
Short-Term and Long-Term Effects after Exposure to Ionizing Radiation and Visible Light on Retina and Retinal Pigment Epithelium of Mouse Eye


Abstract
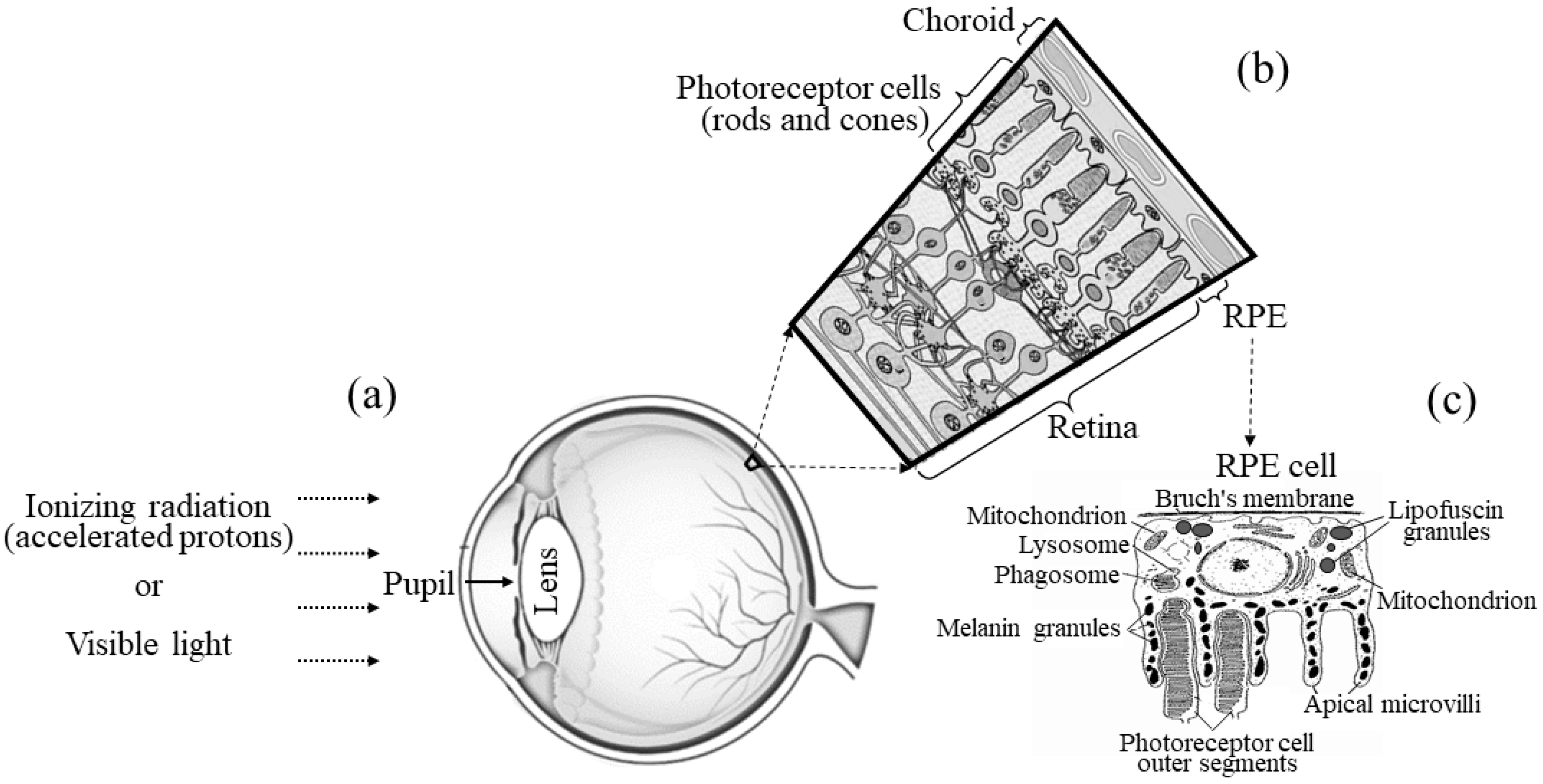
1. Introduction
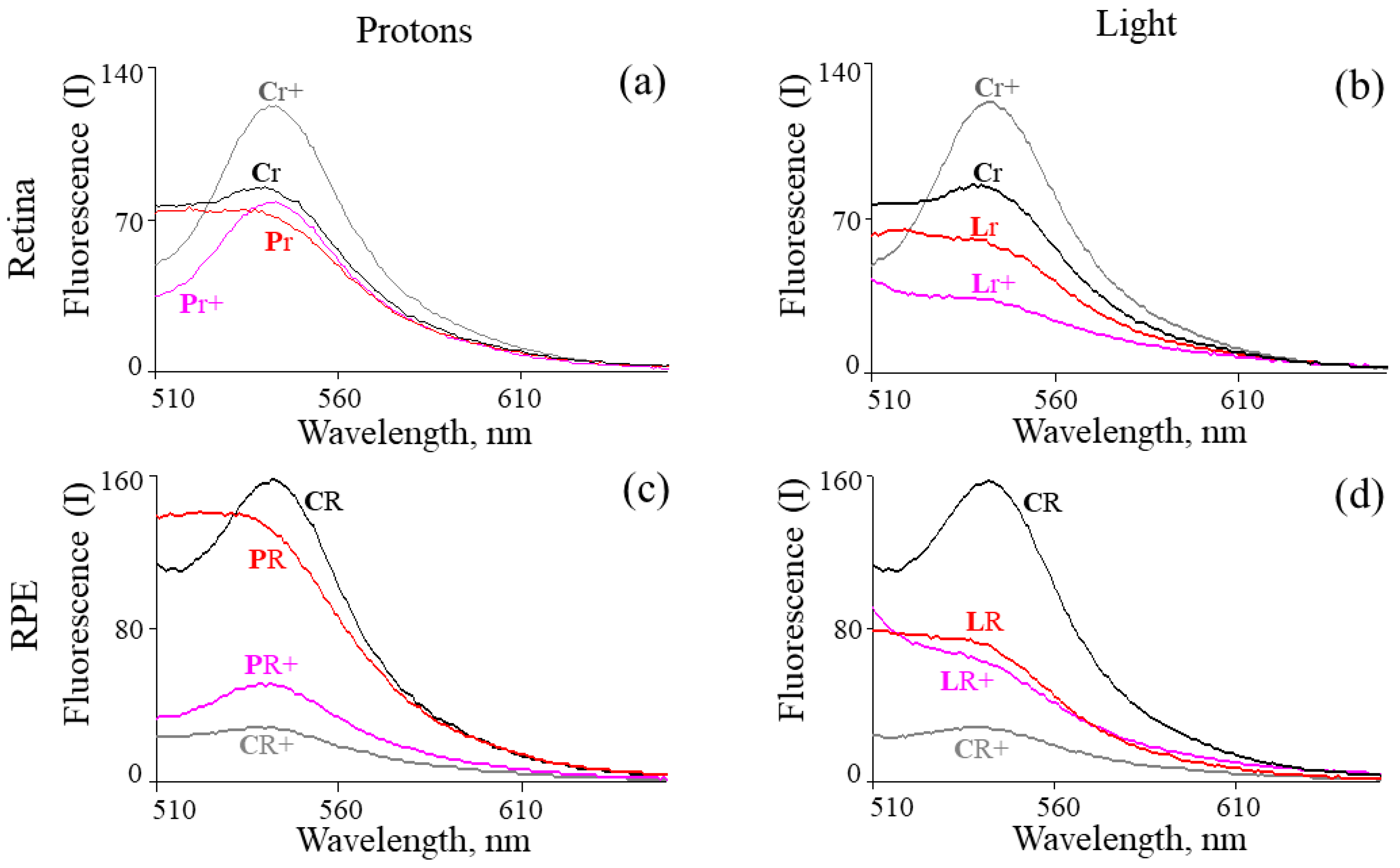
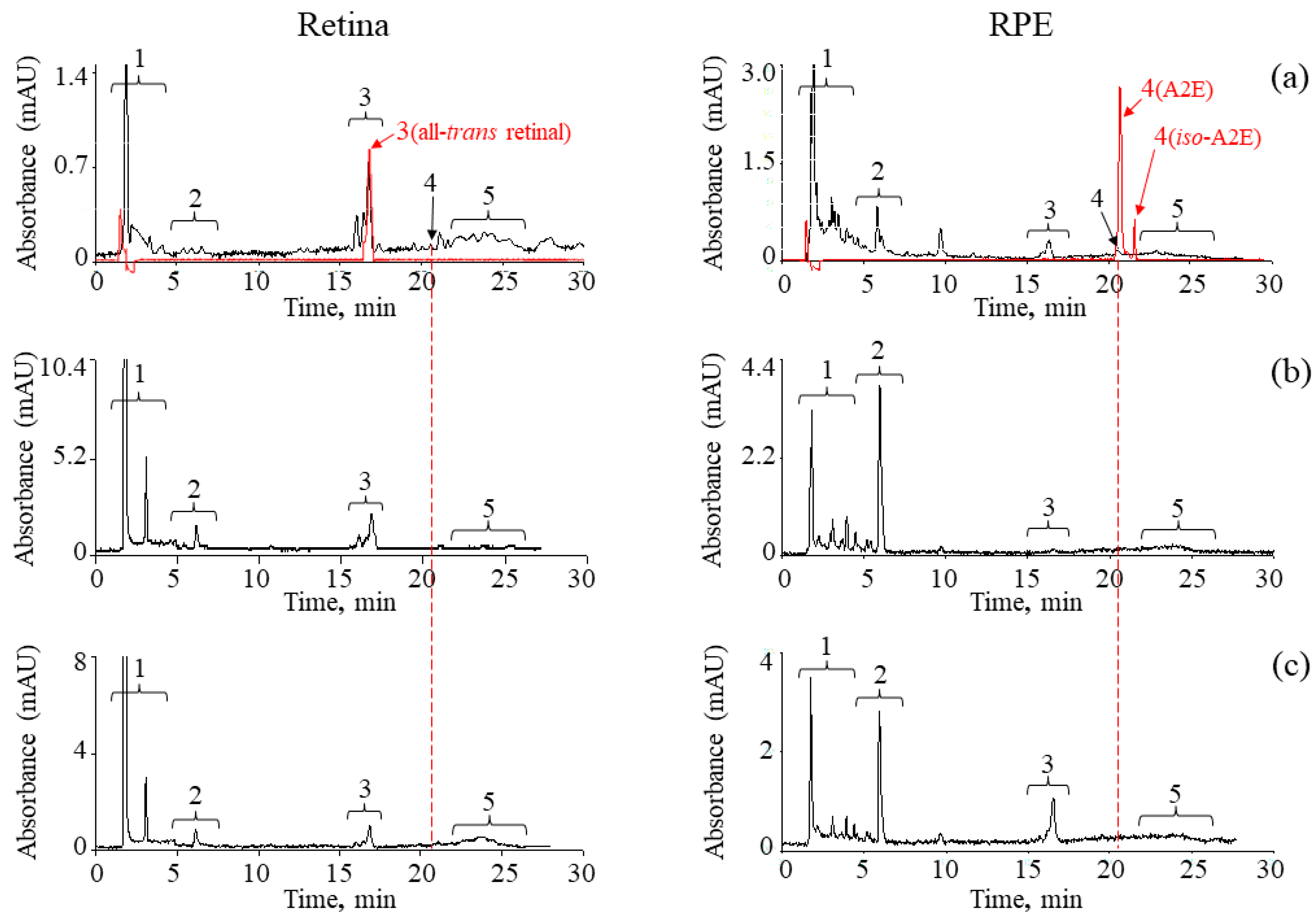
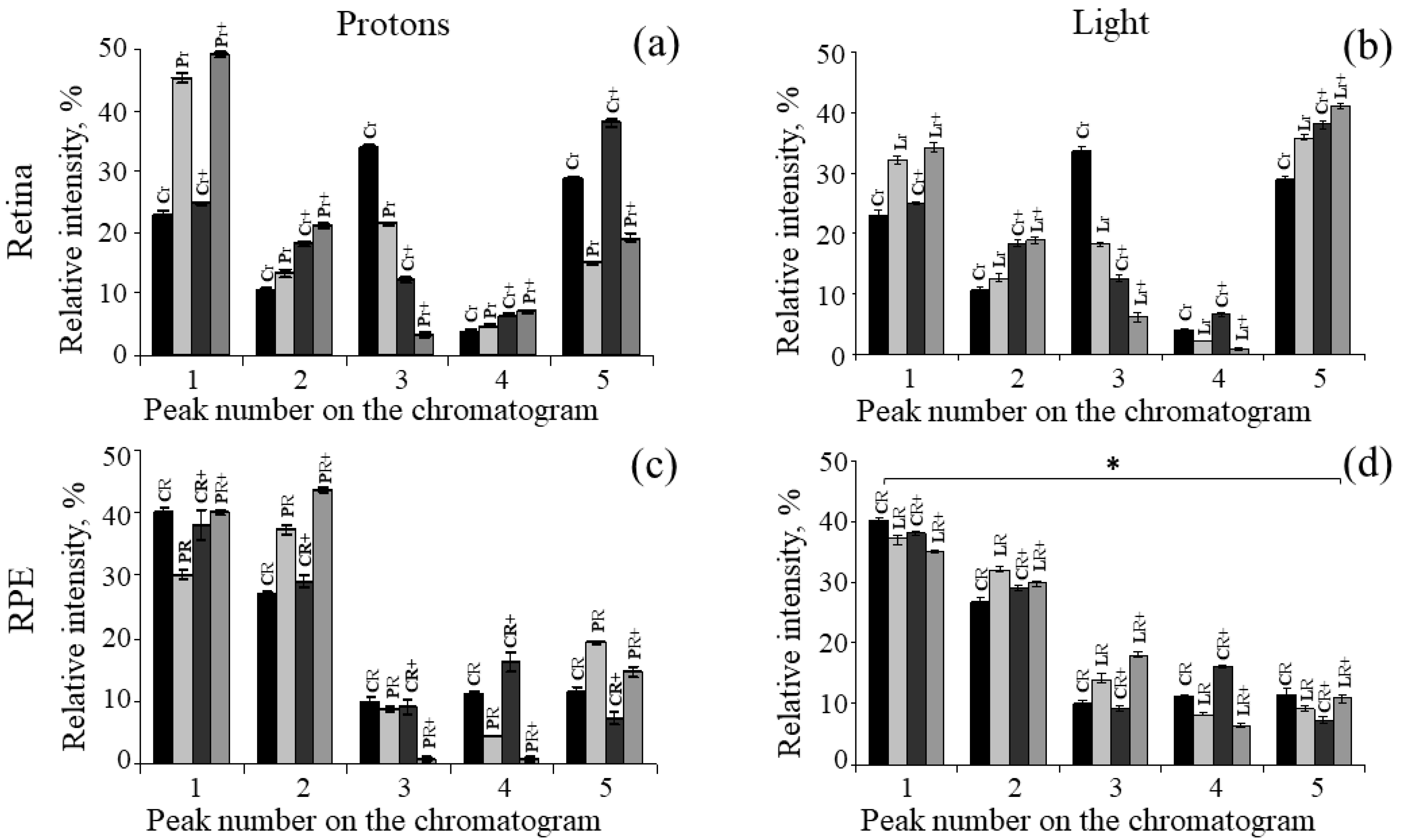
2. Results
3. Discussion
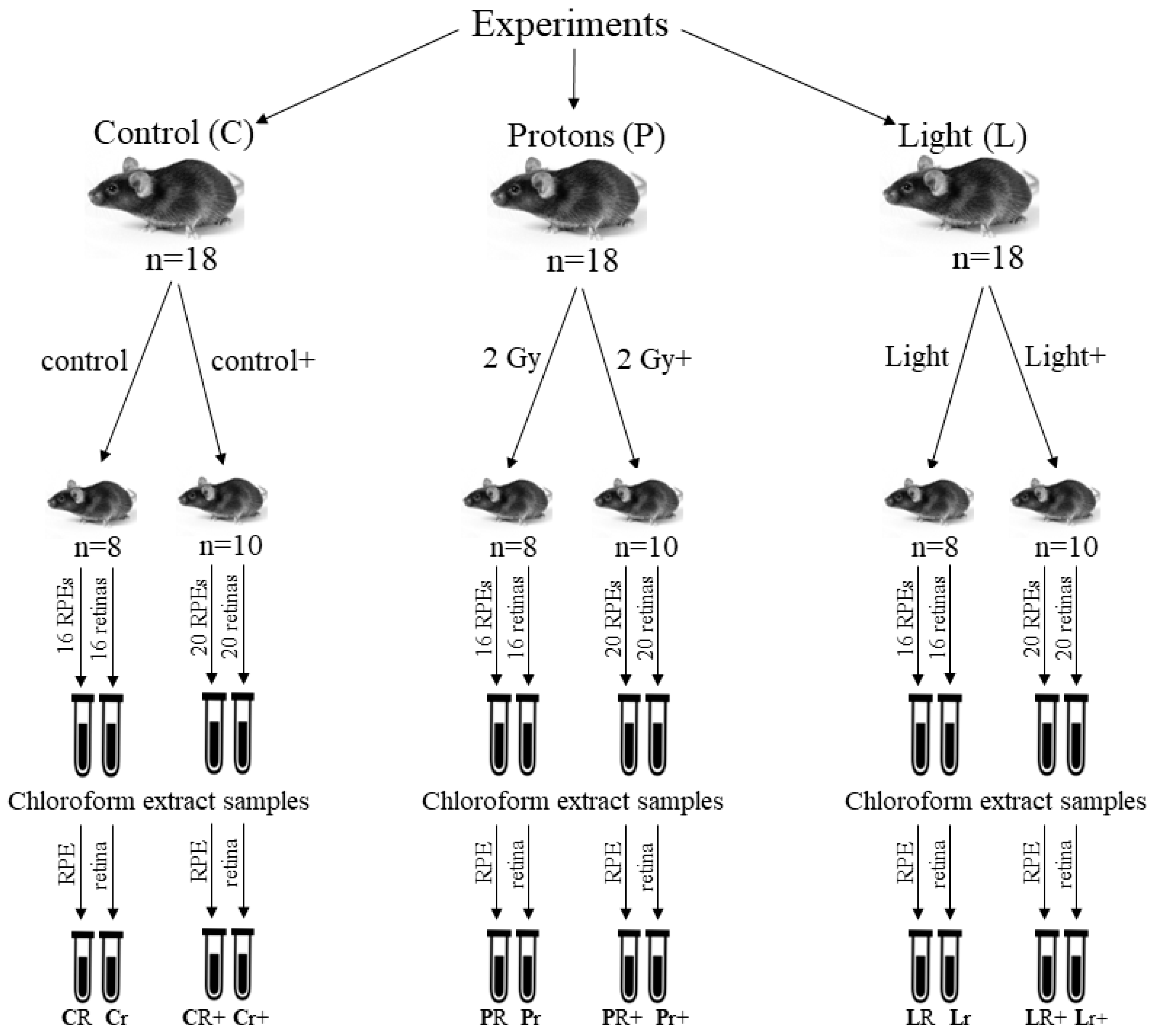
4. Materials and Methods
4.1. Reagents
4.2. Animals
4.3. Irradiation of Mice with Accelerated Protons
4.4. Irradiation of Mice with Visible Light
4.5. Fluorescent Analysis of Chloroform Extracts of Retinoids from the Retina and RPE
4.6. High Performance Liquid Chromatography (HPLC)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kennedy, A.R. Biological effects of space radiation and development of effective countermeasures. Life Sci. Space Res. 2014, 1, 10–43. [Google Scholar] [CrossRef]
- Sanzari, J.K.; Wan, X.S.; Wroe, A.J.; Rightnar, S.; Cengel, K.A.; Diffenderfer, E.S.; Krigsfeld, G.S.; Gridley, D.S.; Kennedy, A.R. Acute hematological effects of solar particle event proton radiation in the porcine model. Radiat. Res. 2013, 180, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Finnberg, N.; Wambi, C.; Kennedy, A.R.; El-Deiry, W.S. The effects of antioxidants on gene expression following gamma-radiation (GR) and proton radiation (PR) in mice in vivo. Cell Cycle 2013, 12, 2241–2247. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kennedy, A.R. Effects of dietary supplements on the space radiation-induced reduction in total antioxidant status in CBA mice. Radiat. Res. 2006, 165, 373–378. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Powell, D.R.; Li, Z.; Bell, J.S.K.; Barwick, B.G.; Feng, H.; McCrary, M.R.; Dwivedi, B.; Kowalski, J.; Dynan, W.S.; et al. Galactic cosmic radiation induces persistent epigenome alterations relevant to human lung cancer. Sci. Rep. 2018, 8, 6709. [Google Scholar] [CrossRef] [PubMed]
