
Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
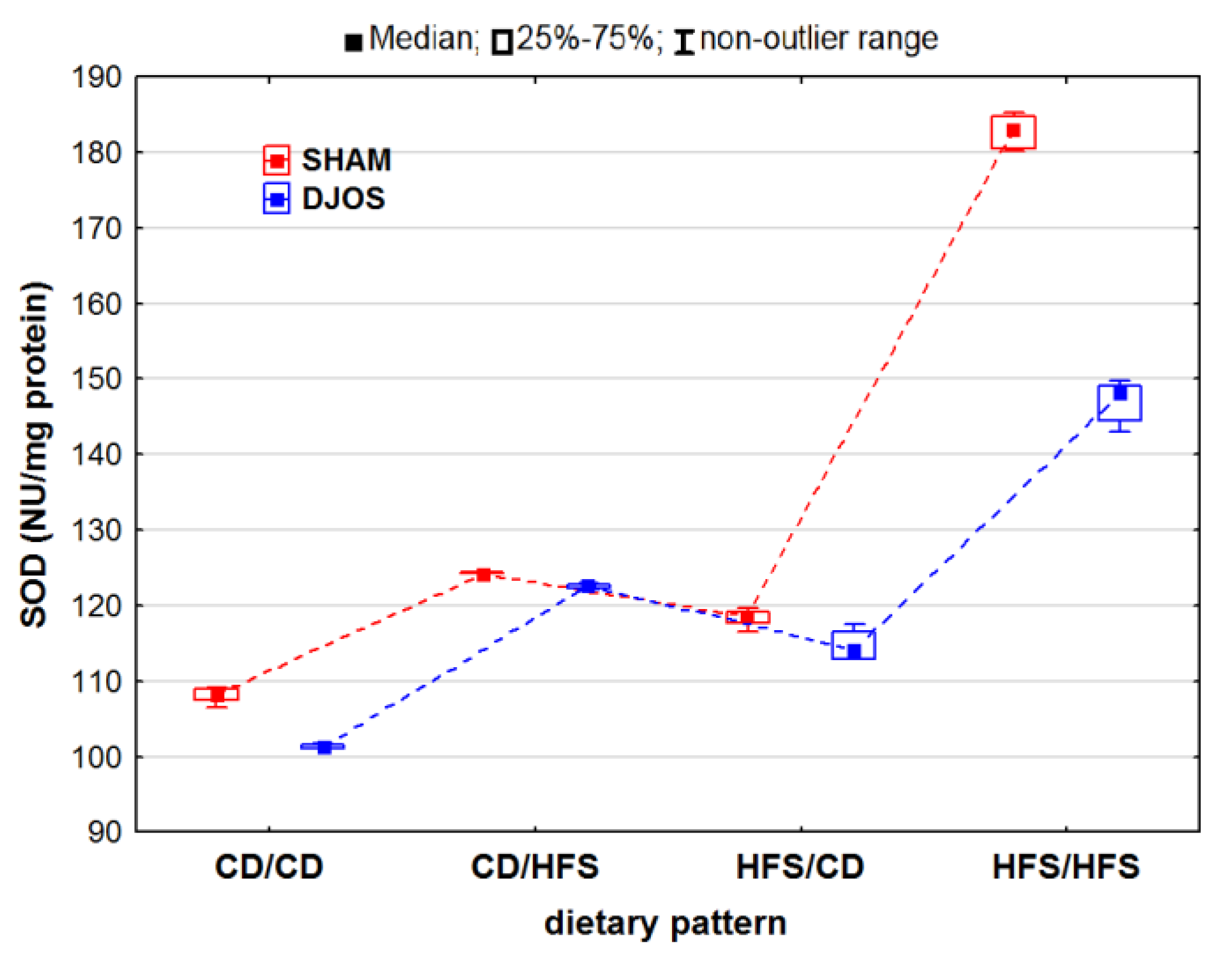
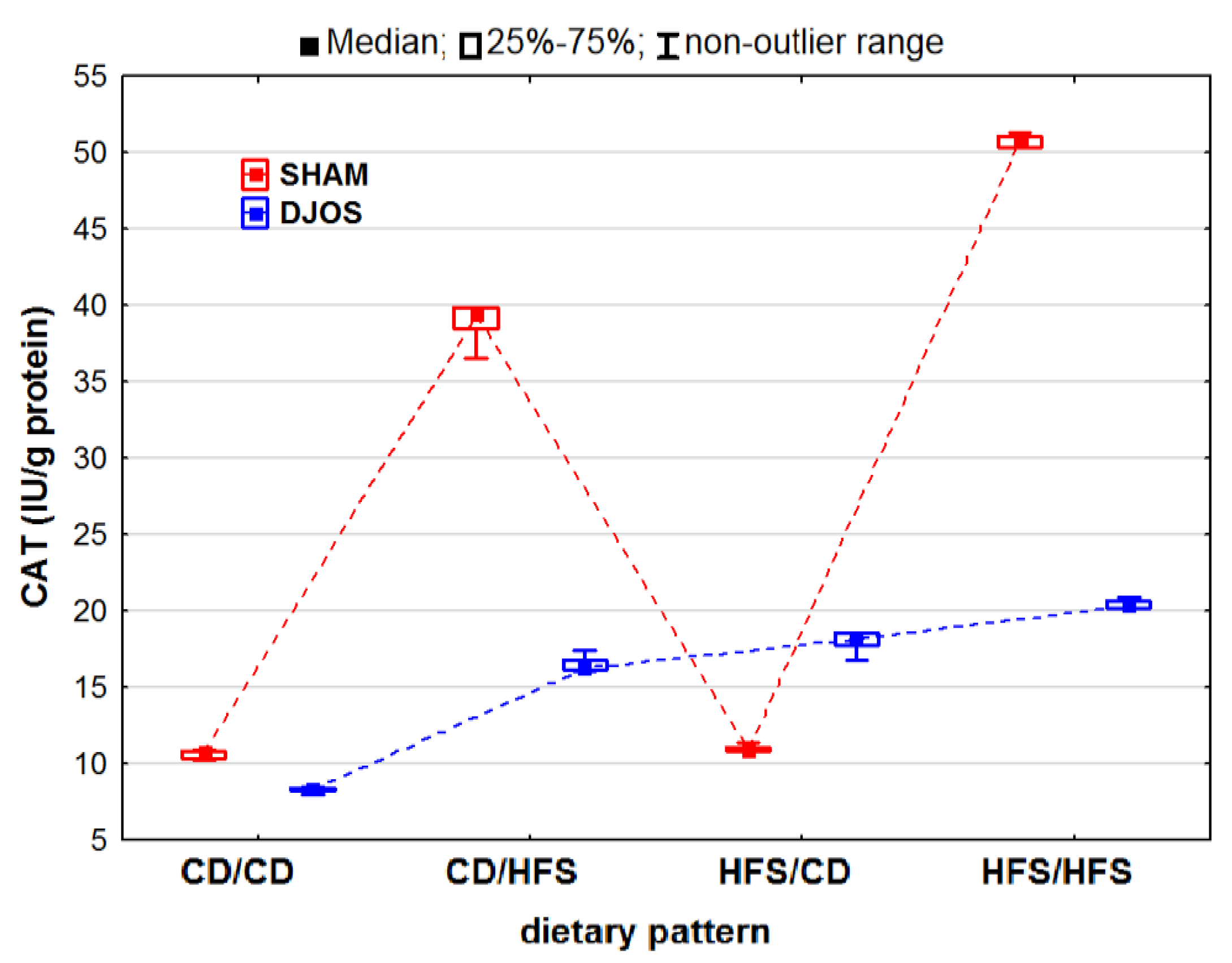
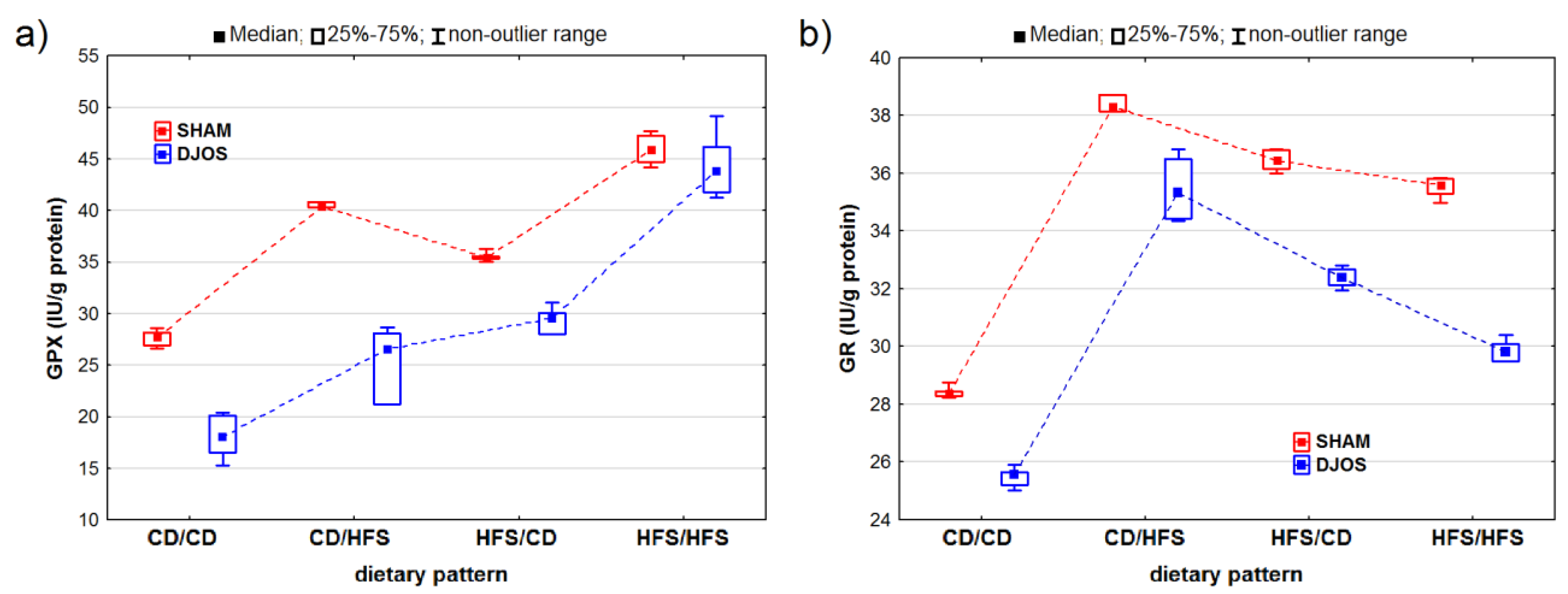
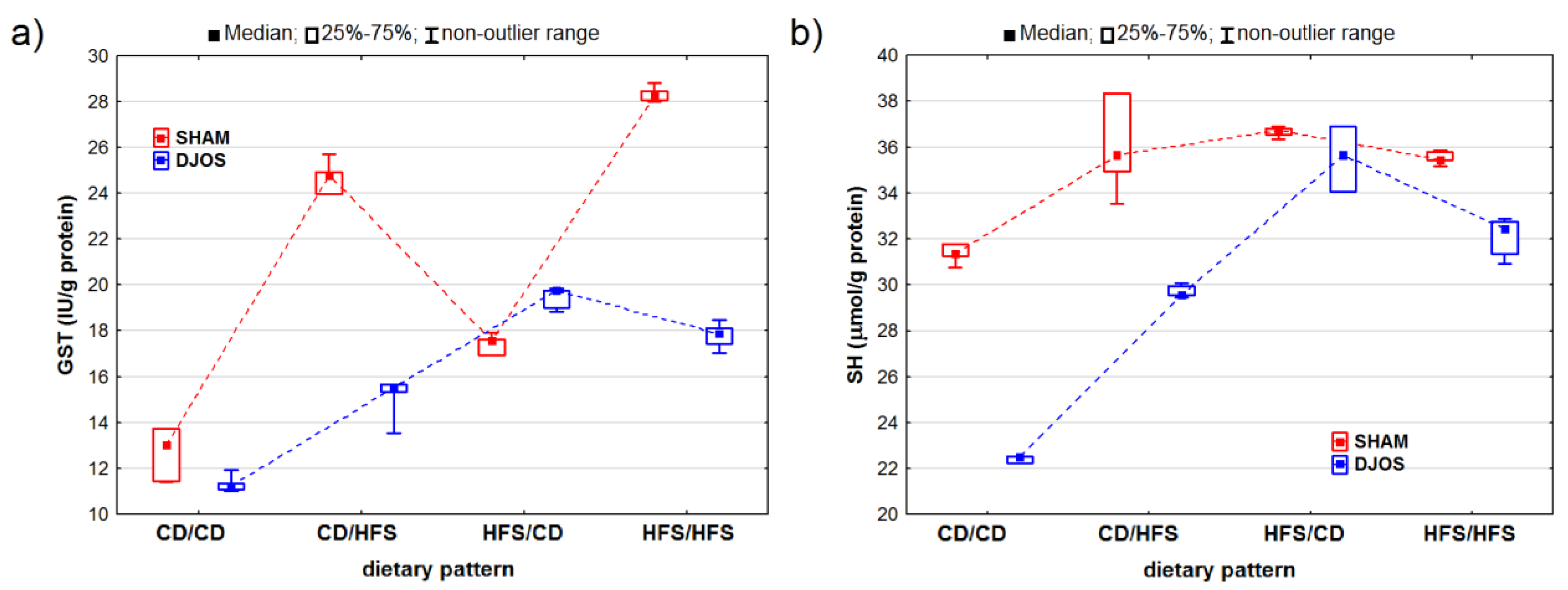
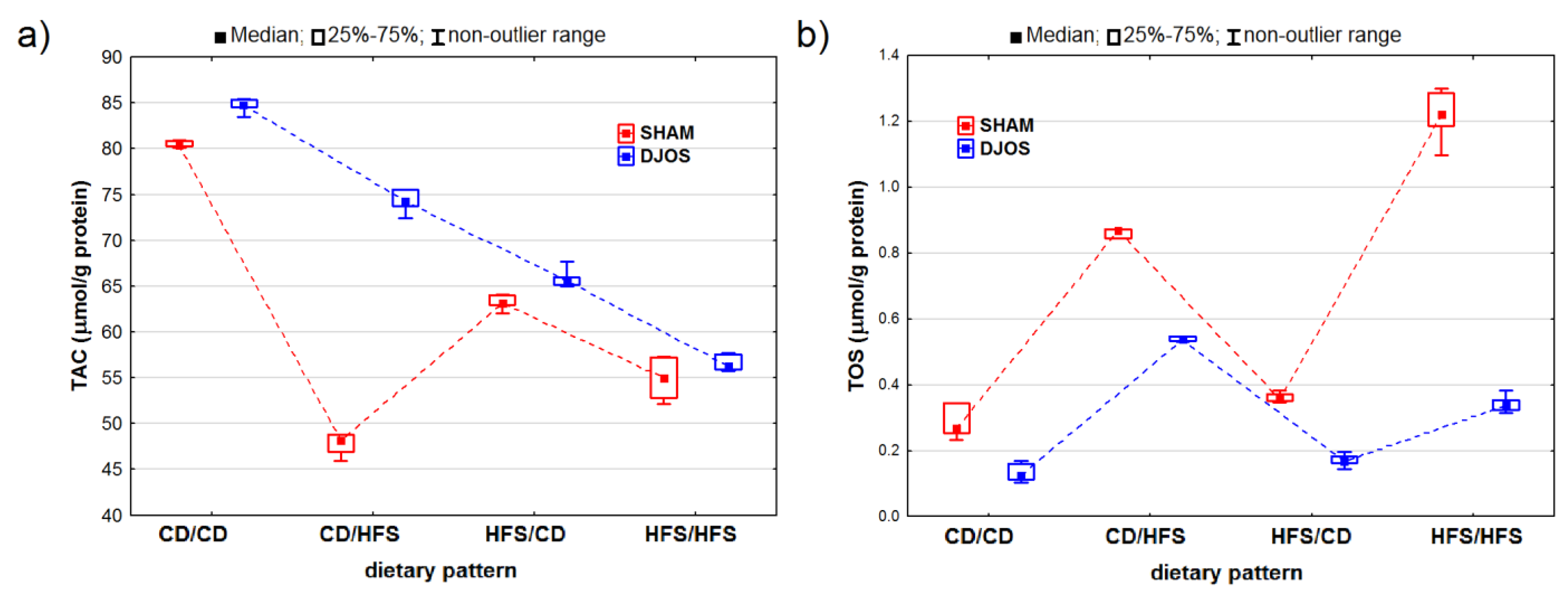
2. Results
2.1. Antioxidant Status Markers
2.2. Cholesterol, Triglyceride (TG), and HDL Levels in the Serum
3. Discussion
4. Materials and Methods
4.1. Ethical Permission
4.2. Study Subject
4.3. Study Design
4.4. Experimental Procedures
4.4.1. Control (SHAM) and Duodenojejunal Omega Switch (DJOS) Surgery
4.4.2. Sample Collection and Preparation
4.4.3. Antioxidant Status Markers Analysis
Superoxide Dismutase (SOD) (EC 1.15.1.1) Activity
Catalase (CAT) Activity (EC 1.11.1.6)
Glutathione Peroxidase (GPx) Activity (EC 1.11.1.9)
Glutathione Reductase (GR) Activity (EC 1.8.1.7)
Glutathione-S Transferase (GST) Activity (EC 2.5.1.18)
Sulfhydryl Groups (SH) Concentration
Total Oxidant Status (TOS) and Total Antioxidant Capacity (TAC)
Malondialdehyde (MDA) Concentration
4.4.4. Cholesterol, Triglycerides (TG), and HDL Concentrations
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Świderska, M.; Maciejczyk, M.; Zalewska, A.; Pogorzelska, J.; Flisiak, R.; Chabowski, A. Oxidative stress biomarkers in the serum and plasma of patients with non-alcoholic fatty liver disease (NAFLD). Can plasma AGE be a marker of NAFLD? Oxidative stress biomarkers in NAFLD patients. Free Radic. Res. 2019, 53, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Stefater, M.A.; Wilson-Perez, H.E.; Chambers, A.P.; Sandoval, D.A.; Seeley, R.J. All bariatric surgeries are not created equal: Insights from mechanistic comparisons. Endocr. Rev. 2012, 33, 595–622. [Google Scholar] [CrossRef]
- Stygar, D.; Sawczyn, T.; Skrzep-Poloczek, B.; Owczarek, A.J.; Matysiak, N.; Michalski, M.; Mielańczyk, Ł.; Bażanów, B.; Ziora, P.; Choręza, P.; et al. The effects of duodenojejunal omega switch in combination with high-fat diet and control diet on incretins, body weight, and glucose tolerance in Sprague-Dawley rats. Obes Surg. 2018, 28, 748–759. [Google Scholar] [CrossRef]
- Grueneberger, J.M.; Fritz, T.; Zhou, C.; Meyer, S.; Karcz-Socha, I.; Sawczyn, T.; Stygar, D.; Goos, M.; Hopt, U.T.; Küsters, S. Long segment ileal transposition leads to early amelioration of glucose control in the diabetic obese Zucker rat. Videosurgery Other Miniinvasive Tech. 2013, 8, 130–138. [Google Scholar] [CrossRef]
- Grueneberger, J.M.; Karcz-Socha, I.; Sawczyn, T.; Kosmowski, J.; Stygar, D.; Goos, M.; Küsters, S.; Zwirska-Korczala, K.; Marjanovic, G.; Keck, T.; et al. Systematic ileal transposition in Zucker rats shows advantage for long segment distal transposition. Surgery 2014, 155, 165–172. [Google Scholar] [CrossRef]
- Stygar, D.; Skrzep-Poloczek, B.; Romuk, E.; Chełmecka, E.; Poloczek, J.; Sawczyn, T.; Maciarz, J.; Kukla, M.; Karcz, K.W.; Jochem, J. The influence of high-fat, high-sugar diet and bariatric surgery on HSP70 and HSP90 plasma and liver concentrations in diet-induced obese rats. Cell Stress Chaperones 2019, 24, 427–439. [Google Scholar] [CrossRef]
