova, H.R.; Jiang, J.; Wang, W.; Tsirulnikov, K.; Kayık, G.; Khan, H.M.; Azimov, R.; Abuladze, N.; Kao, L.; Newman, D.; et al. CryoEM structures of anion exchanger 1 capture multiple states of inward- and outward-facing conformations. Commun. Biol. 2022, 5, 1372. [Google Scholar] [CrossRef]
- Kostopoulou, E.; Kalaitzopoulou, E.; Papadea, P.; Skipitari, M.; Rojas Gil, A.P.; Spiliotis, B.E.; Georgiou, C.D. Oxidized lipid-associated protein damage in children and adolescents with type 1 diabetes mellitus: New diagnostic/prognostic clinical markers. Pediatr. Diabetes 2021, 22, 1135–1142. [Google Scholar] [CrossRef]
- Veitch, S.; Njock, M.S.; Chandy, M.; Siraj, M.A.; Chi, L.; Mak, H.; Yu, K.; Rathnakumar, K.; Perez-Romero, C.A.; Chen, Z.; et al. MiR-30 promotes fatty acid beta-oxidation and endothelial cell dysfunction and is a circulating biomarker of coronary microvascular dysfunction in pre-clinical models of diabetes. Cardiovasc. Diabetol. 2022, 21, 31. [Google Scholar] [CrossRef] [PubMed]
- Griesser, E.; Vemula, V.; Raulien, N.; Wagner, U.; Reeg, S.; Grune, T.; Fedorova, M. Cross-talk between lipid and protein carbonylation in a dynamic cardiomyocyte model of mild nitroxidative stress. Redox Biol 2017, 11, 438–455. [Google Scholar] [CrossRef]
- Schöttker, B.; Xuan, Y.; Gào, X.; Anusruti, A.; Brenner, H. Oxidatively Damaged DNA/RNA and 8-Isoprostane Levels Are Associated with the Development of Type 2 Diabetes at Older Age: Results From a Large Cohort Study. Diabetes Care 2020, 43, 130–136. [Google Scholar] [CrossRef]
- Leinisch, F.; Mariotti, M.; Hägglund, P.; Davies, M.J. Structural and functional changes in RNAse A originating from tyrosine and histidine cross-linking and oxidation induced by singlet oxygen and peroxyl radicals. Free Radic. Biol. Med. 2018, 126, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Martinez, G.R.; Medeiros, M.H.; Di Mascio, P. Singlet molecular oxygen generated by biological hydroperoxides. J. Photochem. Photobiol. B 2014, 139, 24–33. [Google Scholar] [CrossRef]
- Umeno, A.; Fukui, T.; Hashimoto, Y.; Kataoka, M.; Hagihara, Y.; Nagai, H.; Horie, M.; Shichiri, M.; Yoshino, K.; Yoshida, Y. Early diagnosis of type 2 diabetes based on multiple biomarkers and non-invasive indices. J. Clin. Biochem. Nutr. 2018, 62, 187–194. [Google Scholar] [CrossRef]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Inside the neutrophil phagosome: Oxidants, myeloperoxidase, and bacterial killing. Blood 1998, 92, 3007–3017. [Google Scholar] [CrossRef]
- Badwey, J.A.; Karnovsky, M.L. Active oxygen species and the functions of phagocytic leukocytes. Annu. Rev. Biochem. 1980, 49, 695–726. [Google Scholar] [CrossRef] [PubMed]
- Kanofsky, J.R. Singlet oxygen production from the peroxidase-catalyzed oxidation of indole-3-acetic acid. J. Biol. Chem. 1988, 263, 14171–14175. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Martinez, G.R.; Rettori, D.; Augusto, O.; Medeiros, M.H.; Di Mascio, P. Linoleic acid hydroperoxide reacts with hypochlorous acid, generating peroxyl radical intermediates and singlet molecular oxygen. Proc. Natl. Acad. Sci. USA 2006, 103, 293–298. [Google Scholar] [CrossRef]