- Sonnenfeld, G.; Butel, J.S.; Shearer, W.T. Effects of the space flight environment on the immune system. Rev. Environ. Health 2003, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.E.; Stowe, R.P.; Pierson, D.L.; Sams, C.F. Immune system dysregulation following short-vs longduration spaceflight. Aviat. Space Environ. Med. 2008, 79, 835–843. [Google Scholar] [CrossRef] [PubMed]
- von Sonntag, C. The Chemical Basis of Radiation Biology; Taylor&Francis: London, UK, 1987. [Google Scholar]
- Alwood, J.S.; Tran, L.H.; Schreurs, A.S.; Shirazi-Fard, Y.; Kumar, A.; Hilton, D.; Tahimic, C.G.T.; Globus, R.K. Dose- and ion-dependent effects in the oxidative stress response to space-like radiation exposure in the skeletal system. Int. J. Mol. Sci. 2017, 18, 2117. [Google Scholar] [CrossRef]
- Guan, J.; Wan, X.S.; Zhou, Z.; Ware, J.; Donahue, J.J.; Biaglow, J.E.; Kennedy, A.R. Effects of dietary supplements on space radiation-induced oxidative stress in Sprague-Dawley rats. Radiat. Res. 2004, 162, 572–579. [Google Scholar] [CrossRef]
- Kobashigawa, S.; Suzuki, K.; Yamashita, S. Ionizing radiation accelerates Drp1-dependent mitochondrial fission, which involves delayed mitochondrial reactive oxygen species production in normal human fibroblast-like cells. Biochem. Biophys. Res. Commun. 2011, 414, 795–800. [Google Scholar] [CrossRef]
- Strauss, O. The retinal pigment epithelium in visual function. Physiol. Rev. 2005, 85, 845–881. [Google Scholar] [CrossRef]
- Yakovleva, M.A.; Feldman, T.B.; Lyakhova, K.N.; Utina, D.M.; Kolesnikova, I.A.; Vinogradova, Y.V.; Molokanov, A.G.; Ostrovsky, M.A. Ionized radiation-mediated retinoid oxidation in the retina and retinal pigment epithelium of the murine eye. Radiat. Res. 2022, 197, 270–279. [Google Scholar] [CrossRef]
- Gorgels, T.G.; van der Pluijm, I.; Brandt, R.M.; Garinis, G.A.; van Steeg, H.; van den Aardweg, G.; Jansen, G.H.; Ruijter, J.M.; Bergen, A.A.; van Norren, D.; et al. Retinal degeneration and ionizing radiation hypersensitivity in a mouse model for Cockayne syndrome. Mol. Cell. Biol. 2007, 27, 1433–1441. [Google Scholar] [CrossRef]