- Rezzani, R.; Franco, C. Liver, Oxidative stress and metabolic syndromes. Nutrients 2021, 13, 301. [Google Scholar] [CrossRef]
- Klisic, A.; Kavaric, N.; Ninic, A.; Kotur-Stevuljevic, J. Oxidative stress and cardiometabolic biomarkers in patients with non-alcoholic fatty liver disease. Sci. Rep. 2021, 11, 18455. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Takaki, A.; Kawai, D.; Yamamoto, K. Multiple hits, including oxidative stress, as pathogenesis and treatment target in non-alcoholic steatohepatitis (NASH). Int. J. Mol. Sci. 2013, 14, 20704–20728. [Google Scholar] [CrossRef] [PubMed]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.M.; Fletcher, J.A.; Thyfault, J.P.; Rector, R.S. The role of angiotensin II in nonalcoholic steatohepatitis. Mol. Cell. Endocrinol. 2013, 378, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Clark, S.E.; Morris, E.M.; Thyfault, J.P.; Uptergrove, G.M.; Whaley-Connell, A.T.; Ferrario, C.M.; Sowers, J.R.; Ibdah, J.A. Angiotensin II-induced non-alcoholic fatty liver disease is mediated by oxidative stress in transgenic TG(mRen2)27(Ren2) rats. J. Hepatol. 2008, 49, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Patrick, L. Nonalcoholic fatty liver disease: Relationship to insulin sensitivity and oxidative stress. Treatment approaches using vitamin E, magnesium, and betaine. Altern. Med. Rev. 2002, 7, 276–291. [Google Scholar] [PubMed]
