

Investigation of the Effects of Monomeric and Dimeric Stilbenoids on Bacteria-Induced Cytokines and LPS-Induced ROS Formation in Bone Marrow-Derived Dendritic Cells

 , , , , , ,
, , , , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Methylation of Resveratrol Diminishes Its Inhibitory Effect on the L. Acidophilus NCFM-Induced Cytokine Response
2.2. Monomer Derivatives Show Potent IL-12 Inhibition but Weak Inhibition of IL-10 and TNF-α Production in bmDCs Stimulated with E. Coli Nissle 1917
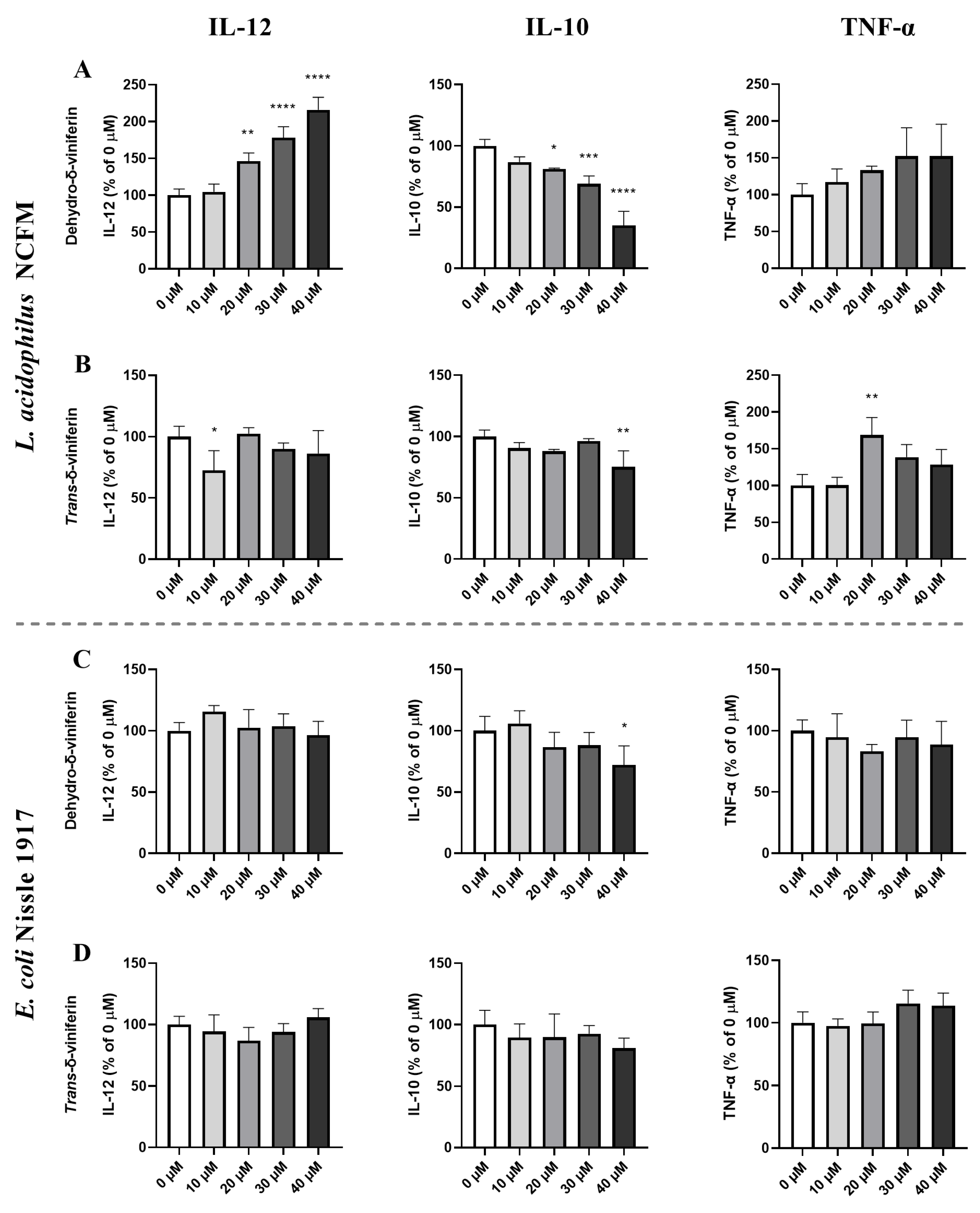
2.3. Effect of Dimeric Stilbenoids on the Bacterially Induced Cytokine Response
2.4. Number and Position of Hydroxy Groups Affect Stilbenoids Inhibition of Intracellular ROS
2.5. Determination of the Total Antioxidant Capacity of Resveratrol and Resveratrol Derivatives
2.6. Resveratrol and Dehydro-δ-Viniferin Affect Map Kinases Divergently
- The striking difference in L. acidophilus NCFM-induced IL-12 production elicited by resveratrol and dehydro-δ-viniferin prompted us to investigate the mechanisms underlying the divergent activities of the two stilbenoids. Accordingly, it has been investigated how inhibitors of the Jun N-terminal kinase (JNK) and p38 affected the IL-12 and IL-10 responses to stimulation with L. acidophilus NCFM and how L. acidophilus (used alone or after a resveratrol or dehydro-δ-viniferin pretreatment) affected the expression of dual specificity phosphatase 1 (Dusp1) (Figure 6). While JNK inhibition gave a potent inhibition of IL-12 (73%) and IL-10 (96%) production, inhibition of p38 inhibition had only minor effects on IL-12 (11% inhibition), but strongly impaired IL-10 production (76% inhibition, Figure 6A). Thus, JNK activation seems indispensable for L. acidophilus NCFM-induced effects on IL-12 production.