- Ponomarev, A.L.; Nounu, H.N.; Hussein, H.F.; Kim, M.-H.Y.; Atwell, W.; Cucinotta, F.A. NASA-Developed ProE-Based Tool for the Ray-Tracing of Spacecraft Geometry to Determine Radiation Doses and Particle Fluxes in Habitable Areas of Spacecraft and in the Human Body; NASA/TP-2007-214770; NASA/USRA: Cocoa Beach, FL, USA, 2007. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Manuel, F.K.; Jones, J.; Iszard, G.; Murrey, J.; Djojonegro, B.; Wear, M. Space radiation and cataracts in astronauts. Radiat. Res. 2001, 156 Pt 1, 460–466. [Google Scholar] [CrossRef]
- Mao, X.W.; Boerma, M.; Rodriguez, D.; Campbell-Beachler, M.; Jones, T.; Stanbouly, S.; Sridharan, V.; Wroe, A.; Nelson, G.A. Acute effect of low-dose space radiation on mouse retina and retinal endothelial cells. Radiat. Res. 2018, 190, 45–52. [Google Scholar] [CrossRef]
- Mao, X.W.; Archambeau, J.O.; Kubinova, L.; Boyle, S.; Petersen, G.; Grove, R. Quantification of rat retinal growth and vascular population changes after single and split doses of proton irradiation: Translational study using stereology methods. Radiat. Res. 2003, 160, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Pecaut, M.J.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Bouxsein, M.; Jones, T.A.; Moldovan, M.; Cunningham, C.E.; Chieu, J.; et al. Space flight environment induces mitochondrial oxidative damage in ocular tissue. Radiat. Res. 2013, 180, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Favre, C.; Fike, J.R.; Kubinova, L.; Anderson, E.; Campbell-Beachler, M.; Jones, T.; Smith, A.; Rightnar, S.; Nelson, G.A. High-LET radiation-induced response of microvessels in the hippocampus. Radiat. Res. 2010, 173, 486–493. [Google Scholar] [CrossRef] [PubMed]
- van Reyk, D.M.; Gillies, M.C.; Davies, M.J. The retina: Oxidative stress and diabetes. Redox Rep. 2003, 8, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Yakovleva, M.A.; Dontsov, A.E.; Trofimova, N.N.; Sakina, N.L.; Kononikhin, A.S.; Aybush, A.V.; Feldman, T.B.; Ostrovsky, M.A. Lipofuscin granule bisretinoid oxidation in the human retinal pigment epithelium forms cytotoxic carbonyls. Int. J. Mol. Sci. 2022, 23, 222. [Google Scholar] [CrossRef] [PubMed]