- Videla, L.A. Oxidative stress signaling underlying liver disease and hepatoprotective mechanisms. World. J. Hepatol. 2009, 1, 72–78. [Google Scholar] [CrossRef]
- Sumida, Y.; Niki, E.; Naito, Y.; Yoshikawa, T. Involvement of free radicals and oxidative stress in NAFLD/NASH. Free Radic. Res. 2013, 47, 869–880. [Google Scholar] [CrossRef]
- Ferro, D.; Baratta, F.; Pastori, D.; Cocomello, N.; Colantoni, A.; Angelico, F.; Del Ben, M. New insights into the pathogenesis of non-alcoholic fatty liver disease: Gut-derived lipopolysaccharides and oxidative stress. Nutrients 2020, 12, 2762. [Google Scholar] [CrossRef]
- Zhu, R.; Wang, Y.; Zhang, L.; Guo, Q. Oxidative stress and liver disease. Hepatol. Res. 2012, 42, 741–749. [Google Scholar] [CrossRef]
- Diesen, D.L.; Kuo, P.C. Nitric oxide and redox regulation in the liver: Part I. General considerations and redox biology in hepatitis. J. Surg. Res. 2010, 162, 95–109. [Google Scholar] [CrossRef]
- Muriel, P. Role of free radicals in liver diseases. Hepatol. Int. 2009, 3, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The Liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Polimeni, L.; Del Ben, M.; Baratta, F.; Perri, L.; Albanese, F.; Pastori, D.; Violi, F.; Angelico, F. Oxidative stress: New insights on the association of non-alcoholic fatty liver disease and atherosclerosis. World J. Hepatol. 