3. Discussion
4. Materials and Methods
4.1. Generation of Murine Bone Marrow-Derived Dendritic Cells (bmDCs)
4.2. Bacterial Strains and Stilbenoid Synthesis
4.3. Assessment of Bacteria-Induced Cytokine Response
4.4. Assessment of Intracellular ROS Formation
4.5. In Vitro Total Antioxidant Capacity (TAC) Determination
4.6. Assessment of Fold Change in Gene Expression in bmDC
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Remsberg, C.M.; Yáñez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. 2008, 22, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Teuscher, K.B.; Zhang, M.; Ji, H. A Versatile Method to Determine the Cellular Bioavailability of Small-Molecule Inhibitors. J. Med. Chem. 2017, 60, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M.; Nussenzweig, M.C. Avoiding horror autotoxicus: The importance of dendritic cells in peripheral T cell tolerance. Proc. Natl. Acad. Sci. USA 2002, 99, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Mathiesen, R.; Eld, H.M.S.; Sørensen, J.; Fuglsang, E.; Lund, L.D.; Taverniti, V.; Frøkiær, H. Mannan Enhances IL-12 Production by Increasing Bacterial Uptake and Endosomal Degradation in L. acidophilus and S. aureus Stimulated Dendritic Cells. Front. Immunol. 2019, 10, 2646. [Google Scholar] [CrossRef]
- Mikkelsen, M.S.; Jespersen, B.M.; Mehlsen, A.; Engelsen, S.B.; Frøkiær, H. Cereal β-glucan immune modulating activity depends on the polymer fine structure. Food Res. Int. 2014, 62, 829–836. [Google Scholar] [CrossRef]
- Weiss, G.; Maaetoft-Udsen, K.; Stifter, S.A.; Hertzog, P.; Goriely, S.; Thomsen, A.R.; Paludan, S.R.; Frøkiær, H. MyD88 drives the IFN-β response to Lactobacillus acidophilus in dendritic cells through a mechanism involving IRF1, IRF3, and IRF7. J. Immunol. 2012, 189, 2860–2868. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Dvorakova, M.; Landa, P. Anti-inflammatory activity of natural stilbenoids: A review. Pharmacol. Res. 2017, 124, 126–145. [Google Scholar] [CrossRef]
- Mattio, L.M.; Dallavalle, S.; Musso, L.; Filardi, R.; Franzetti, L.; Pellegrino, L.; D’Incecco, P.; Mora, D.; Pinto, A.; Arioli, S. Antimicrobial activity of resveratrol-derived monomers and dimers against foodborne pathogens. Sci. Rep. 2019, 9, 19525. [Google Scholar] [CrossRef]
- Zeuthen, L.H.; Christensen, H.R.; Frøkiaer, H. Lactic acid bacteria inducing a weak interleukin-12 and tumor necrosis factor alpha response in human dendritic cells inhibit strongly stimulating lactic acid bacteria but act synergistically with gram-negative bacteria. Clin. Vaccine Immunol. 2006, 13, 365–375. [Google Scholar] [CrossRef]
- Boye, L.; Welsby, I.; Lund, L.D.; Goriely, S.; Frøkiaer, H. Plasma membrane Toll-like receptor activation increases bacterial uptake but abrogates endosomal Lactobacillus acidophilus induction of interferon-β. Immunology 2016, 149, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Saikh, K.U. MyD88 and beyond: A perspective on MyD88-targeted therapeutic approach for modulation of host immunity. Immunol. Res. 2021, 69, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Tian, S.; Fu, M.; Li, Y.; Sun, Y.; Liu, J.; Liu, Y. Resveratrol protects human bronchial epithelial cells against nickel-induced toxicity via suppressing p38 MAPK, NF-κB signaling, and NLRP3 inflammasome activation. Environ. Toxicol. 2020, 35, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Lv, R.; Wang, L.; Hou, H.; Liu, H.; Shao, S. Resveratrol, an antioxidant, protects spinal cord injury in rats by suppressing MAPK pathway. Saudi J. Biol. Sci. 2018, 25, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Eld, H.M.S.; Nielsen, E.M.; Johnsen, P.R.; Marengo, M.; Kamper, I.W.; Frederiksen, L.; Bonomi, F.; Frees, D.; Iametti, S.; Frøkiær, H. Cefoxitin treatment of MRSA leads to a shift in the IL-12/IL-23 production pattern in dendritic cells by a mechanism involving changes in the MAPK signaling. Mol. Immunol. 2021, 134, 1–12. [Google Scholar] [CrossRef]