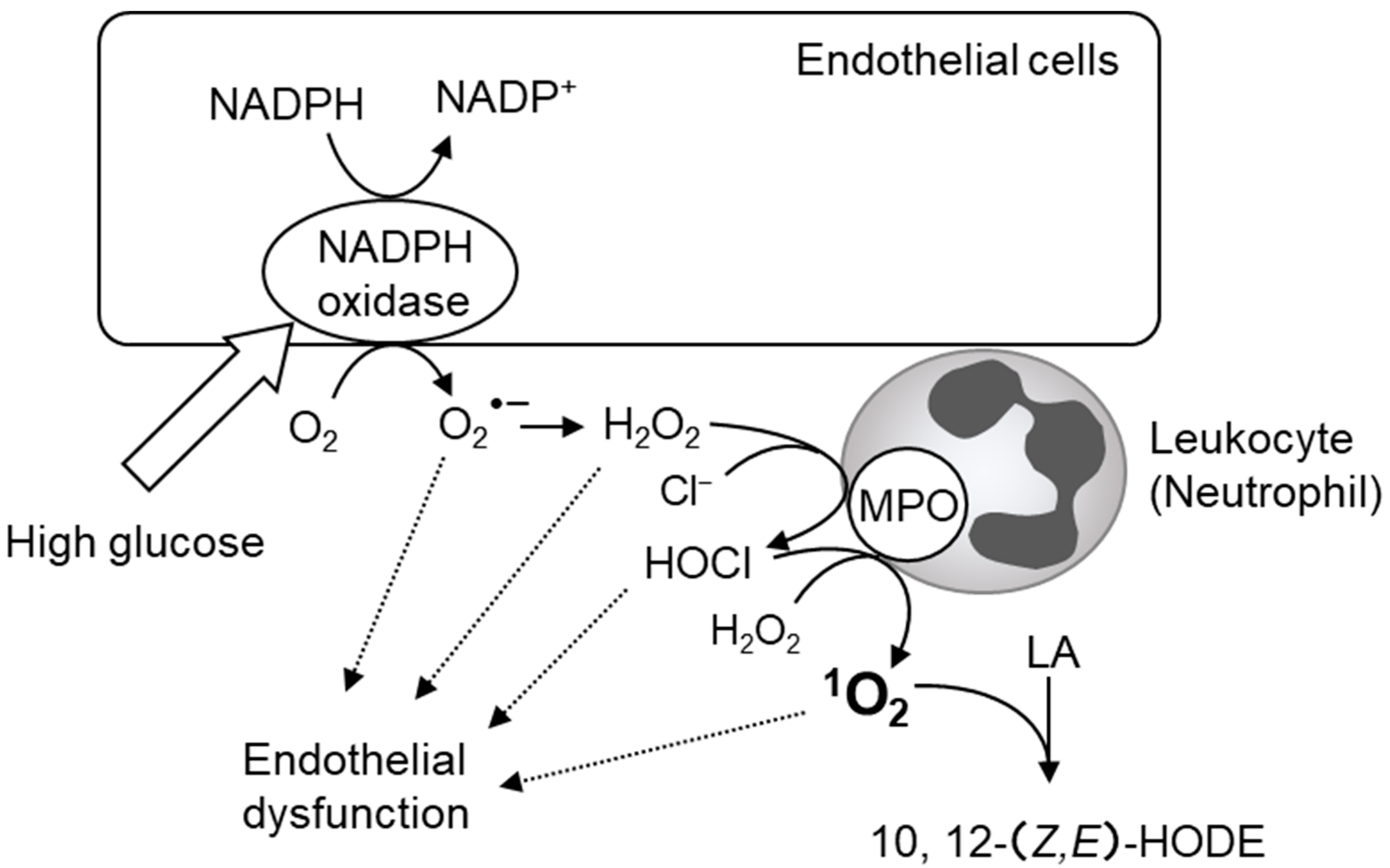
- Tian, R.; Ding, Y.; Peng, Y.Y.; Lu, N. Myeloperoxidase amplified high glucose-induced endothelial dysfunction in vasculature: Role of NADPH oxidase and hypochlorous acid. Biochem. Biophys. Res. Commun. 2017, 484, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, J.; Jennings, L.K. Leukocyte-derived myeloperoxidase amplifies high-glucose--induced endothelial dysfunction through interaction with high-glucose—Stimulated, vascular non—Leukocyte-derived reactive oxygen species. Diabetes 2004, 53, 2950–2959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, J.; Jacobs, J.D.; Jennings, L.K. Interaction of myeloperoxidase with vascular NAD(P)H oxidase-derived reactive oxygen species in vasculature: Implications for vascular diseases. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H2563–H2572. [Google Scholar] [CrossRef] [PubMed]
- Eiserich, J.P.; Baldus, S.; Brennan, M.L.; Ma, W.; Zhang, C.; Tousson, A.; Castro, L.; Lusis, A.J.; Nauseef, W.M.; White, C.R.; et al. Myeloperoxidase, a leukocyte-derived vascular NO oxidase. Science 2002, 296, 2391–2394. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, C.; Klinke, A.; Adam, M.; Baldus, S.; Sperandio, M. Myeloperoxidase: A leukocyte-derived protagonist of inflammation and cardiovascular disease. Antioxid. Redox Signal 2013, 18, 692–713. [Google Scholar] [CrossRef]
- Tian, R.; Jin, Z.; Zhou, L.; Zeng, X.P.; Lu, N. Quercetin Attenuated Myeloperoxidase-Dependent HOCl Generation and Endothelial Dysfunction in Diabetic Vasculature. J. Agric. Food Chem. 2021, 69, 404–413. [Google Scholar] [CrossRef]
- Onyango, A.N. The Contribution of Singlet Oxygen to Insulin Resistance. Oxid. Med. Cell. Longev. 2017, 2017, 8765972. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tamaoki, J.; Nagase, H.; Yamaguchi, M.; Horiguchi, T.; Hozawa, S.; Ichinose, M.; Iwanaga, T.; Kondo, R.; Nagata, M.; et al. Japanese guidelines for adult asthma 2020. Allergol. Int. 2020, 69, 519–548. [Google Scholar] [CrossRef]
- Kaur, R.; Chupp, G. Phenotypes and endotypes of adult asthma: Moving toward precision medicine. J. Allergy Clin. Immunol. 2019, 144, 1–12. [Google Scholar] [CrossRef]
- Hekking, P.W.; Wener, R.R.; Amelink, M.; Zwinderman, A.H.; Bouvy, M.L.; Bel, E.H. The prevalence of severe refractory asthma. J. Allergy Clin. Immunol. 2015, 135, 896–902. [Google Scholar] [CrossRef]