- Dontsov, A.; Yakovleva, M.; Trofimova, N.; Sakina, N.; Gulin, A.; Aybush, A.; Gostev, F.; Vasin, A.; Feldman, T.; Ostrovsky, M. Water-soluble products of photooxidative destruction of the bisretinoid A2E cause proteins modification in the dark. Int. J. Mol. Sci. 2022, 23, 1534. [Google Scholar] [CrossRef]
- Feldman, T.; Ostrovskiy, D.; Yakovleva, M.; Dontsov, A.; Borzenok, S.; Ostrovsky, M. Lipofuscin-mediated photic stress induces a dark toxic effect on ARPE-19 cells. Int. J. Mol. Sci. 2022, 23, 12234. [Google Scholar] [CrossRef] [PubMed]
- Feldman, T.B.; Yakovleva, M.A.; Arbukhanova, P.M.; Borzenok, S.A.; Kononikhin, A.S.; Popov, I.A.; Nikolaev, E.N.; Ostrovsky, M.A. Changes in spectral properties and composition of lipofuscin fluorophores from human retinal pigment epithelium with age and pathology. Anal. Bioanal. Chem. 2015, 407, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yanase, E.; Feng, X.; Siegel, M.M.; Sparrow, J.R. Structural characterization of bisretinoid A2E photocleavage products and implications for age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2010, 107, 7275–7280. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shabat, S.; Itagaki, Y.; Jockusch, S.; Sparrow, J.R.; Turro, N.J.; Nakanishi, K. Formation of a nona-oxirane from A2E, alipofuscin fluorophore related to macular degeneration, and evidence of singlet oxygen involvement. Angew. Chem. Int. Ed. 2002, 41, 814–817. [Google Scholar] [CrossRef]
- Yoon, K.D.; Yamamoto, K.; Ueda, K.; Zhou, J.; Sparrow, J.R. A novel source of methylglyoxal and glyoxal in retina: Implications for age-related macular degeneration. PLoS ONE 2012, 7, e41309. [Google Scholar] [CrossRef] [PubMed]
- Ergin, V.; Ebrahimi, R.; Karasu, C. Carbonyl stress in aging process: Role of vitamins and phytochemicals as redox regulators. Aging Dis. 2013, 4, 276–294. [Google Scholar] [CrossRef]