2015, 7, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Gupta, D.K.; Palma, J.M.; Corpas, F.J. (Eds.) Redox State as a Central Regulator of Plant-Cell Stress Responses, 1st ed.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Geetha, A.; Lakshmi, P.; Jeyachristy, S.A.; Surendran, R. Level of oxidative stress in the red blood cells of patients with liver cirrhosis. Indian J. Med. Res. 2007, 126, 204–210. [Google Scholar] [PubMed]
- Erel, O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin. Biochem. 2004, 37, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.S.; Magalhães, L.M.; Tóth, I.V.; Segundo, M.A. Insights on antioxidant assays for biological samples based on the reduction of copper complexes-the importance of analytical conditions. Int. J. Mol. Sci. 2014, 15, 11387–11402. [Google Scholar] [CrossRef]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Vet. Res. 2016, 12, 166. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of oxidative stress in pathophysiology of nonalcoholic fatty liver disease. Oxid. Med. Cell. Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef]
- Russell, W.M.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Oyanagui, Y. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem. 1984, 142, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Mannervik, B. Glutathione peroxidase. Methods Enzymol. 1985, 113, 490–495. [Google Scholar] [PubMed]
- Carlberg, I.; Mannervik, B. Glutathione reductase. Methods Enzymol. 1985, 113, 484–490. [Google Scholar]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-transferases. Methods Enzymol. 1981, 77, 398–405. [Google Scholar]