- Hammad, H.; Lambrecht, B.N. The basic immunology of asthma. Cell 2021, 184, 1469–1485. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Trevor, J.L.; Deshane, J.S. Refractory asthma: Mechanisms, targets, and therapy. Allergy 2014, 69, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, P.G.; Khashayar, R.; Lazarus, S.C.; Janson, S.; Avila, P.; Boushey, H.A.; Segal, M.; Fahy, J.V. Relationship between airway inflammation, hyperresponsiveness, and obstruction in asthma. J. Allergy Clin. Immunol. 2001, 108, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.E.; Berry, M.A.; Hargadon, B.; McKenna, S.; Shelley, M.J.; Green, R.H.; Brightling, C.E.; Wardlaw, A.J.; Pavord, I.D. Association between neutrophilic airway inflammation and airflow limitation in adults with asthma. Chest 2007, 132, 1871–1875. [Google Scholar] [CrossRef]
- Moore, W.C.; Hastie, A.T.; Li, X.; Li, H.; Busse, W.W.; Jarjour, N.N.; Wenzel, S.E.; Peters, S.P.; Meyers, D.A.; Bleecker, E.R.; et al. Sputum neutrophil counts are associated with more severe asthma phenotypes using cluster analysis. J. Allergy Clin. Immunol. 2014, 133, 1557–1563. [Google Scholar] [CrossRef]
- Jatakanon, A.; Uasuf, C.; Maziak, W.; Lim, S.; Chung, K.F.; Barnes, P.J. Neutrophilic inflammation in severe persistent asthma. Am. J. Respir. Crit. Care Med. 1999, 160, 1532–1539. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Yang, F.; Xu, Y.; Feng, C.; Zhao, Y. The regulatory roles of neutrophils in adaptive immunity. Cell Commun. Signal 2019, 17, 147. [Google Scholar] [CrossRef]
- Giacalone, V.D.; Margaroli, C.; Mall, M.A.; Tirouvanziam, R. Neutrophil Adaptations upon Recruitment to the Lung: New Concepts and Implications for Homeostasis and Disease. Int. J. Mol. Sci. 2020, 21, 851. [Google Scholar] [CrossRef]
- Wills-Karp, M. Neutrophil ghosts worsen asthma. Sci. Immunol. 2018, 3, eaau0112. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yu, M.; Zhong, Y.; Wang, L.; Huang, H. Characteristics and Role of Neutrophil Extracellular Traps in Asthma. Inflammation. 2022, 45, 6–13. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Qin, L.; He, R.; Hu, C. Neutrophil Extracellular Trapping Network Promotes the Pathogenesis of Neutrophil-associated Asthma through Macrophages. Immunol. Invest. 2021, 50, 544–561. [Google Scholar] [CrossRef]
- Varricchi, G.; Modestino, L.; Poto, R.; Cristinziano, L.; Gentile, L.; Postiglione, L.; Spadaro, G.; Galdiero, M.R. Neutrophil extracellular traps and neutrophil-derived mediators as possible biomarkers in bronchial asthma. Clin. Exp. Med. 2022, 22, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Huang, Y.; Li, J.; Huang, J.; Zhang, L.; Feng, J.; Li, J.; Xia, Q.; Zhao, Q.; Huang, L.; et al. Eosinophil extracellular traps drive asthma progression through neuro-immune signals. Nat. Cell Biol. 2021, 23, 1060–1072. [Google Scholar] [CrossRef]
- Michaeloudes, C.; Abubakar-Waziri, H.; Lakhdar, R.; Raby, K.; Dixey, P.; Adcock, I.M.; Mumby, S.; Bhavsar, P.K.; Chung, K.F. Molecular mechanisms of oxidative stress in asthma. Mol. Aspects Med. 2022, 85, 101026. [Google Scholar] [CrossRef]
- Crisford, H.; Sapey, E.; Rogers, G.B.; Taylor, S.; Nagakumar, P.; Lokwani, R.; Simpson, J.L. Neutrophils in asthma: The good, the bad and the bacteria. Thorax 2021, 76, 835–844. [Google Scholar] [CrossRef]