- Schleicher, E.D.; Bierhaus, A.; Haring, H.U.; Nawroth, P.P.; Lehmann, R. Chemistry and pathobiology of advanced glycation end products. Contrib. Nephrol. 2001, 131, 1–9. [Google Scholar] [CrossRef]
- Feldman, T.B.; Yakovleva, M.A.; Larichev, A.V.; Arbukhanova, P.M.; Radchenko, A.S.; Borzenok, S.A.; Kuzmin, V.A.; Ostrovsky, M.A. Spectral analysis of fundus autofluorescence pattern as a tool to detect early stages of degeneration in the retina and retinal pigment epithelium. Eye 2018, 32, 1440–1448. [Google Scholar] [CrossRef]
- Feldman, T.B.; Ostrovsky, M.A.; Yakovleva, M.A.; Larichev, A.V.; Borzenok, S.A.; Arbukhanova, P.M. A Method for the Early Detectionof Age-Related Macular Retinal Dystrophy. RF Patent No. 2651126, 13 February 2017. [Google Scholar]
- Larichev, A.V.; Panchenko, V.Y.; Ostrovsky, M.A.; Feldman, T.B. Optical Device for the Study of the Fundus to Detect Age-Related Macular Dystrophy of the Retina. RF Patent for Utility Model No. 176795, 9 August 2017. [Google Scholar]
- Ferdous, S.; Liao, K.L.; Gefke, I.D.; Summers, V.R.; Wu, W.; Donaldson, K.J.; Kim, Y.-K.; Sellers, J.T.; Dixon, J.A.; Shelton, D.A.; et al. Age-related retinal changes in wild-type C57BL/6J mice between 2 and 32 months. Investig. Ophthalmol. Vis. Sci. 2021, 62, 9. [Google Scholar] [CrossRef]
- Sparrow, J.R.; Wu, Y.; Nagasaki, T.; Yoon, K.D.; Yamamoto, K.; Zhou, J. Fundus autofluorescence and the bisretinoids of retina. Photochem. Photobiol. Sci. 2010, 9, 1480–1489. [Google Scholar] [CrossRef]
- Kim, S.R.; Jang, Y.P.; Jockusch, S.; Fishkin, N.E.; Turro, N.J.; Sparrow, J.R. The all-trans-retinal dimer series of lipofuscin pigments in retinal pigment epithelial cells in a recessive Stargardt disease model. Proc. Natl. Acad. Sci. USA 2007, 104, 19273–19278. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Belli, M.; Indovina, L. The response of living organisms to low radiation environment and its implications in radiation protection. Front. Public Health 2020, 8, 601711. [Google Scholar] [CrossRef] [PubMed]