- Koster, J.F.; Biemond, P.; Swaak, A.J. Intracellular and extracellular sulphydryl levels in rheumatoid arthritis. Ann. Rheum. Dis. 1986, 45, 44–46. [Google Scholar] [CrossRef]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Erel, O. A novel automated method to measure total antioxidant response against potent free radical reactions. Clin. Biochem. 2004, 37, 112–119. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surgery Type | Parameter [mg/dL] | CD/CD | CD/HFS | HFS/CD | HFS/HFS | Pdiet |
|---|---|---|---|---|---|---|
| DJOS | Cholesterol | 35 (33–37) | 38 (37–39) | 44 (44–45) | 55 (54–56) | <0.001 |
| TG | 49 (46–49) | 57 (55–57) | 54 (52–56) | 59 (54–60) | <0.001 | |
| HDL | 16 (15–17) | 20 (19–20) | 23 (22–24) | 20 (19–20) | <0.001 | |
| SHAM | Cholesterol | 41 (40–41) | 63 (63–65) | 50 (50–50) | 77 (74–79) | <0.001 |
| TG | 61 (60–62) | 135 (133–140) | 75 (69–78) | 149 (140–168) | <0.001 | |
| HDL | 11 (11–12) | 13 (13–14) | 36 (30–41) | 12 (10–14) | <0.001 |
| Surgery Type | Parameter | CD/CD vs. CD/HFS | CD/CD vs. HFS/CD | CD/CD vs. HFS/HFS | CD/HFS vs. HFS/CD | CD/HFS vs. HFS/HFS | HFS/CD vs. HFS/HFS |
|---|---|---|---|---|---|---|---|
| DJOS | Cholesterol | 1 | <0.05 | <0.001 | 0.384 | <0.01 | 0.668 |
| TG | <0.01 | 0.156 | <0.001 | 1 | 1 | 0.690 | |
| HDL | 0.081 | <0.001 | 0.106 | 0.149 | 1 | 0.115 | |
| SHAM | Cholesterol | <0.01 | 0.668 | <0.001 | 0.688 | 0.688 | <0.01 |