- Andreadis, A.A.; Hazen, S.L.; Comhair, S.A.; Erzurum, S.C. Oxidative and nitrosative events in asthma. Free Radic Biol. Med. 2003, 35, 213–225. [Google Scholar] [CrossRef]
- Obaid Abdullah, S.; Ramadan, G.M.; Makki Al-Hindy, H.A.; Mousa, M.J.; Al-Mumin, A.; Jihad, S.; Hafidh, S.; Kadhum, Y. Serum Myeloperoxidase as a Biomarker of Asthma Severity Among Adults: A Case Control Study. Rep. Biochem. Mol. Biol. 2022, 11, 182–189. [Google Scholar] [CrossRef]
- Olgart Hoglund, C.; de Blay, F.; Oster, J.P.; Duvernelle, C.; Kassel, O.; Pauli, G.; Frossard, N. Nerve growth factor levels and localisation in human asthmatic bronchi. Eur. Respir. J. 2002, 20, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Azuma, M.; Tsunematsu, T.; Morimoto, Y.; Kondo, M.; Tezuka, T.; Nishioka, Y.; Tsuneyama, K. Neutrophils induce smooth muscle hyperplasia via neutrophil elastase-induced FGF-2 in a mouse model of asthma with mixed inflammation. Clin. Exp. Allergy 2018, 48, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kou, L.; Kong, L. Anti-nerve growth factor antibody improves airway hyperresponsiveness by down-regulating RhoA. J. Asthma 2018, 55, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, S.; Tang, L. Nerve Growth Factor: A Potential Therapeutic Target for Lung Diseases. Int. J. Mol. Sci. 2021, 22, 9112. [Google Scholar] [CrossRef] [PubMed]
- Saffar, A.S.; Ashdown, H.; Gounni, A.S. The molecular mechanisms of glucocorticoids-mediated neutrophil survival. Curr. Drug Targets 2011, 12, 556–562. [Google Scholar] [CrossRef]
- Tamura, H.; Ishikita, H. Quenching of Singlet Oxygen by Carotenoids via Ultrafast Superexchange Dynamics. J. Phys. Chem. A 2020, 124, 5081–5088. [Google Scholar] [CrossRef]
- Stratton, S.P.; Schaefer, W.H.; Liebler, D.C. Isolation and identification of singlet oxygen oxidation products of beta-carotene. Chem. Res. Toxicol. 1993, 6, 542–547. [Google Scholar] [CrossRef]
- Zbyradowski, M.; Duda, M.; Wisniewska-Becker, A.; Heriyanto; Rajwa, W.; Fiedor, J.; Cvetkovic, D.; Pilch, M.; Fiedor, L. Triplet-driven chemical reactivity of β-carotene and its biological implications. Nat. Commun. 2022, 13, 2474. [Google Scholar] [CrossRef]
- Montenegro, M.A.; Nazareno, M.A.; Durantini, E.N.; Borsarelli, C.D. Singlet molecular oxygen quenching ability of carotenoids in a reverse-micelle membrane mimetic system. Photochem. Photobiol. 2002, 75, 353–361. [Google Scholar] [CrossRef]
- Ramel, F.; Birtic, S.; Cuiné, S.; Triantaphylidès, C.; Ravanat, J.L.; Havaux, M. Chemical quenching of singlet oxygen by carotenoids in plants. Plant Physiol. 2012, 158, 1267–1278. [Google Scholar] [CrossRef]
- Kruk, J.; Szymańska, R. Singlet oxygen oxidation products of carotenoids, fatty acids and phenolic prenyllipids. J. Photochem. Photobiol. B 2021, 216, 112148. [Google Scholar] [CrossRef]
- Di Mascio, P.; Devasagayam, T.P.; Kaiser, S.; Sies, H. Carotenoids, tocopherols and thiols as biological singlet molecular oxygen quenchers. Biochem. Soc. Trans. 1990, 18, 1054–1056. [Google Scholar] [CrossRef] [PubMed]