- Chien, T.; Tseng, T.L.; Wang, J.Y.; Shen, Y.T.; Lin, T.H.; Shieh, J.C. Candida albicans DBF4 gene inducibly duplicated by the mini-Urablaster is involved in hypha-suppression. Mutat. Res. 2015, 779, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Kappes, F.; Mundade, A.; Deutzmann, A.; Rosmarin, D.M.; Legendre, M.; Chatain, N.; Al-Obaidi, Z.; Adams, B.S.; Ploegh, H.L.; et al. Intercellular trafficking of the nuclear oncoprotein DEK. Proc. Natl. Acad. Sci. USA 2013, 110, 6847–6852. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.J.; Rakoczy, P.E.; Constable, I.J. Lipofuscin of the retinal pigment epithelium: A review. Eye 1995, 9, 763–771. [Google Scholar] [CrossRef]
- Amoaku, W.M.; Mahon, G.J.; Gardiner, T.A.; Frew, L.; Archer, D.B. Late ultrastructural changes in the retina of the rat following low-dose X-irradiation. Graefe’s Arch. Clin. Exp. Ophthalmol. 1992, 230, 569–574. [Google Scholar] [CrossRef]
- Rózanowska, M.; Sarna, T. Light-induced damage to the retina: Role of rhodopsin chromophore revisited. Photochem. Photobiol. 2005, 81, 1305–1330. [Google Scholar] [CrossRef]
- Feldman, T.B.; Dontsov, A.E.; Yakovleva, M.A.; Ostrovsky, M.A. Photobiology of lipofuscin granules in the retinal pigment epithelium cells of the eye: Norm, pathology, age. Biophys. Rev. 2022, 14, 1051–1065. [Google Scholar] [CrossRef]
- Kubančák, J.; Molokanov, A.G. Measurements of LET spectra of the JINR phasotron radiotherapy proton beam. Probl. At. Sci. Technol. 2013, 88, 90–92. [Google Scholar]
- Mata, N.L.; Weng, J.; Travis, G.H. Biosynthesis of a major lipofuscin fluorophore in mice and humans with ABCR-mediated retinal and macular degeneration. Proc. Natl. Acad. Sci. USA 2000, 97, 7154–7159. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Holz, F.G.; Fleckenstein, M.; Schmitz-Valckenberg, S.; Bird, A.C. Evaluation of fundus autofluorescence images. In Atlas of Fundus Autofluorescence Imaging; Holz, F.G., Schmitz-Valckenberg, S., Spaide, R.F., Bird, A.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 71–76. [Google Scholar] [CrossRef]