| TG | <0.01 | 0.668 | <0.001 | 0.476 | 1 | <0.05 | |
| HDL | 0.176 | <0.001 | 1 | 0.284 | 0.917 | <0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kazura, W.; Michalczyk, K.; Skrzep-Poloczek, B.; Chełmecka, E.; Zalejska-Fiolka, J.; Michalski, M.; Kukla, M.; Jochem, J.; Rutkowski, J.; Stygar, D. Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity. Int. J. Mol. Sci. 2023, 24, 16535. https://doi.org/10.3390/ijms242216535
Kazura W, Michalczyk K, Skrzep-Poloczek B, Chełmecka E, Zalejska-Fiolka J, Michalski M, Kukla M, Jochem J, Rutkowski J, Stygar D. Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity. International Journal of Molecular Sciences. 2023; 24(22):16535. https://doi.org/10.3390/ijms242216535
Chicago/Turabian StyleKazura, Wojciech, Katarzyna Michalczyk, Bronisława Skrzep-Poloczek, Elżbieta Chełmecka, Jolanta Zalejska-Fiolka, Marek Michalski, Michał Kukla, Jerzy Jochem, Jakub Rutkowski, and Dominika Stygar. 2023. "Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity" International Journal of Molecular Sciences 24, no. 22: 16535. https://doi.org/10.3390/ijms242216535
APA StyleKazura, W., Michalczyk, K., Skrzep-Poloczek, B., Chełmecka, E., Zalejska-Fiolka, J., Michalski, M., Kukla, M., Jochem, J., Rutkowski, J., & Stygar, D. (2023). Liver Oxidative Status, Serum Lipids Levels after Bariatric Surgery and High-Fat, High-Sugar Diet in Animal Model of Induced Obesity. International Journal of Molecular Sciences, 24(22), 16535. https://doi.org/10.3390/ijms242216535