- Cantrell, A.; McGarvey, D.J.; Truscott, T.G.; Rancan, F.; Böhm, F. Singlet oxygen quenching by dietary carotenoids in a model membrane environment. Arch. Biochem. Biophys. 2003, 412, 47–54. [Google Scholar] [CrossRef]
- Ukai, N.; Lu, Y.; Etoh, H.; Yagi, A.; Ina, K.; Oshima, S.; Ojima, F.; Sakamoto, H.; Ishiguro, Y. Photosensitized Oxygenation of Lycopene. Biosci. Biotechnol. Biochem. 1994, 58, 1718–1719. [Google Scholar] [CrossRef]
- Naik, P.; Kumar, M. Receptor interactions of constituents of Zingiber officinalis and Solanum lycopersicum on COX. J. Pharmacogn. Phytochem. 2019, 8, 4637–4641. [Google Scholar]
- Hayashi, R.; Hayashi, S.; Machida, S. Changes in Aqueous Humor Lutein Levels of Patients with Cataracts after a 6-Week Course of Lutein-Containing Antioxidant Supplementation. Curr. Eye Res. 2022, 47, 1016–1023. [Google Scholar] [CrossRef]
- Sideri, O.; Tsaousis, K.T.; Li, H.J.; Viskadouraki, M.; Tsinopoulos, I.T. The potential role of nutrition on lens pathology: A systematic review and meta-analysis. Surv. Ophthalmol. 2019, 64, 668–678. [Google Scholar] [CrossRef]
- Groten, K.; Marini, A.; Grether-Beck, S.; Jaenicke, T.; Ibbotson, S.H.; Moseley, H.; Ferguson, J.; Krutmann, J. Tomato Phytonutrients Balance UV Response: Results from a Double-Blind, Randomized, Placebo-Controlled Study. Skin Pharmacol. Physiol. 2019, 32, 101–108. [Google Scholar] [CrossRef]
- Fam, V.W.; Holt, R.R.; Keen, C.L.; Sivamani, R.K.; Hackman, R.M. Prospective Evaluation of Mango Fruit Intake on Facial Wrinkles and Erythema in Postmenopausal Women: A Randomized Clinical Pilot Study. Nutrients 2020, 12, 3381. [Google Scholar] [CrossRef]
- Ouchi, A.; Aizawa, K.; Iwasaki, Y.; Inakuma, T.; Terao, J.; Nagaoka, S.; Mukai, K. Kinetic study of the quenching reaction of singlet oxygen by carotenoids and food extracts in solution. Development of a singlet oxygen absorption capacity (SOAC) assay method. J. Agric. Food Chem. 2010, 58, 9967–9978. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Inokami, Y.; Tokumura, A.; Terao, J.; Suzuki, A. Rate constants for quenching singlet oxygen and activities for inhibiting lipid peroxidation of carotenoids and alpha-tocopherol in liposomes. Lipids 1998, 33, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Foote, C.S.; Ching, T.Y.; Geller, G.G. Chemistry of singlet oxygen. XVIII. Rates of reaction and quenching of alpha-tocopherol and singlet oxygen. Photochem. Photobiol. 1974, 20, 511–513. [Google Scholar] [PubMed]
- Gruszka, J.; Pawlak, A.; Kruk, J. Tocochromanols, plastoquinol, and other biological prenyllipids as singlet oxygen quenchers-determination of singlet oxygen quenching rate constants and oxidation products. Free Radic. Biol. Med. 2008, 45, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Shichiri, M.; Yoshida, Y.; Ishida, N.; Hagihara, Y.; Iwahashi, H.; Tamai, H.; Niki, E. α-Tocopherol suppresses lipid peroxidation and behavioral and cognitive impairments in the Ts65Dn mouse model of Down syndrome. Free Radic. Biol. Med. 2011, 50, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Shichiri, M.; Harada, N.; Ishida, N.; Komaba, L.K.; Iwaki, S.; Hagihara, Y.; Niki, E.; Yoshida, Y. Oxidative stress is involved in fatigue induced by overnight deskwork as assessed by increase in plasma tocopherylhydroqinone and hydroxycholesterol. Biol. Psychol. 2013, 94, 527–533. [Google Scholar] [CrossRef]