- Parish, C.A.; Hashimoto, M.; Nakanishi, K.; Dillon, J.; Sparrow, J. Isolation and one-step preparation of A2E and iso-A2E, fluorophores from human retinal pigment epithelium. Proc. Natl. Acad. Sci. USA 1998, 95, 14609–14613. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chloroform Extract Samples from the Retinas | |||
|---|---|---|---|
| Control Cr | Protons, 2 Gy Pr | Control (after 4.5 months) Cr+ | Protons, 2 Gy (after 4.5 months) Pr+ |
| 1.13 | 1.26 | 0.70 | 0.82 |
| Control Cr | Light Lr | Control (after 4.5 months) Cr+ | Light (after 4.5 months) Lr+ |
| 1.13 | 1.30 | 0.70 | 1.30 |
| Chloroform Extract Samples from the RPEs | |||
| Control CR | Protons, 2 Gy PR | Control (after 4.5 months) CR+ | Protons, 2 Gy (after 4.5 months) PR+ |
| 0.91 | 1.37 | 1.10 | 0.97 |
| Control CR | Light LR | Control (after 4.5 months) CR+ | Light (after 4.5 months) LR+ |
| 0.91 | 1.39 | 1.10 | 1.49 |
| Chloroform Extract Samples from the Retinas | ||||
|---|---|---|---|---|
| № Peak/Group of Peaks * | Control Cr % | Protons, 2 Gy Pr % | Control (after 4.5 Months) Cr+ % | Protons, 2 Gy (after 4.5 Months) Pr+ % |
| 1 | 22.9 ± 0.8 | 45.3 ± 0.7 | 24.8 ± 0.2 | 49.1 ± 0.3 |
| 2 | 10.5 ± 0.5 | 13.4 ± 0.7 | 18.3 ± 0.5 | 21.3 ± 0.5 |
| 3 | 33.7 ± 0.8 | 21.4 ± 0.4 | 12.4 ± 0.6 | 3.3 ± 0.5 |
| 4 | 4.0 ± 0.2 | 4.8 ± 0.1 | 6.5 ± 0.2 | 7.1 ± 0.3 |
| 5 | 28.9 ± 0.5 | 15.1 ± 0.3 | 38.0 ± 0.8 | 19.2 ± 0.7 |
| Chloroform Extract Samples from the RPEs | ||||
| № Peak/Group of Peaks * | Control CR % | Protons, 2 Gy PR % | Control (after 4.5 Months) CR+ % | Protons, 2 Gy (after 4.5 Months) PR+ % |
| 1 | 40.1 ± 0.8 | 30.2 ± 0.6 | 38.1 ± 1.3 | 40.1 ± 0.3 |
| 2 | 26.8 ± 0.5 | 37.2 ± 0.7 | 29.1 ± 0.9 | 43.7 ± 0.5 |
| 3 | 10.1 ± 0.8 | 8.9 ± 0.4 | 9.2 ± 0.4 | 0.7 ± 0.5 |
| 4 | 11.3 ± 0.2 | 4.4 ± 0.1 | 16.3 ± 0.6 | 0.8 ± 0.3 |
| 5 | 11.7 ± 0.5 | 19.3 ± 0.3 | 7.3 ± 0.7 | 14.7 ± 0.7 |
| Chloroform Extract Samples from the Retinas | ||||
|---|---|---|---|---|
| № Peak/Group of Peaks * | Control Cr % | Light Lr % | Control (after 4.5 Months) Cr+ % | Light (after 4.5 Months) Lr+ % |