- Weiss, G.; Christensen, H.R.; Zeuthen, L.H.; Vogensen, F.K.; Jakobsen, M.; Frøkiær, H. Lactobacilli and bifidobacteria induce differential interferon-β profiles in dendritic cells. Cytokine 2011, 56, 520–530. [Google Scholar] [CrossRef]
- Teixeira-Coelho, M.; Guedes, J.; Ferreirinha, P.; Howes, A.; Pedrosa, J.; Rodrigues, F.; Lai, W.S.; Blackshear, P.J.; O’Garra, A.; Castro, A.G.; et al. Differential post-transcriptional regulation of IL-10 by TLR2 and TLR4-activated macrophages. Eur. J. Immunol. 2014, 44, 856–866. [Google Scholar] [CrossRef]
- Eld, H.M.S.; Johnsen, P.R.; Nielsen, E.M.; Jørgensen, F.Z.; Lindstrøm-Svendsen, M.; Baldry, M.; Ingmer, H.; Frøkiær, H. Soluble C-Type Lectin-Receptor Ligands Stimulate ROS Production in Dendritic Cells and Potentiate Killing of MRSA as Well as the MRSA Induced IL-12 Production. Front. Immunol. 2022, 13, 845881. [Google Scholar] [CrossRef]
- Arthur, J.S.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef]
- Huang, G.; Shi, L.Z.; Chi, H. Regulation of JNK and p38 MAPK in the immune system: Signal integration, propagation and termination. Cytokine 2009, 48, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Dias, S.J.; Li, K.; Rimando, A.M.; Dhar, S.; Mizuno, C.S.; Penman, A.D.; Levenson, A.S. Trimethoxy-Resveratrol and Piceatannol Administered Orally Suppress and Inhibit Tumor Formation and Growth in Prostate Cancer Xenografts. Prostate 2013, 73, 1135–1146. [Google Scholar] [CrossRef]
- Fulda, S. Resveratrol and derivatives for the prevention and treatment of cancer. Drug Discov. Today 2010, 15, 757–765. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, D.; Zheng, P.; Yu, J.; He, J.; Mao, X.; Yu, B. The Bidirectional Interactions between Resveratrol and Gut Microbiota: An Insight into Oxidative Stress and Inflammatory Bowel Disease Therapy. Biomed. Res. Int. 2019, 2019, 5403761. [Google Scholar] [CrossRef]
- Li, H.; Jiang, N.; Liang, B.; Liu, Q.; Zhang, E.; Peng, L.; Deng, H.; Li, R.; Li, Z.; Zhu, H. Pterostilbene protects against UVB-induced photo-damage through a phosphatidylinositol-3-kinase-dependent Nrf2/ARE pathway in human keratinocytes. Redox Rep. 2017, 22, 501–507. [Google Scholar] [CrossRef]
- Soeur, J.; Eilstein, J.; Léreaux, G.; Jones, C.; Marrot, L. Skin resistance to oxidative stress induced by resveratrol: From Nrf2 activation to GSH biosynthesis. Free Radic. Biol. Med. 2015, 78, 213–223. [Google Scholar] [CrossRef]
- Wahdan, S.A.; Azab, S.S.; Elsherbiny, D.A.; El-Demerdash, E. Piceatannol protects against cisplatin nephrotoxicity via activation of Nrf2/HO-1 pathway and hindering NF-κB inflammatory cascade. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Diez, C.; Miguel, V.; Mennerich, D.; Kietzmann, T.; Sánchez-Pérez, P.; Cadenas, S.; Lamas, S. Antioxidant responses and cellular adjustments to oxidative stress. Redox Biol. 2015, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Radical chemistry of flavonoid antioxidants. Adv. Exp. Med. Biol. 1990, 264, 165–170. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Tybussek, T.; Herfellner, T.; Schneider, F.; Schweiggert-Weisz, U.; Eisner, P. Radical Scavenging Mechanisms of Phenolic Compounds: A Quantitative Structure-Property Relationship (QSPR) Study. Front. Nutr. 2022, 9, 882458. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Miesbauer, O.; Eisner, P. Common Trends and Differences in Antioxidant Activity Analysis of Phenolic Substances Using Single Electron Transfer Based Assays. Molecules 2021, 26, 1244. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; He, L.; Hou, X.; Wei, J.; Ma, X.; Gao, Z.; Yuan, Y.; Xiao, J.; Li, P.; Yue, T. Relationships between Structure and Antioxidant Capacity and Activity of Glycosylated Flavonols. Foods 2021, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Charrel-Dennis, M.; Latz, E.; Halmen, K.A.; Trieu-Cuot, P.; Fitzgerald, K.A.; Kasper, D.L.; Golenbock, D.T. TLR-independent type I interferon induction in response to an extracellular bacterial pathogen via intracellular recognition of its DNA. Cell Host Microbe 2008, 4, 543–554. [Google Scholar] [CrossRef]
- Lund, L.D.; Ingmer, H.; Frøkiær, H. D-Alanylation of Teichoic Acids and Loss of Poly-N-Acetyl Glucosamine in Staphylococcus aureus during Exponential Growth Phase Enhance IL-12 Production in Murine Dendritic Cells. PLoS ONE 2016, 11, e0149092. [Google Scholar] [CrossRef] [PubMed]
- Mackowiak, P.A. Relationship between growth temperature and shedding of lipopolysaccharides by gram-negative bacilli. Eur. J. Clin. Microbiol. 1984, 3, 406–410. [Google Scholar] [CrossRef]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.M.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.; Furuya, Y.; Roberts, S.; Metzger, D.W. Role of Interleukin-12 in Protection against Pulmonary Infection with Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 6308–6316. [Google Scholar] [CrossRef]
- Christensen, H.R.; Frøkiaer, H.; Pestka, J.J. Lactobacilli differentially modulate expression of cytokines and maturation surface markers in murine dendritic cells. J. Immunol. 2002, 168, 171–178. [Google Scholar] [CrossRef]
- Valli, V.; Gómez-Caravaca, A.M.; Di Nunzio, M.; Danesi, F.; Caboni, M.F.; Bordoni, A. Sugar cane and sugar beet molasses, antioxidant-rich alternatives to refined sugar. J. Agric. Food Chem. 2012, 60, 12508–12515. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnsen, P.R.; Pinna, C.; Mattio, L.; Strube, M.B.; Di Nunzio, M.; Iametti, S.; Dallavalle, S.; Pinto, A.; Frøkiær, H. Investigation of the Effects of Monomeric and Dimeric Stilbenoids on Bacteria-Induced Cytokines and LPS-Induced ROS Formation in Bone Marrow-Derived Dendritic Cells. Int. J. Mol. Sci. 2023, 24, 2731. https://doi.org/10.3390/ijms24032731
Johnsen PR, Pinna C, Mattio L, Strube MB, Di Nunzio M, Iametti S, Dallavalle S, Pinto A, Frøkiær H. Investigation of the Effects of Monomeric and Dimeric Stilbenoids on Bacteria-Induced Cytokines and LPS-Induced ROS Formation in Bone Marrow-Derived Dendritic Cells. International Journal of Molecular Sciences. 2023; 24(3):2731. https://doi.org/10.3390/ijms24032731
Chicago/Turabian StyleJohnsen, Peter Riber, Cecilia Pinna, Luce Mattio, Mathilde Bech Strube, Mattia Di Nunzio, Stefania Iametti, Sabrina Dallavalle, Andrea Pinto, and Hanne Frøkiær. 2023. "Investigation of the Effects of Monomeric and Dimeric Stilbenoids on Bacteria-Induced Cytokines and LPS-Induced ROS Formation in Bone Marrow-Derived Dendritic Cells" International Journal of Molecular Sciences 24, no. 3: 2731. https://doi.org/10.3390/ijms24032731
APA StyleJohnsen, P. R., Pinna, C., Mattio, L., Strube, M. B., Di Nunzio, M., Iametti, S., Dallavalle, S., Pinto, A., & Frøkiær, H. (2023). Investigation of the Effects of Monomeric and Dimeric Stilbenoids on Bacteria-Induced Cytokines and LPS-Induced ROS Formation in Bone Marrow-Derived Dendritic Cells. International Journal of Molecular Sciences, 24(3), 2731. https://doi.org/10.3390/ijms24032731