- Shichiri, M.; Ishida, N.; Hagihara, Y.; Yoshida, Y.; Kume, A.; Suzuki, H. Probucol induces the generation of lipid peroxidation products in erythrocytes and plasma of male cynomolgus macaques. J. Clin. Biochem. Nutr. 2019, 64, 129–142. [Google Scholar] [CrossRef]
- Krasnovsky, A.A.; Kagan, V.E.; Minin, A.A. Quenching of singlet oxygen luminescence by fatty acids and lipids: Contribution of physical and chemical mechanisms. FEBS Lett. 1983, 155, 233–236. [Google Scholar] [CrossRef]
- Chaudhuri, R.K.; Bojanowski, K. Bakuchiol: A retinol-like functional compound revealed by gene expression profiling and clinically proven to have anti-aging effects. Int. J. Cosmet. Sci. 2014, 36, 221–230. [Google Scholar] [CrossRef]
- Chaudhuri, R.K.; Marchio, F. Bakuchiol in the Management of Acne-affected Skin. Cosmet. Toilet. 2011, 126, 502–510. [Google Scholar]
- Dhaliwal, S.; Rybak, I.; Ellis, S.R.; Notay, M.; Trivedi, M.; Burney, W.; Vaughn, A.R.; Nguyen, M.; Reiter, P.; Bosanac, S.; et al. Prospective, randomized, double-blind assessment of topical bakuchiol and retinol for facial photoageing. Br. J. Dermatol. 2019, 180, 289–296. [Google Scholar] [CrossRef]
- Krishnakantha, T.P.; Lokesh, B.R. Scavenging of superoxide anions by spice principles. Indian J. Biochem. Biophys. 1993, 30, 133–134. [Google Scholar]
- Swindell, W.R.; Bojanowski, K.; Chaudhuri, R.K. A Zingerone Analog, Acetyl Zingerone, Bolsters Matrisome Synthesis, Inhibits Matrix Metallopeptidases, and Represses IL-17A Target Gene Expression. J. Invest. Dermatol. 2020, 140, 602–614.e615. [Google Scholar] [CrossRef] [PubMed]
- Das, K.C.; Das, C.K. Curcumin (diferuloylmethane), a singlet oxygen ((1)O(2)) quencher. Biochem. Biophys. Res. Commun. 2002, 295, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.K.; Meyer, T.; Premi, S.; Brash, D. Acetyl zingerone: An efficacious multifunctional ingredient for continued protection against ongoing DNA damage in melanocytes after sun exposure ends. Int. J. Cosmet. Sci. 2020, 42, 36–45. [Google Scholar] [CrossRef]
- Dhaliwal, S.; Rybak, I.; Pourang, A.; Burney, W.; Haas, K.; Sandhu, S.; Crawford, R.; Sivamani, R.K. Randomized double-blind vehicle controlled study of the effects of topical acetyl zingerone on photoaging. J. Cosmet. Dermatol. 2021, 20, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef] [PubMed]













| Detection Method of 1O2 | Principle | Pros/Cons | Ref. |
|---|---|---|---|
| Near-infrared luminescence | 1O2 emission at 1270 nm |
| [54,57,58,59] |