| 1 | 22.9 ± 0.8 | 32.1 ± 0.8 | 24.8 ± 0.2 | 34.2 ± 0.9 |
| 2 | 10.5 ± 0.5 | 12.5 ± 0.6 | 18.3 ± 0.5 | 18.7 ± 0.7 |
| 3 | 33.7 ± 0.8 | 18.2 ± 0.6 | 12.4 ± 0.6 | 5.2 ± 0.3 |
| 4 | 4.0 ± 0.2 | 2.1 ± 0.1 | 6.5 ± 0.2 | 0.8 ± 0.1 |
| 5 | 28.9 ± 0.5 | 35.1 ± 0.3 | 38.0 ± 0.8 | 41.1 ± 0.7 |
| Chloroform Extract Samples from the RPEs | ||||
| № Peak/Group of Peaks * | Control CR % | Light LR % | Control (after 4.5 Months) CR+ % | Light (after 4.5 Months) LR+ % |
| 1 | 40.1 ± 0.8 | 37.1 ± 0.6 | 38.1 ± 1.3 | 35.1 ± 0.7 |
| 2 | 26.8 ± 0.5 | 32.1 ± 0.9 | 29.1 ± 0.9 | 29.8 ± 0.8 |
| 3 | 10.1 ± 0.8 | 13.3 ± 0.5 | 9.2 ± 0.4 | 17.7 ± 0.6 |
| 4 | 11.3 ± 0.2 | 8.3 ± 0.2 | 16.3 ± 0.6 | 6.5 ± 0.4 |
| 5 | 11.7 ± 0.5 | 9.2 ± 0.3 | 7.3 ± 0.7 | 10.9 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feldman, T.; Yakovleva, M.; Utina, D.; Ostrovsky, M. Short-Term and Long-Term Effects after Exposure to Ionizing Radiation and Visible Light on Retina and Retinal Pigment Epithelium of Mouse Eye. Int. J. Mol. Sci. 2023, 24, 17049. https://doi.org/10.3390/ijms242317049
Feldman T, Yakovleva M, Utina D, Ostrovsky M. Short-Term and Long-Term Effects after Exposure to Ionizing Radiation and Visible Light on Retina and Retinal Pigment Epithelium of Mouse Eye. International Journal of Molecular Sciences. 2023; 24(23):17049. https://doi.org/10.3390/ijms242317049
Chicago/Turabian StyleFeldman, Tatiana, Marina Yakovleva, Dina Utina, and Mikhail Ostrovsky. 2023. "Short-Term and Long-Term Effects after Exposure to Ionizing Radiation and Visible Light on Retina and Retinal Pigment Epithelium of Mouse Eye" International Journal of Molecular Sciences 24, no. 23: 17049. https://doi.org/10.3390/ijms242317049
APA StyleFeldman, T., Yakovleva, M., Utina, D., & Ostrovsky, M. (2023). Short-Term and Long-Term Effects after Exposure to Ionizing Radiation and Visible Light on Retina and Retinal Pigment Epithelium of Mouse Eye. International Journal of Molecular Sciences, 24(23), 17049. https://doi.org/10.3390/ijms242317049