| ESR spectroscopy | Measurement of nitroxide radicals produced by the reaction of 1O2 with sterically hindered secondary amine probes |
| [48,60,61,65,66] |
| Fluorescent probes | [67,68,69] | ||
| 9,10-dimethylanthracene (DMA) | React with 1O2 and form stable endoperoxides with high-fluorescence quantum yield (high sensitivity, simplicity of data acquisition, and high spatial resolution in microscopic imaging) |
| [68,70,71,72] |
| 9-[2-(3-Carboxy-9,10-diphenyl)anthryl]-6-hydroxy-3H-xanthen-3-ones (DPAXs) | |||
| Singlet Oxygen Sensor Green (SOSG) |
| [73,74,75] | |
| SOSG-based nanosensors NanoSOSG SPR-SOSG PAM-SOSG |
| [50,78,79] | |
| Si-DMA |
| [76,77] | |
| 1O2-mediated peroxidation products | |||
| 10- and 12-(Z,E)-HODEs | Products mediated by the reaction of 1O2 with linoleic acid |
| [83,89,90,91,92,93] |
| Cholesterol 5α-OOH Cholesterol 6α-OOH Cholesterol 6β-OOH | Products mediated by the reaction of 1O2 with cholesterol |
| [52] |
| 1O2-Mediated Oxidation Product | Sample Species | Disease/Model | Ref. |
|---|---|---|---|
| 10- and/or 12-(Z,E)-HODE | Human | Borderline diabetes/oral glucose tolerance test | [92,93] |
| Human | High serum levels in patients with primary open-angle glaucoma | [94] | |
| Pig | Increased by kidney tissue damage | [95] | |
| Mouse | High in the plasma of model mice with prediabetes | [83,96] | |
| Mouse | Increased in the lungs of model mice with asthma and positively correlated with MPO activity and nerve growth factor (NGF) | [97] | |
| Cholesterol 5α-hydroperoxide | Human | Alcoholic patients | [98] |
| Rat | Pheophorubide a and visible light irradiation | [99] | |
| Mouse | UVA irradiation of hairless mice | [100] | |
| Mouse | Effect of beta-carotene on UVA irradiation of hairless mice | [101,102] | |
| Cholesterol 5,6-secosterol | Human | Cholesterol 5,6-secosterol detected in atherosclerotic plaques | [103] |
| Human | Cortex in patients with Alzheimer’s disease | [104] | |
| Rat | Higher in the plasma of amyotrophic lateral sclerosis (ALS) rats than before disease onset | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murotomi, K.; Umeno, A.; Shichiri, M.; Tanito, M.; Yoshida, Y. Significance of Singlet Oxygen Molecule in Pathologies. Int. J. Mol. Sci. 2023, 24, 2739. https://doi.org/10.3390/ijms24032739
Murotomi K, Umeno A, Shichiri M, Tanito M, Yoshida Y. Significance of Singlet Oxygen Molecule in Pathologies. International Journal of Molecular Sciences. 2023; 24(3):2739. https://doi.org/10.3390/ijms24032739
Chicago/Turabian StyleMurotomi, Kazutoshi, Aya Umeno, Mototada Shichiri, Masaki Tanito, and Yasukazu Yoshida. 2023. "Significance of Singlet Oxygen Molecule in Pathologies" International Journal of Molecular Sciences 24, no. 3: 2739. https://doi.org/10.3390/ijms24032739
APA StyleMurotomi, K., Umeno, A., Shichiri, M., Tanito, M., & Yoshida, Y. (2023). Significance of Singlet Oxygen Molecule in Pathologies. International Journal of Molecular Sciences, 24(3), 2739. https://doi.org/10.3390/ijms24032739